
Methylglyoxal Levels in Human Colorectal Precancer and Cancer: Analysis of Tumor and Peritumor Tissue

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Selection and Data Collection
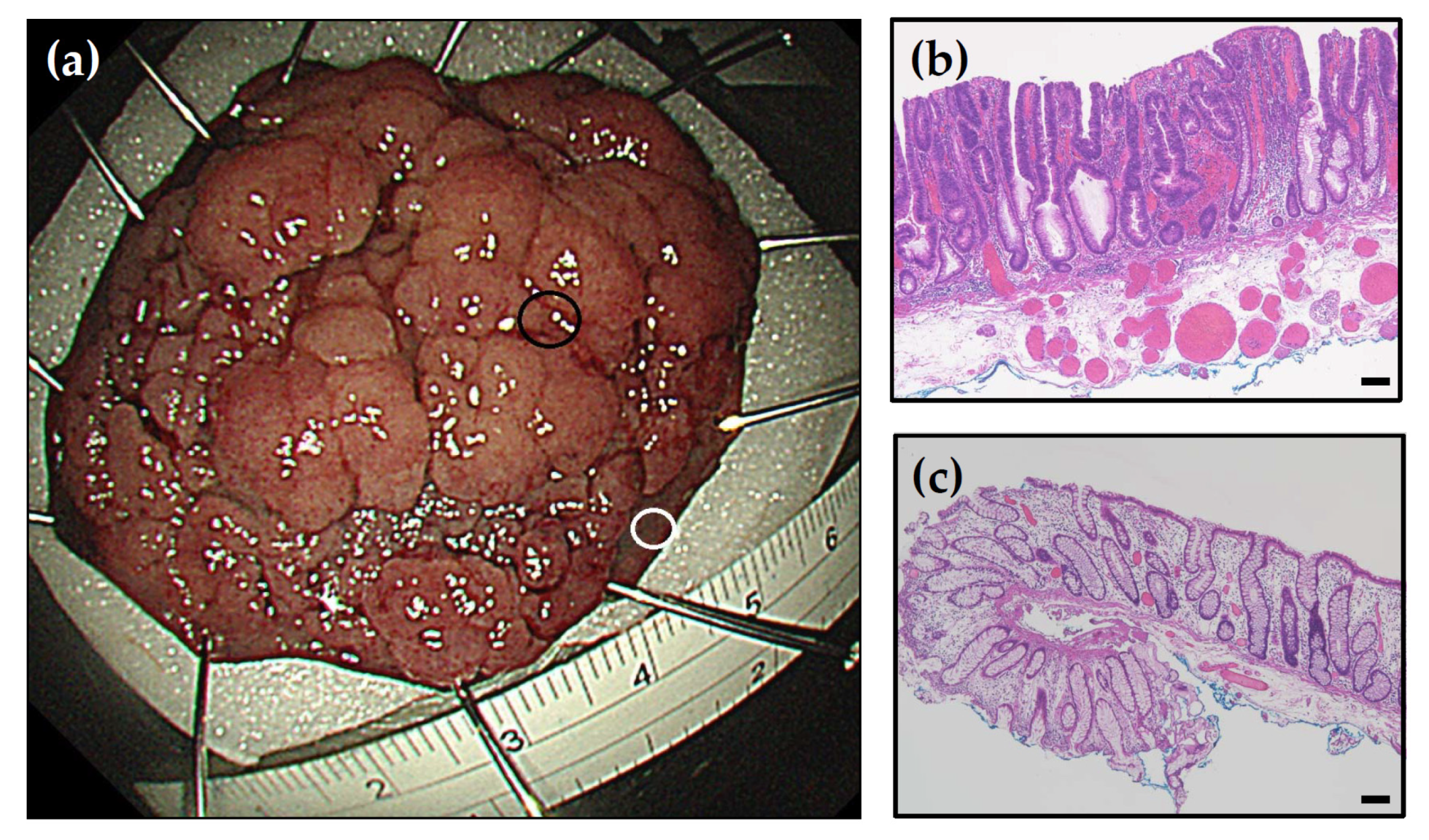
2.2. Tissue Collection
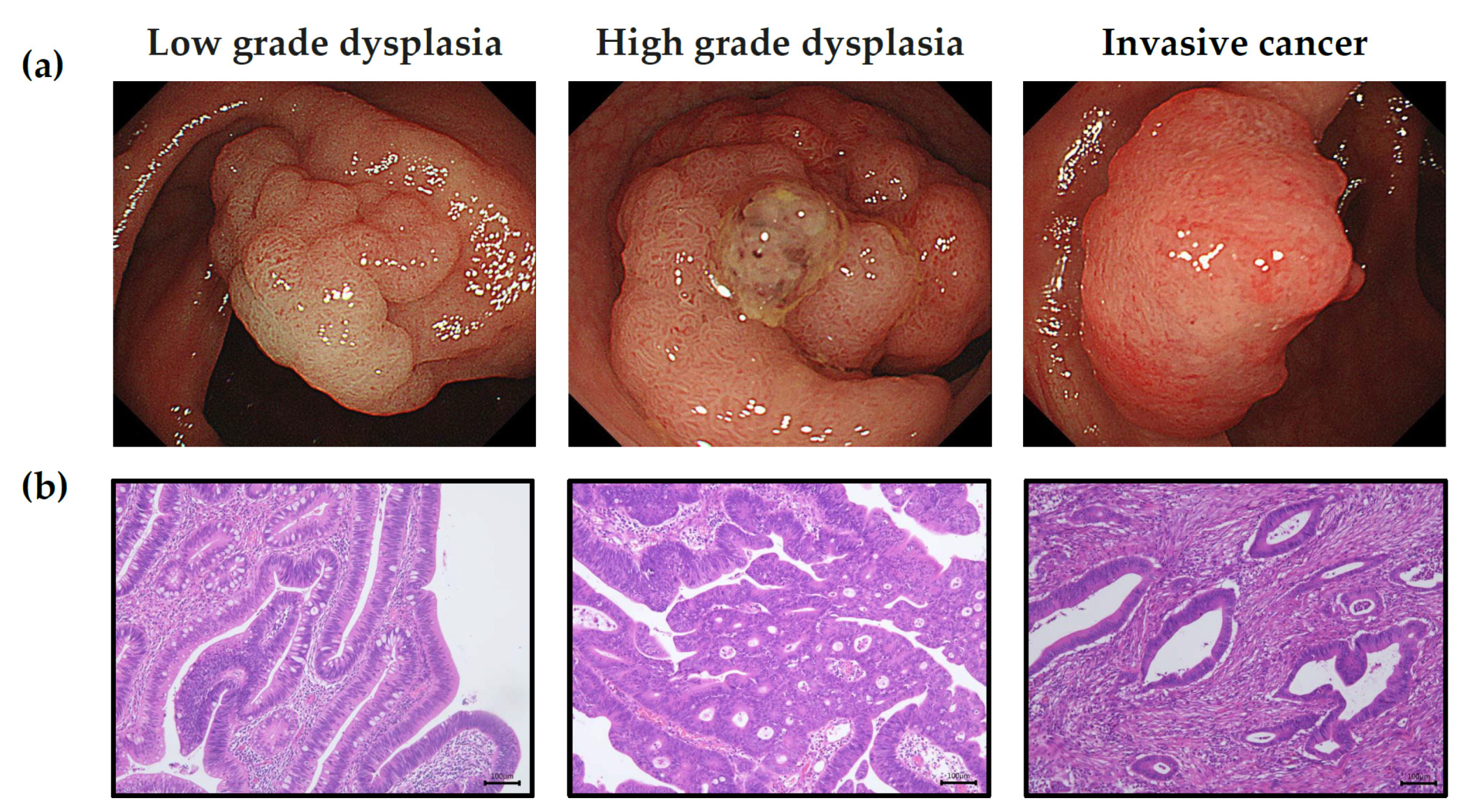
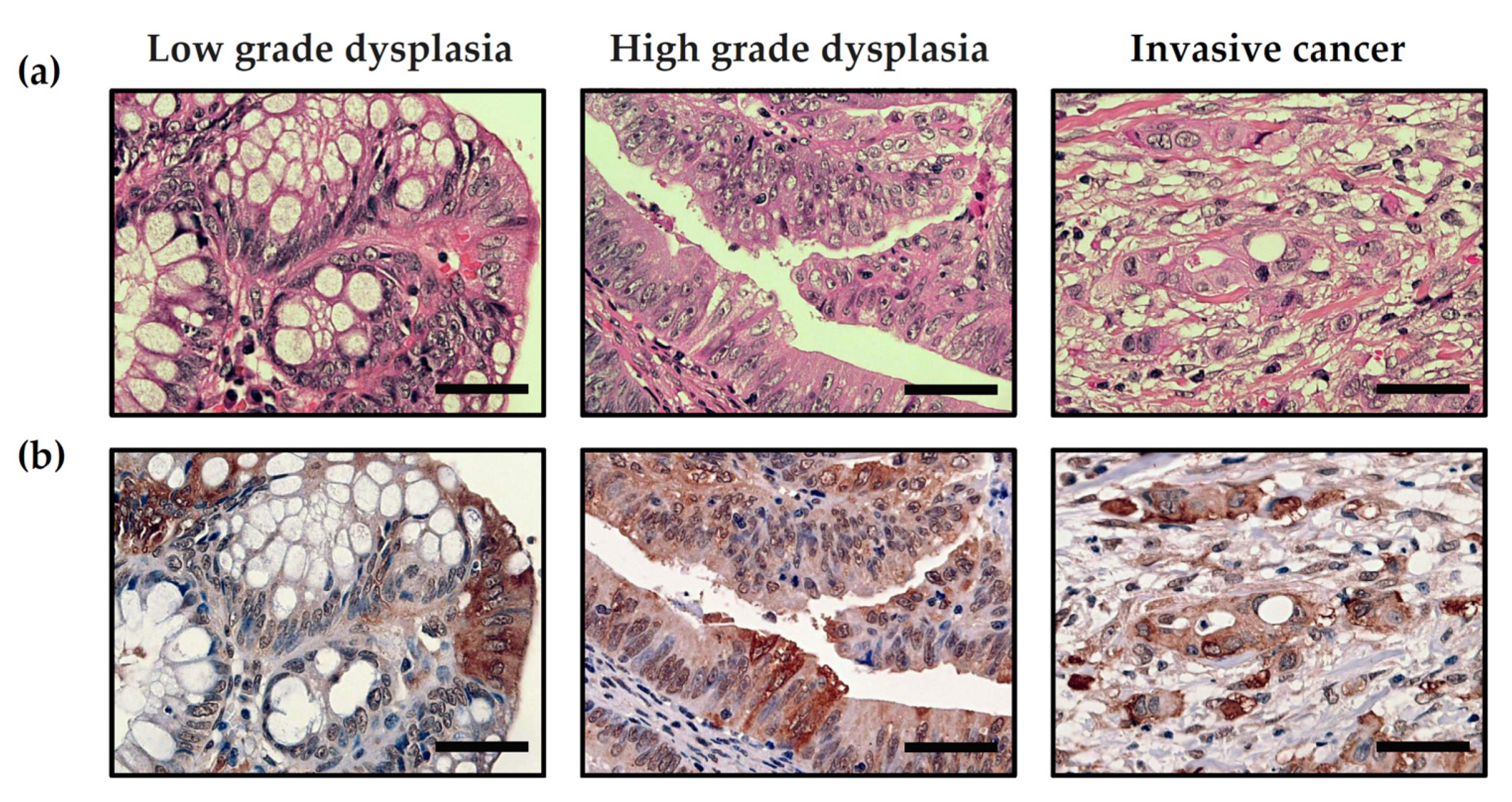
2.3. Pathology
2.4. Total Protein Assay
2.5. Quantification of MGO Concentrations by Fluorescent HPLC Analysis
2.6. Immunohistochemistry
2.7. Statistical Analyses
3. Results
3.1. Demographic Data for the Different Groups of Colorectal Neoplasms
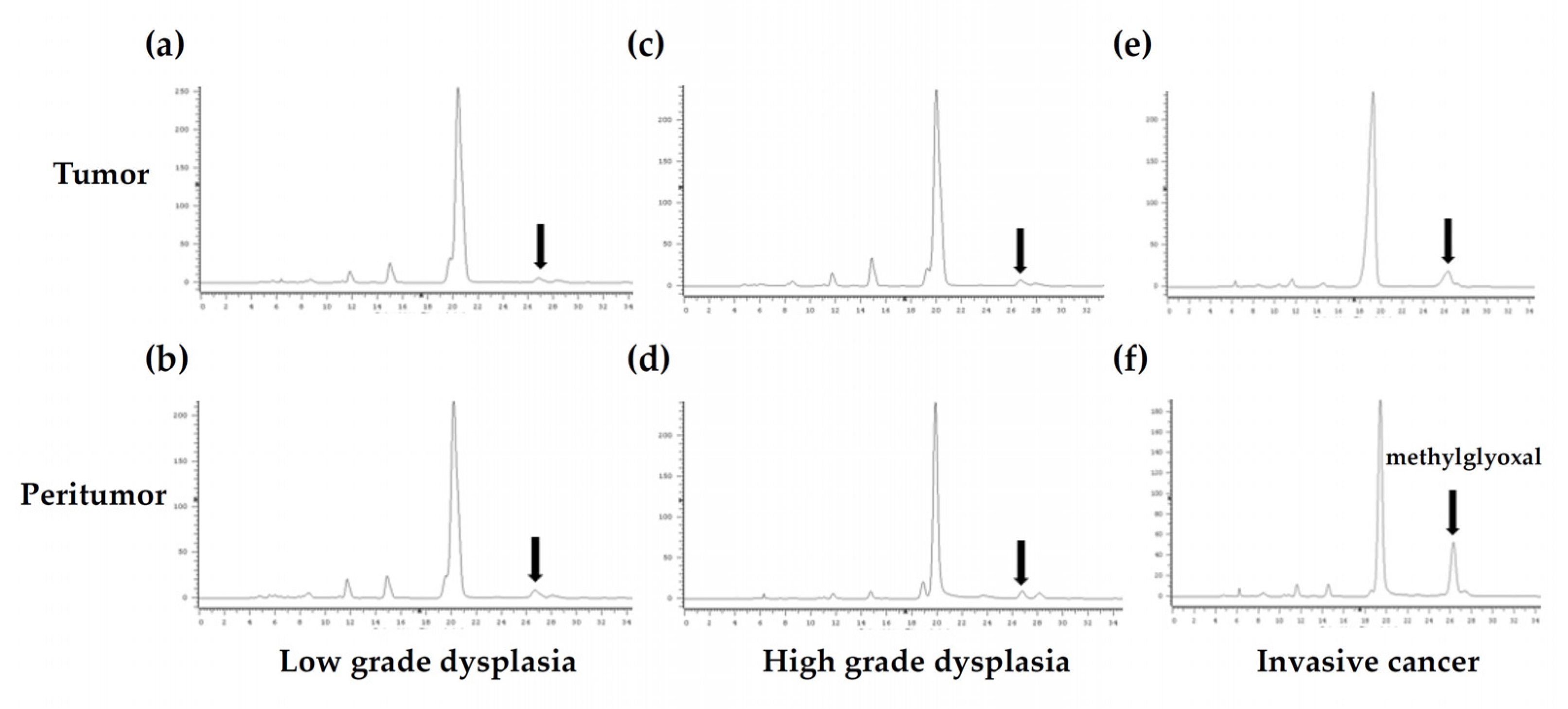
3.2. Fluorescent HPLC Results and Method Validation in Human Colon Samples
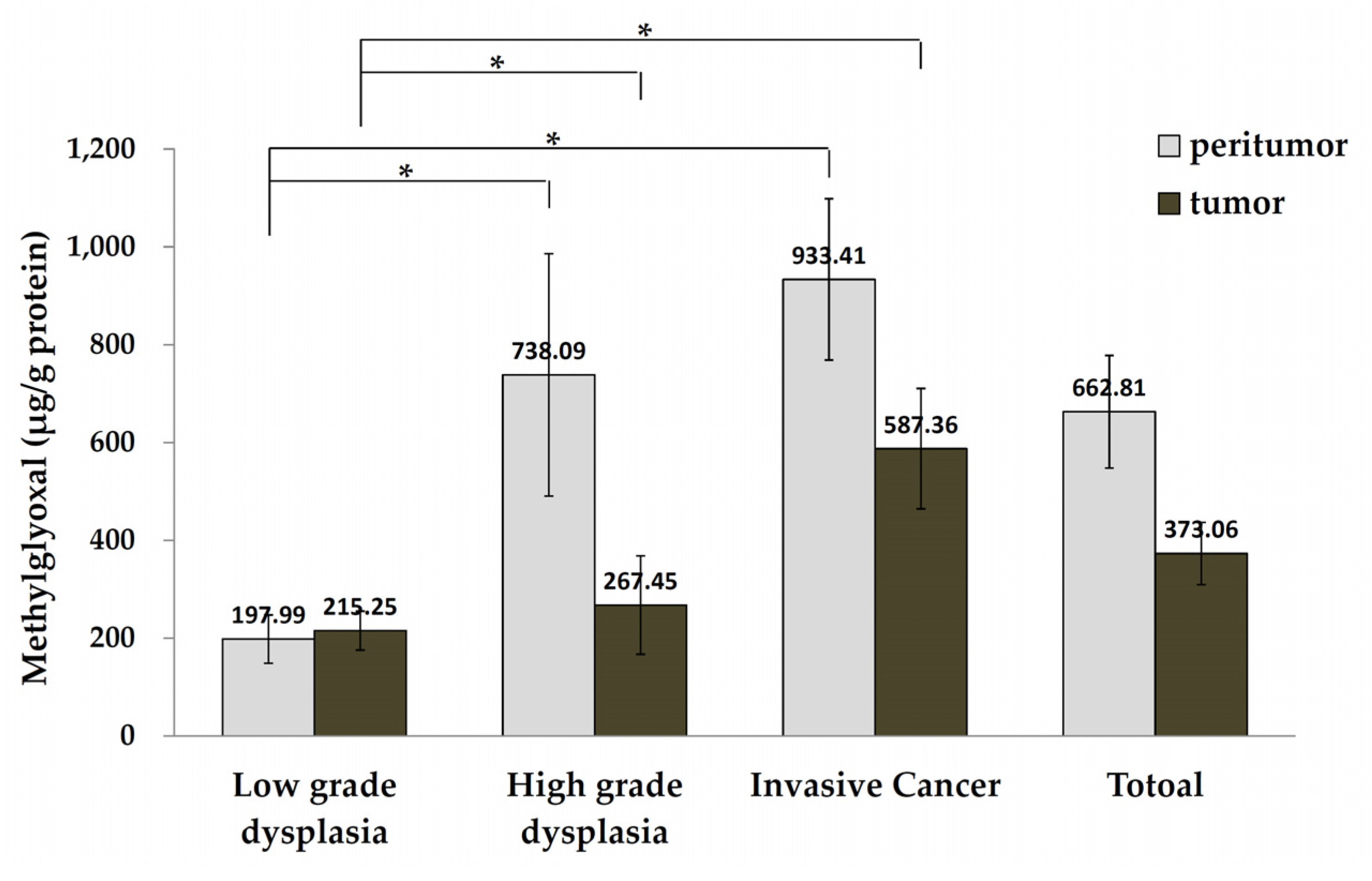
3.3. MGO Levels in Tumor and Peritumor Human Colon Tissue
3.4. Evidence of MGO Dicarbonyl Stress in Human CRC Tumors
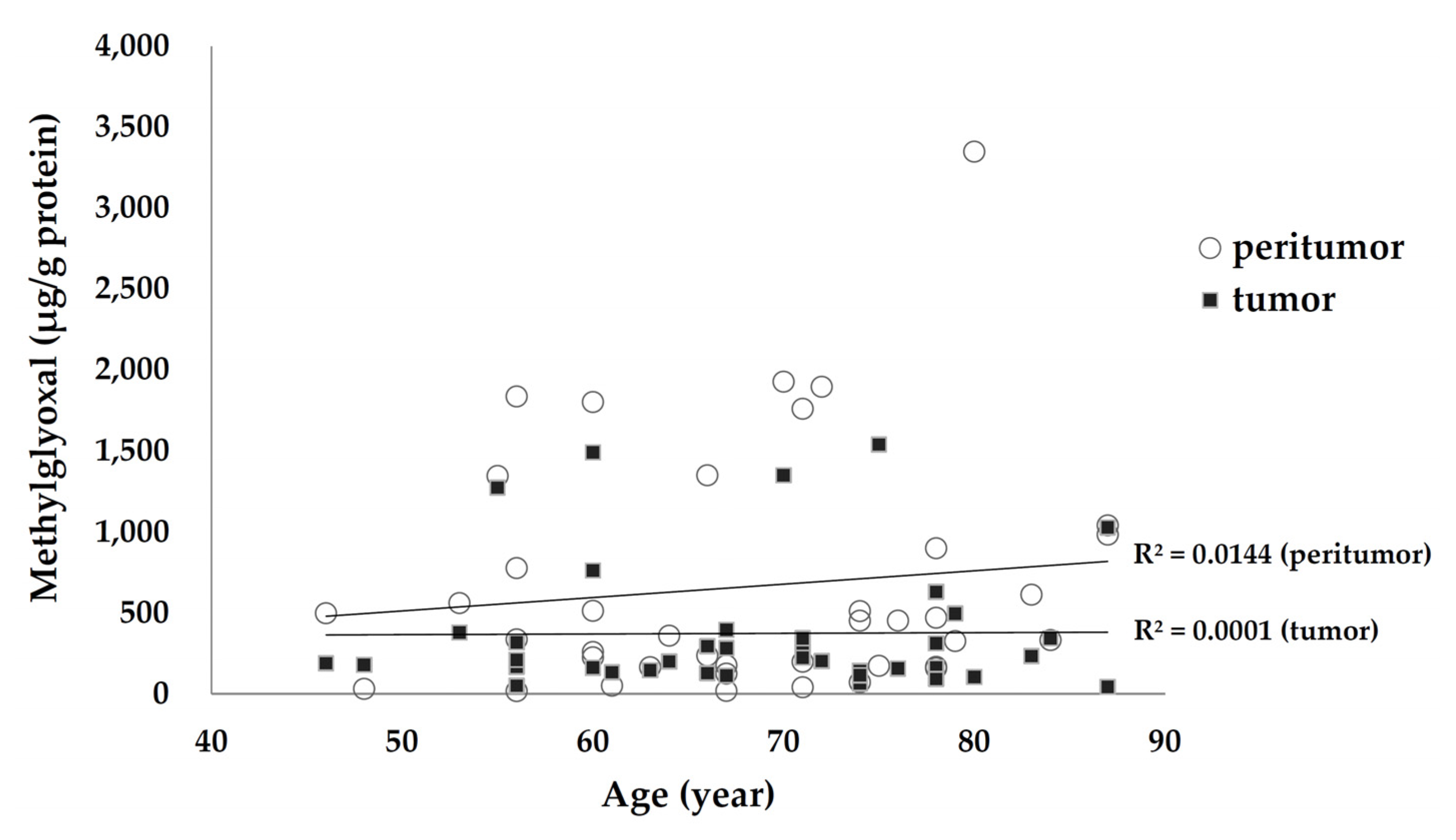
3.5. Association of Tissue MGO Levels with Age and Underlying Diseases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. The Global Cancer Observatory. Available online: http://gco.iarc.fr/today/data/factsheets/cancers/39-All-cancers-fact-sheet.pdf (accessed on 7 April 2021).
- Ikeda, Y.; Mori, M.; Shibahara, K.; Iwashita, A.; Haraguchi, Y.; Saku, M. The role of adenoma for colorectal cancer development: Differences in the distribution of adenoma with low-grade dysplasia, high-grade dysplasia, and cancer that invades the submucosa. Surgery 2002, 131, S105–S108. [Google Scholar] [CrossRef]
- Terzic, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedzwiedzka, E.; Arlukowicz, T.; Przybylowicz, K.E. A Review of Colorectal Cancer in Terms of Epidemiology, Risk Factors, Development, Symptoms and Diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef] [PubMed]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef] [PubMed]
- Turk, Z.; Cavlovic-Naglic, M.; Turk, N. Relationship of methylglyoxal-adduct biogenesis to LDL and triglyceride levels in diabetics. Life Sci. 2011, 89, 485–490. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2012, 42, 1133–1142. [Google Scholar] [CrossRef]
- Nedic, O.; Rattan, S.I.; Grune, T.; Trougakos, I.P. Molecular effects of advanced glycation end products on cell signalling pathways, ageing and pathophysiology. Free Radic. Res. 2013, 47 (Suppl. 1), 28–38. [Google Scholar] [CrossRef]
- Nigro, C.; Leone, A.; Fiory, F.; Prevenzano, I.; Nicolo, A.; Mirra, P.; Beguinot, F.; Miele, C. Dicarbonyl Stress at the Crossroads of Healthy and Unhealthy Aging. Cells 2019, 8, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazawa, N.; Abe, M.; Souma, T.; Tanemoto, M.; Abe, T.; Nakayama, M.; Ito, S. Methylglyoxal augments intracellular oxidative stress in human aortic endothelial cells. Free Radic. Res. 2010, 44, 101–107. [Google Scholar] [CrossRef]
- Liu, B.F.; Miyata, S.; Hirota, Y.; Higo, S.; Miyazaki, H.; Fukunaga, M.; Hamada, Y.; Ueyama, S.; Muramoto, O.; Uriuhara, A.; et al. Methylglyoxal induces apoptosis through activation of p38 mitogen-activated protein kinase in rat mesangial cells. Kidney Int. 2003, 63, 947–957. [Google Scholar] [CrossRef] [Green Version]
- Chiavarina, B.; Nokin, M.J.; Bellier, J.; Durieux, F.; Bletard, N.; Sherer, F.; Lovinfosse, P.; Peulen, O.; Verset, L.; Dehon, R.; et al. Methylglyoxal-Mediated Stress Correlates with High Metabolic Activity and Promotes Tumor Growth in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 213. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, S.; Tew, K.D. Analysis of glyoxalase-I from normal and tumor tissue from human colon. Biochim. Biophys. Acta 1993, 1182, 311–316. [Google Scholar] [CrossRef]
- Rabbani, N.; Xue, M.; Weickert, M.O.; Thornalley, P.J. Multiple roles of glyoxalase 1-mediated suppression of methylglyoxal glycation in cancer biology-Involvement in tumour suppression, tumour growth, multidrug resistance and target for chemotherapy. Semin. Cancer Biol. 2018, 49, 83–93. [Google Scholar] [CrossRef]
- Bellahcene, A.; Nokin, M.J.; Castronovo, V.; Schalkwijk, C. Methylglyoxal-derived stress: An emerging biological factor involved in the onset and progression of cancer. Semin. Cancer Biol. 2018, 49, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravichandran, M.; Priebe, S.; Grigolon, G.; Rozanov, L.; Groth, M.; Laube, B.; Guthke, R.; Platzer, M.; Zarse, K.; Ristow, M. Impairing L-Threonine Catabolism Promotes Healthspan through Methylglyoxal-Mediated Proteohormesis. Cell Metab. 2018, 27, 914–925. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.A.; Wu, C.H.; Yen, G.C. Methylglyoxal displays colorectal cancer-promoting properties in the murine models of azoxymethane and CT26 isografts. Free Radic. Biol. Med. 2018, 115, 436–446. [Google Scholar] [CrossRef]
- He, T.; Zhou, H.; Li, C.; Chen, Y.; Chen, X.; Li, C.; Mao, J.; Lyu, J.; Meng, Q.H. Methylglyoxal suppresses human colon cancer cell lines and tumor growth in a mouse model by impairing glycolytic metabolism of cancer cells associated with down-regulation of c-Myc expression. Cancer Biol. Ther. 2016, 17, 955–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Fang, L.; Li, G.; Zhang, J.; Li, C.; Ma, M.; Guan, C.; Bai, F.; Lyu, J.; Meng, Q.H. Synergistic inhibition of colon cancer growth by the combination of methylglyoxal and silencing of glyoxalase I mediated by the STAT1 pathway. Oncotarget 2017, 8, 54838–54857. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fang, L.; Zhang, J.; Li, G.; Ma, M.; Li, C.; Lyu, J.; Meng, Q.H. Blockage of Glyoxalase I Inhibits Colorectal Tumorigenesis and Tumor Growth via Upregulation of STAT1, p53, and Bax and Downregulation of c-Myc and Bcl-2. Int. J. Mol. Sci. 2017, 18, 570. [Google Scholar] [CrossRef] [Green Version]
- Pimentel-Nunes, P.; Dinis-Ribeiro, M.; Ponchon, T.; Repici, A.; Vieth, M.; De Ceglie, A.; Amato, A.; Berr, F.; Bhandari, P.; Bialek, A.; et al. Endoscopic submucosal dissection: European Society of Gastrointestinal Endoscopy (ESGE) Guideline. Endoscopy 2015, 47, 829–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edge, S.B.; Byrd, D.R.; Compton, C.C. American Joint Committee on Cancer (AJCC) Cancer Staging Manual, 7th ed.; Fritz, A.G., Greene, F.L., Trotti, A., Eds.; Springer: New York, NY, USA, 2010. [Google Scholar]
- Brown, K.G.M.; Solomon, M.J.; Mahon, K.; O’Shannassy, S. Management of colorectal cancer. BMJ 2019, 366, l4561. [Google Scholar] [CrossRef]
- Ferlitsch, M.; Moss, A.; Hassan, C.; Bhandari, P.; Dumonceau, J.M.; Paspatis, G.; Jover, R.; Langner, C.; Bronzwaer, M.; Nalankilli, K.; et al. Colorectal polypectomy and endoscopic mucosal resection (EMR): European Society of Gastrointestinal Endoscopy (ESGE) Clinical Guideline. Endoscopy 2017, 49, 270–297. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, S.R.; Bosman, F.T.; Boffeta, P.; Ilyas, M. Carcinoma of the colon and rectum. In WHO Classification of Tumours of the Digestive System, 4th ed.; Bozman, F.T., Carneiro, F., Hruban, R.H., Theise, N.D., Eds.; International Agency for Research on Cancer (IARC): Lyon, France, 2010; pp. 134–146. [Google Scholar]
- Schlemper, R.J.; Riddell, R.H.; Kato, Y.; Borchard, F.; Cooper, H.S.; Dawsey, S.M.; Dixon, M.F.; Fenoglio-Preiser, C.M.; Flejou, J.F.; Geboes, K.; et al. The Vienna classification of gastrointestinal epithelial neoplasia. Gut 2000, 47, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa-Mansilla, A.; Duran-Meras, I.; Salinas, F. High-performance liquid chromatographic-fluorometric determination of glyoxal, methylglyoxal, and diacetyl in urine by prederivatization to pteridinic rings. Anal. Biochem. 1998, 255, 263–273. [Google Scholar] [CrossRef]
- Srivastava, A.; Lauwers, G.Y. Gastric epithelial dysplasia: The Western perspective. Dig. Liver Dis. 2008, 40, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, C.; Morandi, P.; Fante, R.; De Gaetani, C. Gastric dysplasia. A follow-up study. Am. J. Gastroenterol. 1993, 88, 1714–1719. [Google Scholar] [PubMed]
- Fertitta, A.M.; Comin, U.; Terruzzi, V.; Minoli, G.; Zambelli, A.; Cannatelli, G.; Bodini, P.; Bertoli, G.; Negri, R.; Brunati, S.; et al. Clinical significance of gastric dysplasia: A multicenter follow-up study. Endoscopy 1993, 25, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Ikegami, M.; Shimoda, T.; Takagi, N.; Maruyama, M. Long-term follow-up study of gastric adenoma/dysplasia. Endoscopy 2004, 36, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Rugge, M.; Cassaro, M.; Di Mario, F.; Leo, G.; Leandro, G.; Russo, V.M.; Pennelli, G.; Farinati, F. The long term outcome of gastric non-invasive neoplasia. Gut 2003, 52, 1111–1116. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Jeon, S.W.; Jung, M.K.; Cho, C.M.; Tak, W.Y.; Kweon, Y.O.; Kim, S.K.; Choi, Y.H. Long-term follow-up study of gastric intraepithelial neoplasias: Progression from low-grade dysplasia to invasive carcinoma. Eur. J. Gastroenterol. Hepatol. 2008, 20, 966–970. [Google Scholar] [CrossRef]
- De Jong, M.E.; Kanne, H.; Nissen, L.H.C.; Drenth, J.P.H.; Derikx, L.; Hoentjen, F. Increased risk of high-grade dysplasia and colorectal cancer in inflammatory bowel disease patients with recurrent low-grade dysplasia. Gastrointest. Endosc. 2020, 91, 1334–1342. [Google Scholar] [CrossRef]
- Muthusami, S.; Ramachandran, I.K.; Babu, K.N.; Krishnamoorthy, S.; Guruswamy, A.; Queimado, L.; Chaudhuri, G.; Ramachandran, I. Role of Inflammation in the Development of Colorectal Cancer. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 77–90. [Google Scholar] [CrossRef]
- Jess, T.; Loftus, E.V., Jr.; Harmsen, W.S.; Zinsmeister, A.R.; Tremaine, W.J.; Melton, L.J., 3rd; Munkholm, P.; Sandborn, W.J. Survival and cause specific mortality in patients with inflammatory bowel disease: A long term outcome study in Olmsted County, Minnesota, 1940–2004. Gut 2006, 55, 1248–1254. [Google Scholar] [CrossRef] [Green Version]
- Leone, A.; Nigro, C.; Nicolo, A.; Prevenzano, I.; Formisano, P.; Beguinot, F.; Miele, C. The Dual-Role of Methylglyoxal in Tumor Progression—Novel Therapeutic Approaches. Front. Oncol. 2021, 11, 645686. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Chandel, N.S. We need to talk about the Warburg effect. Nat. Metab. 2020, 2, 127–129. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.A.; Wu, C.H.; Lu, C.C.; Hsia, S.M.; Yen, G.C. Glycative stress from advanced glycation end products (AGEs) and dicarbonyls: An emerging biological factor in cancer onset and progression. Mol. Nutr. Food Res. 2016, 60, 1850–1864. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, F.M.; Allen, R.E.; Thornalley, P.J. Inhibition of proliferation of human leukaemia 60 cells by methylglyoxal in vitro. Biochem. Soc. Trans. 1993, 21, 168S. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Edwards, L.G.; Thornalley, P.J. Effect of methylglyoxal on human leukaemia 60 cell growth: Modification of DNA G1 growth arrest and induction of apoptosis. Leuk. Res. 1996, 20, 397–405. [Google Scholar] [CrossRef]
- Halder, J.; Ray, M.; Ray, S. Inhibition of glycolysis and mitochondrial respiration of Ehrlich ascites carcinoma cells by methylglyoxal. Int. J. Cancer 1993, 54, 443–449. [Google Scholar] [CrossRef]
- Thornalley, P.J. Advances in glyoxalase research. Glyoxalase expression in malignancy, anti-proliferative effects of methylglyoxal, glyoxalase I inhibitor diesters and S-D-lactoylglutathione, and methylglyoxal-modified protein binding and endocytosis by the advanced glycation endproduct receptor. Crit. Rev. Oncol. Hematol. 1995, 20, 99–128. [Google Scholar] [CrossRef]
- Oya-Ito, T.; Naito, Y.; Takagi, T.; Handa, O.; Matsui, H.; Yamada, M.; Shima, K.; Yoshikawa, T. Heat-shock protein 27 (Hsp27) as a target of methylglyoxal in gastrointestinal cancer. Biochim. Biophys. Acta 2011, 1812, 769–781. [Google Scholar] [CrossRef]
- Nokin, M.J.; Durieux, F.; Peixoto, P.; Chiavarina, B.; Peulen, O.; Blomme, A.; Turtoi, A.; Costanza, B.; Smargiasso, N.; Baiwir, D.; et al. Methylglyoxal, a glycolysis side-product, induces Hsp90 glycation and YAP-mediated tumor growth and metastasis. elife 2016, 5, e19375. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Orsini, N.; Wolk, A. Diabetes mellitus and risk of colorectal cancer: A meta-analysis. J. Natl. Cancer Inst. 2005, 97, 1679–1687. [Google Scholar] [CrossRef]
- He, X.K.; Su, T.T.; Si, J.M.; Sun, L.M. Metformin Is Associated With Slightly Reduced Risk of Colorectal Cancer and Moderate Survival Benefits in Diabetes Mellitus: A Meta-Analysis. Medicine 2016, 95, e2749. [Google Scholar] [CrossRef]
- Beisswenger, P.J.; Howell, S.K.; Touchette, A.D.; Lal, S.; Szwergold, B.S. Metformin reduces systemic methylglyoxal levels in type 2 diabetes. Diabetes 1999, 48, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Randell, E.; Han, Y.; Adeli, K.; Krahn, J.; Meng, Q.H. Increased plasma methylglyoxal level, inflammation, and vascular endothelial dysfunction in diabetic nephropathy. Clin. Biochem. 2011, 44, 307–311. [Google Scholar] [CrossRef]
- Ogawa, S.; Nakayama, K.; Nakayama, M.; Mori, T.; Matsushima, M.; Okamura, M.; Senda, M.; Nako, K.; Miyata, T.; Ito, S. Methylglyoxal is a predictor in type 2 diabetic patients of intima-media thickening and elevation of blood pressure. Hypertension 2010, 56, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Lapolla, A.; Flamini, R.; Lupo, A.; Arico, N.C.; Rugiu, C.; Reitano, R.; Tubaro, M.; Ragazzi, E.; Seraglia, R.; Traldi, P. Evaluation of glyoxal and methylglyoxal levels in uremic patients under peritoneal dialysis. Ann. N. Y. Acad. Sci. 2005, 1043, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Brenner, T.; Fleming, T.; Uhle, F.; Silaff, S.; Schmitt, F.; Salgado, E.; Ulrich, A.; Zimmermann, S.; Bruckner, T.; Martin, E.; et al. Methylglyoxal as a new biomarker in patients with septic shock: An observational clinical study. Crit. Care 2014, 18, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nee, J.; Chippendale, R.Z.; Feuerstein, J.D. Screening for Colon Cancer in Older Adults: Risks, Benefits, and When to Stop. Mayo Clin. Proc. 2020, 95, 184–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Low-Grade Dysplasia | High-Grade Dysplasia | Invasive Cancer | Total | p-Value * | |
|---|---|---|---|---|---|
| Number | 11 | 14 | 15 | 40 | |
| Age (years) | 67.91 ± 3.17 | 70.50 ± 2.81 | 66.20 ± 2.83 | 68.18 ± 1.67 | 0.408 |
| Sex | 0.830 | ||||
| Female | 4 (36.36) | 5 (35.71) | 4 (26.67) | 13 (32.50) | |
| Male | 7 (63.64) | 9 (64.29) | 11 (73.33) | 27 (67.50) | |
| Diabetes mellitus | 1 (9.09) | 3 (21.43) | 1 (6.67) | 5 (12.50) | 0.448 |
| Hypertension | 2 (18.18) | 3 (21.43) | 3 (20.00) | 8 (20.00) | 0.980 |
| Smoking | 1 (9.09) | 2 (14.29) | 5 (33.33) | 8 (20.00) | 0.250 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, C.-K.; Yang, P.-C.; Tsai, P.-Y.; Yang, H.-Y.; Tsai, K.-F.; Chen, T.-H.; Liao, K.-S.; Chen, C.-Y.; Lee, J.-A. Methylglyoxal Levels in Human Colorectal Precancer and Cancer: Analysis of Tumor and Peritumor Tissue. Life 2021, 11, 1319. https://doi.org/10.3390/life11121319
Chou C-K, Yang P-C, Tsai P-Y, Yang H-Y, Tsai K-F, Chen T-H, Liao K-S, Chen C-Y, Lee J-A. Methylglyoxal Levels in Human Colorectal Precancer and Cancer: Analysis of Tumor and Peritumor Tissue. Life. 2021; 11(12):1319. https://doi.org/10.3390/life11121319
Chicago/Turabian StyleChou, Chu-Kuang, Po-Chun Yang, Pei-Yun Tsai, Hsin-Yi Yang, Kun-Feng Tsai, Tsung-Hsien Chen, Kai-Sheng Liao, Chi-Yi Chen, and Jen-Ai Lee. 2021. "Methylglyoxal Levels in Human Colorectal Precancer and Cancer: Analysis of Tumor and Peritumor Tissue" Life 11, no. 12: 1319. https://doi.org/10.3390/life11121319