
Survivin as a Therapeutic Target for the Treatment of Human Cancer

1
Department of Medicine, Cedars-Sinai Medical Center, Los Angeles, CA 90048, USA
2
Department of Pathology and Laboratory Medicine, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104, USA
*
Authors to whom correspondence should be addressed.
Cancers 2024, 16(9), 1705; https://doi.org/10.3390/cancers16091705
Submission received: 20 March 2024
/
Revised: 17 April 2024
/
Accepted: 25 April 2024
/
Published: 27 April 2024
(This article belongs to the Collection Innovations in Cancer Drug Development Research)
{kind=link}
{kind=link}
Abstract
:Simple Summary
Survivin is overexpressed in a wide variety of human cancers and is associated with increased chemotherapy resistance, recurrence, and shorter patient survival. Although survivin was first identified as an inhibitor of apoptosis based on its sequence homology, recent studies show that survivin primarily plays a role in the regulation of cell division, as a component of the chromosome passenger complex, which can be localized to the centromere, the spindle midzone, or midbody in a cell cycle-dependent manner. Disruption of survivin function generally leads to mitotic catastrophe and is associated with elevated levels of aneuploidy. In contrast, ectopic expression of survivin can promote cell survival under certain conditions and is implicated in mediating resistance to various cancer drugs, possibly by interactions with molecules that modulate apoptotic pathways. A potential role of survivin in the regulation of mitochondrial functions and processes of autophagy has emerged. Because of its prevalent overexpression in cancer and very limited expression in normal tissues, survivin has been proposed as an ideal therapeutic target, and various approaches have been investigated for survivin inhibition. Here we provide a critical review of our current understanding of the role of survivin in promoting malignancy and strategies for the development of survivin-targeted therapy for cancer.
Abstract
Survivin was initially identified as a member of the inhibitor apoptosis (IAP) protein family and has been shown to play a critical role in the regulation of apoptosis. More recent studies showed that survivin is a component of the chromosome passenger complex and acts as an essential mediator of mitotic progression. Other potential functions of survivin, such as mitochondrial function and autophagy, have also been proposed. Survivin has emerged as an attractive target for cancer therapy because its overexpression has been found in most human cancers and is frequently associated with chemotherapy resistance, recurrence, and poor survival rates in cancer patients. In this review, we discuss our current understanding of how survivin mediates various aspects of malignant transformation and drug resistance, as well as the efforts that have been made to develop therapeutics targeting survivin for the treatment of cancer.
1. Survivin and Cancer
Survivin overexpression has been found in most human cancers and is associated with poor prognosis [1,2,3,4,5,6,7,8]. In particular, high levels of survivin expression are linked with the metastasis of various forms of human cancers, including the presence of circulating tumor cells [9,10,11,12,13]. Overexpression of survivin can facilitate the bypassing of cell cycle checkpoints and promote the survival of aneuploid cells [14,15]. Survivin renders cancer cells resistant to radiation [16,17].
While survivin plays an essential role in early embryogenesis [18,19], its expression levels are very low or undetectable in adult tissues and are usually restricted to stem cells and progenitor cells [15,20,21,22,23]. As shown in conditional knockout mice, survivin is required for T-cell development and homeostasis and triggers p53-dependent cell cycle arrest [24,25]. Similarly, survivin also plays an essential role in B-cell expansion [26]. Moreover, survivin is essential for pancreatic beta cell expansion [27,28,29], early brain development [30], and intestinal epithelial progenitor cells [31].
Survivin, encoded by the BIRC5 gene, is a polypeptide of 142 amino acid residues (Figure 1). The transcription of the BIRC5 gene is mediated through a TATA-less promoter that contains multiple Sp1 sites, a CpG island subjected to potential epigenetic modifications, and cell cycle-dependent element (CDE)/cell cycle homology region (CHR) boxes that mediate cell cycle-dependent gene expression [32,33]. Survivin expression can be upregulated by multiple pathways that are commonly activated in human cancers, such as EGFR, p185Her2/neu, PI-3 kinase, MAPK, NF-κB, and mTOR [34,35,36,37,38]. The transcriptional events from the survivin promoter can also be modulated by Wnt/β-catenin [39], notch [40], YAP [41], and hedgehog signaling pathways [42,43]. In addition, survivin is regulated by Forkhead box m1 (Foxm1), a transcriptional factor critical for G1/S transition and mitotic progression [44]. Conversely, survivin expression can be downregulated by several tumor suppressors, such as TP53, PTEN, Rb, and BRCA1 [45,46,47,48], which are frequently silenced in human cancers.
At least five splicing variants of survivin have been described, which include survivin-ΔEx3 [49,50], survivin-2α [51], survivin-3α [52], survivin-2B [53], and survivin-3B [54]. The differential splicing events lead to the generation of proteins with a shortened BIR domain or truncated polypeptides that do not have the intact NES or the coiled-coil region in the c-terminus, which can exhibit distinct localization patterns. For example, survivin-Ex3 lacks the NES but contains a distinct bipartite nuclear localization signal (NLS) that mediates its localization to the nucleus [49]. In contrast, in survivin-2B, the BIR motif is interrupted by an in-frame insertion of a cryptic exon, generating a protein predominantly localized to the cytoplasm. Survivin-Ex3 and survivin-2B showed reduced affinity to CPC and cannot compensate for the loss of survivin functions [54]. The survivin splicing variants have been reported to be associated with certain transformed phenotypes and clinical outcomes [55]. However, it should be noted that these studies were all focused on the detection of the RNA forms of the splicing variants [49,50,52,53,54]. There has been limited success in the development of antibodies specific for the survivin variants [51,53,54]. Overall, the protein forms of the variants at the endogenous level remain to be convincingly demonstrated in cell lines or pathological specimens. Moreover, it is controversial whether the splicing variants play an important role in mediating cellular functions [54,56,57].
2. Role of Survivin in Cell Division
Survivin plays an essential in cell division, and a loss of survivin leads to mitotic failure and cell death [32,58,59] (Figure 2A). Survivin participates in mitotic checkpoint regulation as a component of the chromosomal passenger complex (CPC) that also includes aurora kinase B, INCENP, TD-60, and borealin [60]. The BIR domain of survivin can interact with a phosphorylated form of histone H3 (Histone H3T3ph), which is mediated by the histone H3-associated kinase HASPIN and required for recruitment of the other CPC proteins to the inner centromere [61,62,63]. Interference with the survivin–histone H3T3ph interaction leads to mislocalized aurora kinase B and mitotic defects [61,62,63]. Upon entry into mitosis, survivin is localized to the centromere region in a manner that is dependent on the inner centromere protein (INCENP) and cooperates with Aurora kinase B and other CPC proteins to modulate spindle formation and proper chromosome alignment [64,65]. Thus, the essential role of survivin in CPC assembly and mitotic progression, mediated by the BIR domain, may be exploited for the development of survivin-targeting therapeutics.
When chromosome segregation occurs at the initiation of anaphase, survivin is separated from the centromere but remains in the spindle midzone, subsequently becoming associated with the midbody. The molecular mechanism involved in the relocation of survivin to the midbody is not well understood. Nonetheless, survivin has been shown to interact with non-muscle myosin II, and midbody-localized survivin is implicated in playing a role in the formation of the contractile ring during cytokinesis [66]. Survivin abnormality is commonly accompanied by aneuploidy [32,58,59,67,68], which supports the notion that it plays an essential role in the regulation of mitotic checkpoints and cytokinesis. Notably, a loss of p53 function is required for re-entry into the cell cycle following the depletion of survivin [69,70].
3. Role of Survivin in Apoptosis
Survivin was first identified as a member of the Inhibitor-of-Apoptosis (IAP) protein family, also known as the Baculoviral IAP repeat-containing (BIRC) proteins, based on the presence of a Baculovirus IAP Repeat (BIR) in the N-terminus [32,58] (Figure 1). The IAP family proteins share the common feature of having at least one BIR domain, which consists of ~70 amino acid residues and is involved in mediating protein–protein interactions [71]. While ablation of survivin can lead to apoptosis, overexpression of survivin can protect cells from apoptosis under various experimental conditions [32,58,72,73] (Figure 2B).
Some IAP family proteins can inhibit apoptosis by directly binding to the activated form of caspase and blocking its enzymatic activities [74,75]. For example, the second BIR domain and a linker region of XIAP can directly bind to caspase and hinder substrate access [76,77,78,79]. In addition, some IAPs also contain either a RING domain—which functions as an E3 ubiquitin ligase—or a ubiquitin-associated domain, which mediates the ubiquitin-mediated proteolytic degradation of caspase [80]. In comparison, survivin contains only a BIR domain, and, to date, no compelling evidence is available to show that survivin can directly bind and inhibit caspase activities (Figure 2B).
Survivin has been reported to bind to the mitochondrial protein DIABLO/SMAC [81,82]. DIABLO/SMAC can potentiate certain forms of apoptosis by blocking the action of IAPs and thereby activating caspases [83,84]. Thus, it has been proposed that survivin may inhibit apoptosis by neutralizing the capacities of DIABLO/SMAC to promote apoptotic signaling (Figure 2B). However, it remains to demonstrate that, as endogenous expression levels, the survivin-DIABLO interactions indeed participate in protection from apoptosis [85].
4. Role of Survivin in Mitochondrial Function and Autophagy
Survivin has been shown to inhibit mitochondrial-dependent apoptotic events [73]. It has been reported that the N-terminus of survivin contains a mitochondria-targeting sequence that can direct protein localization to the mitochondria when fused with a reporter protein [86] (Figure 1). Interestingly, survivin can be detected in the mitochondrial fraction in cancer cells but not in non-transformed cells [87,88], which suggests that the role of survivin in mitochondria function may be dependent on cell content, including the genetic or epigenetic background. The overexpression of mitochondria-targeted survivin can protect cells from apoptosis and enhance transformation (Figure 2C), which may involve its binding to another IAP family member XIAP [87,89]. The localization of survivin to the mitochondria can also promote cancer cell invasion and metastasis [90]. The overexpression of survivin appears to alter the dynamic of mitochondrial fission and fusion [91] or inhibit mitophagy [88].
Paradoxically, it has been reported that both the knockdown [90] and overexpression of survivin [88,91] can disable mitochondrial functions and reduce oxidative phosphorylation in cancer cells. It should be noted that the studies on mitochondrial survivin were conducted using a fusion protein of survivin with the mitochondrial targeting sequence of cytochrome c, and the knockdown approach used in the study does not specifically target the mitochondrial pool of the protein [87,88,90]. Clearly, additional work is needed to unravel the mechanism and the biological significance of mitochondrial localization of survivin.
Survivin has been reported to physically interact with several proteins that are involved in autophagy (Figure 2D), a process by which cancer cells can adapt to physiological or pathological challenges by degrading and recycling subcellular components of the cell [92]. For example, beclin, a key regulator of autophagy [93], was found to bind to survivin, which may be involved in regulating survivin protein levels [94]. Intriguingly, ATG5, a protein known for its role in the formation and elongation of autophagosome [95], was reported to form a complex with survivin in the nucleus upon exposure to DNA damage, leading to mitotic catastrophe in an autophagy-independent manner [96]. Conversely, the interaction between survivin and ATG5/ATG12 may also impact autophagy-mediated events responsive to DNA damage [97]. The induced ectopic expression of survivin appears to be required for autophagy induced by the inhibition of glycolysis [98]. However, these observations were made only in cell lines with forced overexpression of survivin. It remains to determine whether survivin directly participates in the regulation of autophagic processes.
5. Survivin Localization
Survivin contains a nuclear export signal (NES) that binds to the nuclear export receptor Crm1, which is required for survivin cytoplasmic localization during the interphase [49,99,100,101]. Alteration of the NES, which is located between BIR and the c-terminal coiled-coil structure, can disrupt the nuclear export and localization of survivin to the centromere or to the midbody but not its homodimerization or binding to several CPC proteins [100,101,102]. The NES mutations caused a shift from a cytoplasmic localization pattern to a nuclear one, which is associated with the loss of survivin function to protect cells from apoptosis induced by genotoxic damage or external stimulus [99,101]. In addition, the nucleus-directed survivin protein appears to enhance cancer cell sensitivity to apoptosis [103,104].
These observations led to the notion that cytoplasmic survivin is primarily involved in protection from apoptosis. However, it should be noted that the survivin localization patterns in the cytoplasm or nucleus may not be a reliable biomarker for clinical outcomes, as it has been linked to both favorable [8,105,106,107] and unfavorable prognosis of cancer patients [108,109,110,111,112,113].
6. Survivin Protein Structure and Post-Translational Modification
The survivin protein structure in the form of a homodimer has been determined by both crystallography [114,115,116] and nuclear magnetic resonance (NMR) [117]. The N-terminal BIR domain consists of a three-stranded β-sheet and four α-helices, with a zinc-binding fold, and the survivin protein forms a bow-tie-shaped dimer via part of the N-terminal region and the linker region between the BIR domain and the C-terminal helix [114,115]. Notably, the ubiquitination of survivin on several lysine residues within the BIR domain is implicated in playing a role in modulating its localization to the centromere [118].
The structure of survivin that features a heteromeric complex formed with borealin and INCENP has also been resolved [119]. In this structure, the C-terminus of survivin, which contains an extended α-helical coiled-coil domain, forms a three-helical bundle with elements of borealin and INCENP in 1:1:1 stoichiometry [119]. These interactions are essential for the central spindle and midbody localization of the complex. More recently, the crystal structure of survivin with the N-terminal tail of histone H3 has also been reported, which identified structural features in the BIR domain that are important for binding to histone H3T3ph [120].
An accumulating body of evidence shows that survivin can be regulated by phosphorylation. Phosphorylation of survivin at threonine 34 by CDK1 has been shown to be important for its anti-apoptotic role [121,122]. In addition, phosphorylation by PKA at serine 20 is also involved in protection against apoptosis by mediating the interaction with XIAP [89]. Moreover, survivin can be phosphorylated by aurora B at threonine 117 and negatively regulates its localization to the centromere region and function in mitosis [123]. CK2 can phosphorylate survivin at threonine 48 in the BIR domain, which is critical for its mitotic and antiapoptotic functions [124]. Furthermore, PLK1 phosphorylates survivin at threonine 20, which seems to be involved in chromosome orientation during mitosis [125].
Survivin is subject to other forms of post-translational modification. For example, survivin can be acetylated at lysine 129 (K129), which affects its homodimerization, binding to Crm1, and nuclear export [126]. Survivin can also be modified via K48- and K63-linked ubiquitination during mitosis, which mediates survivin localization to the centromere and mitotic progression [118].
7. Therapeutic Strategies to Target Survivin
Efforts have been made in recent years to develop therapeutic strategies to disable survivin functionally. However, suvivin is an unconventional drug target, due to its unique structure and lack of enzymatic activity. It is important to include cell-based assays that evaluate the phenotypic changes affected by inhibition of survivin. Because survivin is required for cell division and knockdown of survivin generally causes mitotic failure [32,58,59,67,68], it is plausible that the gross inhibition of survivin by small molecules would lead to similar effects. Currently, only a limited number of survivin inhibitors have been developed with success.
YM155, an imidazolium-based compound identified by a high-throughput screen, is one of the first small-molecule antagonists known for its ability to inhibit survivin expression. YM155 targets the survivin promoter region to inhibit gene transcription [127]. Preclinical research showed that YM155 can effectively decrease survivin expression levels in various cancer cell lines and inhibit tumor growth in xenografts mouse models, including prostate cancer, non-Hodgkin lymphoma, and lung cancer [127,128]. Several phase I and phase II studies showed that YM155 generally shows low toxicity but has limited antitumor efficacy when used as either a single agent or in combination with other therapeutic agents [129,130,131,132,133,134]. However, recent studies indicate that YM155 can elicit DNA damage in cells [135,136,137], which indicates that the compound may target other proteins and signaling events. Indeed, when tested in vitro, YM155 induces cell death without causing any delay in mitosis, despite that it significantly reduces survivin expression [135,136]. These findings suggest that the primary mechanism by which YM155 induces cytotoxicity is likely not through disabling survivin functions.
Several other molecules that suppress survivin expression levels have been described. For example, FL-118 has been identified by an HTS screen of a library of compounds as a molecule that can reduce expression levels of survivin, as well as those of other IAP family members [138]. FL-118-induced cell death accompanies a reduction in BrdU-incorporating cells, but does not with any effect on mitosis [138]. By using a similar approach to screen for drugs that can inhibit survivin promoter activities, a cytotoxic molecule, termed WM127, was also found to be capable of reducing survivin expression levels [139]. WM127 reduces cancer cell proliferation and causes an accumulation of cells in the G2/M stage of the cell cycle [139], although further analysis of its effect on mitotic progression remains to be carried out. In addition, EM-1421 (also known as terameprocol) has been described as a small molecule that targets Sp1-dependent promoters and reduces the expression levels of survivin and the mitosis regulator cdk1 [140,141,142]. Moreover, GDP366 is another small molecule that can reduce survivin expression at both mRNA and protein levels and increase aberrant cell division and polyploid cells [143]. However, because GDP366 can also inhibit the expression of stathmin 1 [143], which encodes a protein that mediates the dynamics of the microtubule network and mitotic progression [144,145,146], the mechanism of action by this molecule remains to be clarified. Thus, these efforts to target survivin expression have identified molecules that are not specific for survivin.
Strategies designed to reduce survivin protein stability have been reported. Heat shock protein 90 can bind to survivin and protect it from proteasomal degradation, and the disruption of this interaction can lead to apoptosis [147]. Sheperdin, a peptidomimetic derived from the survivin region that is sufficient to bind to Hsp90, showed the ability to bind the ATP pocket of HSP90 and disrupt the interaction with several of its client proteins, including survivin [148]. This causes the degradation of survivin, among other proteins, leading to apoptosis in tumor cells [148]. Of note, sheperdin treatment caused rapid cell death without triggering any apparent delay in mitosis [148].
In another example, using a virtual screen of compounds that mimic the DIABLO/SMAC-IAP interaction, a series of small-molecule IAP inhibitors have been developed [149,150]. These molecules can inhibit survivin and, to a lesser extent, XIAP, by downregulating their protein levels and showed efficacy to inhibit tumor growth [149,150]. Other small molecules that block the interaction between survivin and DIABLO/SMAC have been described and showed anti-cancer activities [151,152].
A high-throughput, affinity-based NMR screen has led to the identification of several survivin-binding molecules that bind to the dimer interface [153,154,155,156]. Several of the compounds identified in the screen displayed activities to inhibit the growth of tumor cells and appeared to cause cell cycle delay in the G0/G1 stage, rather than in the mitotic stage [154]. One of the molecules was shown to sensitize colon cancer cells to topoisomerase inhibitor irinotecan [156].
We employed a unique structure-based approach to identify survivin inhibitors that bind directly to the protein and modulate its functions [157,158]. The method, termed Cavity-Induced Allosteric Modulation (CIAM), was previously used to successfully develop inhibitors for other targets, such as TNFR1 [159]. With the CIAM method, we have identified a cavity close to the survivin dimeric interface. The compounds that fit into this cavity in silico were further tested for the ability to bind the survivin protein and affect mitotic arrest, as one would expect from a loss of survivin function. Several compounds identified by this approach, including S12, exhibit efficacy to inhibit the growth of human cancer cell lines both in vitro and in vivo [26,43,157,158]. This was the first set of small-molecule inhibitors that have been shown to directly bind to the intended target site in the survivin protein and cause phenotypic changes in cancer cell cells that are consistent with what is expected from loss of survivin function. Notably, the knockdown of YAP can increase the sensitivity of cancer cells to S12 [158], which suggests that simultaneously targeting survivin and YAP may achieve enhanced therapeutic effects. Finally, S12 has been modified to be potentially used for imaging survivin in tumors by single-photon emission computerized tomography [160].
More recently, a separate group performed a virtual screen of molecules that target the survivin dimeric interface and identified a series of molecules (e.g., LQZ-7F and LQZ-7I) that can induce the proteasomal degradation of survivin [161,162]. These molecules were also shown to cause apoptosis and inhibit tumor growth in xenograft tumor models [161,162]. It is not clear how disruption of the survivin homodimer by these small molecules leads to a reduction in protein stability. The effect of these molecules on cell cycle progression has not been well characterized [161,162].
Survivin-based immunotherapy has been developed. The Cytotoxic T lymphocyte (CTL) response to survivin can be detected in patients [163]. The survivin-derived, MHC-restricted T cell epitope has been identified and can be harnessed to trigger the CTL response against survivin-expressing cancer cells [164]. In addition, DNA vaccine-encoding survivin and CCL12 can trigger a strong immune response against lung cancer cells in an animal model [165]. Recent studies showed that long synthetic peptides derived from the survivin protein can generate both cytotoxic CD8+ and CD4+ T-cell responses, leading to tumor regression and the prevention of relapse in animal models [166]. In particular, the survivin peptide mimic SurVaxM showed efficacy in stimulating anti-tumor immune responses against brain tumors in animal models and early promise in clinical trials [167,168,169]. A whole protein survivin dendritic cell vaccine has also been developed and tested for the treatment of myeloma patients [170,171].
8. Conclusions and Perspective
Because of its high expression levels in most tumors and absence in most normal tissues, survivin has been considered a promising therapeutic target. Studies in the past 25 years have established that survivin plays a vital role in the regulation of cell division as a component of the CPC in the mitotic apparatus. However, despite its structural similarities to the other IAP family proteins, how survivin acts as an inhibitor of apoptosis remains elusive. The overexpression of survivin may lead to aberrant localization of the protein, which can contribute to aspects of tumorigenesis in a cell content-dependent manner. Studies on survivin protein ensembles in various subcellular pools, such as the mitochondria and the interphase or mitotic cytoplasm, may unravel a mechanism by which survivin links mitotic checkpoint regulation with apoptotic pathways. Progress has been made to develop survivin-targeted therapy, including small molecules that directly bind and disable survivin. With the advent of artificial intelligence-aided structural modeling and drug design, it is expected that more survivin-targeting entities will be available for testing. It should be mentioned that well-designed biophysical analysis and cell-based assays are critical for the identification of small molecules that elicit more specific anti-survivin activities. Finally, understanding the signaling pathways that determine cancer cell sensitivity to survivin-targeted therapy may help to develop more effective therapeutic strategies. Conceivably, the combination of survivin inhibition with chemotherapy or other targeted therapeutics may achieve the maximal clinical benefit.
Author Contributions
Conceptualization, Q.W. and M.I.G.; writing—original draft preparation, Q.W.; writing—review and editing, Q.W. and M.I.G.; funding acquisition, M.I.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tanaka, K.; Iwamoto, S.; Gon, G.; Nohara, T.; Iwamoto, M.; Tanigawa, N. Expression of survivin and its relationship to loss of apoptosis in breast. Clin. Cancer Res. 2000, 6, 127–134. [Google Scholar] [PubMed]
- Cohen, C.; Lohmann, C.M.; Cotsonis, G.; Lawson, D.; Santoianni, R. Survivin expression in ovarian carcinoma: Correlation with apoptotic markers and. Mod. Pathol. 2003, 16, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Sarela, A.I.; Verbeke, C.S.; Ramsdale, J.; Davies, C.L.; Markham, A.F.; Guillou, P.J. Expression of survivin, a novel inhibitor of apoptosis and cell cycle regulatory. Br. J. Cancer 2002, 86, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, P.; Kuhnel, T.; Muhr-Wilkenshoff, F.; Heine, B.; Stein, H.; Hopfner, M.; Germer, C.T.; Scherubl, H. Prognostic value of nuclear survivin expression in oesophageal squamous cell. Br. J. Cancer 2003, 88, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Sui, L.; Dong, Y.; Ohno, M.; Watanabe, Y.; Sugimoto, K.; Tokuda, M. Survivin expression and its correlation with cell proliferation and prognosis in epithelial ovarian tumors. Int. J. Oncol. 2002, 21, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Kato, J.; Kuwabara, Y.; Mitani, M.; Shinoda, N.; Sato, A.; Toyama, T.; Mitsui, A.; Nishiwaki, T.; Moriyama, S.; Kudo, J.; et al. Expression of survivin in esophageal cancer: Correlation with the prognosis and response to chemotherapy. Int. J. Cancer 2001, 95, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Oparina, N.; Erlandsson, M.C.; Faldt Beding, A.; Parris, T.; Helou, K.; Karlsson, P.; Einbeigi, Z.; Bokarewa, M.I. Prognostic Significance of BIRC5/Survivin in Breast Cancer: Results from Three Independent Cohorts. Cancers 2021, 13, 2209. [Google Scholar] [CrossRef] [PubMed]
- Tonini, G.; Vincenzi, B.; Santini, D.; Scarpa, S.; Vasaturo, T.; Malacrino, C.; Coppola, R.; Magistrelli, P.; Borzomati, D.; Baldi, A.; et al. Nuclear and cytoplasmic expression of survivin in 67 surgically resected pancreatic cancer patients. Br. J. Cancer 2005, 92, 2225–2232. [Google Scholar] [CrossRef]
- Cao, M.; Yie, S.M.; Wu, S.M.; Chen, S.; Lou, B.; He, X.; Ye, S.R.; Xie, K.; Rao, L.; Gao, E.; et al. Detection of survivin-expressing circulating cancer cells in the peripheral blood of patients with esophageal squamous cell carcinoma and its clinical significance. Clin. Exp. Metastasis 2009, 26, 751–758. [Google Scholar] [CrossRef]
- Yie, S.M.; Lou, B.; Ye, S.R.; He, X.; Cao, M.; Xie, K.; Ye, N.Y.; Lin, R.; Wu, S.M.; Xiao, H.B.; et al. Clinical significance of detecting survivin-expressing circulating cancer cells in patients with non-small cell lung cancer. Lung Cancer 2009, 63, 284–290. [Google Scholar] [CrossRef]
- Goossens-Beumer, I.J.; Zeestraten, E.C.; Benard, A.; Christen, T.; Reimers, M.S.; Keijzer, R.; Sier, C.F.; Liefers, G.J.; Morreau, H.; Putter, H.; et al. Clinical prognostic value of combined analysis of Aldh1, Survivin, and EpCAM expression in colorectal cancer. Br. J. Cancer 2014, 110, 2935–2944. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, T.; Liu, Q.; Jiao, H.; Yang, W.; Liu, X.; Huo, Z. Clinical and prognostic significance of HIF-1alpha, PTEN, CD44v6, and survivin for gastric cancer: A meta-analysis. PLoS ONE 2014, 9, e91842. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Hanna, D.L.; Zhang, W.; Mendez, A.; Yang, D.; El-Khoueiry, R.; Matsusaka, S.; Sunakawa, Y.; Stremitzer, S.; Parekh, A.; et al. Cytokeratin-20 and Survivin-Expressing Circulating Tumor Cells Predict Survival in Metastatic Colorectal Cancer Patients by a Combined Immunomagnetic qRT-PCR Approach. Mol. Cancer Ther. 2015, 14, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Rosa, J.; Canovas, P.; Islam, A.; Altieri, D.C.; Doxsey, S.J. Survivin modulates microtubule dynamics and nucleation throughout the cell cycle. Mol. Biol. Cell 2006, 17, 1483–1493. [Google Scholar] [CrossRef] [PubMed]
- Gianani, R.; Jarboe, E.; Orlicky, D.; Frost, M.; Bobak, J.; Lehner, R.; Shroyer, K.R. Expression of survivin in normal, hyperplastic, and neoplastic colonic mucosa. Hum. Pathol. 2001, 32, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, A.; Zhai, G.G.; Zhang, M.; Malhotra, R.; Latham, D.E.; Delaney, M.A.; Robe, P.; Nestler, U.; Song, Q.; Loeffler, J. Survivin enhances radiation resistance in primary human glioblastoma cells via caspase-independent mechanisms. Oncogene 2004, 23, 7494–7506. [Google Scholar] [CrossRef] [PubMed]
- Rodel, F.; Hoffmann, J.; Distel, L.; Herrmann, M.; Noisternig, T.; Papadopoulos, T.; Sauer, R.; Rodel, C. Survivin as a radioresistance factor, and prognostic and therapeutic target for radiotherapy in rectal cancer. Cancer Res. 2005, 65, 4881–4887. [Google Scholar] [CrossRef] [PubMed]
- Uren, A.G.; Wong, L.; Pakusch, M.; Fowler, K.J.; Burrows, F.J.; Vaux, D.L.; Choo, K.H. Survivin and the inner centromere protein INCENP show similar cell-cycle localization and gene knockout phenotype. Curr. Biol. 2000, 10, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.M.; Pollefeyt, S.; Steiner-Mosonyi, M.; Luo, W.; Devriese, A.; Lupu, F.; Bono, F.; Leducq, N.; Dol, F.; Schaeffer, P.; et al. Deficiency of survivin in transgenic mice exacerbates Fas-induced apoptosis via mitochondrial pathways. Gastroenterology 2002, 123, 619–631. [Google Scholar] [CrossRef]
- Ambrosini, G.; Adida, C.; Altieri, D.C. A novel anti-apoptosis gene, survivin, expressed in cancer and lymphoma. Nat. Med. 1997, 3, 917–921. [Google Scholar] [CrossRef]
- Zhang, T.; Otevrel, T.; Gao, Z.; Ehrlich, S.M.; Fields, J.Z.; Boman, B.M. Evidence that APC regulates survivin expression: A possible mechanism. Cancer Res. 2001, 61, 8664–8667. [Google Scholar] [PubMed]
- Fukuda, S.; Pelus, L.M. Regulation of the inhibitor-of-apoptosis family member survivin in normal cord. Blood 2001, 98, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Pelus, L.M. Survivin, a cancer target with an emerging role in normal adult tissues. Mol. Cancer Ther. 2006, 5, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Conway, E.M.; Kang, C.; Winoto, A. Essential role of survivin, an inhibitor of apoptosis protein, in T cell development, maturation, and homeostasis. J. Exp. Med. 2004, 199, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Bakal, C.; Shahinian, A.; Elia, A.; Wakeham, A.; Suh, W.K.; Duncan, G.S.; Ciofani, M.; Rottapel, R.; Zuniga-Pflucker, J.C.; et al. Survivin loss in thymocytes triggers p53-mediated growth arrest and p53-independent cell death. J. Exp. Med. 2004, 199, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Miletic, A.V.; Jellusova, J.; Cato, M.H.; Lee, C.R.; Baracho, G.V.; Conway, E.M.; Rickert, R.C. Essential Role for Survivin in the Proliferative Expansion of Progenitor and Mature B Cells. J. Immunol. 2016, 196, 2195–2204. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Nishimura, W.; Devor-Henneman, D.; Kusewitt, D.; Wang, H.; Holloway, M.P.; Dohi, T.; Sabo, E.; Robinson, M.L.; Altieri, D.C.; et al. Postnatal expansion of the pancreatic beta-cell mass is dependent on survivin. Diabetes 2008, 57, 2718–2727. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, L.; Schroer, S.; Choi, D.; Chen, P.; Okada, H.; Woo, M. Perinatal survivin is essential for the establishment of pancreatic beta cell mass in mice. Diabetologia 2009, 52, 2130–2141. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, Q.; Wang, X.; Zhu, J.; Xu, K.; Okada, H.; Wang, R.; Woo, M. Survivin is required for beta-cell mass expansion in the pancreatic duct-ligated mouse model. PLoS ONE 2012, 7, e41976. [Google Scholar] [CrossRef]
- Jiang, Y.; de Bruin, A.; Caldas, H.; Fangusaro, J.; Hayes, J.; Conway, E.M.; Robinson, M.L.; Altura, R.A. Essential role for survivin in early brain development. J. Neurosci. 2005, 25, 6962–6970. [Google Scholar] [CrossRef]
- Martini, E.; Wittkopf, N.; Gunther, C.; Leppkes, M.; Okada, H.; Watson, A.J.; Podstawa, E.; Backert, I.; Amann, K.; Neurath, M.F.; et al. Loss of Survivin in Intestinal Epithelial Progenitor Cells Leads to Mitotic Catastrophe and Breakdown of Gut Immune Homeostasis. Cell Rep. 2016, 14, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ambrosini, G.; Chu, E.Y.; Plescia, J.; Tognin, S.; Marchisio, P.C.; Altieri, D.C. Control of apoptosis and mitotic spindle checkpoint by survivin. Nature 1998, 396, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Altieri, D.C. Transcriptional analysis of human survivin gene expression. Biochem. J. 1999, 344 Pt 2, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Vaira, V.; Lee, C.W.; Goel, H.L.; Bosari, S.; Languino, L.R.; Altieri, D.C. Regulation of survivin expression by IGF-1/mTOR signaling. Oncogene 2007, 26, 2678–2684. [Google Scholar] [CrossRef] [PubMed]
- Asanuma, H.; Torigoe, T.; Kamiguchi, K.; Hirohashi, Y.; Ohmura, T.; Hirata, K.; Sato, M.; Sato, N. Survivin expression is regulated by coexpression of human epidermal growth factor. Cancer Res. 2005, 65, 11018–11025. [Google Scholar] [CrossRef] [PubMed]
- Tracey, L.; Perez-Rosado, A.; Artiga, M.J.; Camacho, F.I.; Rodriguez, A.; Martinez, N.; Ruiz-Ballesteros, E.; Mollejo, M.; Martinez, B.; Cuadros, M.; et al. Expression of the NF-kappaB targets BCL2 and BIRC5/Survivin characterizes small. J. Pathol. 2005, 206, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Greene, M.I. EGFR enhances Survivin expression through the phosphoinositide 3 (PI-3) kinase. Exp. Mol. Pathol. 2005, 79, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Bisi, J.; Strum, J.; Liu, L.; Carrick, K.; Graham, K.M.; Treece, A.L.; Hardwicke, M.A.; Dush, M.; Liao, Q.; et al. Regulation of survivin by ErbB2 signaling: Therapeutic implications for. Cancer Res. 2006, 66, 1640–1647. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.J.; Plescia, J.; Clevers, H.; Fearon, E.R.; Altieri, D.C. Survivin and molecular pathogenesis of colorectal cancer. Lancet 2003, 362, 205–209. [Google Scholar] [CrossRef]
- Lee, C.W.; Raskett, C.M.; Prudovsky, I.; Altieri, D.C. Molecular dependence of estrogen receptor-negative breast cancer on a notch-survivin signaling axis. Cancer Res. 2008, 68, 5273–5281. [Google Scholar] [CrossRef]
- Dong, J.; Feldmann, G.; Huang, J.; Wu, S.; Zhang, N.; Comerford, S.A.; Gayyed, M.F.; Anders, R.A.; Maitra, A.; Pan, D. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell 2007, 130, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Vlckova, K.; Ondrusova, L.; Vachtenheim, J.; Reda, J.; Dundr, P.; Zadinova, M.; Zakova, P.; Pouckova, P. Survivin, a novel target of the Hedgehog/GLI signaling pathway in human tumor cells. Cell Death Dis. 2016, 7, e2048. [Google Scholar] [CrossRef] [PubMed]
- Brun, S.N.; Markant, S.L.; Esparza, L.A.; Garcia, G.; Terry, D.; Huang, J.M.; Pavlyukov, M.S.; Li, X.N.; Grant, G.A.; Crawford, J.R.; et al. Survivin as a therapeutic target in Sonic hedgehog-driven medulloblastoma. Oncogene 2015, 34, 3770–3779. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.C.; Chen, Y.J.; Hughes, D.; Petrovic, V.; Major, M.L.; Park, H.J.; Tan, Y.; Ackerson, T.; Costa, R.H. Forkhead box M1 regulates the transcriptional network of genes essential for mitotic progression and genes encoding the SCF (Skp2-Cks1) ubiquitin ligase. Mol. Cell Biol. 2005, 25, 10875–10894. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.; McGuirk, M.; Hockenberry, T.N.; Wu, Q.; Ashar, H.; Black, S.; Wen, S.F.; Wang, L.; Kirschmeier, P.; Bishop, W.R.; et al. Human survivin is negatively regulated by wild-type p53 and participates in. Oncogene 2002, 21, 2613–2622. [Google Scholar] [CrossRef] [PubMed]
- Raj, D.; Liu, T.; Samadashwily, G.; Li, F.; Grossman, D. Survivin repression by p53, Rb and E2F2 in normal human melanocytes. Carcinogenesis 2008, 29, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Plescia, J.; Leav, I.; Li, J.; Languino, L.R.; Altieri, D.C. Endogenous tumor suppression mediated by PTEN involves survivin gene silencing. Cancer Res. 2009, 69, 4954–4958. [Google Scholar] [CrossRef]
- Wang, R.H.; Zheng, Y.; Kim, H.S.; Xu, X.; Cao, L.; Luhasen, T.; Lee, M.H.; Xiao, C.; Vassilopoulos, A.; Chen, W.; et al. Interplay among BRCA1, SIRT1, and Survivin during BRCA1-associated tumorigenesis. Mol. Cell 2008, 32, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.A.; Span, S.W.; Ferreira, C.G.; Kruyt, F.A.; Giaccone, G. CRM1-mediated nuclear export determines the cytoplasmic localization of the antiapoptotic protein Survivin. Exp. Cell Res. 2002, 275, 44–53. [Google Scholar] [CrossRef]
- Mahotka, C.; Wenzel, M.; Springer, E.; Gabbert, H.E.; Gerharz, C.D. Survivin-deltaEx3 and survivin-2B: Two novel splice variants of the apoptosis inhibitor survivin with different antiapoptotic properties. Cancer Res. 1999, 59, 6097–6102. [Google Scholar]
- Caldas, H.; Honsey, L.E.; Altura, R.A. Survivin 2alpha: A novel Survivin splice variant expressed in human malignancies. Mol. Cancer 2005, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Mola, G.; Vela, E.; Fernandez-Figueras, M.T.; Isamat, M.; Munoz-Marmol, A.M. Exonization of Alu-generated splice variants in the survivin gene of human and non-human primates. J. Mol. Biol. 2007, 366, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Caldas, H.; Jiang, Y.; Holloway, M.P.; Fangusaro, J.; Mahotka, C.; Conway, E.M.; Altura, R.A. Survivin splice variants regulate the balance between proliferation and cell death. Oncogene 2005, 24, 1994–2007. [Google Scholar] [CrossRef] [PubMed]
- Noton, E.A.; Colnaghi, R.; Tate, S.; Starck, C.; Carvalho, A.; Ko Ferrigno, P.; Wheatley, S.P. Molecular analysis of survivin isoforms: Evidence that alternatively spliced variants do not play a role in mitosis. J. Biol. Chem. 2006, 281, 1286–1295. [Google Scholar] [CrossRef]
- Pavlidou, A.; Kroupis, C.; Dimas, K. Association of survivin splice variants with prognosis and treatment of breast cancer. World J. Clin. Oncol. 2014, 5, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Kappler, M.; Rot, S.; Taubert, H.; Greither, T.; Bartel, F.; Dellas, K.; Hansgen, G.; Trott, K.R.; Bache, M. The effects of knockdown of wild-type survivin, survivin-2B or survivin-delta3 on the radiosensitization in a soft tissue sarcoma cells in vitro under different oxygen conditions. Cancer Gene Ther. 2007, 14, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Vivas-Mejia, P.E.; Rodriguez-Aguayo, C.; Han, H.D.; Shahzad, M.M.; Valiyeva, F.; Shibayama, M.; Chavez-Reyes, A.; Sood, A.K.; Lopez-Berestein, G. Silencing survivin splice variant 2B leads to antitumor activity in taxane-resistant ovarian cancer. Clin. Cancer Res. 2011, 17, 3716–3726. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ackermann, E.J.; Bennett, C.F.; Rothermel, A.L.; Plescia, J.; Tognin, S.; Villa, A.; Marchisio, P.C.; Altieri, D.C. Pleiotropic cell-division defects and apoptosis induced by interference with survivin function. Nat. Cell Biol. 1999, 1, 461–466. [Google Scholar] [CrossRef]
- Lens, S.M.; Wolthuis, R.M.; Klompmaker, R.; Kauw, J.; Agami, R.; Brummelkamp, T.; Kops, G.; Medema, R.H. Survivin is required for a sustained spindle checkpoint arrest in response to lack of tension. Embo J. 2003, 22, 2934–2947. [Google Scholar] [CrossRef]
- Vader, G.; Kauw, J.J.; Medema, R.H.; Lens, S.M. Survivin mediates targeting of the chromosomal passenger complex to the centromere and midbody. EMBO Rep. 2006, 7, 85–92. [Google Scholar] [CrossRef]
- Kelly, A.E.; Ghenoiu, C.; Xue, J.Z.; Zierhut, C.; Kimura, H.; Funabiki, H. Survivin reads phosphorylated histone H3 threonine 3 to activate the mitotic kinase Aurora B. Science 2010, 330, 235–239. [Google Scholar] [CrossRef]
- Wang, F.; Dai, J.; Daum, J.R.; Niedzialkowska, E.; Banerjee, B.; Stukenberg, P.T.; Gorbsky, G.J.; Higgins, J.M. Histone H3 Thr-3 phosphorylation by Haspin positions Aurora B at centromeres in mitosis. Science 2010, 330, 231–235. [Google Scholar] [CrossRef]
- Yamagishi, Y.; Honda, T.; Tanno, Y.; Watanabe, Y. Two histone marks establish the inner centromere and chromosome bi-orientation. Science 2010, 330, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, S.P.; Carvalho, A.; Vagnarelli, P.; Earnshaw, W.C. INCENP is required for proper targeting of Survivin to the centromeres and the anaphase spindle during mitosis. Curr. Biol. 2001, 11, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.A.; Lan, W.; Powers, S.E.; McCleland, M.L.; Kuang, J.; Stukenberg, P.T. Aurora B kinase exists in a complex with survivin and INCENP and its kinase activity is stimulated by survivin binding and phosphorylation. Mol. Biol. Cell 2002, 13, 3064–3077. [Google Scholar] [CrossRef]
- Babkoff, A.; Cohen-Kfir, E.; Aharon, H.; Ronen, D.; Rosenberg, M.; Wiener, R.; Ravid, S. A direct interaction between survivin and myosin II is required for cytokinesis. J. Cell Sci. 2019, 132, jcs233130. [Google Scholar] [CrossRef]
- Sumi, T.; Hirai, S.; Yamaguchi, M.; Tanaka, Y.; Tada, M.; Yamada, G.; Hasegawa, T.; Miyagi, Y.; Niki, T.; Watanabe, A.; et al. Survivin knockdown induces senescence in TTF-1-expressing, KRAS-mutant lung adenocarcinomas. Int. J. Oncol. 2018, 53, 33–46. [Google Scholar] [CrossRef]
- Garlapati, C.; Joshi, S.; Bhattarai, S.; Krishnamurthy, J.; Turaga, R.C.; Nguyen, T.; Li, X.; Aneja, R. PLK1 and AURKB phosphorylate survivin differentially to affect proliferation in racially distinct triple-negative breast cancer. Cell Death Dis. 2023, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Welm, A.; Bishop, J.M. Cell division and cell survival in the absence of survivin. Proc. Natl. Acad. Sci. USA 2004, 101, 15100–15105. [Google Scholar] [CrossRef]
- Beltrami, E.; Plescia, J.; Wilkinson, J.C.; Duckett, C.S.; Altieri, D.C. Acute ablation of survivin uncovers p53-dependent mitotic checkpoint functions and control of mitochondrial apoptosis. J. Biol. Chem. 2004, 279, 2077–2084. [Google Scholar] [CrossRef]
- Srinivasula, S.M.; Ashwell, J.D. IAPs: What’s in a name? Mol. Cell 2008, 30, 123–135. [Google Scholar] [CrossRef]
- Grossman, D.; Kim, P.J.; Blanc-Brude, O.P.; Brash, D.E.; Tognin, S.; Marchisio, P.C.; Altieri, D.C. Transgenic expression of survivin in keratinocytes counteracts UVB-induced apoptosis and cooperates with loss of p53. J. Clin. Investig. 2001, 108, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Blanc-Brude, O.P.; Mesri, M.; Wall, N.R.; Plescia, J.; Dohi, T.; Altieri, D.C. Therapeutic targeting of the survivin pathway in cancer: Initiation of mitochondrial apoptosis and suppression of tumor-associated angiogenesis. Clin. Cancer Res. 2003, 9, 2683–2692. [Google Scholar] [PubMed]
- Cetraro, P.; Plaza-Diaz, J.; MacKenzie, A.; Abadia-Molina, F. A Review of the Current Impact of Inhibitors of Apoptosis Proteins and Their Repression in Cancer. Cancers 2022, 14, 1671. [Google Scholar] [CrossRef] [PubMed]
- Deveraux, Q.L.; Reed, J.C. IAP family proteins—Suppressors of apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.J.; Renatus, M.; Schwarzenbacher, R.; Zhou, Q.; Sun, C.; Fesik, S.W.; Liddington, R.C.; Salvesen, G.S. Structural basis for the inhibition of caspase-3 by XIAP. Cell 2001, 104, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Park, Y.C.; Rich, R.L.; Segal, D.; Myszka, D.G.; Wu, H. Structural basis of caspase inhibition by XIAP: Differential roles of the linker versus the BIR domain. Cell 2001, 104, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Shiozaki, E.; Srinivasula, S.M.; Wu, Q.; Datta, P.; Alnemri, E.S.; Shi, Y. Structural basis of caspase-7 inhibition by XIAP. Cell 2001, 104, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Deveraux, Q.; Tamm, I.; Welsh, K.; Assa-Munt, N.; Salvesen, G.S.; Reed, J.C. A single BIR domain of XIAP sufficient for inhibiting caspases. J. Biol. Chem. 1998, 273, 7787–7790. [Google Scholar] [CrossRef]
- Suzuki, Y.; Nakabayashi, Y.; Takahashi, R. Ubiquitin-protein ligase activity of X-linked inhibitor of apoptosis protein promotes proteasomal degradation of caspase-3 and enhances its anti-apoptotic effect in Fas-induced cell death. Proc. Natl. Acad. Sci. USA 2001, 98, 8662–8667. [Google Scholar] [CrossRef]
- Song, Z.; Yao, X.; Wu, M. Direct interaction between survivin and Smac/DIABLO is essential for the anti-apoptotic activity of survivin during taxol-induced apoptosis. J. Biol. Chem. 2003, 278, 23130–23140. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Kelly, A.E.; Funabiki, H.; Patel, D.J. Structural basis for recognition of H3T3ph and Smac/DIABLO N-terminal peptides by human Survivin. Structure 2012, 20, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Fang, M.; Li, Y.; Li, L.; Wang, X. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell 2000, 102, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, A.M.; Ekert, P.G.; Pakusch, M.; Silke, J.; Connolly, L.M.; Reid, G.E.; Moritz, R.L.; Simpson, R.J.; Vaux, D.L. Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to and antagonizing IAP proteins. Cell 2000, 102, 43–53. [Google Scholar] [CrossRef] [PubMed]
- McNeish, I.A.; Lopes, R.; Bell, S.J.; McKay, T.R.; Fernandez, M.; Lockley, M.; Wheatley, S.P.; Lemoine, N.R. Survivin interacts with Smac/DIABLO in ovarian carcinoma cells but is redundant in Smac-mediated apoptosis. Exp. Cell Res. 2005, 302, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Dunajova, L.; Cash, E.; Markus, R.; Rochette, S.; Townley, A.R.; Wheatley, S.P. The N-terminus of survivin is a mitochondrial-targeting sequence and Src regulator. J. Cell Sci. 2016, 129, 2707–2712. [Google Scholar] [CrossRef] [PubMed]
- Dohi, T.; Beltrami, E.; Wall, N.R.; Plescia, J.; Altieri, D.C. Mitochondrial survivin inhibits apoptosis and promotes tumorigenesis. J. Clin. Investig. 2004, 114, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Townley, A.R.; Wheatley, S.P. Mitochondrial survivin reduces oxidative phosphorylation in cancer cells by inhibiting mitophagy. J. Cell Sci. 2020, 133, jcs247379. [Google Scholar] [CrossRef] [PubMed]
- Dohi, T.; Xia, F.; Altieri, D.C. Compartmentalized phosphorylation of IAP by protein kinase A regulates cytoprotection. Mol. Cell 2007, 27, 17–28. [Google Scholar] [CrossRef]
- Rivadeneira, D.B.; Caino, M.C.; Seo, J.H.; Angelin, A.; Wallace, D.C.; Languino, L.R.; Altieri, D.C. Survivin promotes oxidative phosphorylation, subcellular mitochondrial repositioning, and tumor cell invasion. Sci. Signal 2015, 8, ra80. [Google Scholar] [CrossRef]
- Hagenbuchner, J.; Kuznetsov, A.V.; Obexer, P.; Ausserlechner, M.J. BIRC5/Survivin enhances aerobic glycolysis and drug resistance by altered regulation of the mitochondrial fusion/fission machinery. Oncogene 2013, 32, 4748–4757. [Google Scholar] [CrossRef]
- Debnath, J.; Gammoh, N.; Ryan, K.M. Autophagy and autophagy-related pathways in cancer. Nat. Rev. Mol. Cell Biol. 2023, 24, 560–575. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.; Fairlie, W.D.; Lee, E.F. BECLIN1: Protein Structure, Function and Regulation. Cells 2021, 10, 1522. [Google Scholar] [CrossRef] [PubMed]
- Niu, T.K.; Cheng, Y.; Ren, X.; Yang, J.M. Interaction of Beclin 1 with survivin regulates sensitivity of human glioma cells to TRAIL-induced apoptosis. FEBS Lett. 2010, 584, 3519–3524. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Zhang, S.; Mizushima, N. Autophagy genes in biology and disease. Nat. Rev. Genet. 2023, 24, 382–400. [Google Scholar] [CrossRef] [PubMed]
- Maskey, D.; Yousefi, S.; Schmid, I.; Zlobec, I.; Perren, A.; Friis, R.; Simon, H.U. ATG5 is induced by DNA-damaging agents and promotes mitotic catastrophe independent of autophagy. Nat. Commun. 2013, 4, 2130. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Chan, H.H.; Chen, S.H.; Sarvagalla, S.; Chen, P.S.; Coumar, M.S.; Cheng, S.M.; Chang, Y.C.; Lin, C.H.; Leung, E.; et al. BIRC5/Survivin is a novel ATG12-ATG5 conjugate interactor and an autophagy-induced DNA damage suppressor in human cancer and mouse embryonic fibroblast cells. Autophagy 2020, 16, 1296–1313. [Google Scholar] [CrossRef] [PubMed]
- Hagenbuchner, J.; Kiechl-Kohlendorfer, U.; Obexer, P.; Ausserlechner, M.J. BIRC5/Survivin as a target for glycolysis inhibition in high-stage neuroblastoma. Oncogene 2016, 35, 2052–2061. [Google Scholar] [CrossRef] [PubMed]
- Colnaghi, R.; Connell, C.M.; Barrett, R.M.; Wheatley, S.P. Separating the anti-apoptotic and mitotic roles of survivin. J. Biol. Chem. 2006, 281, 33450–33456. [Google Scholar] [CrossRef]
- Knauer, S.K.; Bier, C.; Habtemichael, N.; Stauber, R.H. The Survivin-Crm1 interaction is essential for chromosomal passenger complex localization and function. EMBO Rep. 2006, 7, 1259–1265. [Google Scholar] [CrossRef]
- Stauber, R.H.; Rabenhorst, U.; Rekik, A.; Engels, K.; Bier, C.; Knauer, S.K. Nucleocytoplasmic shuttling and the biological activity of mouse survivin are regulated by an active nuclear export signal. Traffic 2006, 7, 1461–1472. [Google Scholar] [CrossRef] [PubMed]
- Knauer, S.K.; Bier, C.; Schlag, P.; Fritzmann, J.; Dietmaier, W.; Rodel, F.; Klein-Hitpass, L.; Kovacs, A.F.; Doring, C.; Hansmann, M.L.; et al. The survivin isoform survivin-3B is cytoprotective and can function as a chromosomal passenger complex protein. Cell Cycle 2007, 6, 1502–1509. [Google Scholar] [CrossRef] [PubMed]
- Temme, A.; Rodriguez, J.A.; Hendruschk, S.; Gunes, S.; Weigle, B.; Schakel, K.; Schmitz, M.; Bachmann, M.; Schackert, G.; Rieber, E.P. Nuclear localization of Survivin renders HeLa tumor cells more sensitive to apoptosis by induction of p53 and Bax. Cancer Lett. 2007, 250, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Connell, C.M.; Colnaghi, R.; Wheatley, S.P. Nuclear survivin has reduced stability and is not cytoprotective. J. Biol. Chem. 2008, 283, 3289–3296. [Google Scholar] [CrossRef] [PubMed]
- Okada, E.; Murai, Y.; Matsui, K.; Isizawa, S.; Cheng, C.; Masuda, M.; Takano, Y. Survivin expression in tumor cell nuclei is predictive of a favorable prognosis in gastric cancer patients. Cancer Lett. 2001, 163, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.M.; O’Driscoll, L.; Purcell, R.; Fitz-Simons, N.; McDermott, E.W.; Hill, A.D.; O’Higgins, N.J.; Parkinson, M.; Linehan, R.; Clynes, M. Prognostic importance of survivin in breast cancer. Br. J. Cancer 2003, 88, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Vischioni, B.; van der Valk, P.; Span, S.W.; Kruyt, F.A.; Rodriguez, J.A.; Giaccone, G. Nuclear localization of survivin is a positive prognostic factor for survival in advanced non-small-cell lung cancer. Ann. Oncol. 2004, 15, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, E.T.; Gonzalez, A.; Massion, P.P.; Chen, H.; Li, M.; Freyer, A.S.; Olson, S.J.; Andersen, J.J.; Shyr, Y.; Carbone, D.P.; et al. Nuclear survivin predicts recurrence and poor survival in patients with resected nonsmall cell lung carcinoma. Cancer 2005, 103, 1685–1692. [Google Scholar] [CrossRef]
- Shirai, K.; Suzuki, Y.; Oka, K.; Noda, S.E.; Katoh, H.; Suzuki, Y.; Itoh, J.; Itoh, H.; Ishiuchi, S.; Sakurai, H.; et al. Nuclear survivin expression predicts poorer prognosis in glioblastoma. J. Neurooncol. 2008, 91, 353–358. [Google Scholar] [CrossRef]
- Preuss, S.F.; Weinell, A.; Molitor, M.; Stenner, M.; Semrau, R.; Drebber, U.; Weissenborn, S.J.; Speel, E.J.; Wittekindt, C.; Guntinas-Lichius, O.; et al. Nuclear survivin expression is associated with HPV-independent carcinogenesis and is an indicator of poor prognosis in oropharyngeal cancer. Br. J. Cancer 2008, 98, 627–632. [Google Scholar] [CrossRef]
- Kim, J.; McNiff, J.M. Nuclear expression of survivin portends a poor prognosis in Merkel cell carcinoma. Mod. Pathol. 2008, 21, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, P.; Griss, S.; Arnold, C.N.; Horsch, D.; Goke, R.; Arnold, R.; Heine, B.; Stein, H.; Zeitz, M.; Scherubl, H. Nuclear survivin is a powerful novel prognostic marker in gastroenteropancreatic neuroendocrine tumor disease. Neuroendocrinology 2005, 81, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.; Yasufuku, K.; Nakajima, T.; Hiroshima, K.; Chiyo, M.; Yoshida, S.; Suzuki, M.; Sekine, Y.; Shibuya, K.; Agamy, G.; et al. Nuclear Survivin in pN2 Non-small Cell Lung Cancer: Prognostic and Clinical Implications. Eur. Respir. J. 2008, 33, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Chantalat, L.; Skoufias, D.A.; Kleman, J.P.; Jung, B.; Dideberg, O.; Margolis, R.L. Crystal structure of human survivin reveals a bow tie-shaped dimer with two unusual alpha-helical extensions. Mol. Cell 2000, 6, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Verdecia, M.A.; Huang, H.; Dutil, E.; Kaiser, D.A.; Hunter, T.; Noel, J.P. Structure of the human anti-apoptotic protein survivin reveals a dimeric arrangement. Nat. Struct. Biol. 2000, 7, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Muchmore, S.W.; Chen, J.; Jakob, C.; Zakula, D.; Matayoshi, E.D.; Wu, W.; Zhang, H.; Li, F.; Ng, S.C.; Altieri, D.C. Crystal structure and mutagenic analysis of the inhibitor-of-apoptosis protein survivin. Mol. Cell 2000, 6, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Nettesheim, D.; Liu, Z.; Olejniczak, E.T. Solution structure of human survivin and its binding interface with Smac/Diablo. Biochemistry 2005, 44, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Vong, Q.P.; Cao, K.; Li, H.Y.; Iglesias, P.A.; Zheng, Y. Chromosome alignment and segregation regulated by ubiquitination of survivin. Science 2005, 310, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Jeyaprakash, A.A.; Klein, U.R.; Lindner, D.; Ebert, J.; Nigg, E.A.; Conti, E. Structure of a Survivin-Borealin-INCENP core complex reveals how chromosomal passengers travel together. Cell 2007, 131, 271–285. [Google Scholar] [CrossRef]
- Niedzialkowska, E.; Wang, F.; Porebski, P.J.; Minor, W.; Higgins, J.M.; Stukenberg, P.T. Molecular basis for phosphospecific recognition of histone H3 tails by Survivin paralogues at inner centromeres. Mol. Biol. Cell 2012, 23, 1457–1466. [Google Scholar] [CrossRef]
- Barrett, R.M.; Osborne, T.P.; Wheatley, S.P. Phosphorylation of survivin at threonine 34 inhibits its mitotic function and enhances its cytoprotective activity. Cell Cycle 2009, 8, 278–283. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.S.; Grossman, D.; Plescia, J.; Li, F.; Zhang, H.; Villa, A.; Tognin, S.; Marchisio, P.C.; Altieri, D.C. Regulation of apoptosis at cell division by p34cdc2 phosphorylation of survivin. Proc. Natl. Acad. Sci. USA 2000, 97, 13103–13107. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, S.P.; Barrett, R.M.; Andrews, P.D.; Medema, R.H.; Morley, S.J.; Swedlow, J.R.; Lens, S.M. Phosphorylation by aurora-B negatively regulates survivin function during mitosis. Cell Cycle 2007, 6, 1220–1230. [Google Scholar] [CrossRef] [PubMed]
- Barrett, R.M.; Colnaghi, R.; Wheatley, S.P. Threonine 48 in the BIR domain of survivin is critical to its mitotic and anti-apoptotic activities and can be phosphorylated by CK2 in vitro. Cell Cycle 2011, 10, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Colnaghi, R.; Wheatley, S.P. Liaisons between survivin and Plk1 during cell division and cell death. J. Biol. Chem. 2010, 285, 22592–22604. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Holloway, M.P.; Ma, L.; Cooper, Z.A.; Riolo, M.; Samkari, A.; Elenitoba-Johnson, K.S.; Chin, Y.E.; Altura, R.A. Acetylation directs survivin nuclear localization to repress STAT3 oncogenic activity. J. Biol. Chem. 2010, 285, 36129–36137. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, T.; Kita, A.; Yamanaka, K.; Mori, M.; Amino, N.; Takeuchi, M.; Tominaga, F.; Hatakeyama, S.; Kinoyama, I.; Matsuhisa, A.; et al. YM155, a novel small-molecule survivin suppressant, induces regression of. Cancer Res. 2007, 67, 8014–8021. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, T.; Okamoto, I.; Suzuki, M.; Nakahara, T.; Yamanaka, K.; Hatashita, E.; Yamada, Y.; Fukuoka, M.; Ono, K.; Nakagawa, K. Radiosensitizing effect of YM155, a novel small-molecule survivin suppressant, in. Clin. Cancer Res. 2008, 14, 6496–6504. [Google Scholar] [CrossRef]
- Giaccone, G.; Zatloukal, P.; Roubec, J.; Floor, K.; Musil, J.; Kuta, M.; van Klaveren, R.J.; Chaudhary, S.; Gunther, A.; Shamsili, S. Multicenter phase II trial of YM155, a small-molecule suppressor of survivin, in patients with advanced, refractory, non-small-cell lung cancer. J. Clin. Oncol. 2009, 27, 4481–4486. [Google Scholar] [CrossRef]
- Kelly, R.J.; Thomas, A.; Rajan, A.; Chun, G.; Lopez-Chavez, A.; Szabo, E.; Spencer, S.; Carter, C.A.; Guha, U.; Khozin, S.; et al. A phase I/II study of sepantronium bromide (YM155, survivin suppressor) with paclitaxel and carboplatin in patients with advanced non-small-cell lung cancer. Ann. Oncol. 2013, 24, 2601–2606. [Google Scholar] [CrossRef]
- Satoh, T.; Okamoto, I.; Miyazaki, M.; Morinaga, R.; Tsuya, A.; Hasegawa, Y.; Terashima, M.; Ueda, S.; Fukuoka, M.; Ariyoshi, Y.; et al. Phase I study of YM155, a novel survivin suppressant, in patients with advanced solid tumors. Clin. Cancer Res. 2009, 15, 3872–3880. [Google Scholar] [CrossRef] [PubMed]
- Tolcher, A.W.; Quinn, D.I.; Ferrari, A.; Ahmann, F.; Giaccone, G.; Drake, T.; Keating, A.; de Bono, J.S. A phase II study of YM155, a novel small-molecule suppressor of survivin, in castration-resistant taxane-pretreated prostate cancer. Ann. Oncol. 2012, 23, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Tolcher, A.W.; Mita, A.; Lewis, L.D.; Garrett, C.R.; Till, E.; Daud, A.I.; Patnaik, A.; Papadopoulos, K.; Takimoto, C.; Bartels, P.; et al. Phase I and pharmacokinetic study of YM155, a small-molecule inhibitor of survivin. J. Clin. Oncol. 2008, 26, 5198–5203. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Zhang, L.; Boufraqech, M.; Liu-Chittenden, Y.; Zhang, Y.; Patel, D.; Davis, S.; Rosenberg, A.; Ylaya, K.; Aufforth, R.; et al. Inhibition of Survivin with YM155 Induces Durable Tumor Response in Anaplastic Thyroid Cancer. Clin. Cancer Res. 2015, 21, 4123–4132. [Google Scholar] [CrossRef]
- Arora, R.; Shuda, M.; Guastafierro, A.; Feng, H.; Toptan, T.; Tolstov, Y.; Normolle, D.; Vollmer, L.L.; Vogt, A.; Domling, A.; et al. Survivin is a therapeutic target in merkel cell carcinoma. Sci. Transl. Med. 2012, 4, 133ra156. [Google Scholar] [CrossRef]
- Glaros, T.G.; Stockwin, L.H.; Mullendore, M.E.; Smith, B.; Morrison, B.L.; Newton, D.L. The “survivin suppressants” NSC 80467 and YM155 induce a DNA damage response. Cancer Chemother. Pharmacol. 2012, 70, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Majera, D.; Mistrik, M. Effect of Sepatronium Bromide (YM-155) on DNA Double-Strand Breaks Repair in Cancer Cells. Int. J. Mol. Sci. 2020, 21, 9431. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.; Cao, S.; Cheng, Q.; Keefe, J.T.; Rustum, Y.M.; Li, F. A novel small molecule FL118 that selectively inhibits survivin, Mcl-1, XIAP and cIAP2 in a p53-independent manner, shows superior antitumor activity. PLoS ONE 2012, 7, e45571. [Google Scholar] [CrossRef]
- Yin, H.; Que, R.; Liu, C.; Ji, W.; Sun, B.; Lin, X.; Zhang, Q.; Zhao, X.; Peng, Z.; Zhang, X.; et al. Survivin-targeted drug screening platform identifies a matrine derivative WM-127 as a potential therapeutics against hepatocellular carcinoma. Cancer Lett. 2018, 425, 54–64. [Google Scholar] [CrossRef]
- Chang, C.C.; Heller, J.D.; Kuo, J.; Huang, R.C. Tetra-O-methyl nordihydroguaiaretic acid induces growth arrest and cellular apoptosis by inhibiting Cdc2 and survivin expression. Proc. Natl. Acad. Sci. USA 2004, 101, 13239–13244. [Google Scholar] [CrossRef]
- Park, R.; Chang, C.C.; Liang, Y.C.; Chung, Y.; Henry, R.A.; Lin, E.; Mold, D.E.; Huang, R.C. Systemic treatment with tetra-O-methyl nordihydroguaiaretic acid suppresses the growth of human xenograft tumors. Clin. Cancer Res. 2005, 11, 4601–4609. [Google Scholar] [CrossRef] [PubMed]
- Heller, J.D.; Kuo, J.; Wu, T.C.; Kast, W.M.; Huang, R.C. Tetra-O-methyl nordihydroguaiaretic acid induces G2 arrest in mammalian cells and exhibits tumoricidal activity in vivo. Cancer Res. 2001, 61, 5499–5504. [Google Scholar] [PubMed]
- Shi, X.; Wang, D.; Ding, K.; Lu, Z.; Jin, Y.; Zhang, J.; Pan, J. GDP366, a novel small molecule dual inhibitor of survivin and Op18, induces cell growth inhibition, cellular senescence and mitotic catastrophe in human cancer cells. Cancer Biol. Ther. 2010, 9, 640–650. [Google Scholar] [CrossRef]
- Belmont, L.D.; Mitchison, T.J. Identification of a protein that interacts with tubulin dimers and increases the catastrophe rate of microtubules. Cell 1996, 84, 623–631. [Google Scholar] [CrossRef]
- Marklund, U.; Larsson, N.; Gradin, H.M.; Brattsand, G.; Gullberg, M. Oncoprotein 18 is a phosphorylation-responsive regulator of microtubule dynamics. EMBO J. 1996, 15, 5290–5298. [Google Scholar] [CrossRef] [PubMed]
- Miceli, C.; Tejada, A.; Castaneda, A.; Mistry, S.J. Cell cycle inhibition therapy that targets stathmin in in vitro and in vivo models of breast cancer. Cancer Gene Ther. 2013, 20, 298–307. [Google Scholar] [CrossRef]
- Fortugno, P.; Beltrami, E.; Plescia, J.; Fontana, J.; Pradhan, D.; Marchisio, P.C.; Sessa, W.C.; Altieri, D.C. Regulation of survivin function by Hsp90. Proc. Natl. Acad. Sci. USA 2003, 100, 13791–13796. [Google Scholar] [CrossRef]
- Plescia, J.; Salz, W.; Xia, F.; Pennati, M.; Zaffaroni, N.; Daidone, M.G.; Meli, M.; Dohi, T.; Fortugno, P.; Nefedova, Y.; et al. Rational design of shepherdin, a novel anticancer agent. Cancer Cell 2005, 7, 457–468. [Google Scholar] [CrossRef]
- Zhao, G.; Wang, Q.; Wu, Z.; Tian, X.; Yan, H.; Wang, B.; Dong, P.; Watari, H.; Pfeffer, L.M.; Guo, Y.; et al. Ovarian Primary and Metastatic Tumors Suppressed by Survivin Knockout or a Novel Survivin Inhibitor. Mol. Cancer Ther. 2019, 18, 2233–2245. [Google Scholar] [CrossRef]
- Wang, J.; Li, W. Discovery of novel second mitochondria-derived activator of caspase mimetics as selective inhibitor of apoptosis protein inhibitors. J. Pharmacol. Exp. Ther. 2014, 349, 319–329. [Google Scholar] [CrossRef]
- Oikawa, T.; Unno, Y.; Matsuno, K.; Sawada, J.; Ogo, N.; Tanaka, K.; Asai, A. Identification of a small-molecule inhibitor of the interaction between Survivin and Smac/DIABLO. Biochem. Biophys. Res. Commun. 2010, 393, 253–258. [Google Scholar] [CrossRef]
- Park, S.H.; Shin, I.; Park, S.H.; Kim, N.D.; Shin, I. An Inhibitor of the Interaction of Survivin with Smac in Mitochondria Promotes Apoptosis. Chem. Asian J. 2019, 14, 4035–4041. [Google Scholar] [CrossRef] [PubMed]
- Wendt, M.D.; Sun, C.; Kunzer, A.; Sauer, D.; Sarris, K.; Hoff, E.; Yu, L.; Nettesheim, D.G.; Chen, J.; Jin, S.; et al. Discovery of a novel small molecule binding site of human survivin. Bioorg Med. Chem. Lett. 2007, 17, 3122–3129. [Google Scholar] [CrossRef] [PubMed]
- Guvenc, H.; Pavlyukov, M.S.; Joshi, K.; Kurt, H.; Banasavadi-Siddegowda, Y.K.; Mao, P.; Hong, C.; Yamada, R.; Kwon, C.H.; Bhasin, D.; et al. Impairment of glioma stem cell survival and growth by a novel inhibitor for Survivin-Ran protein complex. Clin. Cancer Res. 2013, 19, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Chettiar, S.N.; Cooley, J.V.; Park, I.H.; Bhasin, D.; Chakravarti, A.; Li, P.K.; Li, C.; Jacob, N.K. Design, synthesis and biological studies of survivin dimerization modulators that prolong mitotic cycle. Bioorg. Med. Chem. Lett. 2013, 23, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Steigerwald, C.; Rasenberger, B.; Christmann, M.; Tomicic, M.T. Sensitization of colorectal cancer cells to irinotecan by the Survivin inhibitor LLP3 depends on XAF1 proficiency in the context of mutated p53. Arch. Toxicol. 2018, 92, 2645–2648. [Google Scholar] [CrossRef] [PubMed]
- Berezov, A.; Cai, Z.; Freudenberg, J.A.; Zhang, H.; Cheng, X.; Thompson, T.; Murali, R.; Greene, M.I.; Wang, Q. Disabling the mitotic spindle and tumor growth by targeting a cavity-induced allosteric site of survivin. Oncogene 2012, 31, 1938–1948. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.M.; Nagatomo, I.; Suzuki, E.; Mizuno, T.; Kumagai, T.; Berezov, A.; Zhang, H.; Karlan, B.; Greene, M.I.; Wang, Q. YAP modifies cancer cell sensitivity to EGFR and survivin inhibitors and is negatively regulated by the non-receptor type protein tyrosine phosphatase 14. Oncogene 2013, 32, 2220–2229. [Google Scholar] [CrossRef] [PubMed]
- Murali, R.; Cheng, X.; Berezov, A.; Du, X.; Schon, A.; Freire, E.; Xu, X.; Chen, Y.H.; Greene, M.I. Disabling TNF receptor signaling by induced conformational perturbation of tryptophan-107. Proc. Natl. Acad. Sci. USA 2005, 102, 10970–10975. [Google Scholar] [CrossRef]
- Ishikawa, N.; Fuchigami, T.; Mizoguchi, T.; Yoshida, S.; Haratake, M.; Nakayama, M. Synthesis and characterization of radioiodinated 3-phenethyl-2-indolinone derivatives for SPECT imaging of survivin in tumors. Bioorg. Med. Chem. 2018, 26, 3111–3116. [Google Scholar] [CrossRef]
- Qi, J.; Dong, Z.; Liu, J.; Peery, R.C.; Zhang, S.; Liu, J.Y.; Zhang, J.T. Effective Targeting of the Survivin Dimerization Interface with Small-Molecule Inhibitors. Cancer Res. 2016, 76, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Peery, R.; Kyei-Baffour, K.; Dong, Z.; Liu, J.; de Andrade Horn, P.; Dai, M.; Liu, J.Y.; Zhang, J.T. Synthesis and Identification of a Novel Lead Targeting Survivin Dimerization for Proteasome-Dependent Degradation. J. Med. Chem. 2020, 63, 7243–7251. [Google Scholar] [CrossRef]
- Andersen, M.H.; Pedersen, L.O.; Becker, J.C.; Straten, P.T. Identification of a cytotoxic T lymphocyte response to the apoptosis inhibitor protein survivin in cancer patients. Cancer Res. 2001, 61, 869–872. [Google Scholar]
- Hirohashi, Y.; Torigoe, T.; Maeda, A.; Nabeta, Y.; Kamiguchi, K.; Sato, T.; Yoda, J.; Ikeda, H.; Hirata, K.; Yamanaka, N.; et al. An HLA-A24-restricted cytotoxic T lymphocyte epitope of a tumor-associated protein, survivin. Clin. Cancer Res. 2002, 8, 1731–1739. [Google Scholar] [PubMed]
- Xiang, R.; Mizutani, N.; Luo, Y.; Chiodoni, C.; Zhou, H.; Mizutani, M.; Ba, Y.; Becker, J.C.; Reisfeld, R.A. A DNA vaccine targeting survivin combines apoptosis with suppression of angiogenesis in lung tumor eradication. Cancer Res. 2005, 65, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Onodi, F.; Maherzi-Mechalikh, C.; Mougel, A.; Ben Hamouda, N.; Taboas, C.; Gueugnon, F.; Tran, T.; Nozach, H.; Marcon, E.; Gey, A.; et al. High Therapeutic Efficacy of a New Survivin LSP-Cancer Vaccine Containing CD4(+) and CD8(+) T-Cell Epitopes. Front. Oncol. 2018, 8, 517. [Google Scholar] [CrossRef]
- Fenstermaker, R.A.; Ciesielski, M.J.; Qiu, J.; Yang, N.; Frank, C.L.; Lee, K.P.; Mechtler, L.R.; Belal, A.; Ahluwalia, M.S.; Hutson, A.D. Clinical study of a survivin long peptide vaccine (SurVaxM) in patients with recurrent malignant glioma. Cancer Immunol. Immunother. 2016, 65, 1339–1352. [Google Scholar] [CrossRef]
- Ahluwalia, M.S.; Reardon, D.A.; Abad, A.P.; Curry, W.T.; Wong, E.T.; Figel, S.A.; Mechtler, L.L.; Peereboom, D.M.; Hutson, A.D.; Withers, H.G.; et al. Phase IIa Study of SurVaxM Plus Adjuvant Temozolomide for Newly Diagnosed Glioblastoma. J. Clin. Oncol. 2023, 41, 1453–1465. [Google Scholar] [CrossRef]
- Ciesielski, M.J.; Ahluwalia, M.S.; Munich, S.A.; Orton, M.; Barone, T.; Chanan-Khan, A.; Fenstermaker, R.A. Antitumor cytotoxic T-cell response induced by a survivin peptide mimic. Cancer Immunol. Immunother. 2010, 59, 1211–1221. [Google Scholar] [CrossRef]
- Locke, F.L.; Menges, M.; Veerapathran, A.; Coppola, D.; Gabrilovich, D.; Anasetti, C. Survivin-specific CD4+ T cells are decreased in patients with survivin-positive myeloma. J. Immunother. Cancer 2015, 3, 20. [Google Scholar] [CrossRef]
- Freeman, C.L.; Atkins, R.; Varadarajan, I.; Menges, M.; Edelman, J.; Baz, R.; Brayer, J.; Castaneda Puglianini, O.; Ochoa-Bayona, J.L.; Nishihori, T.; et al. Survivin Dendritic Cell Vaccine Safely Induces Immune Responses and Is Associated with Durable Disease Control after Autologous Transplant in Patients with Myeloma. Clin. Cancer Res. 2023, 29, 4575–4585. [Google Scholar] [CrossRef] [PubMed]
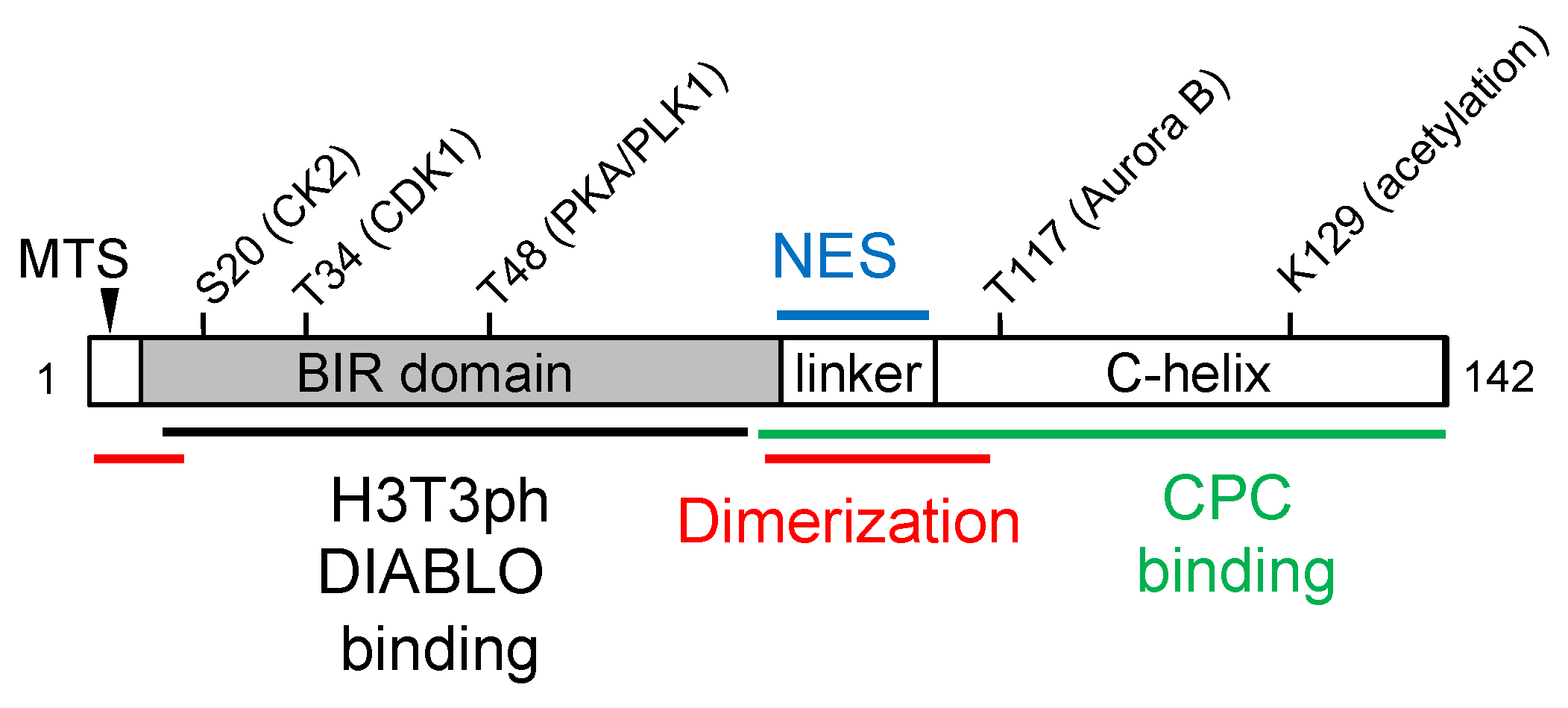
Figure 1.
Schematic representation of survivin structure features involved in dimerization (red), chromosome passenger complex (CPC) binding (green), histone H3 threonine 3 phosphorylated peptide (H3T3ph) and DIABLO binding (black), nuclear export (NES, blue), and mitochondria targeting (MTS, arrowhead). The amino acid residues that have been reported to be modified by acetylation or phosphorylation, as well as the kinases involved, are also indicated.
Figure 1.
Schematic representation of survivin structure features involved in dimerization (red), chromosome passenger complex (CPC) binding (green), histone H3 threonine 3 phosphorylated peptide (H3T3ph) and DIABLO binding (black), nuclear export (NES, blue), and mitochondria targeting (MTS, arrowhead). The amino acid residues that have been reported to be modified by acetylation or phosphorylation, as well as the kinases involved, are also indicated.

Figure 2.
Functions of survivin. (A) Survivin is required for mitosis and cytokinesis. Survivin is associated with INCENP and borealin as components of the chromosome passenger complex (CPC) localized to the centromere during mitosis. Survivin remains associated with the spindle midbody from the anaphase of mitosis until the end of cytokinesis. (B) Survivin can protect cells from apoptosis. The binding of survivin to DIABLO may prevent the latter from inactivating other inhibitor of apoptosis (IAP) family proteins. Alternatively, survivin may directly inhibit caspase activity. (C) Survivin can be localized to the mitochondria and protect cells from mitochondria-mediated apoptosis. (D) Survivin can be associated with beclin or ATG5 and is postulated to be involved in aspects of autophagy.
Figure 2.
Functions of survivin. (A) Survivin is required for mitosis and cytokinesis. Survivin is associated with INCENP and borealin as components of the chromosome passenger complex (CPC) localized to the centromere during mitosis. Survivin remains associated with the spindle midbody from the anaphase of mitosis until the end of cytokinesis. (B) Survivin can protect cells from apoptosis. The binding of survivin to DIABLO may prevent the latter from inactivating other inhibitor of apoptosis (IAP) family proteins. Alternatively, survivin may directly inhibit caspase activity. (C) Survivin can be localized to the mitochondria and protect cells from mitochondria-mediated apoptosis. (D) Survivin can be associated with beclin or ATG5 and is postulated to be involved in aspects of autophagy.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Q.; Greene, M.I. Survivin as a Therapeutic Target for the Treatment of Human Cancer. Cancers 2024, 16, 1705. https://doi.org/10.3390/cancers16091705
AMA Style
Wang Q, Greene MI. Survivin as a Therapeutic Target for the Treatment of Human Cancer. Cancers. 2024; 16(9):1705. https://doi.org/10.3390/cancers16091705
Chicago/Turabian StyleWang, Qiang, and Mark I. Greene. 2024. "Survivin as a Therapeutic Target for the Treatment of Human Cancer" Cancers 16, no. 9: 1705. https://doi.org/10.3390/cancers16091705
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.