Roseburia intestinalis Supplementation Could Reverse the Learning and Memory Impairment and m6A Methylation Modification Decrease Caused by 27-Hydroxycholesterol in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Roseburia Intestinalis Culture
2.2. Animal and Experimental Design
2.3. Neurobehavioral Tests
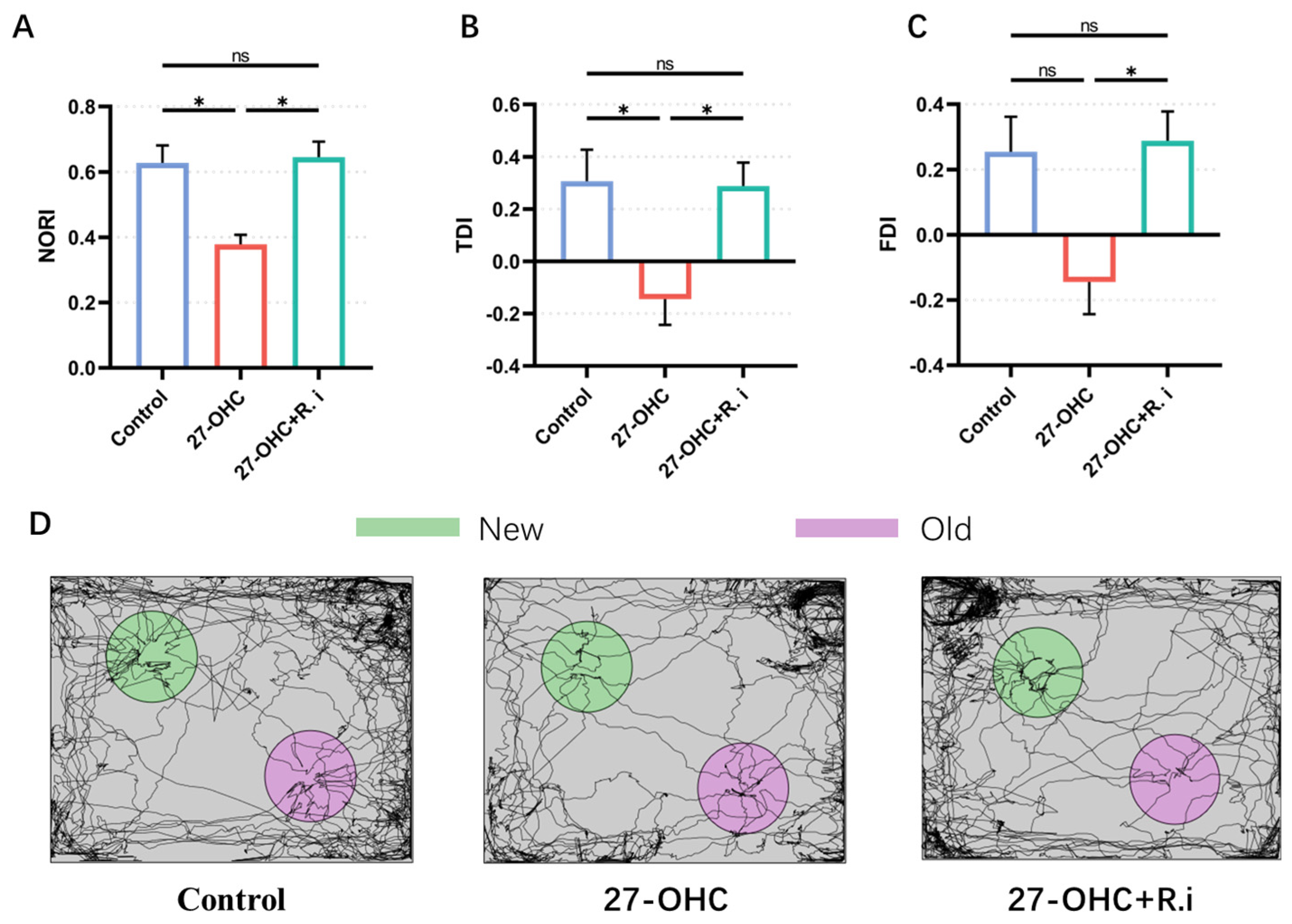
2.3.1. Novel Object Recognition Test
2.3.2. Y Maze Test
2.3.3. Morris Water Maze (MWM) Test
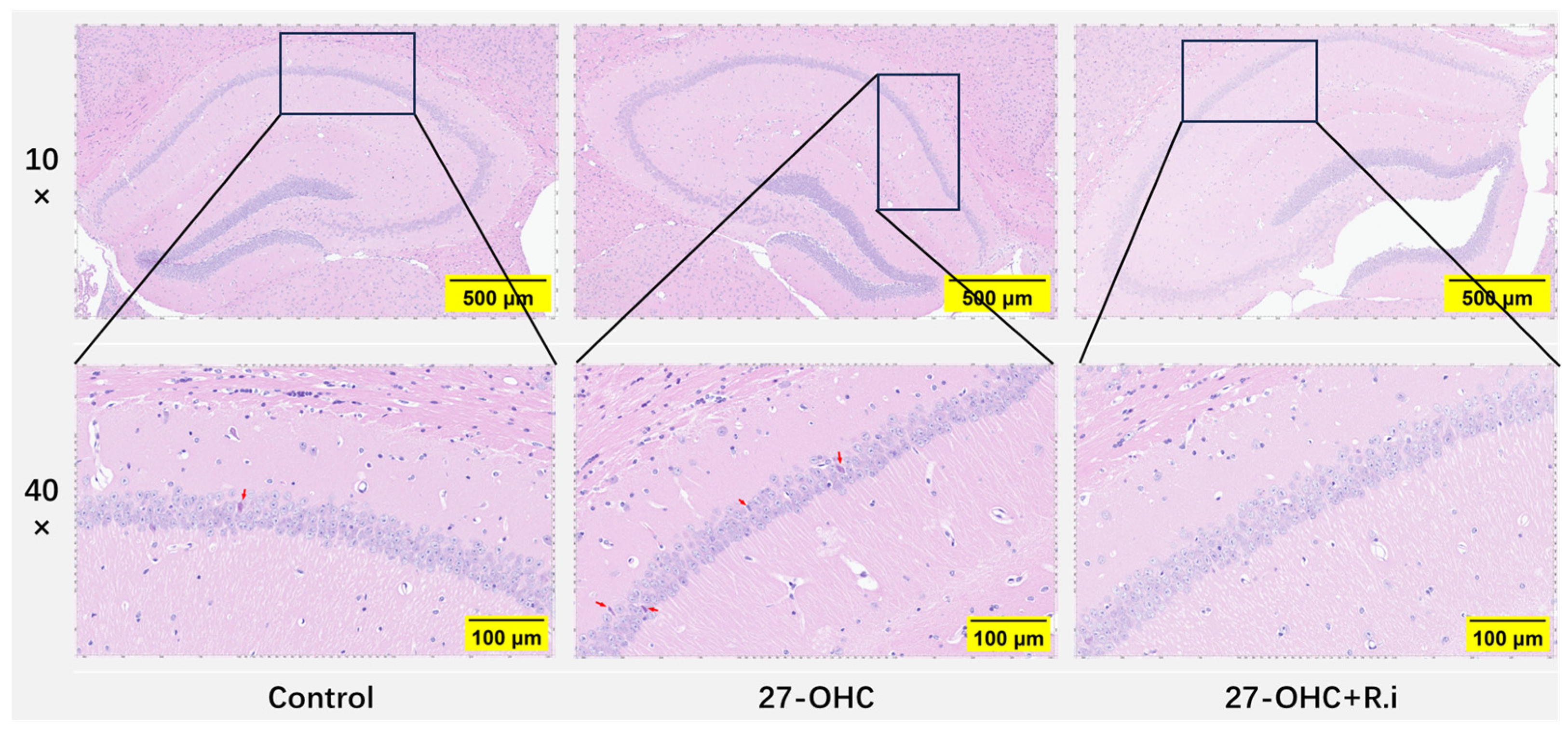
2.4. Hematoxylin-Eosin (HE) Staining
2.5. High-Performance Liquid Chromatography-Mass Spectrometry (HPLC–MS)
2.6. Total RNA m6A Modification Level
2.7. Quantitative Real-Time PCR (RT-PCR)
2.8. Western Blot
2.9. Statistical Analysis
3. Results
3.1. Body Weight and Organ Coefficient Were Not Affected by the 27-OHC Intervention and Roseburia Intestinalis Gavage
3.2. Roseburia Intestinalis Intervention Reversed the Impaired Learning and Memory Ability Caused by 27-OHC Treatment
3.3. Roseburia Intestinalis Intervention Improved the 27-OHC-Induced Morphology Change in the Brain
3.4. Roseburia Intestinalis Intervention Could Restore the Decrease of Synapse-Related Molecules Caused by 27-OHC in Brain Cortex
3.5. Roseburia Intestinalis Complementation Reversed the Abnormal Change in m6A Methylation Modification Level and Its Related Enzymes Caused by 27-OHC
3.6. Roseburia Intestinalis Complementation Improved the Level of 24S-OHC in the Brain
3.7. Roseburia Intestinalis Intervention Saved the Impaired Colon Barrier Function Caused by 27-OHC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alzheimer’s Association. 2023 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2023, 19, 1598–1695. [Google Scholar] [CrossRef] [PubMed]
- Self, W.K.; Holtzman, D.M. Emerging diagnostics and therapeutics for Alzheimer disease. Nat. Med. 2023, 29, 2187–2199. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.; Kales, H.C. New treatments for Alzheimer’s disease. BMJ 2023, 382, 1852. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, J.; Liu, Q. Brain cell type-specific cholesterol metabolism and implications for learning and memory. Trends Neurosci. 2022, 45, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Yeo, I.-J.; Hong, J.-T.; Eo, S.-K.; Lee, D.; Kim, K. Side-Chain Immune Oxysterols Induce Neuroinflammation by Activating Microglia. Int. J. Mol. Sci. 2023, 24, 15288. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.L.; Ferreira, C.N.; Palotás, A.; Rocha, N.P.; Reis, H.J. Role of Oxysterols in the Activation of the NLRP3 Inflammasome as a Potential Pharmacological Approach in Alzheimer’s Disease. Curr. Neuropharmacol. 2023, 21, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Sorboni, S.G.; Moghaddam, H.S.; Jafarzadeh-Esfehani, R.; Soleimanpour, S. A Comprehensive Review on the Role of the Gut Microbiome in Human Neurological Disorders. Clin. Microbiol. Rev. 2022, 35, e0033820. [Google Scholar] [CrossRef]
- Bairamian, D.; Sha, S.; Rolhion, N.; Sokol, H.; Dorothée, G.; Lemere, C.A.; Krantic, S. Microbiota in neuroinflammation and synaptic dysfunction: A focus on Alzheimer’s disease. Mol. Neurodegener. 2022, 17, 19. [Google Scholar] [CrossRef]
- Zhu, G.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Gut Microbiota and its Metabolites: Bridge of Dietary Nutrients and Alzheimer’s Disease. Adv. Nutr. 2023, 14, 819–839. [Google Scholar] [CrossRef]
- Migliore, L.; Coppedè, F. Gene-environment interactions in Alzheimer disease: The emerging role of epigenetics. Nat. Rev. Neurol. 2022, 18, 643–660. [Google Scholar] [CrossRef]
- Maity, S.; Farrell, K.; Navabpour, S.; Narayanan, S.N.; Jarome, T.J. Epigenetic Mechanisms in Memory and Cognitive Decline Associated with Aging and Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 12280. [Google Scholar] [CrossRef] [PubMed]
- Jeremic, D.; Jiménez-Díaz, L.; Navarro-López, J.D. Targeting epigenetics: A novel promise for Alzheimer’s disease treatment. Ageing Res. Rev. 2023, 90, 102003. [Google Scholar] [CrossRef] [PubMed]
- Merino-Serrais, P.; Loera-Valencia, R.; Rodriguez-Rodriguez, P.; Parrado-Fernandez, C.; Ismail, M.A.; Maioli, S.; Matute, E.; Jimenez-Mateos, E.M.; Björkhem, I.; DeFelipe, J.; et al. 27-Hydroxycholesterol Induces Aberrant Morphology and Synaptic Dysfunction in Hippocampal Neurons. Cereb. Cortex 2019, 29, 429–446. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, H.; Hao, L.; Ju, M.; Feng, W.; Xiao, R. The Interaction Effect of 27-Hydroxycholesterol Metabolism Disorder and CYP27A1 Single Nucleotide Polymorphisms in Mild Cognitive Impairment: Evidence from a Case-Control Study. Mol. Nutr. Food Res. 2023, 67, e2200576. [Google Scholar] [CrossRef]
- Liu, Q.; An, Y.; Yu, H.; Lu, Y.; Feng, L.; Wang, C.; Xiao, R. Relationship between oxysterols and mild cognitive impairment in the elderly: A case-control study. Lipids Health Dis. 2016, 15, 177. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cui, S.; Hao, L.; Liu, W.; Wang, L.; Ju, M.; Feng, W.; Xiao, R. Regulation of Th17/Treg Balance by 27-Hydroxycholesterol and 24S-Hydroxycholesterol Correlates with Learning and Memory Ability in Mice. Int. J. Mol. Sci. 2022, 23, 4370. [Google Scholar] [CrossRef]
- Wang, Y.; An, Y.; Ma, W.; Yu, H.; Lu, Y.; Zhang, X.; Wang, Y.; Liu, W.; Wang, T.; Xiao, R. 27-Hydroxycholesterol contributes to cognitive deficits in APP/PS1 transgenic mice through microbiota dysbiosis and intestinal barrier dysfunction. J. NeuroInflamm. 2020, 17, 199. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Sisodia, S.S.; Vassar, R.J. The gut microbiome in Alzheimer’s disease: What we know and what remains to be explored. Mol. Neurodegener. 2023, 18, 9. [Google Scholar] [CrossRef]
- Pellegrini, C.; Fornai, M.; D’Antongiovanni, V.; Antonioli, L.; Bernardini, N.; Derkinderen, P. The intestinal barrier in disorders of the central nervous system. Lancet Gastroenterol. Hepatol. 2023, 8, 66–80. [Google Scholar] [CrossRef]
- Choi, H.; Mook-Jung, I. Functional effects of gut microbiota-derived metabolites in Alzheimer’s disease. Curr. Opin. Neurobiol. 2023, 81, 102730. [Google Scholar] [CrossRef]
- Jin, J.; Xu, Z.; Zhang, L.; Zhang, C.; Zhao, X.; Mao, Y.; Zhang, H.; Liang, X.; Wu, J.; Yang, Y.; et al. Gut-derived β-amyloid: Likely a centerpiece of the gut-brain axis contributing to Alzheimer’s pathogenesis. Gut Microbes 2023, 15, 2167172. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Wang, L.; Ju, M.; Feng, W.; Guo, Z.; Sun, X.; Xiao, R. 27-Hydroxycholesterol impairs learning and memory ability via decreasing brain glucose uptake mediated by the gut microbiota. Biomed. Pharmacother. 2023, 168, 115649. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Zhu, J.; Sun, C.; Li, M.; Liu, J.; Wu, S.; Ning, K.; He, L.-J.; Zhao, X.-M.; Chen, W.-H. GMrepo v2: A curated human gut microbiome database with special focus on disease markers and cross-dataset comparison. Nucleic Acids Res. 2022, 50, D777–D784. [Google Scholar] [CrossRef] [PubMed]
- Verhaar, B.J.H.; Hendriksen, H.M.A.; de Leeuw, F.A.; Doorduijn, A.S.; van Leeuwenstijn, M.; Teunissen, C.E.; Barkhof, F.; Scheltens, P.; Kraaij, R.; van Duijn, C.M.; et al. Gut Microbiota Composition Is Related to AD Pathology. Front. Immunol. 2021, 12, 794519. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Cheng, Y.; Ruan, G.; Fan, L.; Tian, Y.; Xiao, Z.; Chen, D.; Wei, Y. New pathway ameliorating ulcerative colitis: Focus on Roseburia intestinalis and the gut-brain axis. Ther. Adv. Gastroenterol. 2021, 14, 17562848211004469. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Sun, Q.; Zheng, H.; Zhang, Y.; Wang, Y.; Liu, S.; Duan, L. Roseburia hominis Alleviates Neuroinflammation via Short-Chain Fatty Acids through Histone Deacetylase Inhibition. Mol. Nutr. Food Res. 2022, 66, e2200164. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Meng, C.; Li, L.; Feng, S.; Zhu, Y.; Yang, J.; Han, L.; Sun, L.; Lv, W.; Figeys, D.; et al. Positive mood-related gut microbiota in a long-term closed environment: A multiomics study based on the “Lunar Palace 365” experiment. Microbiome 2023, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Ju, Z.; Zheng, M.; Zhang, X.; Zuo, W.; Wang, Y.; Ding, X.; Zhang, X.; Peng, Y.; Li, J.; et al. Loss of the m6A methyltransferase METTL3 in monocyte-derived macrophages ameliorates Alzheimer’s disease pathology in mice. PLoS Biol. 2023, 21, e3002017. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Zhang, F.; Li, Y.; Mo, Y.; Zhang, L.; Li, Q.; Luo, M.; Hou, X.; Du, Z.; Deng, J.; et al. A new perspective on Alzheimer’s disease: m6A modification. Front. Genet. 2023, 14, 1166831. [Google Scholar] [CrossRef]
- Boulias, K.; Greer, E.L. Biological roles of adenine methylation in RNA. Nat. Rev. Genet. 2023, 24, 143–160. [Google Scholar] [CrossRef]
- Castro-Hernández, R.; Berulava, T.; Metelova, M.; Epple, R.; Peña Centeno, T.; Richter, J.; Kaurani, L.; Pradhan, R.; Sakib, M.S.; Burkhardt, S.; et al. Conserved reduction of m6A RNA modifications during aging and neurodegeneration is linked to changes in synaptic transcripts. Proc. Natl. Acad. Sci. USA 2023, 120, e2204933120. [Google Scholar] [CrossRef] [PubMed]
- Kivisäkk, P.; Carlyle, B.C.; Sweeney, T.; Quinn, J.P.; Ramirez, C.E.; Trinbetta, B.A.; Mendes, M.; Brock, M.; Rubel, C.; Czerkowicz, J.; et al. Increased levels of the synaptic proteins PSD-95, SNAP-25, and neurogranin in the cerebrospinal fluid of patients with Alzheimer’s disease. Alzheimer’s Res. Ther. 2022, 14, 58. [Google Scholar] [CrossRef]
- Wang, C.; Kavalali, E.; Monteggia, L. BDNF Signaling in Context: From Synaptic Regulation to Psychiatric Disorders. Cell 2022, 185, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.; Zhu, S.; Peng, J.; Lang, Z.; Zhou, X.; Liao, H.; Zou, J.; Zeng, P.; Tan, S. The hippocampal FTO-BDNF-TrkB pathway is required for novel object recognition memory reconsolidation in mice. Transl. Psychiatry 2023, 13, 349. [Google Scholar] [CrossRef] [PubMed]
- Merkurjev, D.; Hong, W.-T.; Iida, K.; Oomoto, I.; Goldie, B.J.; Yamaguti, H.; Ohara, T.; Kawaguchi, S.-Y.; Hirano, T.; Martin, K.C.; et al. Synaptic N6-methyladenosine (m6A) epitranscriptome reveals functional partitioning of localized transcripts. Nat. Neurosci. 2018, 21, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- .Knight, H.M.; Öz, M.D.; PerezGrovas-Saltijeral, A. Dysregulation of RNA modification systems in clinical populations with neurocognitive disorders. Neural Regen. Res. 2024, 19, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Wellman, A.S.; Metukuri, M.R.; Kazgan, N.; Xu, X.; Xu, Q.; Ren, N.S.X.; Czopik, A.; Shanahan, M.T.; Kang, A.; Chen, W.; et al. Intestinal Epithelial Sirtuin 1 Regulates Intestinal Inflammation during Aging in Mice by Altering the Intestinal Microbiota. Gastroenterology 2017, 153, 772–786. [Google Scholar] [CrossRef] [PubMed]
- Cornuault, J.K.; Moncaut, E.; Loux, V.; Mathieu, A.; Sokol, H.; Petit, M.-A.; De Paepe, M. The enemy from within: A prophage of Roseburia intestinalis systematically turns lytic in the mouse gut, driving bacterial adaptation by CRISPR spacer acquisition. ISME J. 2020, 14, 771–787. [Google Scholar] [CrossRef]
- Mehla, J.; Lacoursiere, S.; Stuart, E.; McDonald, R.J.; Mohajerani, M.H. Gradual Cerebral Hypoperfusion Impairs Fear Conditioning and Object Recognition Learning and Memory in Mice: Potential Roles of Neurodegeneration and Cholinergic Dysfunction. J. Alzheimer’s Dis. 2018, 61, 283–293. [Google Scholar] [CrossRef]
- Bak, J.; Pyeon, H.-I.; Seok, J.-I.; Choi, Y.-S. Effect of rotation preference on spontaneous alternation behavior on Y maze and introduction of a new analytical method, entropy of spontaneous alternation. Behav. Brain Res. 2017, 320, 219–224. [Google Scholar] [CrossRef]
- Wang, T.; Feng, W.; Ju, M.; Yu, H.; Guo, Z.; Sun, X.; Yang, K.; Liu, M.; Xiao, R. 27-hydroxycholesterol causes cognitive deficits by disturbing Th17/Treg balance and the related immune responses in mild cognitive impairment patients and C57BL/6J mice. J. Neuroinflamm. 2023, 20, 305. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, R.; Jiang, H.; Farhat, N.Y.; Carrillo-Carrasco, N.; Woolery, M.; Ottinger, E.; Porter, F.D.; Schaffer, J.E.; Ory, D.S.; Jiang, X. A validated LC-MS/MS assay for quantification of 24(S)-hydroxycholesterol in plasma and cerebrospinal fluid. J. Lipid Res. 2015, 56, 1222–1233. [Google Scholar] [CrossRef]
- Gamba, P.; Giannelli, S.; Staurenghi, E.; Testa, G.; Sottero, B.; Biasi, F.; Poli, G.; Leonarduzzi, G. The Controversial Role of 24-S-Hydroxycholesterol in Alzheimer’s Disease. Antioxidants 2021, 10, 740. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; An, Y.; Zhang, D.; Yu, H.; Zhang, X.; Wang, Y.; Tao, L.; Xiao, R. 27-Hydroxycholesterol Alters Synaptic Structural and Functional Plasticity in Hippocampal Neuronal Cultures. J. Neuropathol. Exp. Neurol. 2019, 78, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Shafik, A.M.; Zhang, F.; Guo, Z.; Dai, Q.; Pajdzik, K.; Li, Y.; Kang, Y.; Yao, B.; Wu, H.; He, C.; et al. N6-methyladenosine dynamics in neurodevelopment and aging, and its potential role in Alzheimer’s disease. Genome Biol. 2021, 22, 17. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.-J.; You, X.-J.; Ding, J.-H.; Zhang, Y.-F.; Yuan, B.-F.; Feng, Y.-Q. Identification of Inosine and 2′-O-Methylinosine Modifications in Yeast Messenger RNA by Liquid Chromatography-Tandem Mass Spectrometry Analysis. Anal. Chem. 2022, 94, 4747–4755. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xie, J.; Tan, L.; Lu, Y.; Shen, N.; Li, J.; Hu, H.; Li, H.; Li, X.; Cheng, L. N6-methyladenosine-modified circRIMS2 mediates synaptic and memory impairments by activating GluN2B ubiquitination in Alzheimer’s disease. Transl. Neurodegener. 2023, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Li, R.; Zhang, Z.; Liu, J.; Du, J.; Wei, H. Identification of altered exosomal microRNAs and mRNAs in Alzheimer’s disease. Ageing Res. Rev. 2022, 73, 101497. [Google Scholar] [CrossRef]
- Balusu, S.; Horré, K.; Thrupp, N.; Craessaerts, K.; Snellinx, A.; Serneels, L.; T’Syen, D.; Chrysidou, I.; Arranz, A.M.; Sierksma, A.; et al. MEG3 activates necroptosis in human neuron xenografts modeling Alzheimer’s disease. Science 2023, 381, 1176–1182. [Google Scholar] [CrossRef]
- Nie, K.; Ma, K.; Luo, W.; Shen, Z.; Yang, Z.; Xiao, M.; Tong, T.; Yang, Y.; Wang, X. Roseburia intestinalis: A Beneficial Gut Organism From the Discoveries in Genus and Species. Front. Cell. Infect. Microbiol. 2021, 11, 757718. [Google Scholar] [CrossRef]
- Kang, X.; Liu, C.; Ding, Y.; Ni, Y.; Ji, F.; Lau, H.C.H.; Jiang, L.; Sung, J.J.; Wong, S.H.; Yu, J. Roseburia intestinalis generated butyrate boosts anti-PD-1 efficacy in colorectal cancer by activating cytotoxic CD8+ T cells. Gut 2023, 72, 2112–2122. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, K.; Krautkramer, K.A.; Org, E.; Romano, K.A.; Kerby, R.L.; Vivas, E.I.; Mehrabian, M.; Denu, J.M.; Bäckhed, F.; Lusis, A.J.; et al. Interactions between Roseburia intestinalis and diet modulate atherogenesis in a murine model. Nat. Microbiol. 2018, 3, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.; Jeon, K.; Moon, S.; Lee, K.; Kim, W.-K.; Jeong, H.; Cha, K.H.; Lim, M.Y.; Kang, W.; Kweon, M.-N. Roseburia spp. Abundance Associates with Alcohol Consumption in Humans and Its Administration Ameliorates Alcoholic Fatty Liver in Mice. Cell Host Microbe 2020, 27, 25–40.e6. [Google Scholar] [CrossRef] [PubMed]
- Ruan, G.; Chen, M.; Chen, L.; Xu, F.; Xiao, Z.; Yi, A.; Tian, Y.; Ping, Y.; Lv, L.; Cheng, Y.; et al. Roseburia intestinalis and Its Metabolite Butyrate Inhibit Colitis and Upregulate TLR5 through the SP3 Signaling Pathway. Nutrients 2022, 14, 3041. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pan, S.; Luo, W.; Shen, Z.; Meng, X.; Xiao, M.; Tan, B.; Nie, K.; Tong, T.; Wang, X. Roseburia intestinalis-derived flagellin ameliorates colitis by targeting miR-223-3p-mediated activation of NLRP3 inflammasome and pyroptosis. Mol. Med. Rep. 2020, 22, 2695–2704. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.M.; Mulder, I.E.; Travis, A.J.; Lan, A.; Cerf-Bensussan, N.; Gaboriau-Routhiau, V.; Garden, K.; Logan, E.; Delday, M.I.; Coutts, A.G.P.; et al. Human Gut Symbiont Roseburia hominis Promotes and Regulates Innate Immunity. Front. Immunol. 2017, 8, 1166. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; He, M.; Sun, Q.; Wang, Y.; Zhang, J.; Fang, Y.; Liu, S.; Duan, L. Roseburia hominis Increases Intestinal Melatonin Level by Activating p-CREB-AANAT Pathway. Nutrients 2021, 14, 117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Peng, Q.; Huo, D.; Jiang, S.; Ma, C.; Chang, H.; Chen, K.; Li, C.; Pan, Y.; Zhang, J. Melatonin Regulates the Neurotransmitter Secretion Disorder Induced by Caffeine through the Microbiota-Gut-Brain Axis in Zebrafish (Danio rerio). Front. Cell Dev. Biol. 2021, 9, 678190. [Google Scholar] [CrossRef]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut-Brain Axis. J. Cell Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Quantifier Transition | Qualifier Transition | DP | CE | Retention Time (min) |
|---|---|---|---|---|---|
| 27-OHC | 307 → 124 | 307 → 490.2 | 130 | 20, 10 | 9.6 |
| 27-OHC-D5 | 309.6 → 124 | 309.6 → 495.2 | 130 | 20, 10 | 9.37 |
| 24S-OHC | 307 → 124 | 307 → 490 | 130 | 20, 10 | 8.87 |
| 24S-OHC-D7 | 310.7 → 124 | 310.7 → 497.3 | 130 | 20, 10 | 8.81 |
| Primer | Forward Sequence (5′–3′) | Reverse Sequence (5′–3′) |
|---|---|---|
| METTL4 | GAAAGGATGGAGGCCAGGAC | TCTCGACAGCCTCTCCTACC |
| METTL14 | CTGAGAGTG CGGATAGCATTG | GAGCAGATGTATCATAGGAAGCC |
| FTO | TCACAGACGTGGTTTCCGAG | ACCACTGGGTTGAGAGGAGT |
| WTAP | TAATGGCGAAGTGTCGAATG | CTGCTGTCGTGTCTCCTTCA |
| YTHDF1 | CTGCAGTTAAGACGGTGGGT | TAGCAATGGCTGCCCATGAA |
| BDNF | TAACGGCGGCAGACAAAAAGA | GAAGTATTGCTTCAGTTGGCCT |
| PSD-95 | CGATTACCACTTTGTCTCCTCCC | ACGGATGAAGATGGCGATAGG |
| β-actin | ATGACCCAAGCCGAGAAGG | TGCAATGACGTGAGGAACACT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Zhou, C.; Ju, M.; Feng, W.; Guo, Z.; Qi, C.; Yang, K.; Xiao, R. Roseburia intestinalis Supplementation Could Reverse the Learning and Memory Impairment and m6A Methylation Modification Decrease Caused by 27-Hydroxycholesterol in Mice. Nutrients 2024, 16, 1288. https://doi.org/10.3390/nu16091288
Sun X, Zhou C, Ju M, Feng W, Guo Z, Qi C, Yang K, Xiao R. Roseburia intestinalis Supplementation Could Reverse the Learning and Memory Impairment and m6A Methylation Modification Decrease Caused by 27-Hydroxycholesterol in Mice. Nutrients. 2024; 16(9):1288. https://doi.org/10.3390/nu16091288
Chicago/Turabian StyleSun, Xuejing, Cui Zhou, Mengwei Ju, Wenjing Feng, Zhiting Guo, Chengyan Qi, Kexin Yang, and Rong Xiao. 2024. "Roseburia intestinalis Supplementation Could Reverse the Learning and Memory Impairment and m6A Methylation Modification Decrease Caused by 27-Hydroxycholesterol in Mice" Nutrients 16, no. 9: 1288. https://doi.org/10.3390/nu16091288