
Mechanism of Action of Melatonin as a Potential Adjuvant Therapy in Inflammatory Bowel Disease and Colorectal Cancer
Abstract
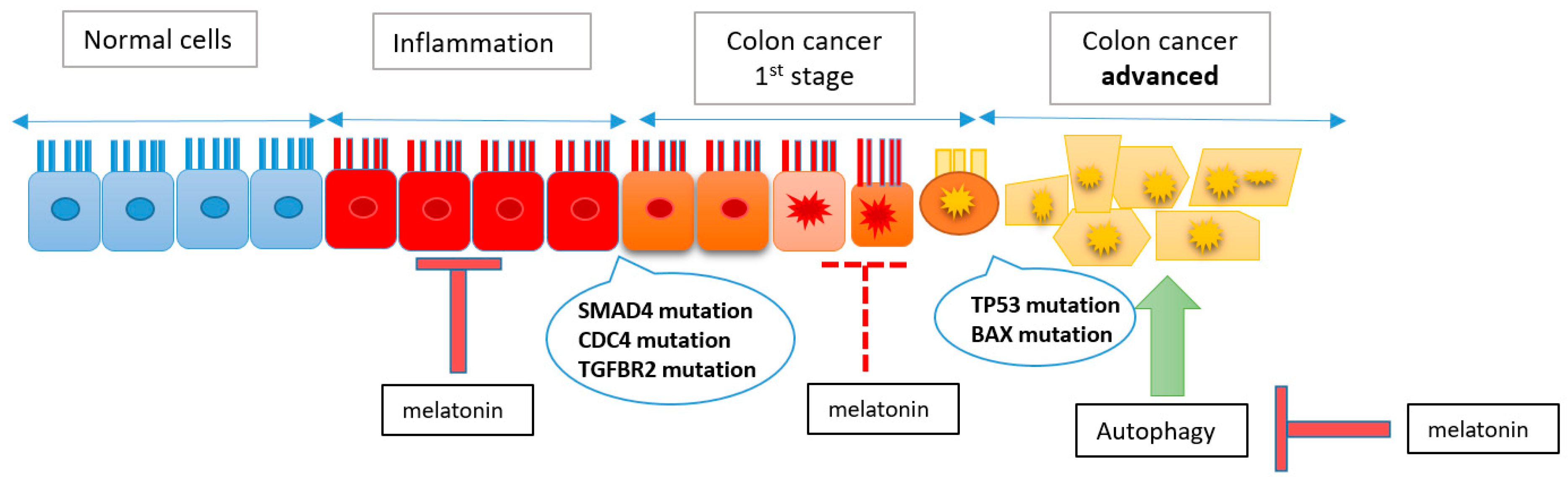
:1. Introduction
2. Melatonin: Structure and Function
3. Melatonin in the Gut
4. Melatonin and GM Interplay
4.1. Melatonin and Dysbiosis
4.2. Circadian Rhythm and Microbiota
4.3. Anti-Oxidation Effect of Melatonin
5. Melatonin and IBD
5.1. Intestinal Barrier, Melatonin, and IBD
5.2. Intestinal Microbiota, Melatonin, and IBD
6. Melatonin as Anticancer Medication in CRC
6.1. Proliferation Inhibitors
6.2. Apoptosis Activation and Melatonin
6.3. Angiogenesis Inhibition and Melatonin
6.4. Melatonin as an Antioxidant
7. Immune System, Melatonin, and IBD
7.1. Immunomodulation
7.2. Neutrophils
7.3. Macrophages
7.4. Natural Killers (NKs)
7.5. T Lymphocytes
8. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pandi-Perumal, S.R.; Zisapel, N.; Srinivasan, V.; Cardinali, D.P. Melatonin and Sleep in Aging Population. Exp. Gerontol. 2005, 40, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s Most Versatile Biological Signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef] [PubMed]
- Claustrat, B.; Leston, J. Melatonin: Physiological Effects in Humans. Neurochirurgie 2015, 61, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Karasek, M.; Reiter, R.J.; Cardinali, D.P.; Pawlikowski, M. Future of Melatonin as a Therapeutic Agent. Neuro Endocrinol. Lett. 2002, 23 (Suppl. S1), 118–121. [Google Scholar] [PubMed]
- Bruner, L.P.; White, A.M.; Proksell, S. Inflammatory Bowel Disease. Prim. Care 2023, 50, 411–427. [Google Scholar] [CrossRef] [PubMed]
- Sairenji, T.; Collins, K.L.; Evans, D.V. An Update on Inflammatory Bowel Disease. Prim. Care Clin. Off. Pract. 2017, 44, 673–692. [Google Scholar] [CrossRef] [PubMed]
- Moura, F.A.; de Andrade, K.Q.; dos Santos, J.C.F.; Araújo, O.R.P.; Goulart, M.O.F. Antioxidant Therapy for Treatment of Inflammatory Bowel Disease: Does It Work? Redox Biol. 2015, 6, 617–639. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Zubero, E.; López-Pingarrón, L.; Alatorre-Jiménez, M.A.; Ochoa-Moneo, P.; Buisac-Ramón, C.; Rivas-Jiménez, M.; Castán-Ruiz, S.; Antoñanzas-Lombarte, Á.; Tan, D.-X.; García, J.J.; et al. Melatonin’s Role as a Co-Adjuvant Treatment in Colonic Diseases: A Review. Life Sci. 2017, 170, 72–81. [Google Scholar] [CrossRef]
- Vaghari-Tabari, M.; Moein, S.; Alipourian, A.; Qujeq, D.; Malakoti, F.; Alemi, F.; Yousefi, B.; Khazaie, S. Melatonin and Inflammatory Bowel Disease: From Basic Mechanisms to Clinical Application. Biochimie 2023, 209, 20–36. [Google Scholar] [CrossRef]
- Mehrzadi, S.; Sheibani, M.; Koosha, F.; Alinaghian, N.; Pourhanifeh, M.H.; Tabaeian, S.A.P.; Reiter, R.J.; Hosseinzadeh, A. Protective and Therapeutic Potential of Melatonin against Intestinal Diseases: Updated Review of Current Data Based on Molecular Mechanisms. Expert. Rev. Gastroenterol. Hepatol. 2023, 17, 1011–1029. [Google Scholar] [CrossRef] [PubMed]
- Dyson, J.K.; Rutter, M.D. Colorectal Cancer in Inflammatory Bowel Disease: What Is the Real Magnitude of the Risk? World J. Gastroenterol. 2012, 18, 3839–3848. [Google Scholar] [CrossRef] [PubMed]
- Baidoun, F.; Elshiwy, K.; Elkeraie, Y.; Merjaneh, Z.; Khoudari, G.; Sarmini, M.T.; Gad, M.; Al-Husseini, M.; Saad, A. Colorectal Cancer Epidemiology: Recent Trends and Impact on Outcomes. Curr. Drug Targets 2021, 22, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Bretthauer, M.; Løberg, M.; Wieszczy, P.; Kalager, M.; Emilsson, L.; Garborg, K.; Rupinski, M.; Dekker, E.; Spaander, M.; Bugajski, M.; et al. Effect of Colonoscopy Screening on Risks of Colorectal Cancer and Related Death. N. Engl. J. Med. 2022, 387, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Colon Cancer Treatment—NCI. Available online: https://www.cancer.gov/types/colorectal/patient/colon-treatment-pdq (accessed on 21 January 2024).
- Kvietkauskas, M.; Zitkute, V.; Leber, B.; Strupas, K.; Stiegler, P.; Schemmer, P. The Role of Melatonin in Colorectal Cancer Treatment: A Comprehensive Review. Ther. Adv. Med. Oncol. 2020, 12, 1758835920931714. [Google Scholar] [CrossRef] [PubMed]
- Iravani, S.; Eslami, P.; Dooghaie Moghadam, A.; Moazzami, B.; Mehrvar, A.; Hashemi, M.R.; Mansour-Ghanaei, F.; Mansour-Ghanaei, A.; Majidzadeh-A, K. The Role of Melatonin in Colorectal Cancer. J. Gastrointest. Cancer 2020, 51, 748–753. [Google Scholar] [CrossRef] [PubMed]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Melo-González, F.; Sepúlveda-Alfaro, J.; Schultz, B.M.; Suazo, I.D.; Boone, D.L.; Kalergis, A.M.; Bueno, S.M. Distal Consequences of Mucosal Infections in Intestinal and Lung Inflammation. Front. Immunol. 2022, 13, 877533. [Google Scholar] [CrossRef]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A Review of Colorectal Cancer in Terms of Epidemiology, Risk Factors, Development, Symptoms and Diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef]
- Li, J.; Zhang, A.; Wu, F.; Wang, X. Alterations in the Gut Microbiota and Their Metabolites in Colorectal Cancer: Recent Progress and Future Prospects. Front. Oncol. 2022, 12, 841552. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. Bacterial Infection and Microbiota in Carcinogenesis and Tumor Development. Front. Cell. Infect. Microbiol. 2023, 13, 1294082. [Google Scholar] [CrossRef] [PubMed]
- Masters, A.; Pandi-Perumal, S.R.; Seixas, A.; Girardin, J.-L.; McFarlane, S.I. Melatonin, the Hormone of Darkness: From Sleep Promotion to Ebola Treatment. Brain Disord. Ther. 2014, 4, 1000151. [Google Scholar] [CrossRef]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; Fougerou, C. Melatonin: Pharmacology, Functions and Therapeutic Benefits. Curr. Neuropharmacol. 2017, 15, 434–443. [Google Scholar] [CrossRef]
- Kvetnoy, I.; Ivanov, D.; Mironova, E.; Evsyukova, I.; Nasyrov, R.; Kvetnaia, T.; Polyakova, V. Melatonin as the Cornerstone of Neuroimmunoendocrinology. Int. J. Mol. Sci. 2022, 23, 1835. [Google Scholar] [CrossRef]
- Arendt, J.; Aulinas, A. Physiology of the Pineal Gland and Melatonin. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Choi, D. Potency of Melatonin in Living Beings. Dev. Reprod. 2013, 17, 149–177. [Google Scholar] [CrossRef]
- When Our Cells Sleep: Quiescence and Renaissance | Institut Curie. Available online: https://institut-curie.org/page/when-our-cells-sleep-quiescence-and-renaissance (accessed on 18 April 2024).
- Peuhkuri, K.; Sihvola, N.; Korpela, R. Dietary Factors and Fluctuating Levels of Melatonin. Food Nutr. Res. 2012, 56, 17252. [Google Scholar] [CrossRef]
- Rzepka-Migut, B.; Paprocka, J. Melatonin-Measurement Methods and the Factors Modifying the Results. A Systematic Review of the Literature. Int. J. Environ. Res. Public Health 2020, 17, 1916. [Google Scholar] [CrossRef] [PubMed]
- Saidi, O.; Rochette, E.; Doré, É.; Maso, F.; Raoux, J.; Andrieux, F.; Fantini, M.L.; Merlin, E.; Pereira, B.; Walrand, S.; et al. Randomized Double-Blind Controlled Trial on the Effect of Proteins with Different Tryptophan/Large Neutral Amino Acid Ratios on Sleep in Adolescents: The Protmorpheus Study. Nutrients 2020, 12, 1885. [Google Scholar] [CrossRef]
- Voiculescu, S.; Zygouropoulos, N.; Zahiu, C.; Zagrean, A. Role of Melatonin in Embryo Fetal Development. J. Med. Life 2014, 7, 488–492. [Google Scholar]
- Rath, M.F.; Coon, S.L.; Amaral, F.G.; Weller, J.L.; Møller, M.; Klein, D.C. Melatonin Synthesis: Acetylserotonin O-Methyltransferase (ASMT) Is Strongly Expressed in a Subpopulation of Pinealocytes in the Male Rat Pineal Gland. Endocrinology 2016, 157, 2028–2040. [Google Scholar] [CrossRef]
- Soliman, A.; Lacasse, A.-A.; Lanoix, D.; Fagundes, L.; Boulard, V.; Vaillancourt, C. Placental Melatonin System Is Present throughout Pregnancy and Regulates Villous Trophoblast Differentiation. J. Pineal Res. 2015, 59, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Q.; Fichna, J.; Bashashati, M.; Li, Y.-Y.; Storr, M. Distribution, Function and Physiological Role of Melatonin in the Lower Gut. World J. Gastroenterol. WJG 2011, 17, 3888. [Google Scholar] [CrossRef]
- Iesanu, M.I.; Zahiu, C.D.M.; Dogaru, I.-A.; Chitimus, D.M.; Pircalabioru, G.G.; Voiculescu, S.E.; Isac, S.; Galos, F.; Pavel, B.; O’Mahony, S.M.; et al. Melatonin-Microbiome Two-Sided Interaction in Dysbiosis-Associated Conditions. Antioxidants 2022, 11, 2244. [Google Scholar] [CrossRef]
- Chojnacki, C.; Wiśniewska-Jarosińska, M.; Kulig, G.; Majsterek, I.; Reiter, R.J.; Chojnacki, J. Evaluation of Enterochromaffin Cells and Melatonin Secretion Exponents in Ulcerative Colitis. World J. Gastroenterol. 2013, 19, 3602–3607. [Google Scholar] [CrossRef]
- Stefulj, J.; Hörtner, M.; Ghosh, M.; Schauenstein, K.; Rinner, I.; Wölfler, A.; Semmler, J.; Liebmann, P.M. Gene Expression of the Key Enzymes of Melatonin Synthesis in Extrapineal Tissues of the Rat. J. Pineal Res. 2001, 30, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Paulose, J.K.; Wang, C.; O’Hara, B.F.; Cassone, V.M. The Effects of Aging on Sleep Parameters in a Healthy, Melatonin-Competent Mouse Model. Nat. Sci. Sleep 2019, 11, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Maitra, S.K. Gut Melatonin in Vertebrates: Chronobiology and Physiology. Front. Endocrinol. 2015, 6, 112. [Google Scholar] [CrossRef]
- Liu, J.; Clough, S.J.; Hutchinson, A.J.; Adamah-Biassi, E.B.; Popovska-Gorevski, M.; Dubocovich, M.L. MT1 and MT2 Melatonin Receptors: A Therapeutic Perspective. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 361–383. [Google Scholar] [CrossRef]
- Jockers, R.; Maurice, P.; Boutin, J.A.; Delagrange, P. Melatonin Receptors, Heterodimerization, Signal Transduction and Binding Sites: What’s New? Br. J. Pharmacol. 2008, 154, 1182–1195. [Google Scholar] [CrossRef]
- Parandavar, N.; Hojat, M.; Abdali, K.; Keshtgar, S.; Emamghoreishi, M.; Yeganeh, B.S. The Effect of Melatonin on the Lipid Levels in Menopausal Women: A Double-Blind, Controlled, Clinical Trial. J. Educ. Health Promot. 2018, 7, 144. [Google Scholar] [CrossRef]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin Membrane Receptors in Peripheral Tissues: Distribution and Functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef]
- Neish, A.S. Microbes in Gastrointestinal Health and Disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, F.; Gao, Z.; Huang, W.; Zhang, X.; Liu, F.; Yi, H.; Guan, J.; Wu, X.; Xu, H.; et al. Melatonin Attenuates Chronic Intermittent Hypoxia-Induced Intestinal Barrier Dysfunction in Mice. Microbiol. Res. 2023, 276, 127480. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lai, J.; Ma, F.; Cai, Y.; Li, S.; Feng, Z.; Lu, Z.; Liu, X.; Ke, Q.; Hao, H.; et al. Maternal Melatonin Supplementation Shapes Gut Microbiota and Protects against Inflammation in Early Life. Int. Immunopharmacol. 2023, 120, 110359. [Google Scholar] [CrossRef]
- Diez-Echave, P.; Vezza, T.; Algieri, F.; Ruiz-Malagón, A.J.; Hidalgo-García, L.; García, F.; Moron, R.; Sánchez, M.; Toral, M.; Romero, M.; et al. The Melatonergic Agonist Agomelatine Ameliorates High Fat Diet-Induced Obesity in Mice through the Modulation of the Gut Microbiome. Biomed. Pharmacother. 2022, 153, 113445. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez-Rodriguez, A.; Roman, P.; Rueda-Ruzafa, L.; Campos-Rios, A.; Cardona, D. Changes in Gut Microbiota and Multiple Sclerosis: A Systematic Review. Int. J. Environ. Res. Public Health 2023, 20, 4624. [Google Scholar] [CrossRef]
- Muñoz-Jurado, A.; Escribano, B.M.; Caballero-Villarraso, J.; Galván, A.; Agüera, E.; Santamaría, A.; Túnez, I. Melatonin and Multiple Sclerosis: Antioxidant, Anti-Inflammatory and Immunomodulator Mechanism of Action. Inflammopharmacology 2022, 30, 1569–1596. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, T.; Cao, M.; Yuan, C.; Reiter, R.J.; Zhao, Z.; Zhao, Y.; Chen, L.; Fan, W.; Wang, X.; et al. Gut Microbiota Dysbiosis Induced by Decreasing Endogenous Melatonin Mediates the Pathogenesis of Alzheimer’s Disease and Obesity. Front. Immunol. 2022, 13, 900132. [Google Scholar] [CrossRef] [PubMed]
- Ballester, P.; Zafrilla, P.; Arcusa, R.; Galindo, A.; Cerdá, B.; Marhuenda, J. Food as a Dietary Source of Melatonin and Its Role in Human Health: Present and Future Perspectives. In Current Topics in Functional Food; IntechOpen: Rijeka, Croatia, 2022. [Google Scholar] [CrossRef]
- Bubenik, G.A.; Brown, G.M. Pinealectomy Reduces Melatonin Levels in the Serum but Not in the Gastrointestinal Tract of Rats. Neurosignals 1997, 6, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Bubenik, G.A. Localization, Physiological Significance and Possible Clinical Implication of Gastrointestinal Melatonin. Biol. Signals Recept. 2001, 10, 350–366. [Google Scholar] [CrossRef] [PubMed]
- Rosenwasser, A.M.; Turek, F.W. Neurobiology of Circadian Rhythm Regulation. Sleep Med. Clin. 2015, 10, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.E.; Al-Ghoul, W.M.; Gillette, M.U.; Dubocovich, M.L. Activation of MT(2) Melatonin Receptors in Rat Suprachiasmatic Nucleus Phase Advances the Circadian Clock. Am. J. Physiol. Cell Physiol. 2001, 280, C110–C118. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Weaver, D.R.; Jin, X.; Shearman, L.P.; Pieschl, R.L.; Gribkoff, V.K.; Reppert, S.M. Molecular Dissection of Two Distinct Actions of Melatonin on the Suprachiasmatic Circadian Clock. Neuron 1997, 19, 91–102. [Google Scholar] [CrossRef]
- Ashkar, F.; Wu, J. Effects of Food Factors and Processing on Protein Digestibility and Gut Microbiota. J. Agric. Food Chem. 2023, 71, 8685–8698. [Google Scholar] [CrossRef] [PubMed]
- Bubenik, G.A. Gastrointestinal Melatonin: Localization, Function, and Clinical Relevance. Dig. Dis. Sci. 2002, 47, 2336–2348. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Delgado, R.C.; Espinosa-Tanguma, R.; Valdés Abadía, B.; Ramírez-Plascencia, O.D.; Escobar, C.; Saderi, N. Feeding during the Resting Phase Causes Gastrointestinal Tract Dysfunction and Desynchronization of Metabolic and Neuronal Rhythms in Rats. Neurogastroenterol. Motil. 2023, 35, e14687. [Google Scholar] [CrossRef]
- Matenchuk, B.A.; Mandhane, P.J.; Kozyrskyj, A.L. Sleep, Circadian Rhythm, and Gut Microbiota. Sleep Med. Rev. 2020, 53, 101340. [Google Scholar] [CrossRef]
- Chen, M.; Wang, Z.; Tan, D.S.; Wang, X.; Ye, Z.; Xie, Z.; Zhang, D.; Wu, D.; Zhao, Y.; Qu, Y.; et al. The Causal Relationship between the Morning Chronotype and the Gut Microbiota: A Bidirectional Two-Sample Mendelian Randomization Study. Nutrients 2023, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Xiang, X.; Shi, Y.; Qiu, A.; Luo, X.; Xie, J.; Russell, R.; Zhang, D. Chronic Jet Lag Alters Gut Microbiome and Mycobiome and Promotes the Progression of MAFLD in HFHFD-Fed Mice. Front. Microbiol. 2023, 14, 1295869. [Google Scholar] [CrossRef] [PubMed]
- Siebieszuk, A.; Sejbuk, M.; Witkowska, A.M. Studying the Human Microbiota: Advances in Understanding the Fundamentals, Origin, and Evolution of Biological Timekeeping. Int. J. Mol. Sci. 2023, 24, 16169. [Google Scholar] [CrossRef] [PubMed]
- Mehrzadi, S.; Pourhanifeh, M.H.; Mirzaei, A.; Moradian, F.; Hosseinzadeh, A. An Updated Review of Mechanistic Potentials of Melatonin against Cancer: Pivotal Roles in Angiogenesis, Apoptosis, Autophagy, Endoplasmic Reticulum Stress and Oxidative Stress. Cancer Cell Int. 2021, 21, 188. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Gao, M.; Zhang, S.; Dai, T.; Wang, F.; Geng, J.; Rao, J.; Qin, X.; Qian, J.; Zuo, L.; et al. Sleep Deprivation Impairs Intestinal Mucosal Barrier by Activating Endoplasmic Reticulum Stress in Goblet Cells. Am. J. Pathol. 2024, 194, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. The Role of Aeromonas-Goblet Cell Interactions in Melatonin-Mediated Improvements in Sleep Deprivation-Induced Colitis. Oxid. Med. Cell. Longev. 2022, 2022, 8133310. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin Ameliorates Corticosterone-Mediated Oxidative Stress-Induced Colitis in Sleep-Deprived Mice Involving Gut Microbiota. Oxid. Med. Cell. Longev. 2021, 2021, 9981480. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Cai, R.; Shaoyong, W.; Wang, G.; Yan, W.; He, Z.; Li, R.; Chao, M.; Zhao, T.; Deng, L.; et al. Melatonin Promotes Gut Anti-Oxidative Status in Perinatal Rat by Remodeling the Gut Microbiome. Redox Biol. 2023, 65, 102829. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Mechanisms of Melatonin in Obesity: A Review. Int. J. Mol. Sci. 2021, 23, 218. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Cabrera, J.; D’Arpa, D.; Sainz, R.M.; Mayo, J.C.; Ramos, S. The Oxidant/Antioxidant Network: Role of Melatonin. Biol. Signals Recept. 1999, 8, 56–63. [Google Scholar] [CrossRef]
- Bonmatí-Carrión, M.-Á.; Rol, M.-A. Melatonin as a Mediator of the Gut Microbiota–Host Interaction: Implications for Health and Disease. Antioxidants 2024, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Antoni, L.; Nuding, S.; Wehkamp, J.; Stange, E.F. Intestinal Barrier in Inflammatory Bowel Disease. World J. Gastroenterol. 2014, 20, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Park, H.; Choe, B.-H.; Kang, B. The Role and Function of Mucins and Its Relationship to Inflammatory Bowel Disease. Front. Med. 2022, 9, 848344. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.; Shin, D.-H.; Lee, D.; Kang, S.-M.; Seok, J.-H.; Kang, H.Y.; Jeung, E.-B. Expression of Claudins, Occludin, Junction Adhesion Molecule A and Zona Occludens 1 in Canine Organs. Mol. Med. Rep. 2016, 14, 3697–3703. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Park, Y.S.; Baik, H.-W.; Jun, J.H.; Kim, E.K.; Sull, J.W.; Sung, H.J.; Choi, J.W.; Chung, S.H.; Gye, M.C.; et al. Melatonin Modulates Adiponectin Expression on Murine Colitis with Sleep Deprivation. World J. Gastroenterol. 2016, 22, 7559–7568. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Chung, S.-H.; Lee, S.-K.; Kim, J.-H.; Kim, J.-B.; Kim, T.-K.; Kim, D.-S.; Baik, H.-W. Melatonin Improves Experimental Colitis with Sleep Deprivation. Int. J. Mol. Med. 2015, 35, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.; Dahlgren, D.; Lennernäs, H.; Sjöblom, M. Melatonin-Activated Receptor Signaling Pathways Mediate Protective Effects on Surfactant-Induced Increase in Jejunal Mucosal Permeability in Rats. Int. J. Mol. Sci. 2021, 22, 10762. [Google Scholar] [CrossRef] [PubMed]
- Akcan, A.; Kucuk, C.; Sozuer, E.; Esel, D.; Akyildiz, H.; Akgun, H.; Muhtaroglu, S.; Aritas, Y. Melatonin Reduces Bacterial Translocation and Apoptosis in Trinitrobenzene Sulphonic Acid-Induced Colitis of Rats. World J. Gastroenterol. 2008, 14, 918–924. [Google Scholar] [CrossRef]
- Mannino, G.; Caradonna, F.; Cruciata, I.; Lauria, A.; Perrone, A.; Gentile, C. Melatonin Reduces Inflammatory Response in Human Intestinal Epithelial Cells Stimulated by Interleukin-1β. J. Pineal Res. 2019, 67, e12598. [Google Scholar] [CrossRef]
- Favero, G.; Franceschetti, L.; Bonomini, F.; Rodella, L.F.; Rezzani, R. Melatonin as an Anti-Inflammatory Agent Modulating Inflammasome Activation. Int. J. Endocrinol. 2017, 2017, 1835195. [Google Scholar] [CrossRef]
- Ikram, M.; Park, H.Y.; Ali, T.; Kim, M.O. Melatonin as a Potential Regulator of Oxidative Stress, and Neuroinflammation: Mechanisms and Implications for the Management of Brain Injury-Induced Neurodegeneration. J. Inflamm. Res. 2021, 14, 6251–6264. [Google Scholar] [CrossRef] [PubMed]
- Celiberto, L.S.; Graef, F.A.; Healey, G.R.; Bosman, E.S.; Jacobson, K.; Sly, L.M.; Vallance, B.A. Inflammatory Bowel Disease and Immunonutrition: Novel Therapeutic Approaches through Modulation of Diet and the Gut Microbiome. Immunology 2018, 155, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Peng, K.; Xiao, S.; Long, Y.; Yu, Q. The Role of Lactobacillus in Inflammatory Bowel Disease: From Actualities to Prospects. Cell Death Discov. 2023, 9, 361. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Choi, J.W.; Jhun, J.; Kwon, J.Y.; Lee, B.-I.; Yang, C.W.; Park, S.-H.; Cho, M.-L. Lactobacillus Acidophilus Improves Intestinal Inflammation in an Acute Colitis Mouse Model by Regulation of Th17 and Treg Cell Balance and Fibrosis Development. J. Med. Food 2018, 21, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Kim, S.H.; Park, J.W.; Kho, Y.; Seok, P.R.; Shin, J.-H.; Choi, Y.J.; Jun, J.-H.; Jung, H.C.; Kim, E.K. Melatonin in the Colon Modulates Intestinal Microbiota in Response to Stress and Sleep Deprivation. Intest. Res. 2020, 18, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lang, R.; Guo, S.; Luo, X.; Li, H.; Liu, C.; Dong, W.; Bao, C.; Yu, Y. Intestinal Microbiota and Melatonin in the Treatment of Secondary Injury and Complications after Spinal Cord Injury. Front. Neurosci. 2022, 16, 981772. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Kim, S.; Son, M.; Cheon, J.H.; Park, Y.S. Melatonin Controls Microbiota in Colitis by Goblet Cell Differentiation and Antimicrobial Peptide Production through Toll-like Receptor 4 Signalling. Sci. Rep. 2020, 10, 2232. [Google Scholar] [CrossRef] [PubMed]
- Lutfi, E.; Basili, D.; Falcinelli, S.; Morillas, L.; Carnevali, O.; Capilla, E.; Navarro, I. The Probiotic Lactobacillus Rhamnosus Mimics the Dark-Driven Regulation of Appetite Markers and Melatonin Receptors’ Expression in Zebrafish (Danio rerio) Larvae: Understanding the Role of the Gut Microbiome. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2021, 256, 110634. [Google Scholar] [CrossRef]
- Williams, B.B.; Benschoten, A.H.V.; Cimermancic, P.; Donia, M.S.; Zimmermann, M.; Taketani, M.; Ishihara, A.; Kashyap, P.C.; Fraser, J.S.; Fischbach, M.A. Discovery and Characterization of Gut Microbiota Decarboxylases That Can Produce the Neurotransmitter Tryptamine. Cell Host Microbe 2014, 16, 495. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial Tryptophan Catabolites in Health and Disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef]
- Liu, B.; Fan, L.; Wang, Y.; Wang, H.; Yan, Y.; Chen, S.; Hung, I.; Liu, C.; Wei, H.; Ge, L.; et al. Gut Microbiota Regulates Host Melatonin Production through Epithelial Cell MyD88. Gut Microbes 2024, 16, 2313769. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; He, M.; Sun, Q.; Wang, Y.; Zhang, J.; Fang, Y.; Liu, S.; Duan, L. Roseburia Hominis Increases Intestinal Melatonin Level by Activating P-CREB-AANAT Pathway. Nutrients 2021, 14, 117. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-N.; Yang, H.-W.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.; Tain, Y.-L. Melatonin Prevents Chronic Kidney Disease-Induced Hypertension in Young Rat Treated with Adenine: Implications of Gut Microbiota-Derived Metabolites. Antioxidants 2021, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Gut Microbiota-Derived Metabolites Mediate the Neuroprotective Effect of Melatonin in Cognitive Impairment Induced by Sleep Deprivation. Microbiome 2023, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, J. Bile Acid Metabolism and Circadian Rhythms. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 319, G549–G563. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, W. Knockdown of Galectin-1 Facilitated Cisplatin Sensitivity by Inhibiting Autophagy in Neuroblastoma Cells. Chem. Biol. Interact. 2019, 297, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zou, D.; Yang, X.; Xue, X.; Zuo, L.; Zhou, Q.; Hu, R.; Wang, Y. Melatonin Inhibits Colon Cancer RKO Cell Migration by Downregulating Rho-Associated Protein Kinase Expression via the P38/MAPK Signaling Pathway. Mol. Med. Rep. 2017, 16, 9383–9392. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.-B.; Wei, X.; Hu, R.-L.; Yang, X.-P.; Zuo, L.; Zhang, S.-M.; Zhu, H.-Q.; Zhou, Q.; Gui, S.-Y.; Wang, Y. Melatonin Inhibits the Migration of Colon Cancer RKO Cells by Down-Regulating Myosin Light Chain Kinase Expression through Cross-Talk with P38 MAPK. Asian Pac. J. Cancer Prev. 2015, 16, 5835–5842. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N.; Popovich, I.G.; Zabezhinski, M.A. Melatonin and Colon Carcinogenesis: I. Inhibitory Effect of Melatonin on Development of Intestinal Tumors Induced by 1,2-Dimethylhydrazine in Rats. Carcinogenesis 1997, 18, 1549–1553. [Google Scholar] [CrossRef]
- Park, S.-Y.; Jang, W.-J.; Yi, E.-Y.; Jang, J.-Y.; Jung, Y.; Jeong, J.-W.; Kim, Y.-J. Melatonin Suppresses Tumor Angiogenesis by Inhibiting HIF-1alpha Stabilization under Hypoxia. J. Pineal Res. 2010, 48, 178–184. [Google Scholar] [CrossRef]
- León, J.; Casado, J.; Jiménez Ruiz, S.M.; Zurita, M.S.; González-Puga, C.; Rejón, J.D.; Gila, A.; Muñoz de Rueda, P.; Pavón, E.J.; Reiter, R.J.; et al. Melatonin Reduces Endothelin-1 Expression and Secretion in Colon Cancer Cells through the Inactivation of FoxO-1 and NF-Κβ. J. Pineal Res. 2014, 56, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Polidarová, L.; Houdek, P.; Sumová, A. Chronic Disruptions of Circadian Sleep Regulation Induce Specific Proinflammatory Responses in the Rat Colon. Chronobiol. Int. 2017, 34, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Pandi-Perumal, S.R.; Brzezinski, A.; Bhatnagar, K.P.; Cardinali, D.P. Melatonin, Immune Function and Cancer. Recent. Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Kossoy, G.; Ben-Hur, H.; Popovich, I.; Zabezhinski, M.; Anisimov, V.; Zusman, I. Melatonin and Colon Carcinogenesis. IV. Effect of Melatonin on Proliferative Activity and Expression of Apoptosis-Related Proteins in the Spleen of Rats Exposed to 1,2-Dimethylhydrazine. Oncol. Rep. 2000, 7, 1401–1405. [Google Scholar] [CrossRef] [PubMed]
- Farriol, M.; Venereo, Y.; Orta, X.; Castellanos, J.M.; Segovia-Silvestre, T. In Vitro Effects of Melatonin on Cell Proliferation in a Colon Adenocarcinoma Line. J. Appl. Toxicol. 2000, 20, 21–24. [Google Scholar] [CrossRef]
- Winczyk, K.; Pawlikowski, M.; Lawnicka, H.; Kunert-Radek, J.; Spadoni, G.; Tarzia, G.; Karasek, M. Effects of Melatonin and Melatonin Receptors Ligand N-[(4-Methoxy-1H-Indol-2-Yl)Methyl]Propanamide on Murine Colon 38 Cancer Growth in Vitro and in Vivo. Neuro Endocrinol. Lett. 2002, 23 (Suppl. S1), 50–54. [Google Scholar] [PubMed]
- Hong, Y.; Won, J.; Lee, Y.; Lee, S.; Park, K.; Chang, K.-T.; Hong, Y. Melatonin Treatment Induces Interplay of Apoptosis, Autophagy, and Senescence in Human Colorectal Cancer Cells. J. Pineal Res. 2014, 56, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.-Y.; Li, W.-M.; Zhou, L.-L.; Lu, Q.-N.; He, W. Melatonin Induces Apoptosis of Colorectal Cancer Cells through HDAC4 Nuclear Import Mediated by CaMKII Inactivation. J. Pineal Res. 2015, 58, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Chovancova, B.; Hudecova, S.; Lencesova, L.; Babula, P.; Rezuchova, I.; Penesova, A.; Grman, M.; Moravcik, R.; Zeman, M.; Krizanova, O. Melatonin-Induced Changes in Cytosolic Calcium Might Be Responsible for Apoptosis Induction in Tumour Cells. Cell Physiol. Biochem. 2017, 44, 763–777. [Google Scholar] [CrossRef]
- Yun, C.W.; Kim, S.; Lee, J.H.; Lee, S.H. Melatonin Promotes Apoptosis of Colorectal Cancer Cells via Superoxide-Mediated ER Stress by Inhibiting Cellular Prion Protein Expression. Anticancer. Res. 2018, 38, 3951–3960. [Google Scholar] [CrossRef]
- Kannen, V.; Marini, T.; Zanette, D.L.; Frajacomo, F.T.; Silva, G.E.B.; Silva, W.A.; Garcia, S.B. The Melatonin Action on Stromal Stem Cells within Pericryptal Area in Colon Cancer Model under Constant Light. Biochem. Biophys. Res. Commun. 2011, 405, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Louise Jones, J. Jekyll and Hyde: The Role of the Microenvironment on the Progression of Cancer. J. Pathol. 2011, 223, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Su, S.-C.; Hsieh, M.-J.; Yang, W.-E.; Chung, W.-H.; Reiter, R.J.; Yang, S.-F. Cancer Metastasis: Mechanisms of Inhibition by Melatonin. J. Pineal Res. 2017, 62, e12370. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-R.; Chu, Y.-T.; Chang, C.-L.; Yip, H.-K.; Chen, H.-H. ZNF746 Plays Cardinal Roles on Colorectal Cancer (CRC) Cell Invasion and Migration and Regulates Mitochondrial Dynamics and Morphological Changes of CRC Cells-Role of Combined Melatonin and 5-FU Regimen. J. Cell. Biochem. 2024, 125, e30507. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Zhou, W.; Li, X.; Du, J.; Li, X.; Hao, H. Melatonin Inhibits Proliferation and Viability and Promotes Apoptosis in Colorectal Cancer Cells via Upregulation of the microRNA-34a/449a Cluster. Mol. Med. Rep. 2021, 23, 187. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Guo, Y.; Hong, F.; Xu, P.; Zhai, Y. Therapeutic Potential of Melatonin in Colorectal Cancer: Focus on Lipid Metabolism and Gut Microbiota. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2022, 1868, 166281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Q.; Shi, S.-S.; Li, Y.-F.; Yang, Y.; Bai, P.; Qiao, C.-H. Melatonin Inhibits Gastric Cancer Cell Proliferation by Suppressing Exosome miR-27b-3p Expression. Anticancer. Res. 2023, 43, 4413–4424. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, H.; Dan, Z.; Shu, C.; Zhu, R.; Yang, Q.; Wang, Y.; Zhu, H. Melatonin Potentiates Sensitivity to 5-Fluorouracil in Gastric Cancer Cells by Upregulating Autophagy and Downregulating Myosin Light-Chain Kinase. J. Cancer 2023, 14, 2608–2618. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.-J.; Zong, W.-X. The Cellular Decision between Apoptosis and Autophagy. Chin. J. Cancer 2013, 32, 121–129. [Google Scholar] [CrossRef]
- Bento, C.F.; Renna, M.; Ghislat, G.; Puri, C.; Ashkenazi, A.; Vicinanza, M.; Menzies, F.M.; Rubinsztein, D.C. Mammalian Autophagy: How Does It Work? Annu. Rev. Biochem. 2016, 85, 685–713. [Google Scholar] [CrossRef]
- Gao, J.; Hou, Y.; Yang, X.; Liu, J.; Zhang, Y. Melatonin Enhances the Sensitivity of Colorectal Cancer Cells to 5-Fluorouracil through the Regulation of the miR-532-3p/β-Catenin Pathway. Environ. Toxicol. 2024, 39, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, C.; Goel, A. A Combined Treatment with Melatonin and Andrographis Promotes Autophagy and Anticancer Activity in Colorectal Cancer. Carcinogenesis 2022, 43, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Go, G.; Lee, S.H. PrPC Regulates the Cancer Stem Cell Properties via Interaction with C-Met in Colorectal Cancer Cells. Anticancer. Res. 2021, 41, 3459–3470. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yoon, Y.M.; Han, Y.-S.; Yun, C.W.; Lee, S.H. Melatonin Promotes Apoptosis of Oxaliplatin-Resistant Colorectal Cancer Cells through Inhibition of Cellular Prion Protein. Anticancer. Res. 2018, 38, 1993–2000. [Google Scholar] [CrossRef] [PubMed]
- Angiogenesis Inhibitors—NCI. Available online: https://www.cancer.gov/about-cancer/treatment/types/immunotherapy/angiogenesis-inhibitors-fact-sheet (accessed on 9 January 2024).
- Goradel, N.H.; Asghari, M.H.; Moloudizargari, M.; Negahdari, B.; Haghi-Aminjan, H.; Abdollahi, M. Melatonin as an Angiogenesis Inhibitor to Combat Cancer: Mechanistic Evidence. Toxicol. Appl. Pharmacol. 2017, 335, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Gil-Martín, E.; Egea, J.; Reiter, R.J.; Romero, A. The Emergence of Melatonin in Oncology: Focus on Colorectal Cancer. Med. Res. Rev. 2019, 39, 2239–2285. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, D.; Sharda, N.; Giri, B.; Hassan, M.S.; Singh, D.; Tarasiewicz, A.; Lohr, C.; von Holzen, U.; Kristian, T.; Waddell, J.; et al. Melatonin and Andrographolide Synergize to Inhibit the Colospheroid Phenotype by Targeting Wnt/Beta-Catenin Signaling. J. Pineal Res. 2022, 73, e12808. [Google Scholar] [CrossRef] [PubMed]
- Kvietkauskas, M.; Zitkute, V.; Leber, B.; Strupas, K.; Stiegler, P.; Schemmer, P. Dietary Melatonin and Glycine Decrease Tumor Growth through Antiangiogenic Activity in Experimental Colorectal Liver Metastasis. Nutrients 2021, 13, 2035. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zhou, L.; Huang, Z.; Li, B.; Nice, E.C.; Xu, J.; Huang, C. Antioxidant Therapy in Cancer: Rationale and Progress. Antioxidants 2022, 11, 1128. [Google Scholar] [CrossRef]
- Trivedi, P.P.; Jena, G.B.; Tikoo, K.B.; Kumar, V. Melatonin Modulated Autophagy and Nrf2 Signaling Pathways in Mice with Colitis-Associated Colon Carcinogenesis. Mol. Carcinog. 2016, 55, 255–267. [Google Scholar] [CrossRef]
- Collin, A.; Kohan, R.; Tolosa de Talamoni, N.; Picotto, G. Melatonin Enhances Anti-Tumoral Effects of Menadione on Colon Cancer Cells. Anticancer Agents Med. Chem. 2022, 22, 2411–2418. [Google Scholar] [CrossRef] [PubMed]
- Chok, K.C.; Koh, R.Y.; Ng, M.G.; Ng, P.Y.; Chye, S.M. Melatonin Induces Autophagy via Reactive Oxygen Species-Mediated Endoplasmic Reticulum Stress Pathway in Colorectal Cancer Cells. Molecules 2021, 26, 5038. [Google Scholar] [CrossRef] [PubMed]
- Abadi, S.H.M.H.; Shirazi, A.; Alizadeh, A.M.; Changizi, V.; Najafi, M.; Khalighfard, S.; Nosrati, H. The Effect of Melatonin on Superoxide Dismutase and Glutathione Peroxidase Activity, and Malondialdehyde Levels in the Targeted and the Non-Targeted Lung and Heart Tissues after Irradiation in Xenograft Mice Colon Cancer. Curr. Mol. Pharmacol. 2018, 11, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Yu, I.; Dakwar, A.; Takabe, K. Immunotherapy: Recent Advances and Its Future as a Neoadjuvant, Adjuvant, and Primary Treatment in Colorectal Cancer. Cells 2023, 12, 258. [Google Scholar] [CrossRef] [PubMed]
- Vijayalaxmi; Thomas, C.R.; Reiter, R.J.; Herman, T.S. R.; Reiter, R.J.; Herman, T.S. Melatonin: From Basic Research to Cancer Treatment Clinics. J. Clin. Oncol. 2002, 20, 2575–2601. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Liu, G.; Chen, S.; Yin, J.; Wang, J.; Tan, B.; Wu, G.; Bazer, F.W.; Peng, Y.; Li, T.; et al. Melatonin Signaling in T Cells: Functions and Applications. J. Pineal Res. 2017, 62, e12394. [Google Scholar] [CrossRef] [PubMed]
- Wallace, K.L.; Zheng, L.-B.; Kanazawa, Y.; Shih, D.Q. Immunopathology of Inflammatory Bowel Disease. World J. Gastroenterol. 2014, 20, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Okeke, E.B.; Uzonna, J.E. The Pivotal Role of Regulatory T Cells in the Regulation of Innate Immune Cells. Front. Immunol. 2019, 10, 680. [Google Scholar] [CrossRef] [PubMed]
- Kałużna, A.; Olczyk, P.; Komosińska-Vassev, K. The Role of Innate and Adaptive Immune Cells in the Pathogenesis and Development of the Inflammatory Response in Ulcerative Colitis. J. Clin. Med. 2022, 11, 400. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Muñoz, F.; Dominguez-Lopez, A.; Yamamoto-Furusho, J.K. Role of Cytokines in Inflammatory Bowel Disease. World J. Gastroenterol. 2008, 14, 4280–4288. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.J. Pro-Inflammatory Cytokines in the Pathogenesis of IBD. Gastroenterology 2011, 140, 1756–1767. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Ding, D.; Bai, D.; Zhu, Y.; Sun, W.; Sun, Y.; Zhang, D. Melatonin Biosynthesis Pathways in Nature and Its Production in Engineered Microorganisms. Synth. Syst. Biotechnol. 2022, 7, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Vico, A.; Lardone, P.J.; Álvarez-Sánchez, N.; Rodríguez-Rodríguez, A.; Guerrero, J.M. Melatonin: Buffering the Immune System. Int. J. Mol. Sci. 2013, 14, 8638. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, Z. Inflammatory Bowel Disease Related Innate Immunity and Adaptive Immunity. Am. J. Transl. Res. 2016, 8, 2490–2497. [Google Scholar] [PubMed]
- Wéra, O.; Lancellotti, P.; Oury, C. The Dual Role of Neutrophils in Inflammatory Bowel Diseases. J. Clin. Med. 2016, 5, 118. [Google Scholar] [CrossRef] [PubMed]
- Villanacci, V.; Del Sordo, R.; Parigi, T.L.; Leoncini, G.; Bassotti, G. Inflammatory Bowel Diseases: Does One Histological Score Fit All? Diagnostics 2023, 13, 2112. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Miralda, I.; Armstrong, C.L.; Uriarte, S.M.; Bagaitkar, J. The Roles of NADPH Oxidase in Modulating Neutrophil Effector Responses. Mol. Oral. Microbiol. 2019, 34, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Ivetic, A.; Hoskins Green, H.L.; Hart, S.J. L-Selectin: A Major Regulator of Leukocyte Adhesion, Migration and Signaling. Front. Immunol. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Cuzzocrea, S. Antiinflammatory Activity of Melatonin in Central Nervous System. Curr. Neuropharmacol. 2010, 8, 228. [Google Scholar] [CrossRef]
- Kirchgesner, J.; Svrcek, M.; Le Gall, G.; Landman, C.; Dray, X.; Bourrier, A.; Nion-Larmurier, I.; Hoyeau, N.; Sokol, H.; Seksik, P.; et al. Nancy Index Scores of Chronic Inflammatory Bowel Disease Activity Associate with Development of Colorectal Neoplasia. Clin. Gastroenterol. Hepatol. 2020, 18, 150–157.e1. [Google Scholar] [CrossRef]
- Seoane-Viaño, I.; Gómez-Lado, N.; Lázare-Iglesias, H.; Rey-Bretal, D.; Lamela-Gómez, I.; Otero-Espinar, F.; Mendez, J.; Antúnez-López, J.; Pasín, M.C.; Aguiar, P.; et al. Evaluation of the Therapeutic Activity of Melatonin and Resveratrol in Inflammatory Bowel Disease: A Longitudinal PET/CT Study in an Animal Model. Int. J. Pharm. 2019, 572, 118713. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Liu, S.; Li, S.; Yang, J.; Hu, G.; Xu, C.; Song, W. Psychological Stress in Inflammatory Bowel Disease: Psychoneuroimmunological Insights into Bidirectional Gut–Brain Communications. Front. Immunol. 2022, 13, 1016578. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Chen, S.; Zeng, S.; Zhao, Y.; Zhu, C.; Deng, B.; Zhu, G.; Yin, Y.; Wang, W.; Hardeland, R.; et al. Melatonin in Macrophage Biology: Current Understanding and Future Perspectives. J. Pineal Res. 2019, 66, e12547. [Google Scholar] [CrossRef] [PubMed]
- Leopold Wager, C.M.; Wormley, F.L. Classical versus Alternative Macrophage Activation: The Ying and the Yang in Host Defense against Pulmonary Fungal Infections. Mucosal Immunol. 2014, 7, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Dev, K.; Agarwal, B.; Das, P.; Syed, M.A. Macrophages: Their Role, Activation and Polarization in Pulmonary Diseases. Immunobiology 2018, 223, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Arango Duque, G.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, H.; Wang, X.; Jiang, G.; Liu, H.; Zhang, G.; Wang, H.; Fang, R.; Bu, X.; Cai, S.; et al. TGF-β Induces M2-like Macrophage Polarization via SNAIL-Mediated Suppression of a pro-Inflammatory Phenotype. Oncotarget 2016, 7, 52294–52306. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Ismail, N. M1 and M2 Macrophages Polarization via mTORC1 Influences Innate Immunity and Outcome of Ehrlichia Infection. J. Cell. Immunol. 2020, 2, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Guo, J.; Yan, W.; Xu, L. Macrophage Polarization in Inflammatory Bowel Disease. Cell Commun. Signal 2023, 21, 367. [Google Scholar] [CrossRef]
- Ames, C.; Boland, E.; Szentirmai, É. Effects of Macrophage Depletion on Sleep in Mice. PLoS ONE 2016, 11, e0159812. [Google Scholar] [CrossRef]
- Zheng, N.; Long, Y.; Bai, Z.; Li, J.; Wang, H.; Song, D.-D.; Liu, H.-L.; Shi, J.-H.; Zhao, S. Melatonin as an Immunomodulator in CD19-Targeting CAR-T Cell Therapy: Managing Cytokine Release Syndrome. J. Transl. Med. 2024, 22, 58. [Google Scholar] [CrossRef] [PubMed]
- Rébé, C.; Ghiringhelli, F. STAT3, a Master Regulator of Anti-Tumor Immune Response. Cancers 2019, 11, 1280. [Google Scholar] [CrossRef] [PubMed]
- Lünemann, A.; Lünemann, J.D.; Münz, C. Regulatory NK-Cell Functions in Inflammation and Autoimmunity. Mol. Med. 2009, 15, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Steel, A.W.; Mela, C.M.; Lindsay, J.O.; Gazzard, B.G.; Goodier, M.R. Increased Proportion of CD16(+) NK Cells in the Colonic Lamina Propria of Inflammatory Bowel Disease Patients, but Not after Azathioprine Treatment. Aliment. Pharmacol. Ther. 2011, 33, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Mallick, B.; Malik, S. Use of Azathioprine in Ulcerative Colitis: A Comprehensive Review. Cureus 2022, 14, e24874. [Google Scholar] [CrossRef] [PubMed]
- Mah, A.Y.; Cooper, M.A. Metabolic Regulation of Natural Killer Cell IFN-γ Production. Crit. Rev. Immunol. 2016, 36, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Overview of Azathioprine and Mercaptopurine Use in Inflammatory Bowel Disease—UpToDate. Available online: https://www.uptodate.com/contents/overview-of-azathioprine-and-mercaptopurine-use-in-inflammatory-bowel-disease#H3912407284 (accessed on 20 January 2024).
- Fathollahi, A.; Aslani, S.; Mostafaei, S.; Rezaei, N.; Mahmoudi, M. The Role of Killer-Cell Immunoglobulin-like Receptor (KIR) Genes in Susceptibility to Inflammatory Bowel Disease: Systematic Review and Meta-Analysis. Inflamm. Res. 2018, 67, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin in Aging and Disease—Multiple Consequences of Reduced Secretion, Options and Limits of Treatment. Aging Dis. 2011, 3, 194–225. [Google Scholar] [PubMed]
- Miller, S.C.; Pandi, P.S.R.; Esquifino, A.I.; Cardinali, D.P.; Maestroni, G.J.M. The Role of Melatonin in Immuno-Enhancement: Potential Application in Cancer. Int. J. Exp. Pathol. 2006, 87, 81–87. [Google Scholar] [CrossRef]
- Li, M.; Hao, B.; Zhang, M.; Reiter, R.J.; Lin, S.; Zheng, T.; Chen, X.; Ren, Y.; Yue, L.; Abay, B.; et al. Melatonin Enhances Radiofrequency-Induced NK Antitumor Immunity, Causing Cancer Metabolism Reprogramming and Inhibition of Multiple Pulmonary Tumor Development. Signal Transduct. Target. Ther. 2021, 6, 330. [Google Scholar] [CrossRef]
- Coppin, E.; Florentin, J.; Vasamsetti, S.B.; Arunkumar, A.; Sembrat, J.; Rojas, M.; Dutta, P. Splenic Hematopoietic Stem Cells Display a Pre-Activated Phenotype. Immunol. Cell Biol. 2018, 96, 772–784. [Google Scholar] [CrossRef] [PubMed]
- Berisha, A.; Shutkind, K.; Borniger, J.C. Sleep Disruption and Cancer: Chicken or the Egg? Front. Neurosci. 2022, 16, 856235. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Esquifino, A.I.; Perumal, S.P.; Miller, S.C. Melatonin, Immune Function and Aging. Immun. Ageing 2005, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Nedkova-Hristova, V.; Vélez-Santamaría, V.; Casasnovas, C. Myasthenia Gravis Exacerbation after Melatonin Administration: Case Series from a Tertiary Referral Centre. BMC Neurol. 2020, 20, 403. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M. Impact of Melatonin on Immunity: A Review. Open Med. 2013, 8, 369–376. [Google Scholar] [CrossRef]
- Liu, H.; Xu, L.; Wei, J.-E.; Xie, M.-R.; Wang, S.-E.; Zhou, R.-X. Role of CD4+ CD25+ Regulatory T Cells in Melatonin-Mediated Inhibition of Murine Gastric Cancer Cell Growth in Vivo and in Vitro. Anat. Rec. 2011, 294, 781–788. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Alonso-González, C.; González-González, A.; Menéndez-Menéndez, J.; Cos, S.; Martínez-Campa, C. Melatonin as an Adjuvant to Antiangiogenic Cancer Treatments. Cancers 2021, 13, 3263. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.-G.; Tang, S.-T.; Tseng, H.-P.; Wu, K.K. Melatonin Suppresses Macrophage Cyclooxygenase-2 and Inducible Nitric Oxide Synthase Expression by Inhibiting P52 Acetylation and Binding. Blood 2006, 108, 518–524. [Google Scholar] [CrossRef]
- Talib, W.H.; Alsayed, A.R.; Abuawad, A.; Daoud, S.; Mahmod, A.I. Melatonin in Cancer Treatment: Current Knowledge and Future Opportunities. Molecules 2021, 26, 2506. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Melatonin Receptor | MT1 | MT2 | MT3 |
|---|---|---|---|
| Previously named | Mel1A | Mel1B | Mel1C |
| Coupled Receptors/Binding site | G protein-coupled Receptors | Quinone reductase 2 (QR2) | |
| Function/mechanism |
|
|
|
| Abundance | Ileum, colon and mucosa |
| Ileum and Colon |
| Species | Influence of Gut Bacteria on Melatonin and Melatonin Precursors | References |
|---|---|---|
| Helicobacter pylori (H.Pylori) |
| [38] |
| Lactobacillus rhamnosus |
| [92] |
| Clostredium sporogenes Ruminococcus gnavus |
| [93] |
| E.coli Bacteroides |
| [94] |
| E.coli or Lactobacillus rhamnosus |
| [95] |
| Roseburia hominis |
| [96] |
| Metabolites | Effect | References |
|---|---|---|
| SCFA |
| [37] |
| TMAO |
| [97] |
| Aeromonas Veronii LPS |
| [98] |
| Bile acids |
| [99] |
| Study | Effect of Melatonin |
|---|---|
| Gao et al., [100] | Contributes to the activity of 5-FU in inhibiting CC cell migration |
| Liu et al., [101] | Inhibits RKO CC migration by attenuating ROCK expression (through p38/MAPK) |
| Zou et al., [102] | Reduces CC cell proliferation by inactivating p38/MAPK |
| Anisimov et al., [103] | Reduces the depth of invasion of CC in vivo |
| Park et al., [104] | Reduces the expression of VEGF by destabilizing HIF-1α and by acting on HIF-1α activity in CC cells. |
| Léon et al., [105] | Inhibits angiogenesis by blocking ET-1 release from CC cells |
| Palidarova et al., [106] and Srinivasan et al., [107] | Affects the immunity by targeting specific cells (such as TH cell) to produce Interleukins to attenuate CC development |
| Kossoy et al., [108] | Acts on the lymphatic system of the host to exert its anti-carcinogenic effect |
| Farriol et al., [109] | Antiproliferative activity by decreasing cell growth in non-hormonal depedent colon cells |
| Winczyk et al., [110] | Oncostatic effect through MT2 receptors acting on RZR/ROR α nuclear receptors. |
| Hong et al., [111] | Induces colon cell death programs and stops mitotic activity through G1-phase arrest |
| Wei et al., [112] | Contributes to apoptosis in CC through HDAC4 nuclear import |
| Chovancova et al., [113] | Induces apoptosis in CC cells through Na/Ca exchanger type 1 and IP3 type 1 receptors/ |
| Yun et al., [114] | Induces mitochondria-mediated cell apoptosis in CC cells through PrPC pathway |
| Kannen et al., [115] | Limits CC progression by controlling malignant lesions in CC through CD68+ and CD133+ cluster cells |
| Immune Factors/Cells | Effect of IBD | Effect of Melatonin |
|---|---|---|
| Th17 | - | Inhibition of differentiation |
| T-reg | - | Increase production |
| IL-17 | Increase in number | Remarkable reduction in number |
| IL-23 | Increase in number | Mild reduction in number |
| IL-10 | Reduction in number | Increase production |
| IL-6 | Increase in number | Significant reduction in number |
| TNF-alpha | Increase in number | Significant decrease in number |
| CD4+T | Increased in number, main drivers of inflammation | Induces in-vitro proliferation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurjus, A.; El Masri, J.; Ghazi, M.; El Ayoubi, L.M.; Soueid, L.; Gerges Geagea, A.; Jurjus, R. Mechanism of Action of Melatonin as a Potential Adjuvant Therapy in Inflammatory Bowel Disease and Colorectal Cancer. Nutrients 2024, 16, 1236. https://doi.org/10.3390/nu16081236
Jurjus A, El Masri J, Ghazi M, El Ayoubi LM, Soueid L, Gerges Geagea A, Jurjus R. Mechanism of Action of Melatonin as a Potential Adjuvant Therapy in Inflammatory Bowel Disease and Colorectal Cancer. Nutrients. 2024; 16(8):1236. https://doi.org/10.3390/nu16081236
Chicago/Turabian StyleJurjus, Abdo, Jad El Masri, Maya Ghazi, Lemir Majed El Ayoubi, Lara Soueid, Alice Gerges Geagea, and Rosalyn Jurjus. 2024. "Mechanism of Action of Melatonin as a Potential Adjuvant Therapy in Inflammatory Bowel Disease and Colorectal Cancer" Nutrients 16, no. 8: 1236. https://doi.org/10.3390/nu16081236