
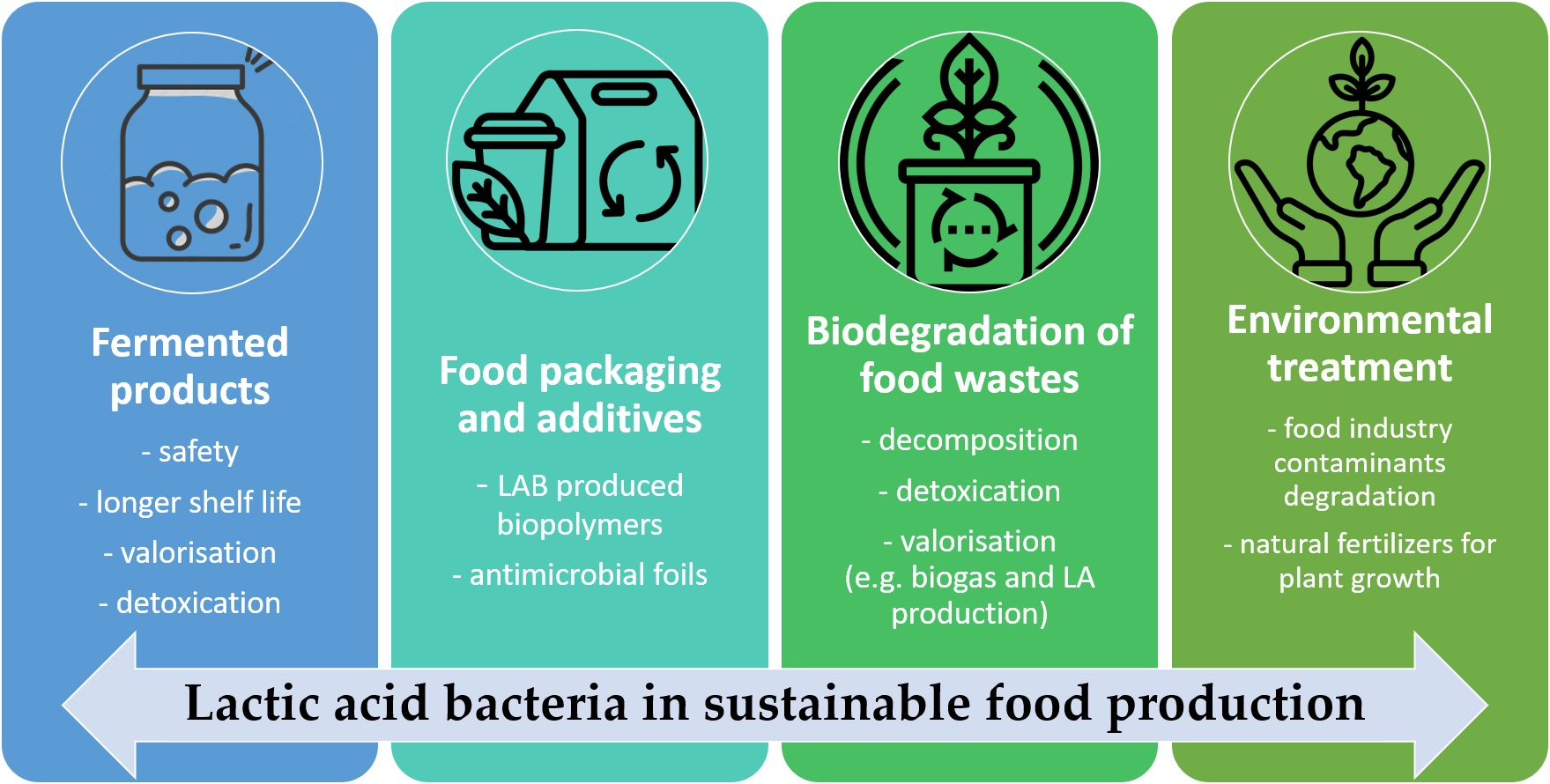
Lactic Acid Bacteria in Sustainable Food Production


Department of Biotechnology, Microbiology and Human Nutrition, Faculty of Food Science and Biotechnology, University of Life Science in Lublin, 8 Skromna Street, 20-704 Lublin, Poland
*
Author to whom correspondence should be addressed.
Sustainability 2024, 16(8), 3362; https://doi.org/10.3390/su16083362
Submission received: 27 February 2024
/
Revised: 12 April 2024
/
Accepted: 15 April 2024
/
Published: 17 April 2024
(This article belongs to the Special Issue Food, Supply Chains, and Sustainable Development)
Abstract
:The main tenets of the sustainable food production model are to reduce the adverse environmental impacts of production and to use available resources more efficiently. The sustainable food production model allows companies to adapt their strategies to current challenges and requirements while maintaining long-term production stability and competitiveness. To ensure that sustainable food chain participants implement appropriate practices, research is being conducted to develop new solutions. Among the important issues that are of great interest to researchers is the use of lactic acid bacteria (LAB). These bacteria play a pivotal role in sustainable food production, encompassing environmental, economic, and social aspects. The following article highlights recent innovations and advancements in LAB applications, contributing to enhanced efficiency and sustainable development of food products. By fermenting food, LAB effectively enhances food safety, prolong shelf life, and augment nutritional values, while simultaneously eliminating or outcompeting foodborne pathogens, thus preventing food poisoning. This article underscores these often-overlooked aspects of LAB, such as the critical role of fermented food in sustaining humanity during challenging times, by providing essential nutrients, and supporting health through its unique preservative and probiotic properties. It also points out the lesser-known applications of these microorganisms, including the degradation of organic waste or biogas and bioplastics production.

{kind=link}
1. Introduction
Sustainable food production is based on meeting three main assumptions: it is environmentally and climate friendly, economically viable and socially acceptable. The deployment of lactic acid bacteria in food production, food packaging and food waste management aligns with all of these assumptions [1]. In the context of escalating demands for sustainable alimentary production, lactic acid bacteria (LAB) serve as a bridge linking traditional food production methods, supporting the longevity and safety of consumable goods, with a modern approach. The use of this natural process, starting from spontaneous fermentation and the back-slopping method, was probably the first biotechnological thought [2].
Subsequently, the development of starter cultures has facilitated augmented consistency and safety, contributing to the improvement of taste, texture, and durability of this food. The name LAB suggests their most important feature, the production of lactic acid, which determines the transition of LAB from an insignificant part of the microorganism population in the raw product to dominating in the product with completely altered attributes. The activity of microorganisms not only refers to food preservation and ensuring its safety for consumption by improving nutritional value and bioavailability but also minimizes waste and environmental impact [3]. The modern approach to fermentation using LAB highlights their key role in creating more sustainable food production systems, firstly by reducing the negative ecological impact associated with traditional food production methods, and secondly, by providing nutritionally superior products [4]. Innovative applications such as the use of microalgal fermentation to expand the range of substrates with incredible nutritional value; proteins, lipids, pigments, carotenoids, vitamins, polyunsaturated fatty acids, and steroids, all of which can be further augmented through LAB fermentation, underscoring the innovation in leveraging LAB for expanded nutritional and environmental benefits [5]. LAB can be employed in food waste treatment processes to enhance the removal of organic pollutants and nutrients. Their ability to ferment and metabolize various compounds makes them valuable in the treatment of food waste and to convert them into valuable products such as lactic acid, animal feed or soil conditioner. In addition to their role in managing pollution through degradation of some compounds in the environment, microorganisms serve as the biosynthetic engines for the production of bioplastics [6]. Moreover, they can be exploited for biogas production, also based on waste materials as substrates.
There is a lack of previous papers in the literature that summarise all these aspects of the use of lactic acid bacteria in sustainable food production. Hence, this study aims to provide a broad overview of their versatile application in food production. This review highlights mankind’s recognition centuries ago of the potential of LAB in food preservation right through to recycling and valorisation of food waste, making it possible to obtain food additives, ingredients and packaging or certain other products, including lactic acid and biogas.
2. The Role of Lactic Acid Bacteria in Food Fermentation
If LAB were to vanish suddenly, it would pose significant challenges to understanding the profound effects on various aspects of our daily lives. This includes impacts on food production and its related economic sectors, human and animal health, ecosystems, the environment, and even the realms of science and biotechnology. Historically, preventing food spoilage has been a key function of LAB [7]. These microorganisms, which represent a small fraction of the bacterial populations plant-based products, can dominate their ecological niche when provided with optimal conditions such as appropriate osmotic environments, absence of oxygen, and moderate temperatures, thereby initiating fermentation. This task of fermentation, sometimes occurring spontaneously and at other times directed by human intervention, has been crucial in sustaining ecological balance and ensuring the safety of our food supply [8]. It is natural to conclude that spontaneous fermentation, characterized by reliance on the native bacterial flora of raw materials and the surrounding environment without the intervention of added cultures, was historically the first fermentation method.
2.1. Traditional Applications of LAB in Food Fermentation
What conditions must be met for spontaneous fermentation to occur? Depending on the raw material, bacteria such as LAB, yeasts, or even molds necessary to start fermentation should be present on their surface. For the growth and development of microorganisms, the appropriate water activity is necessary, about 0.95 or higher [9]. Of course, the optimal temperature depends on the process, for example, for black olives fermentation it is 25 °C [10], in the case of yogurt we have a temperature range of 37–46 °C [11], 30–35 °C for sourdough LAB fermentation [12]. The initial pH also matters, a range from 4.5–6.5 is considered optimal for LAB at the start of lactic acid fermentation. Often overlooked is the issue that the raw material must contain fermentable sugars which would be substrates in the formation of lactic acid and other metabolites. A distinct physico-chemical property is the incorporation of salt, indicative of human activity’s role in the fermentation process. Typical addition of salt is a concentration of 2–5% in relation to the weight of vegetables, or it may be increased to 8% if whole vegetables are fermented, e.g., cucumbers, where as a result of the diffusion of water from the vegetable to the brine, the salt concentration will decrease. Salt is added to vegetables to create an environment that favors the growth of LAB by inhibiting undesirable microorganisms. The concentration of salt significantly affects the growth dynamics of LAB, increasing their quantity, which in practice improves the quality of the final fermented product [13].
Fermentation is a natural process used by humans for at least 6000 years [14], carried out in individual households to survive unfavorable periods without access to fresh plant-based produce [15]. At some point, the possibility of controlling this process through back-sloping was recognized. This technique is characterized by a straightforward methodology that entails the incorporation of a portion of previously fermented material into a new batch of raw material specifically prepared for the process of fermentation. This approach has several advantages, it quickly induces the growth of appropriate microorganisms. It has been demonstrated that, in comparison with spontaneous fermentation, the quantity of LAB was significantly higher, by as much as an entire logarithmic cycle [16]. The expedited development of the appropriate microorganisms considerably enhances the probability of inhibiting the growth of deleterious spoilage bacteria, owing to the fact that the added bacteria are already well-adapted to the substrate of the fermented product and are capable of producing inhibitory lactic acid at a more rapid pace. To this day, products made using the back-slopping method are appreciated for their tradition, such as sourdough bread, beers, and cheeses [14]. The acceleration of the fermentation process is not the only advantage; it also includes the enhancement of the nutritional value of the product [17]. Concurrently, the practice of back-sloping disrupts the balance of the autochthonous microbiota, which then begins the struggle for dominance and adapts to the individual compounds from which they derive energy. The introduced strains have had the opportunity to undergo the process many times, making them “super strains” for that specific environment. Nevertheless, the method is not perfect because, just as it allows for the introduction of beneficial strains, it also increases the risk of contamination resulting in a lack of repeatability [18]. The succession of various species and even genera of LAB during the fermentation process is well-known, thus raising the question of whether strains adapted to the final stages of fermentation are indeed suitable for ensuring the desired quality and consistency of the fermented product.
The bridge connecting traditional fermentations with a more modern approach is the use of starter cultures. Tailored strains are used to obtain a repeatable final effect. Their application enhances product consistency, predictability, safety by inhibiting harmful microorganisms, accelerates fermentation, and improves the flavor, texture, and shelf life of fermented products [19,20]. Starter cultures are predominantly used in the dairy industry, playing a pivotal role in directing the fermentation process to achieve desired outcomes in dairy products. There are few products that are still produced through spontaneous fermentation in this industry, but they mainly occur on a small scale, including kefir from kefir grains, traditional koumiss fermented mare’s milk drink, and shubat obtained from camel’s milk [21]. These specific microorganisms are intentionally introduced to convert lactose into lactic acid, a critical reaction facilitating milk protein coagulation, essential for manufacturing products like cheese and yogurt [22]. Beyond acidification, these cultures significantly contribute to the development of distinctive aromas, flavors, textures, and nutritional profiles by producing beneficial antimicrobial substances [23]. The selection of such a strain is a continuous search, where the bacterium should exhibit a range of characteristics beyond the obvious production of the desired metabolite. It is essential that the bacterium not only produces the desired metabolite but is also examined for resistance to oxidative stress, lag phase duration, resistance to low pH, and optimal temperature conditions [24]. The starter strain must be adapted to variable, often extreme physicochemical environmental conditions at different stages of fermentation—from milk to yogurt or cheese, from raw to sauerkraut cabbage. To explore the most commonly utilized LAB strains in the production of yogurt, specifically Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus, it’s essential to highlight their co-cultivation synergy. This relationship significantly contributes to the fermentation process, enhancing yogurt’s flavor and texture [25]. Lactococcus lactis and Limosilactobacillus fermentum are identified as key contributors to the flavor and aroma profiles of cheeses [26,27]. In the case of sauerkraut, Leuconostoc mesenteroides is predominantly used in the initial stages of fermentation due to its rapid growth and lactic acid production, which quickly diminishes in favor of others genre of bacteria [28]. Furthermore, the highly versatile Lactiplantibacillus plantarum, present in almost every environment occupied by LAB and characteristic of the final stages of plant fermentation. The same strains are also prominent in kimchi production, along with Weissella koreensis—a strain dominant in all samples of kimchi fermented at low temperatures, suggesting its significant role in the fermentation process [29]. As a functional starter culture in fermented meat products, the species name Latilactobacillus sakei often emerges, predominantly in the domain of fermented sausage production. This microorganism is acclaimed for its remarkable adaptability to the meat environment, valued for its production of enzymes, which are responsible for developing desired aromatic compounds during the maturation of fermented meat products. The proteolytic breakdown of proteins during the fermentation process of traditional meat products allows for the release of bioactive peptides, including valuable antioxidant peptides and angiotensin-I-converting enzyme inhibitors [30,31]. During the fermentation of dry sausages, the succession of microorganisms depends on maturation; in addition to the Lactobacillus genus, Leuconostoc, Lactococcus, Pediococcus, Weissella, and Enterococcus also appear [32]. The methodology applied in the crafting of dry fermented meat products typically involves mild to no heat exposure, thus optimizing the viability of LAB under such conditions. Currently, much research focuses on adding probiotic strains. For example, the quality of Italian salami sausage has been shown to improve with the addition of Lacticaseibacillus casei and FOS [33]. The LAB metabolism is extremely important in obtaining taste and color, Tian et al. have shown that strains of Lactobacillus helveticus contributed to the formation of 72 flavor compounds, and the amount of alkanes, ketones, acids increased [34]. Furthermore, strategic manipulation of various L. sakei strains causes changes not only in the taste and aroma of fermented sausages but also significantly influences the formation of color [35]. A notable instance includes findings by Ameer et al. (2021) proved that the use of the L. sakei S3 strain improves the values of red color [36].
Therefore, these are only some of the LAB strains present in widely-consumed fermented products. Remarkably, each nation contributes in food sciences by its distinct tradition and modern innovations to these processes. This diversity in fermentation practices underscores the profound and intrinsic value of fermentation traditions, illustrating their broad application across various cultural and technological contexts, thereby affirming their consistently key role in the development of societies over the centuries.
2.2. The Role of LAB in Enhancing the Shelf Life and Safety of Fermented Food Products
The main factor contributing to the popularity of the widespread use of the fermentation process is its ability to extend the shelf life of food products. This is achieved through the metabolism of lactic acid bacteria, which, as the name suggests, produce lactic acid which lowers the pH of the environment which effectively inhibits the growth of spoilage-causing microorganisms, thereby extending the shelf life of the product [37]. The impact of LAB on the shelf life and quality of stored foods lies in their primary metabolism. The metabolic activity of LAB allows them to utilize sugars such as lactose, fructose, glucose and galactose into significant amounts of lactic acid along with secondary metabolites. Lactic fermentation bacteria, depending on the amount of primary metabolites produced, can be divided into homofermentative and heterofermentative bacteria. Homofermentative bacteria in the Embden-Meyerhof-Parnas pathway convert one molecule of glucose into two molecules of lactic acid, a more energy-efficient process, as it produces 2 molecules of ATP from one molecule of glucose. This metabolic efficiency is crucial to their ecological dominance, as the rapid drop in pH caused by lactic acid synthesis is crucial during the initial stages of fermentation, effectively suppressing the competitive and abundant native food surface microflora [38].
Conversely, heterofermentative LAB metabolize glucose primarily via the pentose phosphate pathway. This route yields lactic acid along with diverse metabolites like carbon dioxide, ethanol, or acetic acid, but only one ATP molecule per glucose molecule—indicating a lower energy efficiency compared to homofermentative bacteria.
However, the spectrum of metabolic outputs enriches the sensory qualities of food, offering advantages for specific fermentation applications [39]. The longevity and microbiological safety of fermented food are ensured by LAB not only through organic acids by heterofermentative pathway LAB produce a range of antimicrobial metabolites such as carbon dioxide, ethanol, diacetyl, hydrogen peroxide, acetaldehyde, and acetoin [7]. Ethanol damages the cell membranes of microorganisms, carbon dioxide, by dissolving, creates an anaerobic environment inhibiting the growth of aerobic spoilage bacteria, while hydrogen peroxide acts as a preservative in fermented products, impairing microbial cell membranes and DNA integrity [40].
The antimicrobial effect of lactic acid against an unfavorable part of the bacterial community, is triggered precisely by LAB activity or naturally occurring organic acids. The mechanism involves disrupting the metabolism and cell membrane integrity of pathogens [41]. Undissociated, lipophilic forms of weekly organic acids penetrate microbial cells by simple diffusion, disrupting the internal pH balance. Once inside the cell, at the neutral pH of the cytosol, it dissociates, leading to the accumulation of acid anions in the cytosol. Inhibiting growth through mechanisms such as enzyme denaturation, disruption of proton motive force and ATP depletion and accumulation of reactive oxygen species (ROS) [42]. The yeast Saccharomyces cerevisiae, which often co-ferments foods alongside LAB, adapts to this acid by activating the H+-ATPase Pma1 pump to pump out protons, which helps restore pH balance [43].
Even LAB are sensitive to high concentrations of lactic acid. One of the researchers explaining why LAB die from their own metabolite is Rallu et al. [44]. On the acid-resistant L. lactis strain MG1363, they examined the molecular basis of protection by the acid. The L. lactis strain has several defense mechanisms, for example, (p)ppGpp intracellular phosphate and guanine nucleotide pools signal indirection of the stress response. In one study, Matsumoto, et al. [45] investigated the acid tolerance of various strains of bifidobacteria by testing their survival rates under acidic conditions (pH 2–5). They found that Bifidobacterium lactis and Bifidobacterium animalis strains showed higher acid tolerance compared to other strains, with significant survival rates even after exposure to pH 3–5 for 3 h. It was hypothesized that this higher acid tolerance is related to the activity of H+-ATPase, an enzyme that helps maintain intracellular pH by expelling protons from the cell. The H+-ATPase activity of the more tolerant strains was higher at pH 4 than at pH 5, suggesting a proportional response to stress [45]. In a study conducted by Lowe al. [46], three LAB strains selected for their lactic acid generating efficiency or enzymatic activity during the malting process were used to evaluate their impact on malt and wort quality. Comparisons were made between wort obtained using LAB and malt wort with the addition of lactic acid. The results showed that the use of LAB improved the levels of β-glucanase in malt, although reduced malt crispness was observed. However, lactic acid is not enough to stop food spoilage microflora retroactive use of this organic acid has been noted during fermentation of sourdough, pickling [47] and pickled cucumbers [48]. Besides lactic acid, LAB are known to produce acetic acid, malic acid. These acids can beneficially affect the shelf life of fermented food and inhibit the growth of yeasts or molds. When added to the fermentation of cassava foliage along with the strain L. plantarum, they quickly lowered the pH and increased the lactic acid content [49]. Citric acids are a valuable component in the fermentation niche’s metabolic cycle, as they can be utilized by other microorganisms as a carbon source for energy acquisition.
Acetic acid is formed from ethanol or acetaldehyde and has a wide range of uses beyond food preservation, acting as an emulsifier or flavor enhancer. It is commonly known as an acid with strong antibacterial properties [50]. Studies have shown that acetic acid is effective against L. monocytogenes at equal pH values compared to lactic, citric, malic, and hydrochloric acids [51]. Analogous findings by Al-Rousan et al. [52] found that acetic acid substantially inhibited the proliferation of Salmonella and E. coli more effectively than citric acid, even at a concentration of 0.4%. Unlike the production of other organic acids, lactic acid bacteria produce acetic acid in relatively small amounts. As the name implies, when it comes to acetic acid production, acetic acid bacteria (AAB) far outperform LAB in the production of acetic acid, as the fermentation environment is a melting pot of different species, a pattern of co-operation between these groups of bacteria has been observed. In a study conducted by Xia et al. [53] on the fermentation of aged Shanxi vinegar, antagonistic interactions were noted between the two groups of microoganisms. The acetic acid bacterium Acetobacter pasteurianus, which was the predominant strain in this group, hindered the growth of L. helveticus only in the fermentation medium and not in vitro.
When discussing fermentation, it’s difficult to attribute significance to a single organic acid, as it typically involves a blend of several acids. Ozcelik et al. [54] showed that with various fish additives to MRS broth, Pediococcus acidilactici produced a significantly larger amount of lactic acid in tilapia broth than in MRS, reaching levels of 1211.97 mg/L compared to 305.09 mg/L in MRS. The same bacteria also produced a high concentration of propionic acid (3747.28 mg/L) in sea bass broth. When grown in trout broth, L. plantarum exhibited low acetic acid production levels (0.62 mg/L). In contrast, Streptococcus thermophilus showed no production of lactic, acetic, or butyric acids in tilapia broth. High concentrations of butyric acid (>400 mg/L) were observed with Lactobacillus delbrueckii subsp. lactis and Lactobacillus acidophilus. In a related finding, Al-Rousan et al. [52] reported that acetic acid was more effective at inhibiting the growth of Salmonella and E. coli than citric acid, even at a low concentration of 0.4%.
Compounds contributing to the enhanced microbiological stability of fermented products can be reuterin and reutericyclin, synthesized by Limosilactobacillus reuteri strains. It is a compound with a broad spectrum of activity against Gram-positive and negative bacteria, fungi, and protozoa [40]. The mechanism of action is not fully understood; it may involve inhibiting DNA synthesis in cells, oxidative damage through the depletion of free sulfhydryl groups.
These compounds exert their bacteriostatic effects through the alteration of thiol groups in proteins, preventing the growth of bacteria, including Escherichia coli and Clostridium difficile [55,56]. Additionally, combining reuterin with diacetyl shows increased antimicrobial activity against foodborne pathogens, especially under acidic conditions, notably against Listeria monocytogenes [57]. Reuterin biosynthesis occurs under conditions of glucose deficiency, with glycerol acting as the precursor substrate. This biochemical pathway was exploited by incorporating L. reuteri into cheese, which by secreting reuterin, significantly reduced the number of Clostridium tyrobutyricum spores [58]. A similar addition of starter and substrate was used in preserving ground beef, where reuterin killed the E. coli O157:H7 strain [59].
The peak of antimicrobial effects observed in the environment of fermented foods due to LAB is attributed to their production of bacteriocins, which are peptides of low molecular weight. Their production indicates the evolutionary success of the producing strains, significantly enhancing the quality and safety of food [60]. In a study conducted by Grosu-Tudor [61], it was discovered that only about 20% of analyzed strains demonstrated antimicrobial activity, mainly against Bacillus strains and other LAB, which in most cases could be attributed to the production of organic acids. Only for six strains was it determined that antibacterial activity resulted from bacteriocin production was identified in merely 2% of the strains. These were 6 LAB strains isolated from Romanian traditional fermented vegetables and grains, including bors, a sourdough derived from wheat bran and maize flour. These specific strains manifested inhibitory actions against L. delbrueckii subsp. bulgaricus LMG 6901T, B. cereus CBAB, B. subtilis ATCC6633, Bacillus sp., L. monocytogenes, and Staphylococcus aureus, demonstrating the undiminished potential of bacteriocins as core antimicrobial agents in fermented food systems. Devi et al. evaluated the ability of LAB cultures (Pediococcus acidilactici, Enterococcus faecium, Lactiplantibacillus plantarum) to produce a bacteriocin similar to PA-1 pediocin during fermentation of soy milk. Together, these isolates were capable of producing bacteriocins and fermenting soy milk, showing longevity even after 15 days and suppression activity against L. monocytogenes. Confirming the use of such a consortium as a starter culture to advance shelf-life [62]. Numerous studies have focused on enhancing bacteriocin production through modifications in growth medium composition and alteration in physiological conditions, in order to enhance bacteriocin yields. An additional approach has involved the introduction of extra stimuli. For L. plantarum NC8, a starter culture used in Spanish-style green olive fermentation, and Leuconostoc citreum GJ7, a kimchi isolate, this is achieved by adding specific adjunct cultures that stimulate bacteriocin production [63,64]. When it comes to meats, it’s also worth noting that the presence of LAB strains in the fermentation process is crucial for enhancing the safety of the product at every stage until the end phases of storage. The L. sakei genus, along with others such as Latilactobacillus curvatus and Pediococcus pentosaceus, not solely facilitates the enhancement of the sensory profile but also acts as a protection against spoilage and pathogenic microorganisms, thus elevating the product’s safety and prolonging its shelf life. The esteemed L. sakei species, which is highly regarded in meat production, produces a range of bacteriocins Sakacin A, B, G, K, M, T, P, Q (and more), with a diverse spectrum of action against ‘spoilage-type’ microorganisms like Listeria, as well as Streptococcus, Lactococcus [30]. In 2021, Abitayeva et al. described a novel bacteriocin-like peptide, Sak-59, derived from the aforementioned microorganism, strain B-RKM 0559 originating from traditional Kazakh horse meat Kazy [65]. From a traditional Chinese product, Steamed Roast Duck called Nanjing, a class III bacteriocin Y19-2 was identified, exhibiting not only a broad antimicrobial spectrum of action, including the previously mentioned microorganisms but also efficacy against E. coli. It has been shown to be heat resistant by retaining 86% of its activity post-exposure to 121 °C for 30 min [66]. The antimicrobial properties of L. sakei isolated from Italian salami were “encapsulated” by creating a bioplastic film utilizing the supernatant of this strain containing bacteriocins, which allowed for a 3 log cycle increase in microbiological stability for the tested group of microorganisms [67]. The pragmatic relevance of bacteriocins in food safety and their application includes the use of LAB starter cultures proven to produce bacteriocins, using bacteriocins as food additives, and in films creating functional packaging extending the shelf life of the packaged food. Bacteriocins as peptides can be sensitive to pH, temperature, storage conditions, and even the composition of the food matrix wherein they reside. Nonetheless, this approach harbors several constraints beyond legal regulations, including the specific spectrum of bacteriocin activity, which limits their broader applicability, and the potential for the development of resistance to bacteriocins cannot be excluded.
2.3. Enhancing Nutritional Value of Food through LAB Fermentation
Improving the nutritional profile of food through lactic acid bacteria fermentation is pivotal for sustainable development, combating nutritional deficiencies, and influencing public health, for instance, by relieving healthcare systems. This improvement practically impacts every aspect of life. However, it’s noteworthy that both fermented products and fresh vegetables or fruits serve as appropriate sources of nutrients and bioactive components, but fermented food boasts an extended shelf life [68].
Enhancing the nutritional value of fermented food is achievable due to the specific metabolic characteristics of individual species, often genetically linked to a particular ecological niche. The dynamic changes in metabolite quantities, driven by microorganism activity in interaction with the raw material matrix, indicate exploring various pathways during fermentation to achieve evolutionary success, which we can leverage in many fields [69]. Firstly, by metabolizing a range of complex compounds, LAB release a series of metabolites that are easier for higher organisms to assimilate. We often overlook that we naturally obtain lactose-free products through the breakdown of this compound in fermented dairy [70]. It’s worth expanding on the topic that fermentation reduces carbohydrate amounts, which are converted into organic acids. This process is particularly desired in sourdough, where the pH reduction favors the hydrolysis of nutrients and increases the availability of vitamins and minerals in bread [71].
LAB’s own metabolism can contribute to many aspects of increasing the bioavailability and activity of bioactive food components, such as an increase in polyphenols, through processes like phytic acid degradation, which in turn affects better mineral assimilation. LAB do not directly increase the number of polyphenols but affect their availability and biological activity, through enzyme secretion and the cooperation of many strains. In a study conducted by Ye and colleagues [72], the impact of LAB fermentation on the chemical composition of broccoli puree (previously autoclaved) was examined. Using seven LAB isolates obtained directly from broccoli, the authors observed a significant increase in the total amount of glucosinolates and ten major polyphenols from values of 55–359 μg/g to 903–3105 μg/g of dry weight. This significant increase suggests the potential application of LAB fermentation to enrich broccoli in glucosinolates and polyphenols, further raising their value as dietary components. A similar improvement was noticed in another study where Ricci and his team [73] assessed the impact of ten Lactobacillus strains on the polyphenol metabolism in black currant juice. After adding LAB, particularly Lacticaseibacillus rhamnosus and L. plantarum, a significant increase in total polyphenol concentration was found, with quercetin-3-O-rutinoside as the most abundant compound. Kwaw and colleagues [74] examined berry juice, where an increase in polyphenol quantity and a change in product color were also noted.
Phytic acid is an antinutrient found in many plant food sources, such as grains and legumes, that can bind minerals (e.g., iron, calcium, magnesium) and limit their bioavailability. During fermentation, LAB can produce phytase enzymes that degrade phytic acid, reducing its concentration in the final product. This process is particularly important in the context of plant protein sources and grains, which often contain high levels of phytates; enzymatic hydrolysis of phytates leads to the release of bound minerals. Mohammadi-Kouchesfahani et al. isolated lactic acid bacteria from sourdough produced by the back-sloping method from whole grain flour and fava beans, to select strains showing phytase activity. These were Weissella confusa mk.zh95 and Pediococcus pentosaceus, where the former strain showed the highest phytase activity. This indicates an interesting dependency on the occurrence of microorganisms with such activity in bread, precisely because of the higher concentration of phytic acid [75]. Rosa-Sibakov et al. [76] studied the reduction of phytic acid in faba bean flour, using enzymatic reaction and fermentation with Lactiplantibacillus plantarum VTT E-78076, achieving an 89% reduction, additionally improving protein digestibility and solubility. Other research successfully reduced phytic acid levels in wheat and sorghum [77], or in sorghum-ogi porridge [78], links to phytase-producing L. plantarum. Fermentation with this strain often appears as a promising way to reduce phytic acid content.
One of the key aspects is LAB’s ability to modify the chemical and physical properties of foods, which may facilitate the release of minerals and their absorption by the human body. For example, studies conducted by Scheers et al. suggest that increased iron bioavailability from vegetables (rich and poor in phytic acid) fermented by LAB likely results from the increase of trivalent iron species (Fe3+) [79]. This study did not observe an improvement in zinc bioavailability, unlike the case studied by Zhang et al. Fermented pea protein concentrates featured improved bioavailability of iron and zinc salts [80].
LAB’s metabolic processes sometimes lead to the de novo synthesis of vitamins. LeBlanc et al. (2013) appreciate this fact because these compounds are exogenous to humans. Fermented milks with high levels of B-group vitamins, such as folates and riboflavin, can be produced thanks to LAB-promoted and potentially bifidobacteria-promoted biosynthesis. Additionally, some LAB strains produce the complex vitamin cobalamin (vitamin B12). It’s important to emphasize that vitamins are usually secondary metabolites; beyond having the appropriate genes for vitamin synthesis, optimal conditions like substrate availability or dysbiosis, the presence of precursors, and suitable physicochemical conditions must occur. Some strains may require additional vitamin sources in the medium to transform them into more bioavailable forms or synthesize them in larger quantities. LAB promote the biosynthesis of high levels of B-group vitamins, such as folates and riboflavin, in fermented milks. LeBlanc et al. showed that the consumption of certain fermented dairy products could cover over 10% of the folate (vitamin B9) requirement for pregnant women [81].
There are many aspects from the breakdown of complex compounds, releasing and improving the profile of nutrients to “supplementation” in the form of vitamins delivered by fermenting LAB. Essentially, each relationship between a microbial consortium and the raw product yields different and unique results, a synergistic mixture of compounds in the product carrying a beneficial impact on human health. In a study conducted by Kiczorowski and others [68], the effect of fermentation on the nutritional value of several vegetables compared to unfermented ones was examined; carrots and peppers after fermentation had more vitamin A and carotene, all fermented vegetables had a lower level of heavy metals (lead, cadmium, and copper). They also indicated that the increase in nutritional value might be related to concentration, as the amount of water decreases after fermentation, except for fat and energy content [68]. Vitamin production makes strains potential starters, Rodrigo-Torres and others [82] isolated L. plantarum from a traditional fermented Argentinian drink, chicha, due to its ability to produce vitamin B2 (riboflavin) and vitamin B9 (folates). These strains also showed antimicrobial properties and antibiotic tolerance, making them promising candidates for designing functional foods. In a study by Kaprasob and others [83], the nutritional value of fermented tropical drink, cashew apple juice, was examined where an increase in vitamin C and phenolic compounds was observed. Meanwhile, strains Weissella cibaria and Leuconostoc citreum, obtained from sourdough, are producers of dextran and riboflavin (vitamin B2). These strains showed the potential to produce high levels of riboflavin and exopolysaccharide (EPS), which can be used for biofortification and as natural thickening agents desired in the baking industry [84]. Rheological and sensory properties have a huge impact on product attractiveness. Juvonen et al. [85] likely wanted to study the impact of fermentation by various LAB strains producing EPS on the rheological, chemical, and sensory properties of carrot puree. Fermentation with selected LAB strains modified the perceived texture of the liquid carrot model. The production of low-branched dextran by Weissella confusa and Leuconostoc lactis strains resulted in a dense texture along with a pleasant aroma and taste. This study indicates the potential of LAB fermentation as a natural method for replacing hydrocolloid additives in vegetable-containing products, not just carrots [85].
In conclusion, it’s worth mentioning that not only does fermentation itself improve the biochemical composition of food products, but the bacteria involved also play a significant role. Consuming fermented products delivers significant amounts of probiotics that improve the state of the gut microbiota, lower the risk of cardiovascular diseases, and type 2 diabetes [86]. Finally, the prevailing concept of consuming the right amounts of probiotics and caring for maintenance of gastrointestinal microflora is notably widespread today. Despite the advancement of science, we continue to explore whether it is beneficial to enrich the gut microbiota through consistent intake of fermented foods. In 2020, Pasolli et al. published an expansive study wherein, upon the analysis of 9445 human fecal metagenomes, the prevalence and abundance of LAB were proven to be low, and the species identified in food only partially overlap with those in the intestines [87]. This could suggest a disadvantage associated with the consumption of fermented products, yet comparative genomic analyses substantiated that generally, the origin of intestinal strains predominantly originates from dietary sources, especially in Europe. What further proves the inseparable bond of consumed microorganisms with the gut microbiota is that a connection of four metagenome-assembled genomes, pertaining to the species Limosilactobacillus mucosae, typically isolated only from the intestines of mammals, was reconstructed from kombucha, kimchi, and sauerkraut [87]. This may argue in favor of consuming fermented products, especially since there is no unequivocal evidence of the impact of probiotic supplementation on the condition of the gut microbiota [88]. Although the evidence confirms the potential of probiotics, there are several limitations to the study methodology, such as the probiotic effects being strain-dependent, and the composition and variability of an individual’s gut microbiota vary significantly among individuals, which significantly complicates research due to the need to obtain vast metagenomic data.
2.4. LAB in Detoxification of Food Products
Since exposure to toxic substances through the diet is recognised as a significant health problem, various methods of minimising toxic content in food have been investigated, including the use of LAB [89]. LAB can be employed as a natural treatment to reduce the harmfulness of toxic substances produced during food processing, introduced from raw materials used in food production or as food additives. The mechanisms of interaction between LAB and chemical food contaminants are diverse and depend on the specific strain, type of contaminant and physicochemical conditions. It should be mentioned at the outset that many research attempting to confirm the effectiveness of microbial-based strategies tested in vitro and in vivo have been developed using aqueous solutions of toxic compounds, with fewer studies dedicated to contaminants delivered directly by the contaminated food matrix [90]. In the manufacture of food, the use of certain additives such as potassium nitrate and sodium nitrite can lead to the production of several toxic substances. These compounds are formed by the interaction of food ingredients during the application of certain food processing methods, such as frying, smoking, baking, pickling, fermentation or heating. The most widely known of these compounds are heterocyclic amines (HA), biogenic amines, N-nitrosamines (NA), acrylamide and polycyclic aromatic hydrocarbons (PAH). These compounds have been extensively studied due to their strong carcinogenic effects and widespread occurrence. Recent studies have shown that LAB can reduce the amount of harmful substances in food by producing enzymes (e.g., biogenic amine oxidase), thus decreasing the presence of toxic substances in food products. In addition, LAB may increase antioxidant capacity and the ability to suppress bacteria that cause food spoilage or produce harmful substances [89]. LAB can significantly lower the N-nitrosamine content of fermented meat products. Such an ability has been demonstrated for L. curvatus and Lactiplantibacillus pentosus, among others [91]. There are several methods by which LAB reduces NA content in food, including the direct reduction of NA content by metabolism or adsorption. NA removal was demonstrated for L. pentosus R3, in which reduced NAs were not detected in whole cell extracts, indicating that cellular metabolism may be the mode of their reduction. In Bifidobacterium longum bb536, NA metabolism also may be mediated by the intracellular enzymes [89]. Moreover, the ability to reduce NA up to 50% was observed for L. rhamnosus LOCK 0900, L. rhamnosus LOCK 0908, L. casei LOCK 0919, and Levilactobacillus brevis 0945. The results showed that the concentration of both culture supernatants and membrane extracts of these four strains can reduce the content of N-nitrosodimethylamine (NDMA), suggesting that the reduction occurs through adsorption or metabolism [92]. LAB can also indirectly reduce NA content in food through precursor reduction and by antioxidant effects. In a study by [91], NMDA was shown to be decreased due to the reduction of its precursors in kimchi after inoculation with strains of L. sakei, L. curvatus, L. brevis, Leuconostoc carnosum, Leuconostoc mesenteroides, L. plantarum and L. sakei [91]. Similar results were obtained by Liao et al. [93], who observed that L. plantarum significantly reduced the levels of NDMA and its precursors in the traditional Chinese fermented fish [93]. It has been reported that LABs also have the ability to reduce HA concentrations in foods, mainly through its adsorption [89]. This ability has been demonstrated for L. acidophilus IFO 13951 and Bifidobacterium bifidum IFO 14252, among others, and the main HA-binding component was identified to be a peptidoglycan contained in the cell wall [94]. Stidl et al. (2008) examined the capacity of eight LAB strains to reduce various HA in cooked meat and the highest HA removal capacity was observed for L. helveticus and Sterptococcus thermophilus [95]. In recent years, the possibility of employing LAB to reduce PAHs in food has attracted much interest because it is a natural method which is more acceptable to consumers than available chemical and physical methods [96]. An example of such an application is the use of LAB for the surface treatment of cold-smoked sausage before and after smoking, which can significantly reduce the PAH content of the sausage [97]. The study by Zhao et al. (2013) and Lo et al., (2004) shown that that the main mechanism by which LAB reduces PAHs and is physical adsorption and the main binding site for this group of compounds is a peptidoglycan on the cell wall [98,99]. Examples of LAB with proven ability to reduce benzo[a]pyrene, a member of the PHAs, are L. plantarum 121, L. pentosus ML32 and Bifidobacterium lactis Bb-12 [89]. LAB has also been shown to be a natural agent feasible for reducing the amount of acrylamide commonly present in food and highly toxic to humans. The mechanism through which LAB reduces acrylamide involves direct adsorption and reduced bioavailability of acrylamide through physical binding. Acrylamide binding is associated with teichoic acids present in the bacterial cell wall [100]. Zhang et al. (2017) demonstrated the ability of four LAB strains (L. casei ATCC393, L. plantarum 1.0065, L. acidophilus LADS1.0307 and S. thermophilus KLDS1.0316) to bind acrylamide [101]. They also observed that the highest carbohydrate content in the peptidoglycan structure was positively correlated with the ability of the peptidoglycan to bind acrylamide [102]. LAB can also be used to eliminate mycotoxins from food, including aflatoxin B1, which is a carcinogen produced by certain Aspergillus species. An example of a LAB capable of binding aflatoxin B1, which may be found in contaminated food, is L. rhamnosus strain GG [103].
2.5. Innovative LAB Fermentation Methods Supporting Sustainable Food Production
The development of fermentation methods is key to more sustainable food production, in the face of global challenges related to the wellbeing of the planet and the welfare of succeeding generations. The incorporation of contemporary LAB-based fermentation techniques and the further search for new ones are gaining new significance, offering the potential to create more sustainable food production systems. Such technologies have a dual purpose: firstly, to reduce the negative ecological impacts associated with conventional food production methods, and secondly to provide the most nutritionally valuable products possible. Central to this foundation for further development in this direction are “microbioreactors,” meaning LAB strains. The search for new strains continues, evidenced by the continuous emergence of new LAB species, such as Ligilactobacillus pabuli—a new lactic acid bacteria strain. The typical strain AF129T was isolated from alfalfa silage in Japan [104], from lychees in Taiwan Leuconostoc litchii was obtained [105], and the fructophilic bacteria with specific growth requirements, Philodulcilactobacillus myokoensis, from fermented vegetable extracts also in Japan [106]. In the last five years, we have acquired several promising starter cultures, for the production of feta cheese, Lacticaseibacillus paracasei K5 was chosen. Furthermore, a novel immobilization technique on lignin-free wheat bran was developed for this strain, enhancing the aromatic profile of the cheese and diminishing the likelihood of spoilage or pathogenic microorganisms [107]. Meanwhile, LAB capable of lowering cholesterol levels were found in fermented camel milk in Mongolia [108].
An interesting way to utilize the expansion of substrate sources, with the growing human population increasing the demand for sustainable and safe protein sources, is the fermentation of algae. Garofalo and colleagues [109] describe a recently proposed method for conducting sustainable practices related to food production using microalgae, such as Chlorella vulgaris, Arthrospira platensis, and Dunaliella salina, through LAB, as a source of valuable compounds, including proteins, lipids, pigments, carotenoids, vitamins, polyunsaturated fatty acids, and steroids. The technology of microalgal fermentation using LAB may offer an efficient and economical solution to improve digestibility and nutritional value. Since algae are already valuable food themselves, through fermentation, we are able to increase the amount of protein and improve its bioavailability; increase the amount of bioactive peptides; improve the profile of phenolic compounds; improve the lipid profile; increase the amount of pigments; produce functional metabolites. A. platensis presents itself as a promising substrate for microbial growth and fermentation, enabling the production of valuable fermented foods and beverages from a nutritional, nutraceutical, and economic perspective. The use of algae as a supplement for LAB growth is not a new idea, as early as 1998, Parada and others in an in vitro study, successfully improved the growth parameters of LAB by adding A. platensis filtrate to the laboratory medium [110]. It is worth mentioning that the addition of microalgae significantly changes the dynamics and success of microorganisms in the fermented product, hence optimization studies are necessary for each product to achieve repeatable process results. Promising results were obtained in the case of dairy products, such as Ayran or probiotic yogurt. Research conducted by Çelekli and others [111] showed the beneficial effect of A. platensis addition on the growth and survival of L. delbrueckii spp. bulgaricus, L. acidophilus, S. thermophilus, and Bifidobacterium lactis. This is due to the rich nitrogen content in spirulina, such as proteins, peptides, and free amino acids, as well as minerals, B-group vitamins, exopolysaccharides, adenine, and other organic components that stimulate LAB growth. However, this solution is not without flaws, both in Ayran and in another dairy product, probiotic yogurt, the viscosity worsened, proportionally with the amount of added spirulina [112]. Moreover, there is a need for further research to identify and purify specific bioactive compounds resulting from microalgal fermentation that are significant for various biological functions. Another obstacle is the opinions and sensory acceptance of products supplemented with spirulina, hence future research aimed at improving sensory qualities is necessary [109].
Searching for new food sources is not the only issue as the needs of the constantly evolving human population grow; the problem of managing waste from the food industry is also increasing. Lactic acid bacteria possess the capacity to convert waste into beneficial products, notably lactic acid. In a study conducted by Uwamahoro et al. on the lactic acid production potential from bacterial strains originating from food waste, it was found that the Weissella viridescens WJ39 strain had the highest potential for lactic acid production. It’s important to note that when utilizing waste, the efficiency of metabolite production is not the only concern; the fact that waste is being processed and converted into valuable compounds is crucial [113]. Moreover, further results also indicate the impact of LAB on the food industry and environment, namely, the produced lactic acid had an antimicrobial effect. The same lactic acid lowered the pH of the soil and improved the availability of phosphorus, opening up possibilities for waste bioconversion to enhance soil quality and enrich it with nutrients. The use of lactic acid bacteria in the management of food waste emphasizes their importance in creating new innovative and ecological solutions that support sustainable development and efficient management of natural resources.
3. Environmental Impact of Lactic Acid Bacteria
LAB are microorganisms suitable for application in sustainable food production because their use contributes to reducing the environmental footprint of food production. Mentioned in the previous chapter, the widely used fermentation of various products with LAB not only extends the shelf life of these foods, but also contributes to reducing food waste by preventing spoilage. However, this is not the only possible manner in which LAB bacteria can be involved in environmental protection. Research indicated that they have a multifaceted role in waste reduction, energy efficiency and environmentally friendly practices. Details are described in the following section.
3.1. Biodegradation of Organic Waste
The problem of increasing food waste is growing each year as the world’s population expands. Food waste is becoming one of the leading environmental problems, with the waste and losses produced at every point in the food supply chain. Multiple waste management methods have been developed for the safe disposal of food waste, however, they involve diverse problems such as high costs, environmental pollution and toxic side products. Food waste (FW) comprises edible food that, being produced in excessive quantities, is lost in the supply chain [114]. FW is composed of 30–60% starch, 5–10% protein and 10–40% lipids (w/w). Food waste is generated not only at the consumption, retail and wholesale level, but also during food processing and production [115]. Furthermore, discarded food is associated with excessive consumption of energy and water, which are needed for production, transport and distribution of food. Food waste management is also an extremely relevant issue, since food waste decays easily and generates an unpleasant odor that is detrimental to environmental quality [116]. The range of the problem is evidenced by intimidating statistics. The Food and Agriculture Organisation of the United Nations (FAO) has estimated that, worldwide, about one-third of the food produced for human consumption is wasted or lost. Estimates suggest that 1.6 billion tonnes of FW are generated globally each year, thus valuing the global economy at USD 2.6 trillion per year [117]. In the countries of the European Union alone, an estimated 88 million tonnes of food is wasted annually. The global food supply chain significantly differs according to the types of FW, post-harvest waste and overall consumer behavior. In the EU, in the food supply chain, about 40% of FW is generated at the retail and consumer level. One quarter of FW is generated at the household level and in the post-harvest and processing season this percentage increases to 40% [115]. There is therefore an urgent need to develop closed loop solutions that beneficially exploit and diminish FW in order to mitigate the related negative impacts [117].
In order to reduce the overall amount of waste and minimize the environmental impact, the use of microorganisms, such as lactic acid bacteria, is a promising and relevant solution. The high adaptability of LAB to inhabit various plant niches and substrates, together with their extensive enzymatic machinery, cause this group of bacteria to be considered cell factories for the production of various chemicals of interest. Considering the composition of the waste matrix and the metabolic characteristics of LAB, these bacteria will be able to conduct controlled fermentation of substrates contained in FW [118]. Plant-based food waste contains simple carbohydrates, and the high moisture and solids content enhances the accessibility of nutrients to microorganisms. In consideration of these attributes, these substrates are particularly suitable for processing via anaerobic digestion. According to their adaptations and metabolic capabilities, LAB are capable of fermenting organic materials and decomposing them into simple compounds. This process can not only be exploited to transform the food matrix during fermentation, but this property can also be harnessed to decompose organic waste, such as food or agricultural waste. LAB can be encountered in any environment rich in carbohydrates. This can include waste substrates comprising these substances, in particular food residues, which provide an excellent substrate for LAB growth. This application of LAB can allow not only cost reduction and waste recycling, but also offers the possibility of producing derived products. Certainly, the fermentation of FW using LAB opens up opportunities for a sustainable, closed-loop economy.
In the context of biodegradation of compounds present in organic waste, certain physiological and metabolic characteristics of LAB are crucial. LAB produce energy in the form of ATP mainly by fermenting sugars since they do not synthesize components of the respiratory chains (cytochromes, porphyrins) and are unable to generate ATP through a proton gradient mechanism. Thus, LAB do not utilize oxygen, but they are able to grow in its presence, and protection from oxygen by-products is ensured by peroxidases [119]. LAB, depending on the specific strain, have the ability to decompose various polysaccharides, which provides them with energy, while a by-product can be the production of substances useful to humans [120]. Disaccharides are hydrolysed by LAB to monosaccharides, which then proceed into their respective metabolic pathways. Many LABs are also able to digest pentoses, converting them to lactate and acetate, without releasing CO2 [119]. The degradation of macromolecular proteins is also an important process during FW processing. The LAB proteolytic system degrades proteins into peptides and further converts them into amino acids. They are essential for bacterial growth, but can also be metabolized by LAB into a variety of aromatic compounds such as aldehydes, esters and alcohols. Furthermore, amino acid metabolism is of vital importance for LAB to adapt to the environment, especially in their adaptation to an acidic environment. The NH3 generated in the deamination of amino acids can elevate the pH value inside and outside the cell, thus providing protection of the cell against acid stress [2]. LAB also features enzymes related to lipid metabolism, i.e., lipases that break down lipids to fatty acids and glycerol. This group of microorganisms is capable of performing unique reactions of fatty acid transformation, such as hydration, isomerization, dehydration and saturation. Some side products of their lipid metabolism can be employed for nutraceutical or medicinal purposes (e.g., conjugated linoleic acid) [119].
It has been shown that by employing the proper microorganisms, it is possible to eliminate nutrient-rich FW and use it as a feedstock for the production of materials, compost, chemicals or fuels [115]. In recent years, such valorisation of waste biomass has attracted considerable interest as an effective solution for waste recycling and, simultaneously, as a means of obtaining useful substances at low cost. Depending on the specific properties of FW, the most suitable valorization options are the production of enzymes, exopolysaccharides, biopolymers, biofuels or extraction of various bioactive compounds. Food bio-waste rich in sugars, proteins and lipids is a promising raw material especially for lactic acid production by LAB [121]. Regarded as one of the world’s top ten green molecules of the future, lactic acid has received tremendous attention for its many applications as a chemical intermediate [122]. It has been observed that although FWs are potentially a good medium for lactic acid production, the heterogeneity of the biomass being a mixture of different residues can be a hindrance to scale-up of lactic acid production. Several studies have demonstrated that lactic acid production can be accomplished through fermentation of FW with different microbial consortia fuels [115,123]. In their publication Wang et al. [124] discussed several factors affecting LAB-mediated fermentative lactic acid production and described the feasibility of lactic acid production from starch, cellulose and kitchen wastes. The authors indicated possible future research directions to optimize lactic acid production and the potential for industrial production of lactic acid from organic waste. Dedenaro et al. [125] investigated the potential ricotta cheese whey and pear pomace as a low-cost nutrient source for lactic acid fermentation. Using L. casei and Lactobacillus farciminis under microaerophilic conditions and mild sterility, an overall fermentation yield of 90% and a volumetric yield of 0.42 g lactic acid/L*h was achieved [125]. Another work describes the conversion of food waste to lactic acid using L. casei Shirota. This fermentation resulted in 94.0 g/L and 82.6 g/L of lactic acid with yields of 2.61 g/L*h and 2.50 g/L*h for mixed food waste and bakery waste hydrolysate, respectively [126]. The process of converting FW to lactic acid was also presented in the work of Pleissner et al. [127] where lactic acid was produced from mixed food waste from restaurants through a process of simultaneous saccharification and fermentation at laboratory and technical scale. In contrast to the previously mentioned studies, in this case Lactobacillus sp. strains did not show efficient conversion of food waste into lactic acid (productivity of 0.27–0.53 g/L*h and yield of 0.07–0.14 g/L*h) [127]. However, Al-Dhabi et al. [115] demonstrated that using a batch fermenter and Lacticaseibacillus rhamnosus AW3 isolated from wastewater, lactic acid can be obtained from FW and municipal sludge (MS). Utilizing FW and MS (2:0.5 ratio) with nutrient supplements and appropriate pH, a lactic acid production yield of 28.4 ± 0.87 g/L was achieved [115]. Song et al. [123] addressed the recycling of household FW to produce lactic acid. They took an approach to avoid producing many by-products, and did not require a saccharification process, which increases production costs. FW provided the opportunity for direct lactic acid production through inoculation by Lactobacillus delbrueckii subsp. bulgaricus, Lacticaseibacillus rhamnosus ATCC 7469, and Streptococcus thermophilus. The use of L. rhamnosus ATCC 7469 yielded 30.25 g/L lactic acid (at 37 °C, pH 6.8) from household FW. In the study, it was observed that the large inoculum size and substrate concentration resulted in a high concentration of lactic acid (26.8 g/L on average), but not a high yield of its production (0.20 g/g) [128]. A different approach to the exploitation of FW for lactic acid production was developed by Anagnostopoulou et al. [129], who performed fermentation using native microflora with the addition of two lactic acid bacteria (L. plantarum BS17 and L. casei BP2). The study showed the best lactic acid production (23.07 g/L) at pH 6.5, at 37 °C and a fermentation period of 3.5 days [129]. The literature also reports a study of lactic acid production from food waste under industrial conditions. After initial optimization of the results on a smaller scale, the use of compost as inoculum and the demonstrated optimal conditions for the process (35 °C and pH 5) increased the growth of Lactobacillus sp. resulting in high pilot-scale lactic acid production (68 g/L), yield (0.38 g/gTS) and selectivity (77%). The authors also moved a step further and, to integrate the process into the full value chain, the fermentation residues were further transformed into biogas in an anaerobic digestion process [130]. Lactic acid was also successfully obtained from a mixture of FW supplemented with the addition of digestate as a nitrogen source and sucrose as a readily bioavailable carbon source. Despite the addition of sucrose, it was shown that such lactic acid production was cost-effective. Its value not only exceeded the cost of the sucrose additive, but more importantly, the waste was neutralized in the first place [117]. The aforementioned investigations suggest that FW can be successfully used to produce lactic acid, but this requires the use of a suitable consortium of microorganisms and optimization of fermentation conditions.
Another approach to convert FW is composting, which transforms decaying organic materials into a stabilized form that is suitable for use as a soil conditioner with no potential for phytotoxicity. Asano et al. [131] in their study described LAB as a major system component in anaerobic composting. The waste processing was performed under anaerobic conditions, low pH and high temperature. Through the presence of LAB, the system achieved high stabilization by inhibiting the growth of other microorganisms and preventing waste from putrefaction [131]. Other work examined the effect of inoculation with the lactic acid bacterium Pediococcus acidilactici TM14 on native microorganisms resulting in accelerated composting of food waste. The raw compost material in the study comprised rabbit food mixed with organic acids which simulate food waste. Inoculation of raw compost with P. acidilactici TM14 accelerated the composting process. Applied bacterium synthesized lactic acid in high concentration and inhibited the production of toxic acetic acid, thus increasing the activity of fungi that can decompose organic acids. This resulted in the proliferation of thermophilic bacteria and enhanced degradation of organic matter, which resulted in accelerated composting [116]. As verified in composting, it is also known that homofermentative organisms are better suited for lactic acid production and stabilizing microbial populations. In the context of food waste processing, the abundance of Pediococcus (a homofermentative bacterium) was found to positively influence the efficiency of lactic acid production, which favors a faster composting process. Pediococcus, by producing lactic acid, limits the production of acetic acid, which in turn stimulates the activity of microorganisms necessary for composting. However, the presence of Weissella (a heterofermentative lactic acid bacterium) leads to the generation of acetic acid, which negatively impacts composting microorganisms, inhibiting the decomposition of organic substances. When both types of bacteria coexist in the initial material, the optimal course of composting depends on the proportion of these bacteria [132].
FW processed by LAB can also serve as animal feed. Yang et al. [133] conducted a study to determine the effect of lactic acid bacteria (Ligilactobacillus salivarius) on the physico-chemical and microbiological characteristics of mixed FW (containing bakery by-product, food waste, broiler poultry litter and barley and wheat bran) serving as pig feed. Anaerobic storing of FW with the addition of LAB proved to be an effective storage method. Inoculation with L. salivarius was found to be beneficial in stimulating microbial fermentation and improving the fermentative properties of FW. The inclusion of LAB inoculants in the FW increased the total and lactic acid bacteria counts and further improved the nutritional value resulting from the significant breakdown of fibre into soluble carbohydrates. Therefore, a diet containing mixed FW fermented with LAB can be successfully applied in pigs [133]. Moreover, Seo et al. reported on the use of fermentation by LAB isolated from the intestine and faeces of pigs (including L. plantarum CJY-22, L. brevis CJY-42, L. arizonensis CJY-3 and Pediococcus sp. CJY-41) to convert food waste into probiotic animal feed [134].
3.2. Biogas Production
Directly related to the topic of processing organic food waste is the aspect of bioconversion to biogas. Food waste can be processed using anaerobic digestion (AD), a biological process in which organic substrate is decomposed by microorganisms in the absence of oxygen. This technology is now widely used to produce renewable biogas energy. LAB are employed in AD processes, where organic waste is converted into biogas, which can be used as a renewable energy source for heating, electricity generation, or as a vehicle fuel, promoting energy efficiency and reducing dependence on fossil fuels. To increase the sustainability of the food industry, it is crucial to upgrade AD protocols to achieve higher biogas content and reduce industrial waste pollution. LAB generate substances such as acids, peptides, carbon dioxide or fatty acids that participate in methanogenesis and have the capacity to enhance biomethane production [135]. In their study, Bohn et al., [136] revealed that there is a wide range of LAB in biogas plants, especially species of the genus Lactobacillus with significant abundance (105 copies of each species per ml of fermenter sample) and high viability. The physiological characteristics of LAB, mainly the ability to degrade polymers, utilize carbohydrates and produce acids (lactic and acetic), indicate that these bacteria have the potential to play an active role in fermenters for biogas production. Satpathy et al. [137] investigated the effect of lactic acid on biogas production from substrates such as fresh corn and corn silage. Several types of samples (from an agricultural biogas plant, a wastewater treatment plant and a standardized laboratory reactor) were used as inoculum to study the effect of starter culture on the process. The results of this experiment suggest that lactic acid is an essential intermediate in biogas production and plays an important role in increasing total biogas production. Furthermore, the ability of the starter inoculum to exploit lactic acid is an essential factor in process optimization and increased biogas production [137]. The addition of LAB during biofuel production thus appears to be a beneficial factor. A study by Menardo et al. [138] showed that the addition of LAB slightly increased lactic and acetic acid content in silages containing corn stalks for methane production. While this fermentation did not significantly affect total methane production from corn stalks, it substantially improved the rate of methane production from the same samples. Also, Vervaeren et al. [139] in their study showed that biological additives for corn ensiling affected biomethane production efficiency during AD (up to 22.5% increase). In the study, the addition of homo- and hetero-fermentative LAB with a high ability to produce lactic acid led to successful ensiling, although it reduced biogas and biomethane yields when compared to the use of more complex additives that can promote hydrolysis during AD. Thus, it can be concluded that inoculum for maize ensiling should be considered cautiously, based on whether the ensiled substrate is subsequently subjected to AD [139]. Juodeikiene et al. in their study confirmed the potential of Lactobacillus delbrueckii spp. bulgaricus in biomethane production to reduce dairy wastewater pollution. The bacterium was found to produce high levels of volatile acidic compounds and, among the strains tested, was found to be the most suitable for dairy wastewater treatment, resulting in increased methanogenesis and biogas yields [135]. Similarly, a study by Vasmara et al. [140] showed that post-fermentation wastewater (also in combination with pig slurry) is a feedstock that can be used to produce biomethane by anaerobic digestion. In this study, biogas was produced from fermentation wastewater from the culture of two LAB strains (Lactococcus lactis subsp. lactis and Streptococcus thermophilus) on digested ricotta cheese whey in monofermentation or co-digestion with pig slurry. High yields of biomethane production were obtained in monofermentation (average 372 mL CH4 g−1 VS) and in co-digestion with pig slurry (average 416 mL CH4 g−1 VS). Co-fermentation halved the duration of AD [140].
Since the economic benefits of AD are sometimes questioned, and in order to increase the economics of the treatment process, attempts are being made to convert FW not only to biogas, but simultaneously to other products such as lactic acid. In their investigations, Kim et al. [141] performed FW fermentation using a native mixed culture containing LAB (predominantly Lactobacilli). The implemented process allowed the recovery of highly purified LA and a solid residue, which was further digested to biogas by AD. It was shown that 47 kg of LA and 54 m3 of biogas may be recovered by the developed process per tonne of FW with a COD removal efficiency of 70%, which provides a higher economic value than biogas extraction itself [141].
3.3. Environmental Treatment and Restoration
Pollution by heavy metals, dyes, pesticides and other hazardous substances is a significant environmental problem and a threat to human health arising from industrial development. Certain LAB strains have been investigated for their potential in environmental restoration projects since this group of bacteria may contribute to the bioremediation of polluted environments by assisting in the degradation of organic pollutants. Bioremediation is a biotechnological process in which environmental contaminants may be decomposed through the action of microorganisms to ensure a clean environment without hazards. This process uses the activity of microorganisms to degrade environmental contaminants that are harmful to human health and transforms them into less toxic or non-toxic forms. Microorganisms have the ability to mineralise toxic wastes into further products, such as biomass and water. Research conducted over recent years has shown that LABs have the ability to degrade most organic waste, as well as other industrial pollutants such as dyes [142]. Moreover, they are known to absorb and accumulate heavy metals, which represent one of the most acute environmental problems and risks to human health associated with industrial progress. Most LAB interactions with metals involve surface processes, i.e., adsorption of metal contaminants to functional groups on the cell surface capable of binding cations [103]. LAB strains are widespread in the soil, where they regulate soil organic matter and the biochemical cycle. They are essential for improving the soil carbon pool, detoxification of hazardous chemicals, mycotoxins and improving plant health [4]. In addition, LAB was found to detoxify and decompose pesticides present in fermented milk and other fermented products [143]. Owing to these properties, LAB can be exploited for the production of commercial biofilters for the treatment of heavy metal and aflatoxin-contaminated water. Studies revealed that LAB-based microcapsules showed desirable biodegradability qualities compared to hydrogel and synthetic polymers and were more efficient [4]. Bioremediation of this type of contamination using LAB possesses a high level of safety and therefore is an important and interesting research topic. In their study, Ameen et al. [144] isolated and characterized metal-resistant LAB that could potentially be used in metal bioremediation. From among the isolates, L. plantarum MF042018 showed a high degree of resistance (up to 500 and 100 ppm, on nickel and chromium, respectively), with a multiple antibiotic resistance index above 0.5. This study also evaluated the biosorption capacity of L. plantarum MF042018 and determined the optimal conditions for this process (pH 2.0 and temperature 22 °C). The results demonstrated that the use of L. plantarum MF042018 is an efficient tool for the treatment of battery production wastewater contaminated with hazardous metals. Thus, this study suggests that L. plantarum MF042018 can be employed as a promising biosorbent for the detoxification of heavy metal contaminated environments, particularly from industrial wastewaters [144]. In another study, LAB strains were obtained from dadih lareh. Among the isolates obtained, one with antimicrobial activity also showed potential as a good probiotic and had the ability to remove heavy metal cadmium ions in the solution [145]. Also, Hasan et al. [146] in their study isolated LAB strains and tested them for their potential usefulness to bioremediate toxic lead and cadmium with their bioavailability and persistence. Five probiotic LAB strains selected based on their heavy metal resistance potential were subjected to additional in vitro assays to test their Pb and Cd binding and removal efficiencies and to assess their bioavailability and survival in a dynamic in vitro gastrointestinal digestion model. The results indicated that all strains had high resistance to tested metals and effectively removed Pb and Cd. Of the isolates tested, Lactobacillus delbrueckii subsp. bulgaricus LDMB02 showed the best Pb and Cd removal rates. Furthermore, this study demonstrated that these strains significantly reduced the bioavailability of Pb and Cd (from 42 to 50% and from 40 to 58%, respectively), and their survival rates in the presence of Pb and Cd, ranged from 80.1 to 85.4% and from 81.5 to 87.5%, respectively. This study therefore demonstrated the high resistance of LAB strains to Pb and Cd with remarkable efficiency in binding and reducing the bioavailability of these metals with high survival rates. This investigation points to the tremendous potential use of LAB as a probiotic that can be incorporated as a food additive and provide affordable option for the protection of human health, especially suitable for use in developing countries where many people are unknowingly exposed to toxic heavy metals on a regular basis [146]. Effective elimination of cadmium and lead from water by LAB strains was also observed in the research performed by Halttunen et al. [147]. In this study, the lactic acid bacteria most effective in removing metals were Bifidobacterium longum 46, Bifidobacterium lactis Bb12 and Limosilactobacillus fermentum ME3. The highest maximum removal capacities of Cd and Pb ions (54.7 mg metal/g and 175.7 mg/g dry biomass, respectively) were obtained for B. longum 46 [147].
Beyond the possibility of applying LAB in effective bioremediation and detoxification of heavy metals and mycotoxins, they can also be employed to produce beneficial industrial and agro-products that are safe for the environment. Accordingly, LABs are widely used in the advancement of agricultural products, as they are safe, environmentally friendly, have low production costs and a rapid development rate. As fertilizers, LAB can accelerate the organic content of soil, foster biodegradation, generate organic acids and bacteriocin metabolites. LAB exhibit antagonistic effects against phytopathogens, inhibiting bacterial and fungal populations in the phyllosphere and rhizosphere. These applications of LAB help protect plants and promote their growth for more sustainable production of plant-based foods, with the goal of reducing the negative impact on the environment [4].
3.4. Biopolymers Production
The excessive use of conventional plastic materials as food packaging leads to numerous ecological challenges, most notably the generation of large amounts of waste, depletion of natural resources, and global warming. The amount of waste generated from synthetic polymers is increasing at an alarming rate. Equally worrying, it has been reported that less than 10% of the synthetic plastic produced are recycled, posing serious sustainability concerns regarding the production of synthetic polymers. Considering the growing concerns about the environmental footprint of packaging waste, approaches are being sought to acquire and apply sustainable and eco-friendly packaging. Biomaterials originating from sustainable and renewable biomass, compared to manufactured petrochemical products, minimize the harmful impacts on the environment. Using biopolymers is thus a favorable method for replacing synthetic polymers in the interest of environmental awareness. Environmentally friendly, biodegradable bioplastics produced by microorganisms offer an alternative to plastics derived from hydrocarbons. These biodegradable polymers include, among others, polyhydroxyalkanoates (PHAs), exopolysaccharides (EPS), as well as polylactic acid (PLA). These bioplastics are stored inside the cells of the microorganisms or secreted from cells and generally serve them as a source of carbon and energy or cell protection [148]. Lactic acid bacteria can also serve as a source of biopolymers. Owing to their good biodegradability and the potential to be derived from low-cost biomass, these polymers are now attracting increasing interest not only in research but also with potential for industrial application. The biopolymers produced by LAB are used in the food industry mainly as food packaging. They provide a healthier and safer alternative to traditional plastics, in addition to effectively extending the shelf life of products, helping to reduce microbial contamination of food during transport and storage and even promoting nutrient preservation. LABs have been well-studied in the context of exopolysaccharide (EPS) production, but substantially less as producers of PLA and PHA [6].
3.4.1. Polylactic Acid (PLA)
Polylactic acid holds the leading position among bio-based plastics produced by LAB. Within the array of biodegradable plastics, PLA is both readily available and safe to be degraded after use with no environmental pollution. Moreover, with regard to various properties relevant for industrial applications, such as biocompatibility, mechanical and physical properties, PLA is on a par to conventional plastics such as Polyethylene terephthalate or polypropylene [149]. Because of the beneficial properties of this material, PLA production using lactic acid accounts for about 35% of the total bioplastics market. Furthermore, its use is environmentally friendly because it can be manufactured from renewable sources, including biomass or agricultural waste (molasses, corn steep liquor, wheat bran etc.) and has the potential to act as a CO2 sink, helping decrease greenhouse gas emissions [150]. PLA-based bioplastics could be an outstanding alternative to existing conventional plastics in a variety of applications, which can function not only to protect the environment from pollution, but also to act as a sustainable and economical product [149]. PLA is a natural product that is mainly applied to produce foils and containers for food packaging (primarily perishables such as fruit and vegetables) or disposable food grade packaging [151]. The use of PLA for food packaging offers a number of advantages over conventional materials because it is biocompatible, safe for consumption, industrially compostable and recyclable. The benefits of employing PLA in food packaging are reflected not only in its environmental properties but also in its ease of processing and excellent transparency. Unfortunately, PLA also has some disadvantages that hinder its industrial use, including poor mechanical properties, sensitivity to thermal degradation and low barrier properties to oxygen and other gasses. However, with these properties, it can serve as packaging for products with a shorter shelf life that do not require high resistance to water or oxygen penetration, such as fruits, vegetables, fresh juices, meat, and dairy products [152]. These limitations mentioned above restrict the wider use of PLA in food packaging and drive research related to enhancing PLA’s properties by combining it with other compounds or employing advanced processing or treatment techniques [153].
Clean and environmentally friendly strategies for LA production are now well established. The conventional synthesis of PLA is based on a biochemical hybrid process in which one of the monomers that comprise PLA (L-lactic acid or D-lactic acid) is produced by microbial fermentation from renewable resources, and subsequently PLA is synthesized by polymerisation or by direct solvent-based azeotropic dehydratve condensation of LA. Clean LA production can be achieved using membrane hybrid reactor systems. The enzymatic polymerisation of LA monomers into PLA is considered one of the most economical and environmentally friendly methods. An alternative to the traditional production process is one-step fermentative PLA production using metabolically modified microorganisms. Among LAB, well-known producers of lactic acid include L. rhamnosus, L. paracasei, L. delbrueckii and L. coryniformis subsp. torquens (Huang et al., 2021 [154]). PLA produced directly via fermentation demonstrates high strength and better performance compared to conventional methods, which is considered significant from a technical and economic point of view [6].
3.4.2. Polyhydroxyalkanoates (PHAs)
Polyhydroxyalkanoates (PHAs) represent a group of naturally occurring high molecular weight biopolymers (approx. 105 Da) produced by microorganisms. PHAs are polyesters produced via bacterial fermentation using pure bacterial cultures cultivated on a variety of renewable sources. This group includes polyhydroxybutyrate (PHB), polyhydroxyvalerate (PHV), and derived polymers, i.e., poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV), which are completely degradable. PHAs show several characteristics comparable to the fossil fuel feedstock-derived plastics, including physicochemical and mechanical properties, that render them potential substitutes for conventional plastics. In addition to their biodegradability, another advantage is the possibility of obtaining PHAs through fermentation of agro-industrial byproducts, e.g., milk and cheese whey [155]. This sustainable alternative to plastics has emerged as an interesting research biomaterial in recent decades because of its wide potential industrial applications [148]. However, the large-scale application of PHAs is still limited by their high production costs. The fermentation of complex low-cost substrates to obtain PHAs reduces manufacturing costs. PHAs can be obtained from cheap substrates by employing mixed cultures of lactic-acid-producing bacteria, including bacteria such as Lactobacillus delbrueckii, Lactococcus lactis or and C. necator. PHA production was also obtained for Leuconostoc mesenteroides, Lactobacillus bulgaricus, and Lactiplantibacillus plantarum [6]. Whereas previously, PHB was also observed to accumulate in cells of L. plantarum, L. brevis, L. acidophilus, L. casei, L. bulgaricus, L. bifidus and L. fermentum. c [156].
It is anticipated that, despite the current technical and economic limitations in this area, the market for food packaging based on PHAs will expand, with a positive impact on environmental protection and the approval of consumers. In the food industry PHAs produced by LAB can be used primarily for packaging. Owing to their excellent moisture and oxygen-reducing properties and high melt strength, PHAs can be incorporated into bioplastics used to make waterproof cardboard boxes or cups. Furthermore, PHA-based bioplastics are regarded as a biodegradable alternative to aluminum foil [6]. Known possibilities for the use of PLA-based food packaging include the production of disposable food containers, utensils and films, bags, tubs, trays, hot and cold cups, cup lids, bottles, yogurt containers, jars, bowls and disposable food packaging [151,152,157]. The development of new bioplastics for broader use in food packaging can be accomplished by designing and formulating new properties to improve its functionality. In food packaging, antimicrobial substances such as silver and copper nanoparticles or bacteriocins can be integrated into PHAs to extend the shelf life of products and reduce microbial contamination, thereby reducing food waste [158].
3.4.3. Exopolysaccharides (EPS)
There is a growing need to eradicate the use of artificial ingredients and additives in the food industry. Therefore, natural food additives have become increasingly popular in recent years to meet consumer demand for safe and natural products. Microbial EPS, due to its technical-functional and rheological properties, can be a valued alternative for improving the physicochemical properties of food products [159]. Bacteria have the potential to produce EPS with a variety of often complex chemical structures that determine their functional qualities and biological activities. Bacterial EPS has a broad range of industrial applications, depending on its physicochemical and structural characteristics. There is great interest in the application of this polymer in the food industry, on account of its structural properties, including emulsification, gelling, sweetening, texturization, water binding capacity as well as its bioactive properties. Studies have also shown the health-promoting potential of EPS, encompassing prebiotic, antioxidant, immunomodulatory, anti-inflammatory and anti-biofilm effects [160].
Despite its appealing properties and wide potential for applications, few known EPSs enter the market because of the relatively high production costs associated with the expense of carbon sources and the production yield of this polymer. Bacterial EPS can be produced ex situ or in situ. Ex situ synthesis involves controlled fermentation where a high-quality, reproducible product is incorporated as ingredient or additive into a food product. This process is easier to control than in situ manufacture and gives specific results in terms of monitored EPS properties. However, the application of in-situ produced EPS is a feasible alternative to replace conventional additives and in order to achieve more environmentally and consumer-friendly ‘clean label’ foods.
Nowadays, LAB-produced EPSs have gained particular interest in the food industry because of their ability to enhance the rheological properties of food products, especially fermented foods [6]. LAB are capable of synthesizing EPS that are either heteropolysaccharides or homopolysaccharides. Tremendous structural diversity has been observed in the EPS produced, and these differences relate to monosaccharide composition, conformation, chain length, molecular weight, size, structure and electrical charge. The most studied EPS from LAB with food applications are dextran, kefiran, inulin, alternan, reuteran, levan and hyaluronic acid [159,161].
It is worth noting that EPS synthesis is strain-dependent and highly influenced by the processing conditions (i.e., temperature, pH, carbon source and nutrients present in the culture medium, incubation time, etc.) [162]. LAB strains produce EPS in amounts ranging from 10 mg/L to 400 mg/L under non-optimized culture conditions, while under favorable cultivation parameters it is possible to obtain multiples of this amount [159]. The majority of EPS-producing LAB belong to the genera Lactobacillus, Lactococcus, Leuconostoc, Streptococcus and Weissella. The ability to produce EPS has been observed for about 30 species among LAB, in particular L. casei, L. plantarum, L. brevis, L.rhamnosus, L. acidophilus, L. helveticus, L. curvatus, L. delbrueckii subsp. bulgaricus, L. johnsonii, etc. [6]. The diversity of EPS produced by LAB and known producers of this polymer has recently been thoroughly reviewed by [160].
These diverse and specific characteristics provide EPS with numerous functional properties, which in turn determine particular applications in the food production. EPS produced by LAB have multiple applications in the food industry, especially in fermented products, where they are produced in situ. In addition, they could be added as food ingredients when produced ex situ. These polymers are considered natural bio-thickeners and natural ingredients for functional foods [6]. Valued in such applications are the ability to form gels, influence the taste, prevent syneresis and the capacity to form films [163]. As food additives, EPSs ensure food products’ creaminess, firmness and mouthfeel. EPS are incorporated into a variety of food products, including bakery products (bread, gluten-free products, sourdough), fermented dairy products (i.e., yoghurt, kefir, cheese, low-calorie quark) and fermented vegetable products (e.g., drinks, vegetable purees and cakes). For example, in their study, Hassan et al. [164] showed that EPS from Streptococcus thermophilus, consisting of galactose, rhamnose and fucose, contributes to increased moisture content in Karish cheese samples. Also in low-fat mozzarella, EPS-producing Streptococcus thermophilus MR-1C was observed to increase moisture retention [165]. Similar investigations were carried out on Cheddar cheeses, for which EPS-producing starter cultures were used, including Lactococcus lactis subsp. cremoris, Weissella and Lactobacillus. These studies also reported increased moisture levels in the cheeses and better structural properties [166,167]. EPS produced by Streptococcus thermophilus has also been used as a stabilizer and viscosity enhancer in ice cream. The developed functional fermented ice creams did not require the addition of artificial stabilizers since the presence of EPS produced in situ by starter cultures improved their physicochemical, microstructural and rheological properties [168].
Another application of exopolysaccharides in the food industry towards sustainable production is the utilization of EPS for food packaging. The subject of interest is the use of EPS produced by LAB to develop new food coatings that can substitute for environmentally unfriendly packaging [6]. The structural integrity and smooth, glossy surface of certain EPS produced from LAB makes them suitable for use in the preparation of edible films/coatings. The structural integrity and smooth, glossy surface of some EPS produced from LAB facilitates their use in the preparation of edible films/coatings, whereas EPS with a highly porous, dull structure is not suitable for film production and requires plasticization [161]. An example of research on the development of an edible film based on cassava starch, sodium carboxymethylcellulose and glycerol with the inclusion of EPS-producing LAB strains (Lactiplantibacillus plantarum and Pediococcus pentosaceus) was reported by Li et al. [169]. The film, by forming a barrier to water and light, extended the shelf life of bananas and additionally exhibited good antioxidant activity. This type of packaging does not contribute to environmental pollution. Moreover, it is safe for consumers and further reduces food waste by keeping food fresh longer.
4. Conclusions
Due to their respective history, LABs are mainly associated with food fermentation and thus preservation, increasing microbiological safety, and improving nutritional value. The traditional use of LAB hides millennia of diversity in cultural heritage and a cross-section of the diets of different nations, while at the same time demonstrating the evolutionary adaptation of LAB to the vastness of natural environments. From this adaptive abundance, we benefit today by constantly expanding their use. LAB’s exploration of food fermentation represents a key intersection of traditional food technology practices and contemporary scientific research, offering deep insights into sustainable food production. Nowadays, thanks to their metabolic capabilities, they contribute to the removal of organic waste, facilitating its recycling and valorisation. The most relevant of the bioproducts obtained using LAB from food waste are LA and biogas. In the sustainable production of both food and food packaging, biopolymers produced by LAB are also significant, providing more environmentally friendly alternatives to conventional plastics. Advancements in the use of LAB for the disposal of food waste, the production of food ingredients/additives and packaging are being made possible through intensive research focused on this topic. Although currently still few of these applications are used on a large scale, optimization of these processes will allow for wider exploitation in the future and potentially the development of new opportunities for LAB applications in sustainable food production at different stages.
Author Contributions
Conceptualization, K.G. and K.R.; writing—original draft preparation, K.G. and K.R.; writing—review and editing, K.G. and K.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AD | anaerobic digestion |
| ATCC | American Type Culture Collection |
| ATP | Adenosine triphosphate |
| ATPase | F0F1-Adenosine triphosphate Synthase |
| CH4 | Methane |
| CO2 | carbon dioxide |
| COD | chemical oxygen demand |
| DNA | Deoxyribonucleic acid |
| EPS | Exopolysaccharide |
| EU | European Union |
| FAO | The Food and Agriculture Organisation |
| FOS | Fructooligosaccharides |
| FW | food waste |
| HA | heterocyclic amines |
| LA | lactic acid |
| LAB | Lactic acid bacteria |
| MRS broth | De Man, Rogosa and Sharpe broth |
| MS | municipal sludge |
| NA | N-nitrosamine |
| NDMA | nitrosodimethylamine |
| NH3 | ammonia |
| PAH | polycyclic aromatic hydrocarbon |
| PHA | polyhydroxyalkanoate |
| PHV | polyhydroxyvalerate |
| PHVB | poly(3-hydroxybutyrate-co-3-hydroxyvalerate) |
| PLA | polylactic acid |
| ROS | reactive oxygen species |
| USD | The United States dolar |
References
- Fanzo, J.; Rudie, C.; Sigman, I.; Grinspoon, S.; Benton, T.G.; Brown, M.E.; Covic, N. Sustainable food systems and nutrition in the 21st century: A report from the 22 nd annual Harvard Nutrition Obesity Symposium. Am. J. Clin. Nutr. 2022, 115, 18–33. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H. El Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef] [PubMed]
- Raman, J.; Kim, J.S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.J.; Kim, S.J. Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations. Int. J. Mol. Sci. 2022, 23, 7784. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Xu, J.L.; Wang, Z. Microalgae Biotechnology for Food, Health and High Value Products; Springer: Berlin/Heidelberg, Germany, 2020; ISBN 9789811501692. [Google Scholar]
- Nicolescu, C.M.; Bumbac, M.; Buruleanu, C.L.; Popescu, E.C.; Stanescu, S.G.; Georgescu, A.A.; Toma, S.M. Biopolymers Produced by Lactic Acid Bacteria: Characterization and Food Application. Polymers 2023, 15, 1539. [Google Scholar] [CrossRef]
- Zapaśnik, A.; Sokołowska, B.; Bryła, M. Role of Lactic Acid Bacteria in Food Preservation and Safety. Foods 2022, 11, 1283. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Mitchell, R. Fermentation and pathogen control: A risk assessment approach. Int. J. Food Microbiol. 2002, 79, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Tucker, G. Pasteurization: Principles and Applications, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; ISBN 9780123849533. [Google Scholar]
- Tassou, C.C.; Panagou, E.Z.; Katsaboxakis, K.Z. Microbiological and physicochemical changes of naturally black olives fermented at different temperatures and NaCl levels in the brines. Food Microbiol. 2002, 19, 605–615. [Google Scholar] [CrossRef]
- Haque, A.; Richardson, R.K.; Morris, E.R. Effect of fermentation temperature on the rheology of set and stirred yogurt. Food Hydrocoll. 2001, 15, 593–602. [Google Scholar] [CrossRef]
- Chung, H.C.; Jeong, B.Y.; Han, G.D. Optimum conditions for combined application of Leuconostoc sp. and Saccharomyces sp. to sourdough. Food Sci. Biotechnol. 2011, 20, 1373–1379. [Google Scholar] [CrossRef]
- Wouters, D.; Bernaert, N.; Anno, N.; Van Droogenbroeck, B.; De Loose, M.; Van Bockstaele, E.; De Vuyst, L. Application’ and validation of autochthonous lactic acid bacteria starter cultures for controlled leek fermentations and their influence on the antioxidant properties of leek. Int. J. Food Microbiol. 2013, 165, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, W.H. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 2002, 75, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Motarjemi, Y. Impact of small scale fermentation technology on food safety in developing countries. Int. J. Food Microbiol. 2002, 75, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Wirawati, C.U.; Sudarwanto, M.B.; Lukman, D.W.; Wientarsih, I.; Srihanto, E.A. Diversity of lactic acid bacteria in dadih produced by either back-slopping or spontaneous fermentation from two different regions of West Sumatra, Indonesia. Vet. World 2019, 12, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, N.; Ananthanarayan, L. Use of the backslopping method for accelerated and nutritionally enriched idli fermentation. J. Sci. Food Agric. 2015, 95, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Budzyńska, A.; Grudlewska-Buda, K.; Wiktorczyk-Kapischke, N.; Andrzejewska, M.; Wałecka-Zacharska, E.; Gospodarek-Komkowska, E. Two Faces of Fermented Foods—The Benefits and Threats of Its Consumption. Front. Microbiol. 2022, 13, 845166. [Google Scholar] [CrossRef] [PubMed]
- García-Díez, J.; Saraiva, C. Use of starter cultures in foods from animal origin to improve their safety. Int. J. Environ. Res. Public Health 2021, 18, 2544. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Potes, M.E.; Elias, M. Role of starter cultures on the safety of fermented meat products. Front. Microbiol. 2019, 10, 853. [Google Scholar] [CrossRef]
- Yu, Z.; Peng, C.; Kwok, L.Y.; Zhang, H. The bacterial diversity of spontaneously fermented dairy products collected in northeast asia. Foods 2021, 10, 2321. [Google Scholar] [CrossRef]
- Malo, P.M.; Urquhart, E.A. Fermented Foods: Use of Starter Cultures. In Encyclopedia of Food and Health; Academic Press: Cambridge, MA, USA, 2015; pp. 681–685. ISBN 9780123849533. [Google Scholar]
- Picon, A. Cheese microbial ecology and safety. In Global Cheesemaking Technology: Cheese Quality and Characteristics; Wiley-IEEE Press: Hoboken, NJ, USA, 2017; pp. 71–99. ISBN 9781119046165. [Google Scholar]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Aghababaie, M.; Khanahmadi, M.; Beheshti, M. Developing a kinetic model for co-culture of yogurt starter bacteria growth in pH controlled batch fermentation. J. Food Eng. 2015, 166, 72–79. [Google Scholar] [CrossRef]
- Frantzen, C.A.; Kleppen, H.P.; Holo, H. Lactococcus lactis diversity in undefined mixed dairy starter cultures as revealed by comparative genome analyses and targeted amplicon sequencing of epsD. Appl. Environ. Microbiol. 2018, 84, e02199-17. [Google Scholar] [CrossRef] [PubMed]
- Pavunc, A.L.; Beganović, J.; Kos, B.; Uroić, K.; Blàžić, M.; Šušković, J. Characterization and application of autochthonous starter cultures for fresh cheese production. Food Technol. Biotechnol. 2012, 50, 141–151. [Google Scholar]
- Xiong, T.; Li, X.; Guan, Q.; Peng, F.; Xie, M. Starter culture fermentation of Chinese sauerkraut: Growth, acidification and metabolic analyses. Food Control 2014, 41, 122–127. [Google Scholar] [CrossRef]
- Mun, S.Y.; Chang, H.C. Characterization of Weissella koreensis sk isolated from kimchi fermented at low temperature (Around 0 °C) based on complete genome sequence and corresponding phenotype. Microorganisms 2020, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; García-Vaquero, M. Bioactive Compounds from Fermented Food Products. In Novel Food Fermentation Technologies; Food Engineering Series; Springer: Berlin/Heidelberg, Germany, 2016; pp. 293–310. [Google Scholar]
- Ferranti, P.; Nitride, C.; Nicolai, M.A.; Mamone, G.; Picariello, G.; Bordoni, A.; Valli, V.; Di Nunzio, M.; Babini, E.; Marcolini, E.; et al. In vitro digestion of Bresaola proteins and release of potential bioactive peptides. Food Res. Int. 2014, 63, 157–169. [Google Scholar] [CrossRef]
- Di Cagno, R.; Chaves Lòpez, C.; Tofalo, R.; Gallo, G.; De Angelis, M.; Paparella, A.; Hammes, W.P.; Gobbetti, M. Comparison of the compositional, microbiological, biochemical and volatile profile characteristics of three Italian PDO fermented sausages. Meat Sci. 2008, 79, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Bis-Souza, C.V.; Penna, A.L.B.; da Silva Barretto, A.C. Applicability of potentially probiotic Lactobacillus casei in low-fat Italian type salami with added fructooligosaccharides: In vitro screening and technological evaluation. Meat Sci. 2020, 168, 108186. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Yang, X.; Zhang, K.; Zhao, Y.; Cheng, F.; Jin, Y. Influence of Lactobacillus helveticus ZF22 and TR1-1-3 strains on the aromatic flavor of fermented sausages. Front. Nutr. 2023, 9, 1058109. [Google Scholar] [CrossRef]
- Ammor, M.S.; Gueimonde, M.; Danielsen, M.; Zagorec, M.; Van Hoek, A.H.A.M.; De Los Reyes-Gavilán, C.G.; Mayo, B.; Margolles, A. Two different tetracycline resistance mechanisms, plasmid-carried tet(L) and chromosomally located transposon-associated tet(M), coexist in Lactobacillus sakei rits 9. Appl. Environ. Microbiol. 2008, 74, 1394–1401. [Google Scholar] [CrossRef]
- Mozaffari, S.; Abedi, K.A.D.; Garmaroudi, A.A. Assessing The Emergency Preparedness of The Educational Hospitals in Kurdistan’s University of Medical Sciences, using The Resilience Engineering Method In 2018. Prev. Nutr. Food Sci. 2021, 26, 476–484. [Google Scholar]
- Sionek, B.; Szydłowska, A.; Küçükgöz, K.; Kołożyn-Krajewska, D. Traditional and New Microorganisms in Lactic Acid Fermentation of Food. Fermentation 2023, 9, 1019. [Google Scholar] [CrossRef]
- Salvetti, E.; Fondi, M.; Fani, R.; Torriani, S.; Felis, G.E. Evolution of lactic acid bacteria in the order lactobacillales as depicted by analysis of glycolysis and pentose phosphate pathways. Syst. Appl. Microbiol. 2013, 36, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Šušković, J.; Kos, B.; Beganović, J.; Pavunc, A.L.; Habjanič, K.; Matoć, S. Antimicrobial activity—The most important property of probiotic and starter lactic acid bacteria. Food Technol. Biotechnol. 2010, 48, 296–307. [Google Scholar]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Peetermans, A.; Foulquié-Moreno, M.R.; Thevelein, J.M. Mechanisms underlying lactic acid tolerance and its influence on lactic acid production in Saccharomyces cerevisiae. Microb. Cell 2021, 8, 111–130. [Google Scholar] [CrossRef]
- Kawazoe, N.; Kimata, Y.; Izawa, S. Acetic acid causes endoplasmic reticulum stress and induces the unfolded protein response in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1192. [Google Scholar] [CrossRef]
- Rallu, F.; Gruss, A.; Maguin, E. Lactococcus lactis and stress. Antonie Van Leeuwenhoek 1996, 70, 243–251. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ohishi, H.; Benno, Y. H+-ATPase activity in Bifidobacterium with special reference to acid tolerance. Int. J. Food Microbiol. 2004, 93, 109–113. [Google Scholar] [CrossRef]
- Lowe, D.P.; Arendt, E.K.; Soriano, A.M.; Ulmer, H.M. The influence of lactic acid bacteria on the quality of malt. J. Inst. Brew. 2005, 111, 42–50. [Google Scholar] [CrossRef]
- Zhang, C.; Brandt, M.J.; Schwab, C.; Gänzle, M.G. Propionic acid production by cofermentation of Lactobacillus buchneri and Lactobacillus diolivorans in sourdough. Food Microbiol. 2010, 27, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Johanningsmeier, S.D.; McFeeters, R.F. Metabolism of lactic acid in fermented cucumbers by Lactobacillus buchneri and related species, potential spoilage organisms in reduced salt fermentations. Food Microbiol. 2013, 35, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yu, Q.; Xu, J.; Sun, H.; Cheng, Q.; Xie, Y.; Wang, C.; Li, P.; Chen, C.; Zheng, Y. Effect of different organic acid additives on the fermentation quality and bacterial community of paper mulberry (Broussonetia papyrifera) silage. Front. Microbiol. 2022, 13, 1038549. [Google Scholar] [CrossRef] [PubMed]
- Punia Bangar, S.; Suri, S.; Trif, M.; Ozogul, F. Organic acids production from lactic acid bacteria: A preservation approach. Food Biosci. 2022, 46, 101615. [Google Scholar] [CrossRef]
- Sorrells, K.M.; Enigl, D.C. Effect of pH, acidulant, sodium chloride, and temperature on the growth of Listeria monocytogenes. J. Food Saf. 1990, 11, 31–37. [Google Scholar] [CrossRef]
- Al-Rousan, W.M.; Olaimat, A.N.; Osaili, T.M.; Al-Nabulsi, A.A.; Ajo, R.Y.; Holley, R.A. Use of acetic and citric acids to inhibit Escherichia coli O157: H7, Salmonella Typhimurium and Staphylococcus aureus in tabbouleh salad. Food Microb. 2018, 73, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Zhang, X.; Xiao, Y.; Sheng, Q.; Tu, L.; Chen, F.; Yan, Y.; Zheng, Y.; Wang, M. Interaction of acetic acid bacteria and lactic acid bacteria in multispecies solid-state fermentation of traditional Chinese cereal vinegar. Front. Microbiol. 2022, 13, 964855. [Google Scholar] [CrossRef] [PubMed]
- Özcelik, S.; Kuley, E.; Özogul, F. Formation of lactic, acetic, succinic, propionic, formic and butyric acid by lactic acid bacteria. LWT 2016, 73, 536–542. [Google Scholar] [CrossRef]
- Vollenweider, S.; Grassi, G.; König, I.; Puhan, Z. Purification and structural characterization of 3-hydroxypropionaldehyde and its derivatives. J. Agric. Food Chem. 2003, 51, 3287–3293. [Google Scholar] [CrossRef]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The antimicrobial compound reuterin (3-hydroxypropionaldehyde) induces oxidative stress via interaction with thiol groups. Microbiology 2010, 156, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Langa, S.; Martín-Cabrejas, I.; Montiel, R.; Landete, J.M.; Medina, M.; Arqués, J.L. Short communication: Combined antimicrobial activity of reuterin and diacetyl against foodborne pathogens. J. Dairy Sci. 2014, 97, 6116–6121. [Google Scholar] [CrossRef]
- Gómez-Torres, N.; Ávila, M.; Gaya, P.; Garde, S. Prevention of late blowing defect by reuterin produced in cheese by a Lactobacillus reuteri adjunct. Food Microbiol. 2014, 42, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Muthukumarasamy, P.; Holley, R.A. Survival of Escherichia coli O157:H7 in dry fermented sausages containing micro-encapsulated probiotic lactic acid bacteria. Food Microbiol. 2007, 24, 82–88. [Google Scholar] [CrossRef]
- Bachmann, H.; Pronk, J.T.; Kleerebezem, M.; Teusink, B. Evolutionary engineering to enhance starter culture performance in food fermentations. Curr. Opin. Biotechnol. 2015, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Grosu-Tudor, S.S.; Stancu, M.M.; Pelinescu, D.; Zamfir, M. Characterization of some bacteriocins produced by lactic acid bacteria isolated from fermented foods. World J. Microbiol. Biotechnol. 2014, 30, 2459–2469. [Google Scholar] [CrossRef]
- Devi, S.M.; Ramaswamy, A.M.; Halami, P.M. In situ production of pediocin PA-1 like bacteriocin by different genera of lactic acid bacteria in soymilk fermentation and evaluation of sensory properties of the fermented soy curd. J. Food Sci. Technol. 2014, 51, 3325–3332. [Google Scholar] [CrossRef]
- Maldonado-Barragán, A.; Caballero-Guerrero, B.; Lucena-Padrós, H.; Ruiz-Barba, J.L. Induction of bacteriocin production by coculture is widespread among plantaricin-producing Lactobacillus plantarum strains with different regulatory operons. Food Microbiol. 2013, 33, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Lee, H.J.; Chang, H.C. Identification of the agent from Lactobacillus plantarum KFRI464 that enhances bacteriocin production by Leuconostoc citreum GJ7. J. Appl. Microbiol. 2007, 103, 2504–2515. [Google Scholar] [CrossRef]
- Abitayeva, G.K.; Urazova, M.S.; Abilkhadirov, A.S.; Sarmurzina, Z.S.; Shaikhin, S.M. Characterization of a new bacteriocin-like inhibitory peptide produced by Lactobacillus sakei B-RKM 0559. Biotechnol. Lett. 2021, 43, 2243–2257. [Google Scholar] [CrossRef]
- Fu, Y.; Zhao, D.; Wang, L.; Jiang, G.; Liu, X. A broad-spectrum novel bacteriocin produced by Lactobacillus sakei in Nanjing Steamed Roast Duck: Purification, antimicrobial characteristics, and antibacterial mechanisms. Food Biosci. 2022, 50, 101995. [Google Scholar] [CrossRef]
- Contessa, C.R.; de Souza, N.B.; Gonçalo, G.B.; de Moura, C.M.; da Rosa, G.S.; Moraes, C.C. Development of active packaging based on agar-agar incorporated with bacteriocin of lactobacillus sakei. Biomolecules 2021, 11, 1869. [Google Scholar] [CrossRef] [PubMed]
- Kiczorowski, P.; Kiczorowska, B.; Samolińska, W.; Szmigielski, M.; Winiarska-Mieczan, A. Effect of fermentation of chosen vegetables on the nutrient, mineral, and biocomponent profile in human and animal nutrition. Sci. Rep. 2022, 12, 13422. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Leeuwendaal, N.K.; Stanton, C.; O’toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef]
- Lau, S.W.; Chong, A.Q.; Chin, N.L.; Talib, R.A.; Basha, R.K. Sourdough microbiome comparison and benefits. Microorganisms 2021, 9, 1355. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.H.; Huang, L.Y.; Terefe, N.S.; Augustin, M.A. Fermentation-based biotransformation of glucosinolates, phenolics and sugars in retorted broccoli puree by lactic acid bacteria. Food Chem. 2019, 286, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Mohammadi-Kouchesfahani, M.; Hamidi-Esfahani, Z.; Azizi, M.H. Isolation and identification of lactic acid bacteria with phytase activity from sourdough. Food Sci. Nutr. 2019, 7, 3700–3708. [Google Scholar] [CrossRef]
- Rosa-Sibakov, N.; Re, M.; Karsma, A.; Laitila, A.; Nordlund, E. Phytic Acid Reduction by Bioprocessing as a Tool to Improve the in Vitro Digestibility of Faba Bean Protein. J. Agric. Food Chem. 2018, 66, 10394–10399. [Google Scholar] [CrossRef] [PubMed]
- Venkatasubbaiah, R.; Rajesh, S.K. Food Processing and Fermentation Studies on Reduction of Phytic Acid in Triticum Aestivum and Sorghum Bicolor (L.). J. Microbiol. Biotechnol. Food Sci. 2020, 10, 166–169. [Google Scholar] [CrossRef]
- Onipede, G.O.; Banwo, K.; Ogunremi, O.R.; Sanni, A.; Polytechnic, T.; State, O. Influence of starter culture lactic acid bacteria on the phytic acid content of sorghum-ogi (an indigenous cereal gruel). Ann. Food Sci. Technol. 2014, 15, 121–134. [Google Scholar]
- Scheers, N.; Rossander-Hulthen, L.; Torsdottir, I.; Sandberg, A.S. Increased iron bioavailability from lactic-fermented vegetables is likely an effect of promoting the formation of ferric iron (Fe3+). Eur. J. Nutr. 2016, 55, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lai, S.; Zhou, Z.; Yang, J.; Liu, H.; Zhong, Z.; Fu, H.; Ren, Z.; Shen, L.; Cao, S.; et al. Screening and evaluation of lactic acid bacteria with probiotic potential from local Holstein raw milk. Front. Microbiol. 2022, 13, 918774. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Torres, L.; Yépez, A.; Aznar, R.; Arahal, D.R. Genomic Insights Into Five Strains of Lactobacillus plantarum With Biotechnological Potential Isolated From chicha, a Traditional Maize-Based Fermented Beverage From Northwestern Argentina. Front. Microbiol. 2019, 10, 2232. [Google Scholar] [CrossRef] [PubMed]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Sarkar, D.; Shetty, K. Fermentation-based biotransformation of bioactive phenolics and volatile compounds from cashew apple juice by select lactic acid bacteria. Process Biochem. 2017, 59, 141–149. [Google Scholar] [CrossRef]
- Llamas-Arriba, M.G.; Hernández-Alcántara, A.M.; Mohedano, M.L.; Chiva, R.; Celador-Lera, L.; Velázquez, E.; Prieto, A.; Dueñas, M.T.; Tamame, M.; López, P. Lactic acid bacteria isolated from fermented doughs in Spain produce dextrans and riboflavin. Foods 2021, 10, 2004. [Google Scholar] [CrossRef]
- Juvonen, R.; Honkapää, K.; Maina, N.H.; Shi, Q.; Viljanen, K.; Maaheimo, H.; Virkki, L.; Tenkanen, M.; Lantto, R. The impact of fermentation with exopolysaccharide producing lactic acid bacteria on rheological, chemical and sensory properties of pureed carrots (Daucus carota L.). Int. J. Food Microbiol. 2015, 207, 109–118. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented foods as a dietary source of live organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef] [PubMed]
- Pasolli, E.; De Filippis, F.; Mauriello, I.E.; Cumbo, F.; Walsh, A.M.; Leech, J.; Cotter, P.D.; Segata, N.; Ercolini, D. Large-scale genome-wide analysis links lactic acid bacteria from food with the gut microbiome. Nat. Commun. 2020, 11, 2610. [Google Scholar] [CrossRef] [PubMed]
- Mörschbächer, A.P.; Pappen, E.; Henriques, J.A.P.; Granada, C.E. Effects of probiotic supplementation on the gut microbiota composition of adults: A systematic review of randomized clinical trials. An. Acad. Bras. Cienc. 2023, 95, e20230037. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Xu, B.; Chen, C.; Li, P.; Luo, H. The function and mechanism of lactic acid bacteria in the reduction of toxic substances in food: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 5950–5963. [Google Scholar] [CrossRef] [PubMed]
- Chiocchetti, G.M.; Jadán-Piedra, C.; Monedero, V.; Zúñiga, M.; Vélez, D.; Devesa, V. Use of lactic acid bacteria and yeasts to reduce exposure to chemical food contaminants and toxicity. Crit. Rev. Food Sci. Nutr. 2019, 59, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kang, K.H.; Kim, S.H.; Lee, S.; Lee, S.H.; Ha, E.S.; Sung, N.J.; Kim, J.G.; Chung, M.J. Lactic acid bacteria directly degrade N-nitrosodimethylamine and increase the nitrite-scavenging ability in kimchi. Food Control 2017, 71, 101–109. [Google Scholar] [CrossRef]
- Nowak, A.; Kuberski, S.; Libudzisz, Z. Probiotic lactic acid bacteria detoxify N-nitrosodimethylamine. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2014, 31, 1678–1687. [Google Scholar] [CrossRef] [PubMed]
- Liao, E.; Xu, Y.; Jiang, Q.; Xia, W. Effects of inoculating autochthonous starter cultures on N-nitrosodimethylamine and its precursors formation during fermentation of Chinese traditional fermented fish. Food Chem. 2019, 271, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.B.; Ohta, Y. Binding of Mutagens by Fractions of the Cell Wall Skeleton of Lactic Acid Bacteria on Mutagens. J. Dairy Sci. 1991, 74, 1477–1481. [Google Scholar] [CrossRef]
- Stidl, R.; Sontag, G.; Koller, V.; Knasmüller, S. Binding of heterocyclic aromatic amines by lactic acid bacteria: Results of a comprehensive screening trial. Mol. Nutr. Food Res. 2008, 52, 322–329. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Mozuriene, E.; Krungleviciute, V.; Novoslavskij, A.; Santini, A.; Rozentale, I.; Juodeikiene, G.; Cizeikiene, D. The impact of lactic acid bacteria with antimicrobial properties on biodegradation of polycyclic aromatic hydrocarbons and biogenic amines in cold smoked pork sausages. Food Control 2017, 71, 285–292. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Krungleviciute, V.; Pugajeva, I.; Zadeike, D.; Juodeikiene, G.; Cizeikiene, D. The Influence of Scalded Flour, Fermentation, and Plants Belonging to Lamiaceae Family on the Wheat Bread Quality and Acrylamide Content. J. Food Sci. 2018, 83, 1560–1568. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, F.; Qi, Y.; Dziugan, P.; Bai, F.; Walczak, P.; Zhang, B. Screening of Lactobacillus strains for their ability to bind Benzo(a)pyrene and the mechanism of the process. Food Chem. Toxicol. 2013, 59, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Lo, P.R.; Yu, R.C.; Chou, C.C.; Huang, E.C. Determinations of the antimutagenic activities of several probiotic bifidobacteria under acidic and bile conditions against benzo[a]pyrene by a modified Ames test. Int. J. Food Microbiol. 2004, 93, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Niño, J.C.; Cavazos-Garduño, A.; Cantú-Cornelio, F.; Gonzalez-Cordova, A.F.; Vallejo-Cordoba, B.; Hernandez-Mendoza, A.; García, H.S. In vitro reduced availability of aflatoxin B1 and acrylamide by bonding interactions with teichoic acids from lactobacillus strains. LWT 2015, 64, 1334–1341. [Google Scholar] [CrossRef]
- Zhao, S.; Zhao, X.; Liu, Q.; Jiang, Y.; Li, Y.; Feng, W.; Xu, H.; Shao, M. Protective effect of Lactobacillus plantarum ATCC8014 on acrylamide-induced oxidative damage in rats. Appl. Biol. Chem. 2020, 63, 43. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, W.; Li, L.; Zhao, H.Y.; Sun, H.Y.; Meng, M.H.; Zhang, S.; Shao, M.L. Key role of peptidoglycan on acrylamide binding by lactic acid bacteria. Food Sci. Biotechnol. 2017, 26, 271–277. [Google Scholar] [CrossRef]
- Lahtinen, S.J.; Haskard, C.A.; Ouwenhand, A.C.; Salminen, S.J.; Ahokas, J.T. Binding of aflatoxin B1 to cell wall components of Lactobacillus rhamnosus strain GG. Food Addit. Contam. 2004, 21, 158–164. [Google Scholar] [CrossRef]
- Tohno, M.; Tanizawa, Y.; Sawada, H.; Sakamoto, M.; Ohkuma, M.; Kobayashi, H. A novel species of lactic acid bacteria, Ligilactobacillus pabuli sp. nov., isolated from alfalfa silage. Int. J. Syst. Evol. Microbiol. 2022, 72, 5587. [Google Scholar] [CrossRef]
- Chen, Y.S.; Wang, L.T.; Wu, Y.C.; Mori, K.; Tamura, T.; Chang, C.H.; Chang, Y.C.; Wu, H.C.; Yi, H.H.; Wang, P.Y. Leuconostoc litchii sp. Nov., a novel lactic acid bacterium isolated from lychee. Int. J. Syst. Evol. Microbiol. 2020, 70, 1585–1590. [Google Scholar] [CrossRef]
- Kouya, T.; Ishiyama, Y.; Ohashi, S.; Kumakubo, R.; Yamazaki, T.; Otaki, T. Philodulcilactobacillus myokoensis gen. nov., sp. nov., a fructophilic, acidophilic, and agar-phobic lactic acid bacterium isolated from fermented vegetable extracts. PLoS ONE 2023, 18, e0286677. [Google Scholar] [CrossRef]
- Terpou, A.; Mantzourani, I.; Galanis, A.; Kanellaki, M.; Bezirtzoglou, E.; Bekatorou, A.; Koutinas, A.A.; Plessas, S. Employment of l. Paracasei k5 as a novel potentially probiotic freeze-dried starter for feta-type cheese production. Microorganisms 2019, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Sarengaole, B.; Tsend-Ayush, C.; Bayinjirigala, J.; Menghebilige, B. Development technology of starter cultures using lactic acid bacteria isolated from fermented Camel milk with cholesterol lowering ability. Mong. J. Chem. 2022, 23, 38–50. [Google Scholar] [CrossRef]
- Garofalo, C.; Norici, A.; Mollo, L.; Osimani, A.; Aquilanti, L. Fermentation of Microalgal Biomass for Innovative Food Production. Microorganisms 2022, 10, 2069. [Google Scholar] [CrossRef] [PubMed]
- Parada, J.L.; De Caire, G.Z.; De Mulé, M.C.Z.; De Cano, M.M.S. Lactic acid bacteria growth promoters from Spirulina platensis. Int. J. Food Microbiol. 1998, 45, 225–228. [Google Scholar] [CrossRef]
- Çelekli, A.; Alslibi, Z.A.; Bozkurt, H.ü. Influence of incorporated Spirulina platensis on the growth of microflora and physicochemical properties of ayran as a functional food. Algal Res. 2019, 44, 101710. [Google Scholar] [CrossRef]
- Atallah, A.A.; Morsy, O.M.; Gemiel, D.G. Characterization of functional low-fat yogurt enriched with whey protein concentrate, Ca-caseinate and spirulina. Int. J. Food Prop. 2020, 23, 1678–1691. [Google Scholar] [CrossRef]
- Uwamahoro, H.P.; Li, F.; Timilsina, A.; Liu, B.; Wang, X.; Tian, Y. An Assessment of the Lactic Acid-Producing Potential of Bacterial Strains Isolated from Food Waste. Microbiol. Res. 2022, 13, 278–291. [Google Scholar] [CrossRef]
- Wolka, K.; Melaku, B. Exploring selected plant nutrient in compost prepared from food waste and cattle manure and its effect on soil properties and maize yield at Wondo Genet, Ethiopia. Environ. Syst. Res. 2015, 4, 9. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Esmail, G.A.; Arasu, M.V. Co-fermentation of food waste and municipal sludge from the Saudi Arabian environment to improve lactic acid production by Lactobacillus rhamnosus AW3 isolated from date processing waste. Sustainability 2020, 12, 6899. [Google Scholar] [CrossRef]
- Tran, Q.N.M.; Mimoto, H.; Nakasaki, K. Inoculation of lactic acid bacterium accelerates organic matter degradation during composting. Int. Biodeterior. Biodegrad. 2015, 104, 377–383. [Google Scholar] [CrossRef]
- Bühlmann, C.H.; Mickan, B.S.; Tait, S.; Batstone, D.J.; Bahri, P.A. Lactic acid production from food waste at an anaerobic digestion biorefinery: Effect of digestate recirculation and sucrose supplementation. Front. Bioeng. Biotechnol. 2023, 11, 1177739. [Google Scholar] [CrossRef]
- Sabater, C.; Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Margolles, A. Valorization of Vegetable Food Waste and By-Products Through Fermentation Processes. Front. Microbiol. 2020, 11, 581997. [Google Scholar] [CrossRef]
- Novik, G.; Meerovskaya, O.; Savich, V. Waste Degradation and Utilization by Lactic Acid Bacteria: Use of Lactic Acid Bacteria in Production of Food Additives, Bioenergy and Biogas. In Food Additives; IntechOpen: London, UK, 2017; ISBN 978-953-51-3490-9. [Google Scholar]
- Velikova, P.; Stoyanov, A.; Blagoeva, G.; Popova, L.; Petrov, K.; Gotcheva, V.; Angelov, A.; Petrova, P. Starch utilization routes in lactic acid bacteria: New insight by gene expression assay. Starch-Stärke 2016, 68, 953–960. [Google Scholar] [CrossRef]
- Tsapekos, P.; Alvarado-Morales, M.; Baladi, S.; Bosma, E.F.; Angelidaki, I. Fermentative Production of Lactic Acid as a Sustainable Approach to Valorize Household Bio-Waste. Front. Sustain. 2020, 1, 538783. [Google Scholar] [CrossRef]
- López-Gómez, J.P.; Pérez-Rivero, C.; Venus, J. Valorisation of solid biowastes: The lactic acid alternative. Process Biochem. 2020, 99, 222–235. [Google Scholar] [CrossRef]
- Song, L.; Yang, D.; Liu, R.; Liu, S.; Dai, L.; Dai, X. Microbial production of lactic acid from food waste: Latest advances, limits, and perspectives. Bioresour. Technol. 2022, 345, 126052. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Wang, Q.H.; Wang, X.M.; Ma, H.Z.; Liu, Y.Y. Process optimization for lactic acid production from kitchen waste. Harbin Gongye Daxue Xuebao/J. Harbin Inst. Technol. 2009, 41, 58–63. [Google Scholar]
- Dedenaro, G.; Costa, S.; Rugiero, I.; Pedrini, P.; Tamburini, E. Valorization of agri-food waste via fermentation: Production of L-lactic acid as a building block for the synthesis of biopolymers. Appl. Sci. 2016, 6, 379. [Google Scholar] [CrossRef]
- Kwan, T.H.; Hu, Y.; Lin, C.S.K. Valorisation of food waste via fungal hydrolysis and lactic acid fermentation with Lactobacillus casei Shirota. Bioresour. Technol. 2016, 217, 129–136. [Google Scholar] [CrossRef]
- Pleissner, D.; Demichelis, F.; Mariano, S.; Fiore, S.; Navarro Gutiérrez, I.M.; Schneider, R.; Venus, J. Direct production of lactic acid based on simultaneous saccharification and fermentation of mixed restaurant food waste. J. Clean. Prod. 2017, 143, 615–623. [Google Scholar] [CrossRef]
- Song, L.; Liu, S.; Liu, R.; Yang, D.; Dai, X. Direct lactic acid production from household food waste by lactic acid bacteria. Sci. Total Environ. 2022, 840, 156479. [Google Scholar] [CrossRef]
- Anagnostopoulou, C.; Kontogiannopoulos, K.N.; Gaspari, M.; Morlino, M.S.; Assimopoulou, A.N.; Kougias, P.G. Valorization of household food wastes to lactic acid production: A response surface methodology approach to optimize fermentation process. Chemosphere 2022, 296, 133871. [Google Scholar] [CrossRef]
- Chenebault, C.; Moscoviz, R.; Trably, E.; Escudié, R.; Percheron, B. Lactic acid production from food waste using a microbial consortium: Focus on key parameters for process upscaling and fermentation residues valorization. Bioresour. Technol. 2022, 354, 127230. [Google Scholar] [CrossRef]
- Asano, R.; Otawa, K.; Ozutsumi, Y.; Yamamoto, N.; Abdel-Mohsein, H.S.; Nakai, Y. Development and analysis of microbial characteristics of an acidulocomposting system for the treatment of garbage and cattle manure. J. Biosci. Bioeng. 2010, 110, 419–425. [Google Scholar] [CrossRef]
- Tran, Q.N.M.; Mimoto, H.; Koyama, M.; Nakasaki, K. Lactic acid bacteria modulate organic acid production during early stages of food waste composting. Sci. Total Environ. 2019, 687, 341–347. [Google Scholar] [CrossRef]
- Yang, S.Y.; Ji, K.S.; Baik, Y.H.; Kwak, W.S.; McCaskey, T.A. Lactic acid fermentation of food waste for swine feed. Bioresour. Technol. 2006, 97, 1858–1864. [Google Scholar] [CrossRef]
- Seo, J.-Y.; Song, I.-G.; Lee, D.-G.; Lee, K.-Y.; Kim, Y.-J. From food wastes into useful probiotics: Development of Lactic acid bacteria as useful probiotics for animal feed without antibiotics using food waste. J. Korea Org. Resour. Recycl. Assoc. 2006, 14, 112–120. [Google Scholar]
- Juodeikiene, G.; Cizeikiene, D.; Glasner, C.; Bartkiene, E.; Dikiy, A.; Shumilina, E.; Ilić, N.; Di Berardino, S.; Foncesa, C. Evaluation of the potential of utilizing lactic acid bacteria and dairy wastewaters for methane production. Energy Explor. Exploit. 2017, 35, 388–402. [Google Scholar] [CrossRef]
- Bohn, J.; Yüksel-Dadak, A.; Dröge, S.; König, H. Isolation of lactic acid-forming bacteria from biogas plants. J. Biotechnol. 2017, 244, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, P.; Steinigeweg, S.; Siefert, E.; Cypionka, H. Effect of Lactate and Starter Inoculum on Biogas Production from Fresh Maize and Maize Silage. Adv. Microbiol. 2017, 7, 358–376. [Google Scholar] [CrossRef]
- Menardo, S.; Balsari, P.; Tabacco, E.; Borreani, G. Effect of Conservation Time and the Addition of Lactic Acid Bacteria on the Biogas and Methane Production of Corn Stalk Silage. Bioenergy Res. 2015, 8, 1810–1823. [Google Scholar] [CrossRef]
- Vervaeren, H.; Hostyn, K.; Ghekiere, G.; Willems, B. Biological ensilage additives as pretreatment for maize to increase the biogas production. Renew. Energy 2010, 35, 2089–2093. [Google Scholar] [CrossRef]
- Vasmara, C.; Marchetti, R.; Carminati, D. Wastewater from the production of lactic acid bacteria as feedstock in anaerobic digestion. Energy 2021, 229, 120740. [Google Scholar] [CrossRef]
- Jung, S.J.; Kim, M.J.; Chae, S.W. Quality and functional characteristics of kimchi made with organically cultivated young Chinese cabbage (olgari-baechu). J. Ethn. Foods 2016, 3, 150–158. [Google Scholar] [CrossRef]
- Paladhi, A.G.; Joshi, J.T.; George, A.; Manohar, M.V.; Vallinayagam, S.; Malik, J.A. Lipase and lactic acid bacteria for biodegradation and bioremediation. In Microbes and Microbial Biotechnology for Green Remediation; Elsevier: Amsterdam, The Netherlands, 2022; pp. 265–286. [Google Scholar]
- Zhou, X.W.; Zhao, X.H. Susceptibility of nine organophosphorus pesticides in skimmed milk towards inoculated lactic acid bacteria and yogurt starters. J. Sci. Food Agric. 2015, 95, 260–266. [Google Scholar] [CrossRef]
- Ameen, F.A.; Hamdan, A.M.; El-Naggar, M.Y. Assessment of the heavy metal bioremediation efficiency of the novel marine lactic acid bacterium, Lactobacillus plantarum MF042018. Sci. Rep. 2020, 10, 314. [Google Scholar] [CrossRef]
- Syukur, S.; Yolanda, S.; Jamsari, J.; Fachrial, E. Isolation, antimicrobial activity and bioremediation of heavy metal cadmium (Cd) by using lactic acid bacteria from dadih origin lareh sago halaban, payakumbuh, west Sumatera, Indonesia. J. Chem. Pharm. Res. 2015, 7, 235–241. [Google Scholar]
- Hasan, M.S.; Islam, M.Z.; Liza, R.I.; Sarker, M.A.H.; Islam, M.A.; Harun-ur-Rashid, M. Novel Probiotic Lactic Acid Bacteria with In Vitro Bioremediation Potential of Toxic Lead and Cadmium. Curr. Microbiol. 2022, 79, 387. [Google Scholar] [CrossRef] [PubMed]
- Halttunen, T.; Salminen, S.; Tahvonen, R. Rapid removal of lead and cadmium from water by specific lactic acid bacteria. Int. J. Food Microbiol. 2007, 114, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Nduko, J.M.; Taguchi, S. Microbial Production of Biodegradable Lactate-Based Polymers and Oligomeric Building Blocks From Renewable and Waste Resources. Front. Bioeng. Biotechnol. 2021, 8, 618077. [Google Scholar] [CrossRef]
- Taib, N.A.A.B.; Rahman, M.R.; Huda, D.; Kuok, K.K.; Hamdan, S.; Bin Bakri, M.K.; Bin Julaihi, M.R.M.; Khan, A. A review on poly lactic acid (PLA) as a biodegradable polymer. Polym. Bull. 2023, 80, 1179–1213. [Google Scholar] [CrossRef]
- Kwan, T.H.; Hu, Y.; Lin, C.S.K. Techno-economic analysis of a food waste valorisation process for lactic acid, lactide and poly(lactic acid) production. J. Clean. Prod. 2018, 181, 72–87. [Google Scholar] [CrossRef]
- Nilsen-Nygaard, J.; Fernández, E.N.; Radusin, T.; Rotabakk, B.T.; Sarfraz, J.; Sharmin, N.; Sivertsvik, M.; Sone, I.; Pettersen, M.K. Current status of biobased and biodegradable food packaging materials: Impact on food quality and effect of innovative processing technologies. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1333–1380. [Google Scholar] [CrossRef]
- Kumari, S.V.G.; Pakshirajan, K.; Pugazhenthi, G. Recent advances and future prospects of cellulose, starch, chitosan, polylactic acid and polyhydroxyalkanoates for sustainable food packaging applications. Int. J. Biol. Macromol. 2022, 221, 163–182. [Google Scholar] [CrossRef]
- Yildirim, S.; Röcker, B.; Pettersen, M.K.; Nilsen-Nygaard, J.; Ayhan, Z.; Rutkaite, R.; Radusin, T.; Suminska, P.; Marcos, B.; Coma, V. Active Packaging Applications for Food. Compr. Rev. Food Sci. Food Saf. 2018, 17, 165–199. [Google Scholar] [CrossRef]
- Huang, S.; Xue, Y.; Yu, B.; Wang, L.; Zhou, C.; Ma, Y. A Review of the Recent Developments in the Bioproduction of Polylactic Acid and Its Precursors Optically Pure Lactic Acids. Molecules 2021, 26, 6446. [Google Scholar] [CrossRef]
- Bosco, F.; Cirrincione, S.; Carletto, R.; Marmo, L.; Chiesa, F.; Mazzoli, R.; Pessione, E. Pha production from cheese whey and “scotta”: Comparison between a consortium and a pure culture of leuconostoc mesenteroides. Microorganisms 2021, 9, 2426. [Google Scholar] [CrossRef]
- Aslim, B.; Çalışkan, F.; Beyatlı, Y.; Gündüz, U. Poly-β-hydroxybutyrate production by lactic acid bacteria. FEMS Microbiol. Lett. 1998, 159, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Mazzoli, R.; Bosco, F.; Mizrahi, I.; Bayer, E.A.; Pessione, E. Towards lactic acid bacteria-based biorefineries. Biotechnol. Adv. 2014, 32, 1216–1236. [Google Scholar] [CrossRef] [PubMed]
- Vijayendra, S.V.N.; Shamala, T.R. Film forming microbial biopolymers for commercial applications-A review. Crit. Rev. Biotechnol. 2014, 34, 338–357. [Google Scholar] [CrossRef]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. Technol. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef]
- Moradi, M.; Guimarães, J.T.; Sahin, S. Current applications of exopolysaccharides from lactic acid bacteria in the development of food active edible packaging. Curr. Opin. Food Sci. 2021, 40, 33–39. [Google Scholar] [CrossRef]
- Torino, M.I.; de Valdez, G.F.; Mozzi, F. Biopolymers from lactic acid bacteria. Novel applications in foods and beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef]
- Ghasemlou, M.; Khodaiyan, F.; Oromiehie, A. Physical, mechanical, barrier, and thermal properties of polyol-plasticized biodegradable edible film made from kefiran. Carbohydr. Polym. 2011, 84, 477–483. [Google Scholar] [CrossRef]
- Hassan, A.N.; Corredig, M.; Frank, J.F.; Elsoda, M. Microstructure and rheology of an acid-coagulated cheese (Karish) made with an exopolysaccharide-producing Streptococcus thermophilus strain and its exopolysaccharide non-producing genetic variant. J. Dairy Res. 2004, 71, 116–120. [Google Scholar] [CrossRef]
- Low, D.; Ahlgren, J.A.; Horne, D.; McMahon, D.J.; Oberg, C.J.; Broadbent, J.R. Role of Streptococcus thermophilus MR-1C capsular exopolysaccharide in cheese moisture retention? Appl. Environ. Microbiol. 1998, 64, 2147–2151. [Google Scholar] [CrossRef]
- Lynch, K.M.; McSweeney, P.L.H.; Arendt, E.K.; Uniacke-Lowe, T.; Galle, S.; Coffey, A. Isolation and characterisation of exopolysaccharide-producing Weissella and Lactobacillus and their application as adjunct cultures in Cheddar cheese. Int. Dairy J. 2014, 34, 125–134. [Google Scholar] [CrossRef]
- Costa, N.E.; Hannon, J.A.; Guinee, T.P.; Auty, M.A.E.; McSweeney, P.L.H.; Beresford, T.P. Effect of exopolysaccharide produced by isogenic strains of Lactococcus lactis on half-fat Cheddar cheese. J. Dairy Sci. 2010, 93, 3469–3486. [Google Scholar] [CrossRef] [PubMed]
- Dertli, E.; Toker, O.S.; Durak, M.Z.; Yilmaz, M.T.; Tatlisu, N.B.; Sagdic, O.; Cankurt, H. Development of a fermented ice-cream as influenced by in situ exopolysaccharide production: Rheological, molecular, microstructural and sensory characterization. Carbohydr. Polym. 2016, 136, 427–440. [Google Scholar] [CrossRef]
- Li, D.; Li, J.; Zhao, F.; Wang, G.; Qin, Q.; Hao, Y. The influence of fermentation condition on production and molecular mass of EPS produced by Streptococcus thermophilus 05-34 in milk-based medium. Food Chem. 2016, 197, 367–372. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rachwał, K.; Gustaw, K. Lactic Acid Bacteria in Sustainable Food Production. Sustainability 2024, 16, 3362. https://doi.org/10.3390/su16083362
AMA Style
Rachwał K, Gustaw K. Lactic Acid Bacteria in Sustainable Food Production. Sustainability. 2024; 16(8):3362. https://doi.org/10.3390/su16083362
Chicago/Turabian StyleRachwał, Kamila, and Klaudia Gustaw. 2024. "Lactic Acid Bacteria in Sustainable Food Production" Sustainability 16, no. 8: 3362. https://doi.org/10.3390/su16083362
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.