
Mitochondrial DNA and Electron Transport Chain Protein Levels Are Altered in Peripheral Nerve Tissues from Donors with HIV Sensory Neuropathy: A Pilot Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. mtDNA Copy Number and Common Deletion Relative Abundance (RA) in DSP
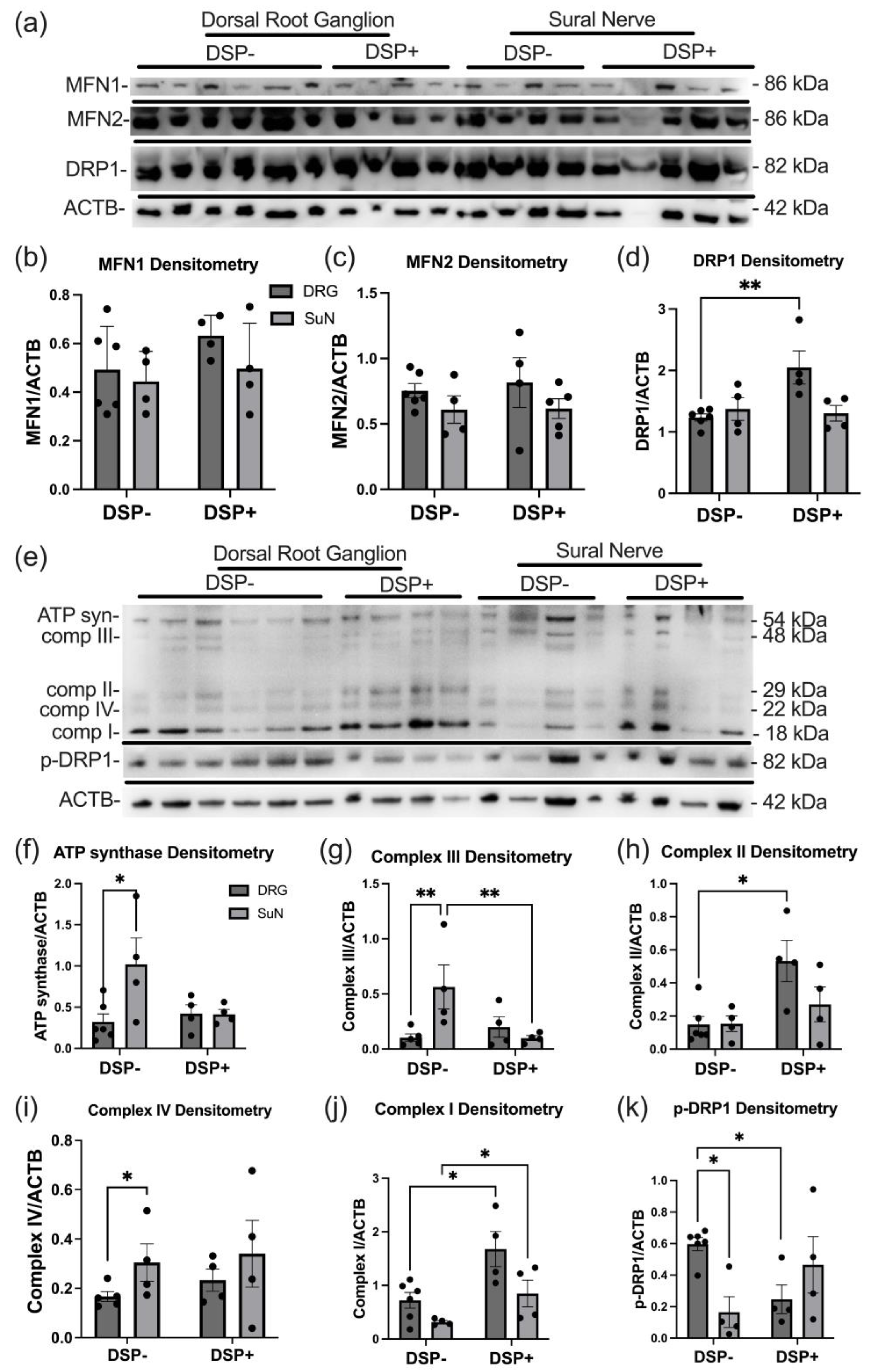
2.2. Mitochondrial Dynamics and ETC Protein Expression Levels Are Altered in DSP
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Phenotype Definitions
4.3. Quantification of mtDNA and the Proportion of Mitochondria Carrying the Common Deletion
4.4. Immunoblot Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collaboration, A.T.C. Life expectancy of individuals on combination antiretroviral therapy in high-income countries: A collaborative analysis of 14 cohort studies. Lancet 2008, 372, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Autenrieth, C.S.; Beck, E.J.; Stelzle, D.; Mallouris, C.; Mahy, M.; Ghys, P. Global and regional trends of people living with HIV aged 50 and over: Estimates and projections for 2000–2020. PLoS ONE 2018, 13, e0207005. [Google Scholar] [CrossRef] [PubMed]
- Tavasoli, A.; Gelman, B.B.; Marra, C.M.; Clifford, D.B.; Iudicello, J.E.; Rubin, L.H.; Letendre, S.L.; Tang, B.; Ellis, R.J. Increasing Neuroinflammation Relates to Increasing Neurodegeneration in People with HIV. Viruses 2023, 15, 1835. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Nelson, A.M. HIV and the spectrum of human disease. J. Pathol. 2015, 235, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Keswani, S.C.; Pardo, C.A.; Cherry, C.L.; Hoke, A.; McArthur, J.C. HIV-associated sensory neuropathies. AIDS 2002, 16, 2105–2117. [Google Scholar] [CrossRef] [PubMed]
- McArthur, J.C.; Brew, B.J.; Nath, A. Neurological complications of HIV infection. Lancet Neurol. 2005, 4, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Cornblath, D.R.; Hoke, A. Recent advances in HIV neuropathy. Curr. Opin. Neurol. 2006, 19, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Vafaei, A.A.; Safakhah, H.A.; Jafari, S.; Tavasoli, A.; Rashidy-Pour, A.; Ghanbari, A.; Seyedinia, S.A.; Tarahomi, P. Role of Cannabinoid Receptors in Crocin-Induced Hypoalgesia in Neuropathic Pain in Rats. J. Exp. Pharmacol. 2020, 12, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Jazebi, N.; Evans, C.; Kadaru, H.S.; Kompella, D.; Raji, M.; Fang, F.; Pappolla, M.; Tang, S.J.; Chung, J.M.; Hammock, B.; et al. HIV-related Neuropathy: Pathophysiology, Treatment and Challenges. J. Neurol. Exp. Neurosci. 2021, 7, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.Q.; Beal, M.F.; Manfredi, G. The role of mitochondria in inherited neurodegenerative diseases. J. Neurochem. 2006, 97, 1659–1675. [Google Scholar] [CrossRef]
- Milligan, E.D.; O’Connor, K.A.; Nguyen, K.T.; Armstrong, C.B.; Twining, C.; Gaykema, R.P.; Holguin, A.; Martin, D.; Maier, S.F.; Watkins, L.R. Intrathecal HIV-1 envelope glycoprotein gp120 induces enhanced pain states mediated by spinal cord proinflammatory cytokines. J. Neurosci. 2001, 21, 2808–2819. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, E.R.; Hulgan, T.; Ellis, R.J.; Samuels, D.C.; Ritchie, M.D.; Haas, D.W.; Kallianpur, A.R.; Bloss, C.S.; Clifford, D.B.; Collier, A.C.; et al. Mitochondrial DNA variation and HIV-associated sensory neuropathy in CHARTER. J. Neurovirol. 2012, 18, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Rozzi, S.J.; Avdoshina, V.; Fields, J.A.; Trejo, M.; Ton, H.T.; Ahern, G.P.; Mocchetti, I. Human Immunodeficiency Virus Promotes Mitochondrial Toxicity. Neurotox. Res. 2017, 32, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Ganta, K.K.; Chaubey, B. Mitochondrial dysfunctions in HIV infection and antiviral drug treatment. Expert Opin. Drug Metab. Toxicol. 2019, 15, 1043–1052. [Google Scholar] [CrossRef]
- Fields, J.A.; Ellis, R.J. HIV in the cART era and the mitochondrial: Immune interface in the CNS. Int. Rev. Neurobiol. 2019, 145, 29–65. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, H.C.; Chen, W.; Borzan, J.; Mankowski, J.L.; Hoke, A. Mitochondrial dysfunction in distal axons contributes to human immunodeficiency virus sensory neuropathy. Ann. Neurol. 2011, 69, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Fields, J.A.; Swinton, M.K.; Carson, A.; Soontornniyomkij, B.; Lindsay, C.; Han, M.M.; Frizzi, K.; Sambhwani, S.; Murphy, A.; Achim, C.L.; et al. Tenofovir disoproxil fumarate induces peripheral neuropathy and alters inflammation and mitochondrial biogenesis in the brains of mice. Sci. Rep. 2019, 9, 17158. [Google Scholar] [CrossRef] [PubMed]
- Youle, M.; Osio, M.; Group, A.S. A double-blind, parallel-group, placebo-controlled, multicentre study of acetyl L-carnitine in the symptomatic treatment of antiretroviral toxic neuropathy in patients with HIV-1 infection. HIV Med. 2007, 8, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Kamerman, P.R.; Moss, P.J.; Weber, J.; Wallace, V.C.; Rice, A.S.; Huang, W. Pathogenesis of HIV-associated sensory neuropathy: Evidence from in vivo and in vitro experimental models. J. Peripher. Nerv. Syst. 2012, 17, 19–31. [Google Scholar] [CrossRef]
- Lipke, K.; Kubis-Kubiak, A.; Piwowar, A. The Influence of Nucleoside Reverse Transcriptase Inhibitors on Mitochondrial Activity, Lipid Content, and Fatty-Acid-Binding Protein Levels in Microglial HMC3 Cells. Pharmaceuticals 2023, 16, 1661. [Google Scholar] [CrossRef]
- Selvaraj, S.; Ghebremichael, M.; Li, M.; Foli, Y.; Langs-Barlow, A.; Ogbuagu, A.; Barakat, L.; Tubridy, E.; Edifor, R.; Lam, W.; et al. Antiretroviral therapy-induced mitochondrial toxicity: Potential mechanisms beyond polymerase-γ inhibition. Clin. Pharmacol. Ther. 2014, 96, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Roca-Bayerri, C.; Robertson, F.; Pyle, A.; Hudson, G.; Payne, B.A.I. Mitochondrial DNA Damage and Brain Aging in Human Immunodeficiency Virus. Clin. Infect. Dis. 2021, 73, e466–e473. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.F.; Millet, A.R.; Tigano, M.; Dubois, S.M.; Crimmins, H.; Babin, L.; Charpentier, M.; Piganeau, M.; Brunet, E.; Sfeir, A. Single-Molecule Analysis of mtDNA Replication Uncovers the Basis of the Common Deletion. Mol. Cell 2017, 65, 527–538.e6. [Google Scholar] [CrossRef] [PubMed]
- Cortopassi, G.A.; Arnheim, N. Detection of a specific mitochondrial DNA deletion in tissues of older humans. Nucleic Acids Res. 1990, 18, 6927–6933. [Google Scholar] [CrossRef] [PubMed]
- Linnane, A.W.; Baumer, A.; Maxwell, R.J.; Preston, H.; Zhang, C.F.; Marzuki, S. Mitochondrial gene mutation: The ageing process and degenerative diseases. Biochem. Int. 1990, 22, 1067–1076. [Google Scholar] [PubMed]
- Yen, T.C.; Su, J.H.; King, K.L.; Wei, Y.H. Ageing-associated 5 kb deletion in human liver mitochondrial DNA. Biochem. Biophys. Res. Commun. 1991, 178, 124–131. [Google Scholar] [CrossRef]
- Nissanka, N.; Moraes, C.T. Mitochondrial DNA heteroplasmy in disease and targeted nuclease-based therapeutic approaches. EMBO Rep. 2020, 21, e49612. [Google Scholar] [CrossRef] [PubMed]
- Sciacco, M.; Bonilla, E.; Schon, E.A.; DiMauro, S.; Moraes, C.T. Distribution of wild-type and common deletion forms of mtDNA in normal and respiration-deficient muscle fibers from patients with mitochondrial myopathy. Hum. Mol. Genet. 1994, 3, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Guerra, F.; Calvani, R.; Coelho-Júnior, H.J.; Leeuwenburgh, C.; Bucci, C.; Marzetti, E. The contribution of mitochondrial DNA alterations to aging, cancer, and neurodegeneration. Exp. Gerontol. 2023, 178, 112203. [Google Scholar] [CrossRef]
- Roda, R.H.; Bargiela, D.; Chen, W.; Perry, K.; Ellis, R.J.; Clifford, D.B.; Bharti, A.; Kallianpur, A.R.; Oliveira, M.F.; Diaz, M.M.; et al. Large Mitochondrial DNA Deletions in HIV Sensory Neuropathy. Neurology 2021, 97, e156–e165. [Google Scholar] [CrossRef]
- Motwani, L.; Asif, N.; Patel, A.; Vedantam, D.; Poman, D.S. Neuropathy in Human Immunodeficiency Virus: A Review of the Underlying Pathogenesis and Treatment. Cureus 2022, 14, e25905. [Google Scholar] [CrossRef]
- Mandal, A.; Drerup, C.M. Axonal Transport and Mitochondrial Function in Neurons. Front. Cell Neurosci. 2019, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- Smith, J. RAWP revisited. Br. Med. J. (Clin. Res. Ed.) 1987, 295, 1015. [Google Scholar] [CrossRef] [PubMed]
- Seager, R.; Lee, L.; Henley, J.M.; Wilkinson, K.A. Mechanisms and roles of mitochondrial localisation and dynamics in neuronal function. Neuronal Signal. 2020, 4, NS20200008. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Zhu, J.; Song, S.; Huang, Y.; Zhang, W.; Sun, Y.; Hao, J.; Yang, X.; Gao, Q.; et al. Neuroinflammation-mediated mitochondrial dysregulation involved in postoperative cognitive dysfunction. Free Radic. Biol. Med. 2022, 178, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Ho, D.H.; Je, A.R.; Lee, H.; Son, I.; Kweon, H.S.; Kim, H.G.; Seol, W. LRRK2 Kinase Activity Induces Mitochondrial Fission in Microglia via Drp1 and Modulates Neuroinflammation. Exp. Neurobiol. 2018, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Nadalutti, C.A.; Ayala-Peña, S.; Santos, J.H. Mitochondrial DNA damage as driver of cellular outcomes. Am. J. Physiol.-Cell Physiol. 2022, 322, C136–C150. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Matute, P.; Pérez-Martínez, L.; Blanco, J.R.; Oteo, J.A. Role of Mitochondria in HIV Infection and Associated Metabolic Disorders: Focus on Nonalcoholic Fatty Liver Disease and Lipodystrophy Syndrome. Oxidative Med. Cell. Longev. 2013, 2013, 493413. [Google Scholar] [CrossRef]
- Roda, R.H.; Hoke, A. Chapter Four-Mitochondrial dysfunction in HIV-induced peripheral neuropathy. In International Review of Neurobiology; Fernyhough, P., Calcutt, N.A., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 145, pp. 67–82. [Google Scholar]
- Fields, J.A.; Serger, E.; Campos, S.; Divakaruni, A.S.; Kim, C.; Smith, K.; Trejo, M.; Adame, A.; Spencer, B.; Rockenstein, E.; et al. HIV alters neuronal mitochondrial fission/fusion in the brain during HIV-associated neurocognitive disorders. Neurobiol. Dis. 2016, 86, 154–169. [Google Scholar] [CrossRef]
- Ferrari, L.F.; Chum, A.; Bogen, O.; Reichling, D.B.; Levine, J.D. Role of Drp1, a Key Mitochondrial Fission Protein, in Neuropathic Pain. J. Neurosci. 2011, 31, 11404–11410. [Google Scholar] [CrossRef]
- Chang, Y.-W.; Tony Yang, T.; Chen, M.-C.; Liaw, Y.g.; Yin, C.-F.; Lin-Yan, X.-Q.; Huang, T.-Y.; Hou, J.-T.; Hung, Y.-H.; Hsu, C.-L.; et al. Spatial and temporal dynamics of ATP synthase from mitochondria toward the cell surface. Commun. Biol. 2023, 6, 427. [Google Scholar] [CrossRef] [PubMed]
- Pardo, C.A.; McArthur, J.C.; Griffin, J.W. HIV neuropathy: Insights in the pathology of HIV peripheral nerve disease. J. Peripher. Nerv. Syst. 2001, 6, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, N.; Zhang, X.; Horng, T. Mitochondrial metabolism regulates macrophage biology. J. Biol. Chem. 2021, 297, 100904. [Google Scholar] [CrossRef] [PubMed]
- Schank, M.; Zhao, J.; Moorman, J.P.; Yao, Z.Q. The Impact of HIV- and ART-Induced Mitochondrial Dysfunction in Cellular Senescence and Aging. Cells 2021, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.M.; Keltner, J.R.; Simmons, A.N.; Franklin, D.; Moore, R.C.; Clifford, D.; Collier, A.C.; Gelman, B.B.; Marra, P.D.C.; McCutchan, J.A.; et al. Paresthesia Predicts Increased Risk of Distal Neuropathic Pain in Older People with HIV-Associated Sensory Polyneuropathy. Pain Med. 2021, 22, 1850–1856. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, B.K.; McMurtray, A.; Davis, J.; Valcour, V.; Watters, M.R.; Shiramizu, B.; Chow, D.C.; Kallianpur, K.; Shikuma, C.M. Incident neuropathy in HIV-infected patients on HAART. AIDS Res. Hum. Retroviruses 2010, 26, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Strain, M.C.; Lada, S.M.; Luong, T.; Rought, S.E.; Gianella, S.; Terry, V.H.; Spina, C.A.; Woelk, C.H.; Richman, D.D. Highly precise measurement of HIV DNA by droplet digital PCR. PLoS ONE 2013, 8, e55943. [Google Scholar] [CrossRef]
- Perez-Santiago, J.; Schrier, R.D.; de Oliveira, M.F.; Gianella, S.; Var, S.R.; Day, T.R.; Ramirez-Gaona, M.; Suben, J.D.; Murrell, B.; Massanella, M.; et al. Cell-free mitochondrial DNA in CSF is associated with early viral rebound, inflammation, and severity of neurocognitive deficits in HIV infection. J. Neurovirol. 2016, 22, 191–200. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sensory Peripheral Neuropathy | Gender (M/F) | Age | ART (Current/Past) | D-Drugs * (Current/Past) | Highly Active ART (Yes/No) | Nadir CD4 | PMI |
|---|---|---|---|---|---|---|---|
| No | 4/2 | 42 ± 10.1 | 4/2 | 1/4 | 5/1 | 14 (2–23) | 17.2 ± 15.2 |
| Yes | 4/1 | 48 ± 12.3 | 4/1 | 2/3 | 3/2 | 8 (7–11) | 24 ± 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boustani, A.; Kulbe, J.R.; Andalibi, M.S.; Pérez-Santiago, J.; Mehta, S.R.; Ellis, R.J.; Fields, J.A. Mitochondrial DNA and Electron Transport Chain Protein Levels Are Altered in Peripheral Nerve Tissues from Donors with HIV Sensory Neuropathy: A Pilot Study. Int. J. Mol. Sci. 2024, 25, 4732. https://doi.org/10.3390/ijms25094732
Boustani A, Kulbe JR, Andalibi MS, Pérez-Santiago J, Mehta SR, Ellis RJ, Fields JA. Mitochondrial DNA and Electron Transport Chain Protein Levels Are Altered in Peripheral Nerve Tissues from Donors with HIV Sensory Neuropathy: A Pilot Study. International Journal of Molecular Sciences. 2024; 25(9):4732. https://doi.org/10.3390/ijms25094732
Chicago/Turabian StyleBoustani, Ali, Jacqueline R. Kulbe, Mohammadsobhan Sheikh Andalibi, Josué Pérez-Santiago, Sanjay R. Mehta, Ronald J. Ellis, and Jerel Adam Fields. 2024. "Mitochondrial DNA and Electron Transport Chain Protein Levels Are Altered in Peripheral Nerve Tissues from Donors with HIV Sensory Neuropathy: A Pilot Study" International Journal of Molecular Sciences 25, no. 9: 4732. https://doi.org/10.3390/ijms25094732