

RadPhysBio: A Radiobiological Database for the Prediction of Cell Survival upon Exposure to Ionizing Radiation

 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Results
2.1. Experimental Data Calculations
2.2. Model Predictions
3. Discussion
4. Methods
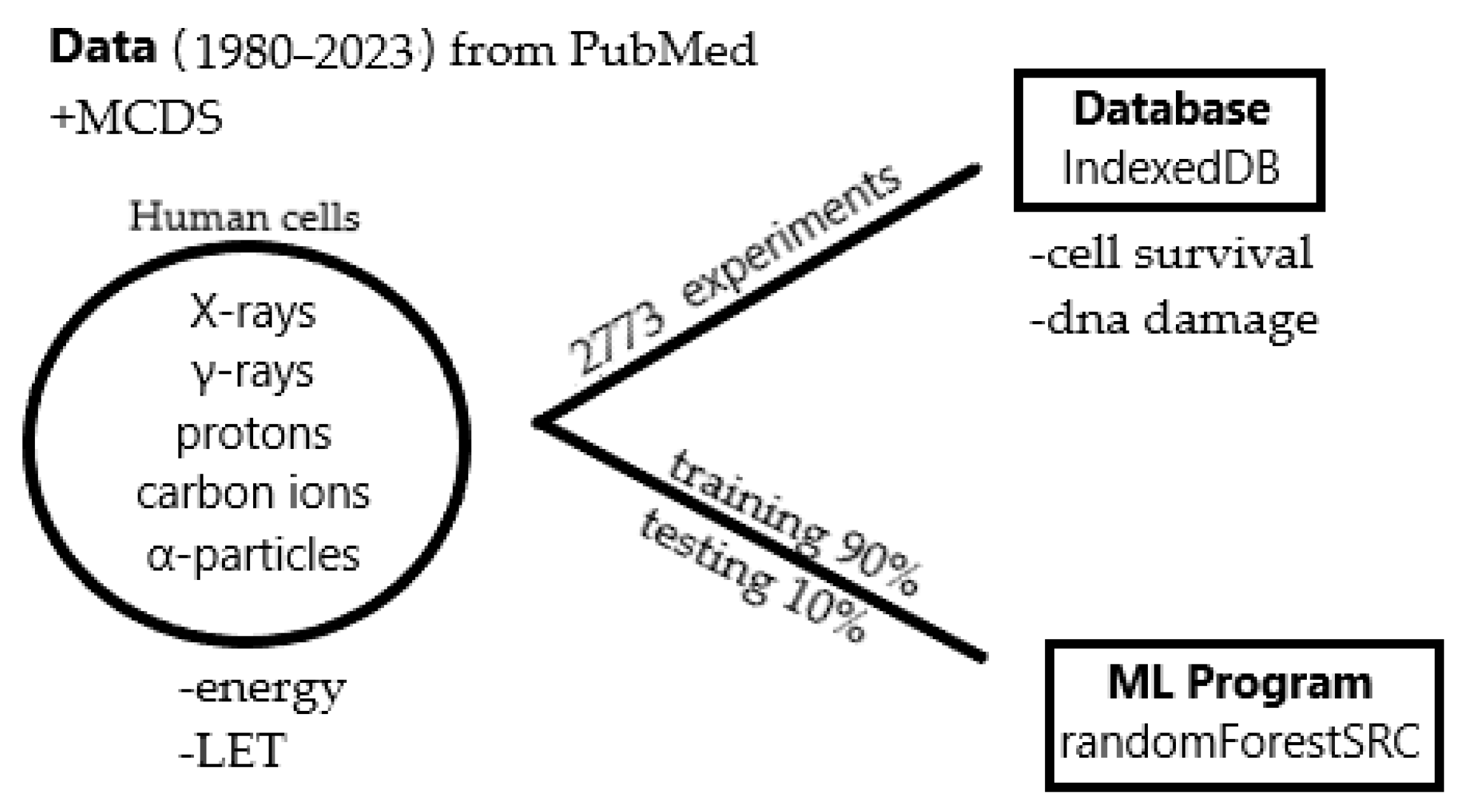
4.1. Data Collection
4.2. DNA Repair Fitting Model
4.3. Database Development
4.4. Machine Learning and Prediction Model
- -
- mtry: Number of variables to possibly split at each node.
- -
- nodesize: Minimum size of the terminal node.
- -
- Performance (qq plots).
- -
- Variable importance [51].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manti, L.; Braselmann, H.; Calabrese, M.L.; Massa, R.; Pugliese, M.; Scampoli, P.; Sicignano, G.; Grossi, G. Effects of Modulated Microwave Radiation at Cellular Telephone Frequency (1.95 Ghz) on X-ray-Induced Chromosome Aberrations in Human Lymphocytes in Vitro. Radiat. Res. 2008, 169, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Mavragani, I.V.; Nikitaki, Z.; Kalospyros, S.A.; Georgakilas, A.G. Ionizing Radiation and Complex DNA Damage: From Prediction to Detection Challenges and Biological Significance. Cancers 2019, 11, 1789. [Google Scholar] [CrossRef] [PubMed]
- Manti, L.; Durante, M.; Grossi, G.; Ortenzia, O.; Pugliese, M.; Scampoli, P.; Gialanella, G. Measurements of Metaphase and Interphase Chromosome Aberrations Transmitted through Early Cell Replication Rounds in Human Lymphocytes Exposed to Low-LET Protons and High-Let 12C Ions. Mutat. Res. 2006, 596, 151–165. [Google Scholar] [CrossRef]
- Friedland, W.; Schmitt, E.; Kundrát, P.; Dingfelder, M.; Baiocco, G.; Barbieri, S.; Ottolenghi, A. Comprehensive Track-Structure Based Evaluation of DNA Damage by Light Ions from Radiotherapy-Relevant Energies Down to Stopping. Sci. Rep. 2017, 7, 45161. [Google Scholar] [CrossRef] [PubMed]
- Moeini, H.; Mokari, M. DNA Damage and Microdosimetry for Carbon Ions: Track Structure Simulations as the Key to Quantitative Modeling of Radiation-Induced Damage. Med. Phys. 2022, 49, 4823–4836. [Google Scholar] [CrossRef] [PubMed]
- Mokari, M.; Moeini, H.; Soleimani, M. Calculation of Microdosimetric Spectra for Protons Using Geant4-DNA and a Μ-Randomness Sampling Algorithm for the Nanometric Structures. Int. J. Radiat. Biol. 2021, 97, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Nikjoo, H.; Taleei, R.; Liamsuwan, T.; Liljequist, D.; Emfietzoglou, D. Perspectives in Radiation Biophysics: From Radiation Track Structure Simulation to Mechanistic Models of DNA Damage and Repair. Radiat. Phys. Chem. 2016, 128, 3–10. [Google Scholar] [CrossRef]
- Hawkins, R.B. A Microdosimetric-Kinetic Model of Cell Death from Exposure to Ionizing Radiation of Any Let, with Experimental and Clinical Applications. Int. J. Radiat. Biol. 1996, 69, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Scholz, M.; Kraft, G. The Physical and Radiobiological Basis of the Local Effect Model: A Response to the Commentary by R. Katz. Radiat. Res. 2004, 161, 612–620. [Google Scholar] [CrossRef]
- McMahon, S.J. The Linear Quadratic Model: Usage, Interpretation and Challenges. Phys. Med. Biol. 2018, 64, 01TR01. [Google Scholar] [CrossRef]
- Papakonstantinou, D.; Zanni, V.; Nikitaki, Z.; Vasileiou, C.; Kousouris, K.; Georgakilas, A.G. Using Machine Learning Techniques for Asserting Cellular Damage Induced by High-LET Particle Radiation. Radiation 2021, 1, 45–64. [Google Scholar] [CrossRef]
- Friedrich, T.; Pfuhl, T.; Scholz, M. Update of the Particle Irradiation Data Ensemble (PIDE) for Cell Survival. J. Radiat. Res. 2021, 62, 645–655. [Google Scholar] [CrossRef]
- Semenenko, V.A.; Stewart, R.D. Fast Monte Carlo Simulation of DNA Damage Formed by Electrons and Light Ions. Phys. Med. Biol. 2006, 51, 1693–1706. [Google Scholar] [CrossRef]
- Emami, B.; Woloschak, G.; Small, W., Jr. Beyond the Linear Quadratic Model: Intraoperative Radiotherapy and Normal Tissue Tolerance. Transl. Cancer Res. 2015, 4, 140–147. [Google Scholar]
- Wilkens, J.J.; Oelfke, U. A Phenomenological Model for the Relative Biological Effectiveness in Therapeutic Proton Beams. Phys. Med. Biol. 2004, 49, 2811–2825. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Goodhead, D.T. Dependence and Independence of Survival Parameters on Linear Energy Transfer in Cells and Tissues. J. Radiat. Res. 2016, 57, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Mokari, M.; Moeini, H.; Farazmand, S. Computational Modeling and a Geant4-DNA Study of the Rejoining of Direct and Indirect DNA Damage Induced by Low Energy Electrons and Carbon Ions. Int. J. Radiat. Biol. 2023, 99, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Bagging Predictors. Mach. Learn. 1996, 24, 123–140. [Google Scholar] [CrossRef]
- Breiman, L. Out-of-Bag Estimation; Statistics Department, University of California Berkeley: Berkeley, CA, USA, 1996. [Google Scholar]
- Jones, L.; Hoban, P.; Metcalfe, P. The Use of the Linear Quadratic Model in Radiotherapy: A Review. Australas. Phys. Eng. Sci. Med. 2001, 24, 132–146. [Google Scholar] [CrossRef]
- Pawlik, T.M.; Keyomarsi, K. Role of Cell Cycle in Mediating Sensitivity to Radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 928–942. [Google Scholar] [CrossRef]
- Lind, B.K.; Brahme, A. The Radiation Response of Heterogeneous Tumors. Phys. Med. 2007, 23, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.J. The Linear-Quadratic Model Is an Appropriate Methodology for Determining Isoeffective Doses at Large Doses per Fraction. Semin. Radiat. Oncol. 2008, 18, 234–239. [Google Scholar] [CrossRef]
- Tobias, C.A. The Repair-Misrepair Model in Radiobiology: Comparison to Other Models. Radiat. Res. Suppl. 1985, 8, S77–S95. [Google Scholar] [CrossRef] [PubMed]
- Curtis, S.B. Lethal and Potentially Lethal Lesions Induced by Radiation—A Unified Repair Model. Radiat. Res. 1986, 106, 252–270. [Google Scholar] [CrossRef] [PubMed]
- Goodhead, D.T.; Belli, M.; Mill, A.J.; Bance, D.A.; Allen, L.A.; Hall, S.C.; Ianzani, F.; Simone, G.; Stevens, D.L.; Stretch, A.; et al. Direct Comparison between Protons and Alpha-Particles of the Same LET: I. Irradiation Methods and Inactivation of Asynchronous V79, HeLa and C3H 10T1/2 cells. Int. J. Radiat. Biol. 1992, 61, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Tsao, D.; Kalogerinis, P.; Tabrizi, I.; Dingfelder, M.; Stewart, R.D.; Georgakilas, A.G. Induction and Processing of Oxidative Clustered DNA Lesions in 56Fe-Ion-Irradiated Human Monocytes. Radiat. Res. 2007, 168, 87–97. [Google Scholar] [CrossRef]
- Friedland, W.; Dingfelder, M.; Jacob, P.; Paretzke, H.G. Calculated DNA Double-Strand Break and Fragmentation Yields after Irradiation with He ions. Radiat. Phys. Chem. 2005, 72, 279–286. [Google Scholar] [CrossRef]
- Baiocco, G.; Barbieri, S.; Babini, G.; Morini, J.; Alloni, D.; Friedland, W.; Kundrát, P.; Schmitt, E.; Puchalska, M.; Sihver, L.; et al. The Origin of Neutron Biological Effectiveness as a Function of Energy. Nat.-Sci. Rep. 2016, 6, 34033. [Google Scholar] [CrossRef] [PubMed]
- Hada, M.; Georgakilas, A.G. Formation of Clustered DNA Damage after High-LET Irradiation: A review. J. Radiat. Res. 2008, 49, 203–210. [Google Scholar] [CrossRef]
- Chatzipapas, K.P.; Papadimitroulas, P.; Emfietzoglou, D.; Kalospyros, S.A.; Hada, M.; Georgakilas, A.G.; Kagadis, G.C. Ionizing Radiation and Complex DNA Damage: Quantifying the Radiobiological Damage Using Monte Carlo Simulations. Cancers 2020, 12, 799. [Google Scholar] [CrossRef]
- Noubissi, F.K.; McBride, A.A.; Leppert, H.G.; Millet, L.J.; Wang, X.; Davern, S.M. Detection and Quantification of γ-H2AX Using a Dissociation Enhanced Lanthanide Fluorescence Immunoassay. Sci. Rep. 2021, 11, 8945. [Google Scholar] [CrossRef]
- Hernández, L.; Terradas, M.; Martín, M.; Tusell, L.; Genescà, A. Highly Sensitive Automated Method for DNA Damage Assessment: Gamma-H2AX Foci Counting and Cell Cycle Sorting. Int. J. Mol. Sci. 2013, 14, 15810–15826. [Google Scholar] [CrossRef] [PubMed]
- Nikitaki, Z.; Pariset, E.; Sudar, D.; Costes, S.V.; Georgakilas, A.G. In Situ Detection of Complex DNA Damage Using Microscopy: A Rough Road Ahead. Cancers 2020, 12, 3288. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.C.; Elkashty, O.; Ramamoorthi, M.; Trinh, N.; Liu, Y.; Sunavala-Dossabhoy, G.; Pranzatelli, T.; Michael, D.G.; Chivasso, C.; Perret, J.; et al. Cross-Contamination of the Human Salivary Gland HSG Cell Line with HeLa Cells: A STR Analysis Study. Oral Dis. 2018, 24, 1477–1483. [Google Scholar] [CrossRef]
- Available online: https://www.atcc.org/the-science/authentication/reclassified-cell-lines (accessed on 10 April 2024).
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res 2022, 50, D20–D26. [Google Scholar] [CrossRef] [PubMed]
- Jones, B. A Simpler Energy Transfer Efficiency Model to Predict Relative Biological Effect for Protons and Heavier Ions. Front. Oncol. 2015, 5, 184. [Google Scholar] [CrossRef] [PubMed]
- Rohatgi, A. WebPlotDigitizer, 2022.
- Van Rossum, G.A.D.; Fred, L. Python 3 Reference Manual; CreateSpace: North Charleston, SC, USA, 2009. [Google Scholar]
- Raybaut, P. Spyder-Documentation; Pythonhosted. Org: Paris, France, 2009. [Google Scholar]
- Anaconda Software Distribution; Anaconda Documentation; Anaconda Inc.: Austin, TX, USA, 2020.
- Sharma-Kuinkel, B.K.; Rude, T.H.; Fowler, V.G., Jr. Pulse Field Gel Electrophoresis. Methods Mol. Biol. 2016, 1373, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Rothkamm, K.; Barnard, S.; Moquet, J.; Ellender, M.; Rana, Z.; Burdak-Rothkamm, S. DNA Damage Foci: Meaning and Significance. Environ. Mol. Mutagen. 2015, 56, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.J.; Stewart, R.D.; Semenenko, V.A.; Sandison, G.A. Combined Use of Monte Carlo DNA Damage Simulations and Deterministic Repair Models to Examine Putative Mechanisms of Cell Killing. Radiat. Res. 2008, 169, 447–459. [Google Scholar] [CrossRef]
- Stewart, R.D.; Yu, V.K.; Georgakilas, A.G.; Koumenis, C.; Park, J.H.; Carlson, D.J. Effects of Radiation Quality and Oxygen on Clustered DNA Lesions and Cell Death. Radiat. Res. 2011, 176, 587–602. [Google Scholar] [CrossRef]
- Taleei, R.; Nikjoo, H. The Non-Homologous End-Joining (NHEJ) Pathway for the Repair of DNA Double-Strand Breaks: I. A Mathematical Model. Radiat. Res. 2013, 179, 530–539. [Google Scholar] [CrossRef] [PubMed]
- MATLAB, 2020a; The Math Works, Inc.: Natick, MA, USA, 2020.
- McNamara, A.L.; Schuemann, J.; Paganetti, H. A Phenomenological Relative Biological Effectiveness (Rbe) Model for Proton Therapy Based on All Published in Vitro Cell Survival Data. Phys. Med. Biol. 2015, 60, 8399–8416. [Google Scholar] [CrossRef] [PubMed]
- Ishwaran, H.; Kogalur, U.B. Random Survival Forests for R. R News 2007, 7, 25–31. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Goldberg, D.E. Genetic Algorithms in Search, Optimization and Machine Learning; Addison-Wesley Publishing Company, Inc.: San Francisco, CA, USA, 1989. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Range of α/β |
|---|---|
| kidney | Late-responding tissue 3–5 |
| lung | |
| bladder | |
| bone | Early-responding tissue 7–10 |
| head and neck | |
| colon | |
| skin |
| Column | Content |
|---|---|
| #ExpID | Running number labelling the database entry |
| PMID | Running number labelling the publication |
| #Exp | Running number labelling the irradiation experiments within a publication |
| CellLine | Name of the irradiated cell line |
| Tissue | Name of cell tissue |
| CellClass | Tumor cells (t) or normal cells (n) |
| CellCycle | Cell cycle phase (phases are provided explicitly in each case, or noted by ‘a’ for ‘asynchronous’ cell lines) |
| Source | Type of radioactive source |
| Energy (MeV) | Specific radiation energy |
| RBE | Relative Biological Effectiveness |
| LET (keV/μm) | Linear Energy Transfer in water |
| IrradiationConditions | Mono-energetic radiation (‘m’), or spread-out Bragg peak (‘s’) |
| DoseRate (Gy/min) | Quantity of radiation delivered per minute of time |
| α | Linear coefficient of the LQ model (in Gy−1) for response to radiation, as given in the corresponding publication, or else from fitting to raw data |
| β | Quadratic coefficient of the LQ model (in Gy−2) for response to radiation, as given in the corresponding publication, or else from fitting to raw data |
| DSBs/(Gbp*Gy) | Number of initial DSBs per Gbp per Gy, as given in the corresponding publication |
| nonDSBClusters/(Gbp*Gy) | Number of initial non-DSB clusters per Gbp per Gy, as given in the corresponding publication |
| DSBs_1%O2 | Number of initial DSBs per Gbp per Gy, calculated by the MCDS simulation code, for the specific energy of each experiment and 1% oxygen concentration in the cell |
| Other_1%O2 | Number of initial non-DSB clusters per Gbp per Gy, calculated by the MCDS simulation code, for the specific energy of each experiment and 1% oxygen concentration in the cell |
| DSBs_20%O2 | Number of initial DSBs per Gbp per Gy, calculated by the MCDS simulation code, for the specific energy of each experiment and 20% oxygen concentration in the cell |
| Other_20%O2 | Number of initial non-DSB clusters per Gbp per Gy, calculated by the MCDS simulation code, for the specific energy of each experiment and 20% oxygen concentration in the cell |
| 1keV_DSBs_1%O2 | Number of initial DSBs per Gbp per Gy, calculated by the MCDS simulation code, for 1 keV energy and 1% oxygen concentration in the cell |
| 1keV_Other_1%O2 | Number of initial non-DSB clusters per Gbp per Gy, calculated by the MCDS simulation code, for 1 keV energy and 1% oxygen concentration in the cell |
| 1keV_DSBs_20%O2 | Number of initial DSBs per Gbp per Gy, calculated by the MCDS simulation code, for 1 keV energy and 20% oxygen concentration in the cell |
| 1keV_Other_20%O2 | Number of initial non-DSB clusters per Gbp per Gy, calculated by the MCDS simulation code, for 1 keV energy and 20% oxygen concentration in the cell |
| 10keV_DSBs_1%O2 | Number of DSBs per Gbp per Gy, calculated by the MCDS simulation code, for 10 keV energy and 1% oxygen concentration in the cell |
| 10keV_Other_1%O2 | Number of initial non-DSB clusters per Gbp per Gy, calculated by the MCDS simulation code, for 10 keV energy and 1% oxygen concentration in the cell |
| 10keV_DSBs_20%O2 | Number of initial DSBs per Gbp per Gy, calculated by the MCDS simulation code, for 10 keV energy and 20% oxygen concentration in the cell |
| 10keV_Other_20%O2 | Number of initial non-DSB clusters per Gbp per Gy, calculated by the MCDS simulation code, for 10 keV energy and 20% oxygen concentration in the cell |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanni, V.; Papakonstantinou, D.; Kalospyros, S.A.; Karaoulanis, D.; Biz, G.M.; Manti, L.; Adamopoulos, A.; Pavlopoulou, A.; Georgakilas, A.G. RadPhysBio: A Radiobiological Database for the Prediction of Cell Survival upon Exposure to Ionizing Radiation. Int. J. Mol. Sci. 2024, 25, 4729. https://doi.org/10.3390/ijms25094729
Zanni V, Papakonstantinou D, Kalospyros SA, Karaoulanis D, Biz GM, Manti L, Adamopoulos A, Pavlopoulou A, Georgakilas AG. RadPhysBio: A Radiobiological Database for the Prediction of Cell Survival upon Exposure to Ionizing Radiation. International Journal of Molecular Sciences. 2024; 25(9):4729. https://doi.org/10.3390/ijms25094729
Chicago/Turabian StyleZanni, Vassiliki, Dimitris Papakonstantinou, Spyridon A. Kalospyros, Dimitris Karaoulanis, Gökay Mehmet Biz, Lorenzo Manti, Adam Adamopoulos, Athanasia Pavlopoulou, and Alexandros G. Georgakilas. 2024. "RadPhysBio: A Radiobiological Database for the Prediction of Cell Survival upon Exposure to Ionizing Radiation" International Journal of Molecular Sciences 25, no. 9: 4729. https://doi.org/10.3390/ijms25094729