
NTRK2 Promotes Sheep Granulosa Cells Proliferation and Reproductive Hormone Secretion and Activates the PI3K/AKT Pathway
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Sheep Granulosa Cells In Vitro
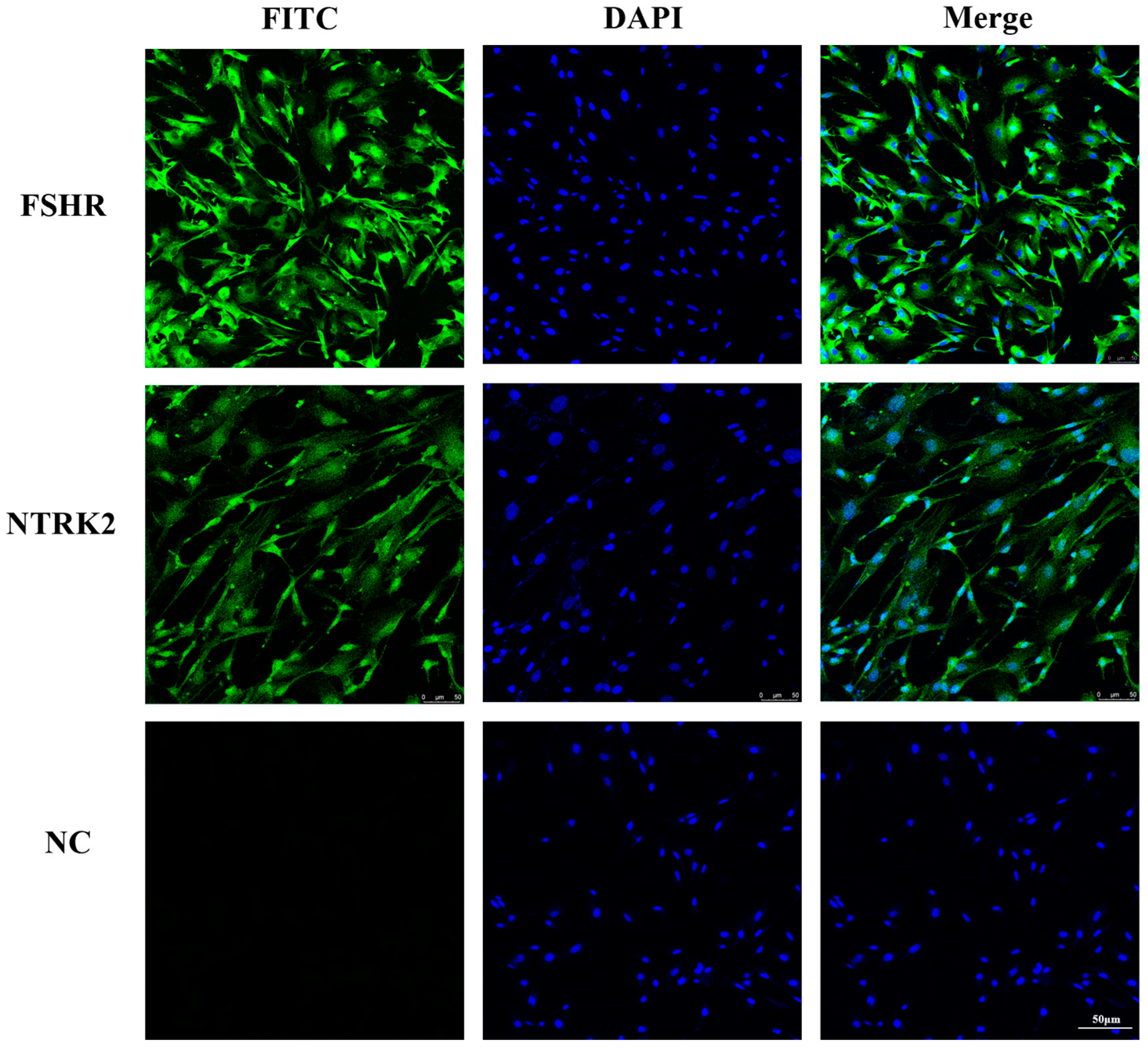
2.2. Granulosa Cell Immunofluorescence Staining
2.3. Plasmid Construction
2.4. Cell Transfection
2.5. RNA Extraction and RT-qPCR
2.6. Protein Extraction and Western Blot Assay
2.7. Cell Counting Experiments
2.8. EdU Assay
2.9. Analysis of Steroid Hormone Secretion
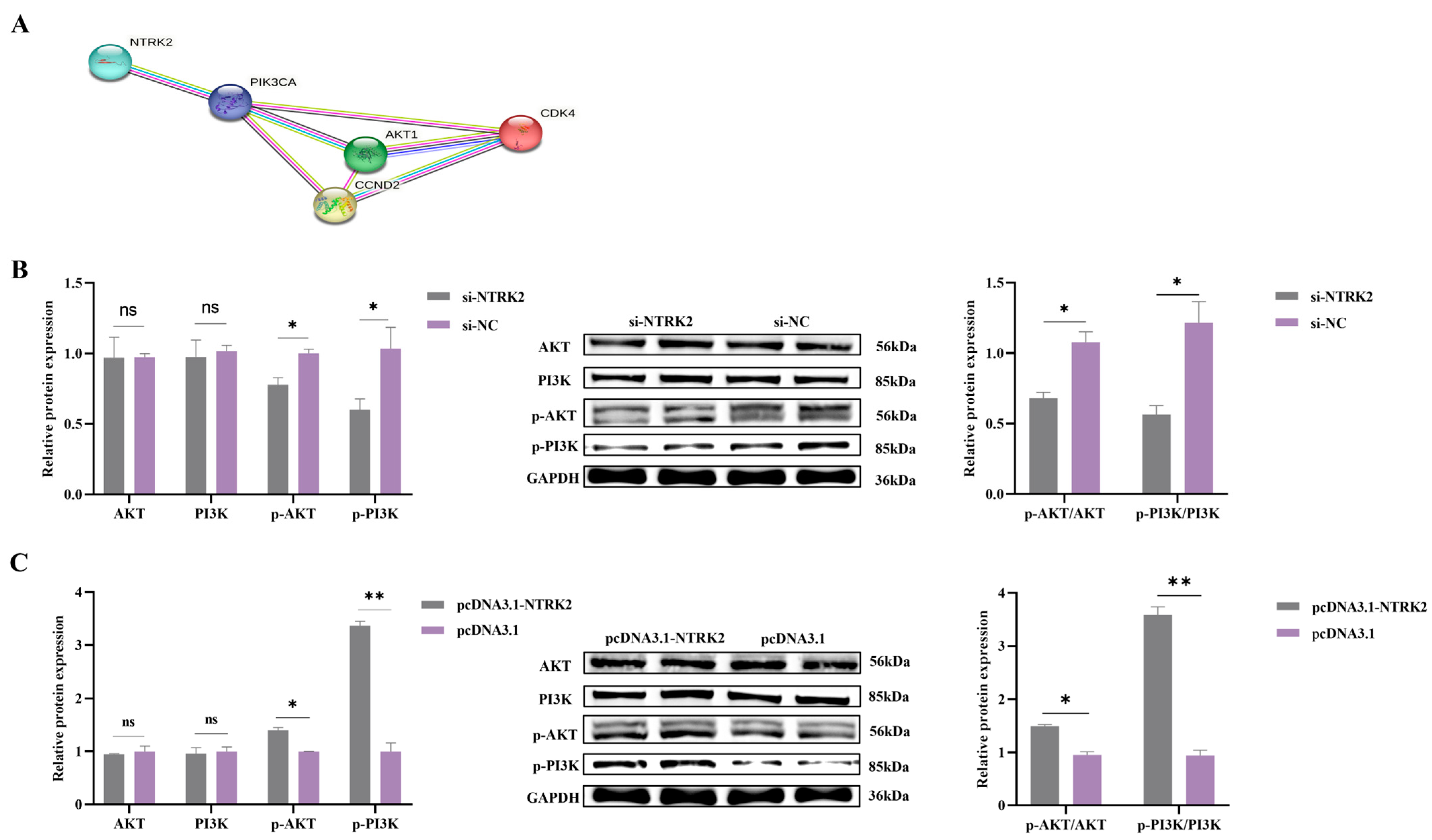
2.10. Construction of Protein–Protein Interaction (PPI) Network
2.11. Statistical Analysis
3. Results
3.1. Identification of Sheep Granulosa Cells and Localization of NTRK2 in Granulosa Cells
3.2. NTRK2 Promotes the Proliferation of Sheep Granulosa Cells
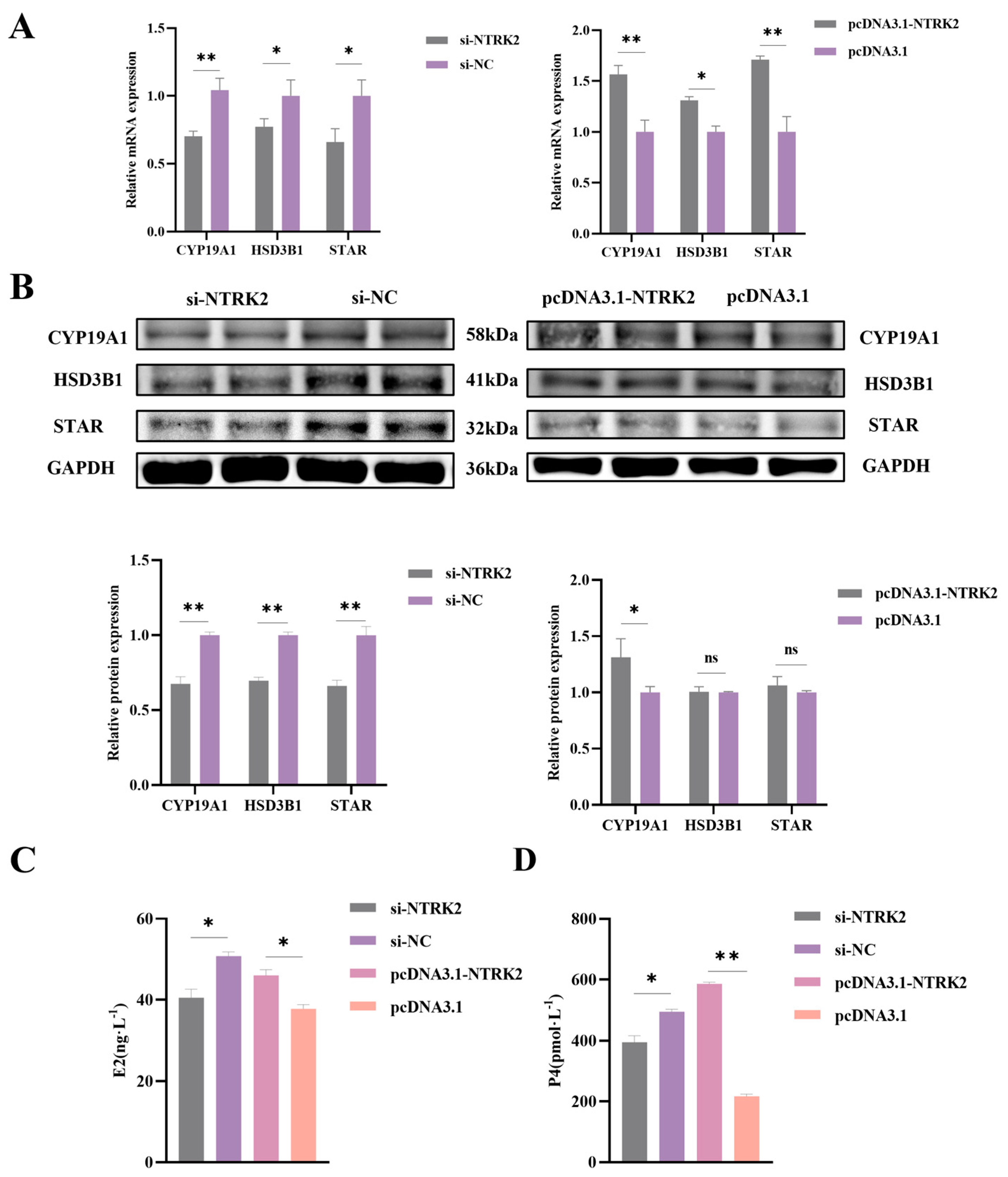
3.3. NTRK2 Promotes Estrogen and Progesterone Secretion in Sheep Granulosa Cells
3.4. NTRK2 Activates the PI3K/AKT Signaling Pathway in Sheep Granulosa Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virant-Klun, I.; Bauer, C.; Ståhlberg, A.; Kubista, M.; Skutella, T. Human oocyte maturation in vitro is improved by co-culture with cumulus cells from mature oocytes. Reprod. Biomed. Online 2018, 36, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Chen, M.J. The Effect of Steroid Hormones on Ovarian Follicle Development. Vitam. Horm. 2018, 107, 155–175. [Google Scholar]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular growth and atresia in mammalian ovaries: Regulation by survival and death of granulosa cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, A. Regulation of ovarian follicular development in primates: Facts and hypotheses. Endocr. Rev. 1996, 17, 121–155. [Google Scholar] [CrossRef] [PubMed]
- Turathum, B.; Gao, E.M.; Chian, R.C. The Function of Cumulus Cells in Oocyte Growth and Maturation and in Subsequent Ovulation and Fertilization. Cells 2021, 10, 2292. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J. Reproduction: Oocytes Call, Granulosa Cells Connect. Curr. Biol. 2018, 28, R354–R356. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Lane, M.; Thompson, J.G. Oocyte-secreted factors: Regulators of cumulus cell function and oocyte quality. Hum. Reprod. Update 2008, 14, 159–177. [Google Scholar] [CrossRef]
- Schulze, J.; Staecker, H.; Wedekind, D.; Lenarz, T.; Warnecke, A. Expression pattern of brain-derived neurotrophic factor and its associated receptors: Implications for exogenous neurotrophin application. Hear. Res. 2022, 413, 108098. [Google Scholar] [CrossRef]
- Hohn, A.; Leibrock, J.; Bailey, K.; Barde, Y.A. Identification and characterization of a novel member of the nerve growth factor/brain-derived neurotrophic factor family. Nature 1990, 344, 339–341. [Google Scholar] [CrossRef]
- Maisonpierre, P.C.; Belluscio, L.; Squinto, S.; Ip, N.Y.; Furth, M.E.; Lindsay, R.M.; Yancopoulos, G.D. Neurotrophin-3: A neurotrophic factor related to NGF and BDNF. Science 1990, 247, 1446–1451. [Google Scholar] [CrossRef]
- Leibrock, J.; Lottspeich, F.; Hohn, A.; Hofer, M.; Hengerer, B.; Masiakowski, P.; Thoenen, H.; Barde, Y.A. Molecular cloning and expression of brain-derived neurotrophic factor. Nature 1989, 341, 149–152. [Google Scholar] [CrossRef]
- Levi-Montalcini, R. The nerve growth factor 35 years later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef]
- Kirstein, M.; Fariñas, I. Sensing life: Regulation of sensory neuron survival by neurotrophins. Cell Mol. Life Sci. 2002, 59, 1787–1802. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Niu, Q.; Xia, T.; Zhou, G.; Li, P.; Zhao, Q.; Xu, C.; Dong, L.; Zhang, S.; Wang, A. ERK1/2-mediated disruption of BDNF-TrkB signaling causes synaptic impairment contributing to fluoride-induced developmental neurotoxicity. Toxicology 2018, 410, 222–230. [Google Scholar] [CrossRef]
- Ka, M.; Kim, W.Y. ANKRD11 associated with intellectual disability and autism regulates dendrite differentiation via the BDNF/TrkB signaling pathway. Neurobiol. Dis. 2018, 111, 138–152. [Google Scholar] [CrossRef]
- Martins da Silva, S.J.; Gardner, J.O.; Taylor, J.E.; Springbett, A.; De Sousa, P.A.; Anderson, R.A. Brain-derived neurotrophic factor promotes bovine oocyte cytoplasmic competence for embryo development. Reproduction 2005, 129, 423–434. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Y.; Li, S.; Chen, P.; Zhang, J.; Feng, L. BDNF promotes mouse follicular development and reverses ovarian aging by promoting cell proliferation. J. Ovarian Res. 2023, 16, 83. [Google Scholar] [CrossRef]
- Esmaeili-Fard, S.M.; Gholizadeh, M.; Hafezian, S.H.; Abdollahi-Arpanahi, R. Genome-wide association study and pathway analysis identify NTRK2 as a novel candidate gene for litter size in sheep. PLoS ONE 2021, 16, e0244408. [Google Scholar] [CrossRef] [PubMed]
- Seifer, D.B.; Feng, B.; Shelden, R.M. Immunocytochemical evidence for the presence and location of the neurotrophin-Trk receptor family in adult human preovulatory ovarian follicles. Am. J. Obstet. Gynecol. 2006, 194, 1129–1134; discussion 1134–1136. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.; Johnson, A.L. Expression and function of brain-derived neurotrophin factor and its receptor, TrkB, in ovarian follicles from the domestic hen (Gallus gallus domesticus). J. Exp. Biol. 2001, 204, 2087–2095. [Google Scholar] [CrossRef]
- Sahu, M.P.; Pazos-Boubeta, Y.; Pajanoja, C.; Rozov, S.; Panula, P.; Castrén, E. Neurotrophin receptor Ntrk2b function in the maintenance of dopamine and serotonin neurons in zebrafish. Sci. Rep. 2019, 9, 2036. [Google Scholar] [CrossRef]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Cometa, M.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; et al. The BDNF/TrkB Neurotrophin System in the Sensory Organs of Zebrafish. Int. J. Mol. Sci. 2022, 23, 2621. [Google Scholar] [CrossRef] [PubMed]
- Dissen, G.A.; Garcia-Rudaz, C.; Ojeda, S.R. Role of neurotrophic factors in early ovarian development. Semin. Reprod. Med. 2009, 27, 24–31. [Google Scholar] [CrossRef]
- Paredes, A.; Romero, C.; Dissen, G.A.; DeChiara, T.M.; Reichardt, L.; Cornea, A.; Ojeda, S.R.; Xu, B. TrkB receptors are required for follicular growth and oocyte survival in the mammalian ovary. Dev. Biol. 2004, 267, 430–449. [Google Scholar] [CrossRef]
- Chen, S.; Wang, F.; Liu, Z.; Zhao, Y.; Jiang, Y.; Chen, L.; Li, C.; Zhou, X. Brain-derived neurotrophic factor promotes proliferation and progesterone synthesis in bovine granulosa cells. J. Cell Physiol. 2019, 234, 8776–8787. [Google Scholar] [CrossRef]
- Mischel, P.S.; Smith, S.G.; Vining, E.R.; Valletta, J.S.; Mobley, W.C.; Reichardt, L.F. The extracellular domain of p75NTR is necessary to inhibit neurotrophin-3 signaling through TrkA. J. Biol. Chem. 2001, 276, 11294–11301. [Google Scholar] [CrossRef]
- Numakawa, T.; Suzuki, S.; Kumamaru, E.; Adachi, N.; Richards, M.; Kunugi, H. BDNF function and intracellular signaling in neurons. Histol. Histopathol. 2010, 25, 237–258. [Google Scholar] [PubMed]
- Qin, X.; Zhao, Y.; Zhang, T.; Yin, C.; Qiao, J.; Guo, W.; Lu, B. TrkB agonist antibody ameliorates fertility deficits in aged and cyclophosphamide-induced premature ovarian failure model mice. Nat. Commun. 2022, 13, 914. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, D.; Wang, D.; Wang, L.; Weng, Y.; Wang, H.; Wu, X.; Wang, Y. Moderate Aerobic Exercise Regulates Follicular Dysfunction by Initiating Brain-Derived Neurotrophic Factor (BDNF)-Mediated Anti-Apoptotic Signaling Pathways in Polycystic Ovary Syndrome. J. Clin. Med. 2022, 11, 5584. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.; Wessels, J.M.; Foster, W.G. Brain-derived neurotrophic factor (BDNF) expression and function in the mammalian reproductive Tract. Hum. Reprod. Update 2020, 26, 545–564. [Google Scholar] [CrossRef]
- Dorfman, M.D.; Kerr, B.; Garcia-Rudaz, C.; Paredes, A.H.; Dissen, G.A.; Ojeda, S.R. Neurotrophins acting via TRKB receptors activate the JAGGED1-NOTCH2 cell-cell communication pathway to facilitate early ovarian development. Endocrinology 2011, 152, 5005–5016. [Google Scholar] [CrossRef] [PubMed]
- Anger, D.L.; Zhang, B.; Boutross-Tadross, O.; Foster, W.G. Tyrosine receptor kinase B (TrkB) protein expression in the human endometrium. Endocrine 2007, 31, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.; Parada, L.F.; Coulier, F.; Barbacid, M. Trkb, a novel tyrosine protein kinase receptor expressed during mouse neural development. EMBO J. 1989, 8, 3701–3709. [Google Scholar] [CrossRef] [PubMed]
- Bilbrough, T.; Piemontese, E.; Seitz, O. Dissecting the role of protein phosphorylation: A chemical biology toolbox. Chem. Soc. Rev. 2022, 51, 5691–5730. [Google Scholar] [CrossRef] [PubMed]
- Hensen, K.; Pook, M.; Sikut, A.; Org, T.; Maimets, T.; Salumets, A.; Kurg, A. Utilising FGF2, IGF2 and FSH in serum-free protocol for long-term in vitro cultivation of primary human granulosa cells. Mol. Cell Endocrinol. 2020, 510, 110816. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Datta, S.R.; Greenberg, M.E. Transcription-dependent and -independent control of neuronal survival by the PI3K-Akt signaling pathway. Curr. Opin. Neurobiol. 2001, 11, 297–305. [Google Scholar] [CrossRef]
- Fiorentino, G.; Cimadomo, D.; Innocenti, F.; Soscia, D.; Vaiarelli, A.; Ubaldi, F.M.; Gennarelli, G.; Garagna, S.; Rienzi, L.; Zuccotti, M. Biomechanical forces and signals operating in the ovary during folliculogenesis and their dysregulation: Implications for fertility. Hum. Reprod. Update 2023, 29, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.Y.; An, X.P.; Fang, F.; Gao, K.X.; Xin, H.Y.; Han, P.; Bao, L.J.; Ma, H.D.; Cao, B.Y. MicroRNA-10b suppresses goat granulosa cell proliferation by targeting brain-derived neurotropic factor. Domest. Anim. Endocrinol. 2016, 54, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Rajkovic, A. Genetics of early mammalian folliculogenesis. Cell Mol. Life Sci. 2006, 63, 579–590. [Google Scholar] [CrossRef]
- McGee, E.A.; Hsueh, A.J. Initial and cyclic recruitment of ovarian follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar] [PubMed]
- Son, W.Y.; Das, M.; Shalom-Paz, E.; Holzer, H. Mechanisms of follicle selection and development. Minerva Ginecol. 2011, 63, 89–102. [Google Scholar] [PubMed]
- Chang, H.M.; Wu, H.C.; Sun, Z.G.; Lian, F.; Leung, P.C.K. Neurotrophins and glial cell line-derived neurotrophic factor in the ovary: Physiological and pathophysiological implications. Hum. Reprod. Update 2019, 25, 224–242. [Google Scholar] [CrossRef]
- Moley, K.H.; Schreiber, J.R. Ovarian follicular growth, ovulation and atresia. Endocrine, paracrine and autocrine regulation. Adv. Exp. Med. Biol. 1995, 377, 103–119. [Google Scholar]
- Streiter, S.; Fisch, B.; Sabbah, B.; Ao, A.; Abir, R. The importance of neuronal growth factors in the ovary. Mol. Hum. Reprod. 2016, 22, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Du, F.; Liu, X.; Ruan, Q.; Wu, Z.; Lei, C.; Deng, Y.; Luo, C.; Jiang, J.; Shi, D.; et al. Brain-derived neurotrophic factor (BDNF) is expressed in buffalo (Bubalus bubalis) ovarian follicles and promotes oocyte maturation and early embryonic development. Theriogenology 2019, 130, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Mirshokraei, P.; Hassanpour, H.; Rahnama, A.; Foster, W.G. Gene expression of BDNF and its receptors, TrkB and p75 in the uterus and oviduct of pregnant and non-pregnant ewes. Res. Vet. Sci. 2013, 95, 164–168. [Google Scholar] [CrossRef]
- Kim, M.; Hyun, S.H. Neurotrophic factors in the porcine ovary: Their effects on follicular growth, oocyte maturation, and developmental competence. Front. Vet. Sci. 2022, 9, 931402. [Google Scholar] [CrossRef]
- Qiu, L.; Zhou, C.; Sun, Y.; Di, W.; Scheffler, E.; Healey, S.; Kouttab, N.; Chu, W.; Wan, Y. Crosstalk between EGFR and TrkB enhances ovarian cancer cell migration and proliferation. Int. J. Oncol. 2006, 29, 1003–1011. [Google Scholar] [CrossRef]
- Cao, M.; Niu, Q.; Xiang, X.; Yuan, C.; Iqbal, T.; Huang, Y.; Tian, M.; Zhao, Z.; Li, C.; Zhou, X. Brain-Derived Neurotrophic Factor Regulates Ishikawa Cell Proliferation through the TrkB-ERK1/2 Signaling Pathway. Biomolecules 2020, 10, 1645. [Google Scholar] [CrossRef]
- Kwapiszewska, G.; Chwalek, K.; Marsh, L.M.; Wygrecka, M.; Wilhelm, J.; Best, J.; Egemnazarov, B.; Weisel, F.C.; Osswald, S.L.; Schermuly, R.T.; et al. BDNF/TrkB signaling augments smooth muscle cell proliferation in pulmonary hypertension. Am. J. Pathol. 2012, 181, 2018–2029. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xin, Q.; Wang, X.; Wang, S.; Wang, H.; Zhang, W.; Yang, Y.; Zhang, Y.; Zhang, Z.; Wang, C.; et al. Estrogen receptors in granulosa cells govern meiotic resumption of pre-ovulatory oocytes in mammals. Cell Death Dis. 2017, 8, e2662. [Google Scholar] [CrossRef]
- Dong, F.; Meng, T.G.; Li, J.; Wang, F.; Li, Y.Y.; Ouyang, Y.C.; Hou, Y.; Wang, Z.B.; Schatten, H.; Sun, Q.Y. Inhibition of CDK4/6 kinases causes production of aneuploid oocytes by inactivating the spindle assembly checkpoint and accelerating first meiotic progression. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119044. [Google Scholar] [CrossRef] [PubMed]
- Moons, D.S.; Jirawatnotai, S.; Tsutsui, T.; Franks, R.; Parlow, A.F.; Hales, D.B.; Gibori, G.; Fazleabas, A.T.; Kiyokawa, H. Intact follicular maturation and defective luteal function in mice deficient for cyclin- dependent kinase-4. Endocrinology 2002, 143, 647–654. [Google Scholar] [CrossRef]
- Richards, J.S.; Hedin, L. Molecular aspects of hormone action in ovarian follicular development, ovulation, and luteinization. Annu. Rev. Physiol. 1988, 50, 441–463. [Google Scholar] [CrossRef] [PubMed]
- Machelon, V.; Emilie, D. Production of ovarian cytokines and their role in ovulation in the mammalian ovary. Eur. Cytokine Netw. 1997, 8, 137–143. [Google Scholar]
- Li, T.; Mo, H.; Chen, W.; Li, L.; Xiao, Y.; Zhang, J.; Li, X.; Lu, Y. Role of the PI3K-Akt Signaling Pathway in the Pathogenesis of Polycystic Ovary Syndrome. Reprod. Sci. 2017, 24, 646–655. [Google Scholar] [CrossRef]
- Li, L.; She, X.; Wang, Z. The Signaling Pathways involved in Ovarian Follicle Development. Front. Physiol. 2021, 12, 730196. [Google Scholar] [CrossRef]
- Monte, A.P.O.; Bezerra, M.É.S.; Menezes, V.G.; Gouveia, B.B.; Barberino, R.S.; Lins, T.L.B.G.; Barros, V.R.P.; Santos, J.M.S.; Donfack, N.J.; Matos, M.H.T. Involvement of Phosphorylated Akt and FOXO3a in the Effects of Growth and Differentiation Factor-9 (GDF-9) on Inhibition of Follicular Apoptosis and Induction of Granulosa Cell Proliferation After In Vitro Culture of Sheep Ovarian Tissue. Reprod Sci. 2021, 28, 2174–2185. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| siRNA | Sequences (5′-3′) |
|---|---|
| siRNA-1 | F: GCUGGCUGAUUGUGGGCUUTT R: AAGCCCACAAUCAGCCAGCTT |
| siRNA-2 | F: GGAGUCUACCAGUGCUGAUTT R: AUCAGCACUGGUAGACUCCTT |
| siRNA-3 | F: GGAUUUGUACUGUCUGGAUTT R: AUCCAGACAGUACAAAUCCTT |
| Negative control (NC) | F: UUCUCCGAACGUGUCACGUTT R: ACGUGACACGUUCGGAGAATT |
| Genes | Accession | Sequences (5′–3′) | Product Length (bp) |
|---|---|---|---|
| NTRK2 | XM_042243072.1 | F: GTTTGGCATGAAAGGCCCAG R: GAGATGTGATGCAGTGGGCT | 72 |
| PI3K | XM_042234194.1 | F: GTGTGGGACTTATTGAGGTGGTGAG R: TGATGGAGTGTGTGGCTGTTGAAC | 106 |
| AKT | NM_001161857.1 | F: GCAGCATCGTGTGGCAGGAC R: GTCTTGGTCAGGTGGCGTAATGG | 140 |
| CCND2 | NM_001127290.1 | F: ATCTCCTGGCAAAGATCACCAACAC R: TGTTCAGCAGCACCACCTCAATC | 81 |
| CDK4 | XM_012158548.4 | F: GCTGCTGCTGGAGATGCTGAC R: CTCTGCGTCACCTTCTGCCTTG | 100 |
| CYP19A1 | XM_042251921.1 | F: AGTCTGGGTATGTGGAGAGGAAAC R: CTGCCAAATCGGGATATGTAGTGAC | 96 |
| HSD3B1 | XM_042255821.1 | F: TGGGTGGGCTCTGAAGAATGG R: TAATTGGTCAGGATGCCGTTGTTC | 135 |
| STAR | NM_001009243.1 | F: AGTCTGGGTATGTGGAGAGGAAAC R: CTGCCAAATCGGGATATGTAGTGAC | 70 |
| RPL19 | XM_027974613.2 | F: ATCGCCAATGCCAACTC R: CCTTTCGCTTACCTATACC | 154 |
| Article Number | Antibodies | Dilution Ratio | Place of Origin |
|---|---|---|---|
| 29961-1-AP | anti-NTRK2 | 1:1000 | Proteintech, Wuhan, China |
| 60225-1-Ig | anti-PI3K | 1:10,000 | Proteintech, Wuhan, China |
| 60004-1-Ig | anti-GAPDH | 1:100,000 | Proteintech, Wuhan, China |
| 10934-1-AP | anti-cyclin-D2 | 1:1000 | Proteintech, Wuhan, China |
| 11026-1-AP | anti-CDK4 | 1:1000 | Proteintech, Wuhan, China |
| 10176-2-AP | anti-AKT | 1:2000 | Proteintech, Wuhan, China |
| 22665-1-AP | anti-FSHR | 1:1000 | Proteintech, Wuhan, China |
| ab182651 | anti-P-PI3K | 1:1000 | Abcam, Shanghai, China |
| ab81283 | anti-P-AKT | 1:1000 | Abcam, Shanghai, China |
| bs-1292R | anti-Aromatase | 1:1000 | Bioss, Beijing, China |
| bs-3906R | anti-HSD3B1 | 1:1000 | Bioss, Beijing, China |
| bs-20387R | anti-STAR | 1:1000 | Bioss, Beijing, China |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Liu, Y.; Wang, P.; Liu, Z.; Zhang, R.; Chu, M.; Zhao, A. NTRK2 Promotes Sheep Granulosa Cells Proliferation and Reproductive Hormone Secretion and Activates the PI3K/AKT Pathway. Animals 2024, 14, 1465. https://doi.org/10.3390/ani14101465
Jia Y, Liu Y, Wang P, Liu Z, Zhang R, Chu M, Zhao A. NTRK2 Promotes Sheep Granulosa Cells Proliferation and Reproductive Hormone Secretion and Activates the PI3K/AKT Pathway. Animals. 2024; 14(10):1465. https://doi.org/10.3390/ani14101465
Chicago/Turabian StyleJia, Yuhang, Yufang Liu, Peng Wang, Ziyi Liu, Runan Zhang, Mingxing Chu, and Ayong Zhao. 2024. "NTRK2 Promotes Sheep Granulosa Cells Proliferation and Reproductive Hormone Secretion and Activates the PI3K/AKT Pathway" Animals 14, no. 10: 1465. https://doi.org/10.3390/ani14101465