
Fecal Microbial Communities of Nellore and Crossbred Beef Calves Raised at Pasture
 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diet
2.3. Sampling
2.4. DNA Extraction and Sequencing
2.5. Bioinformatics
2.6. Statistics
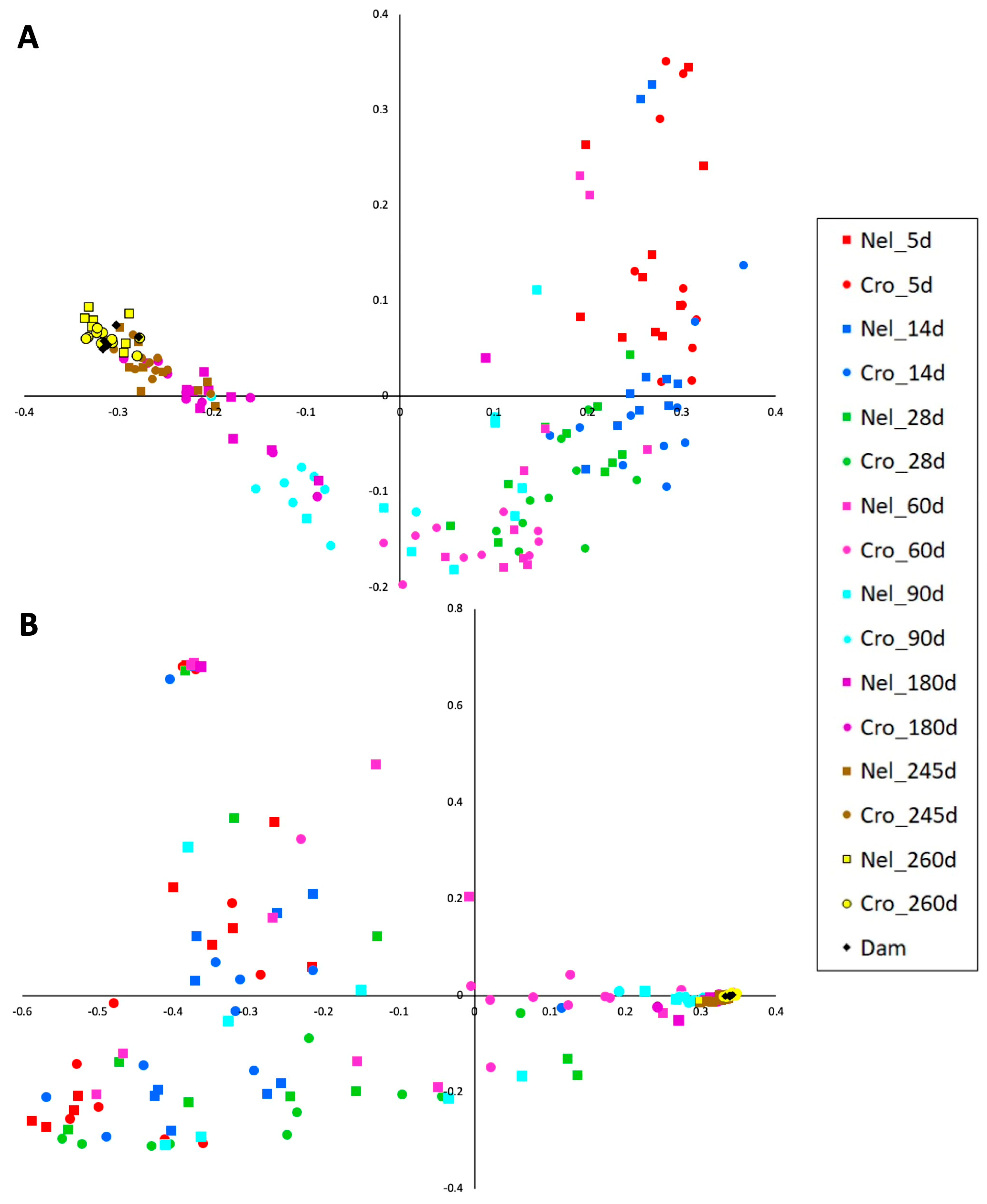
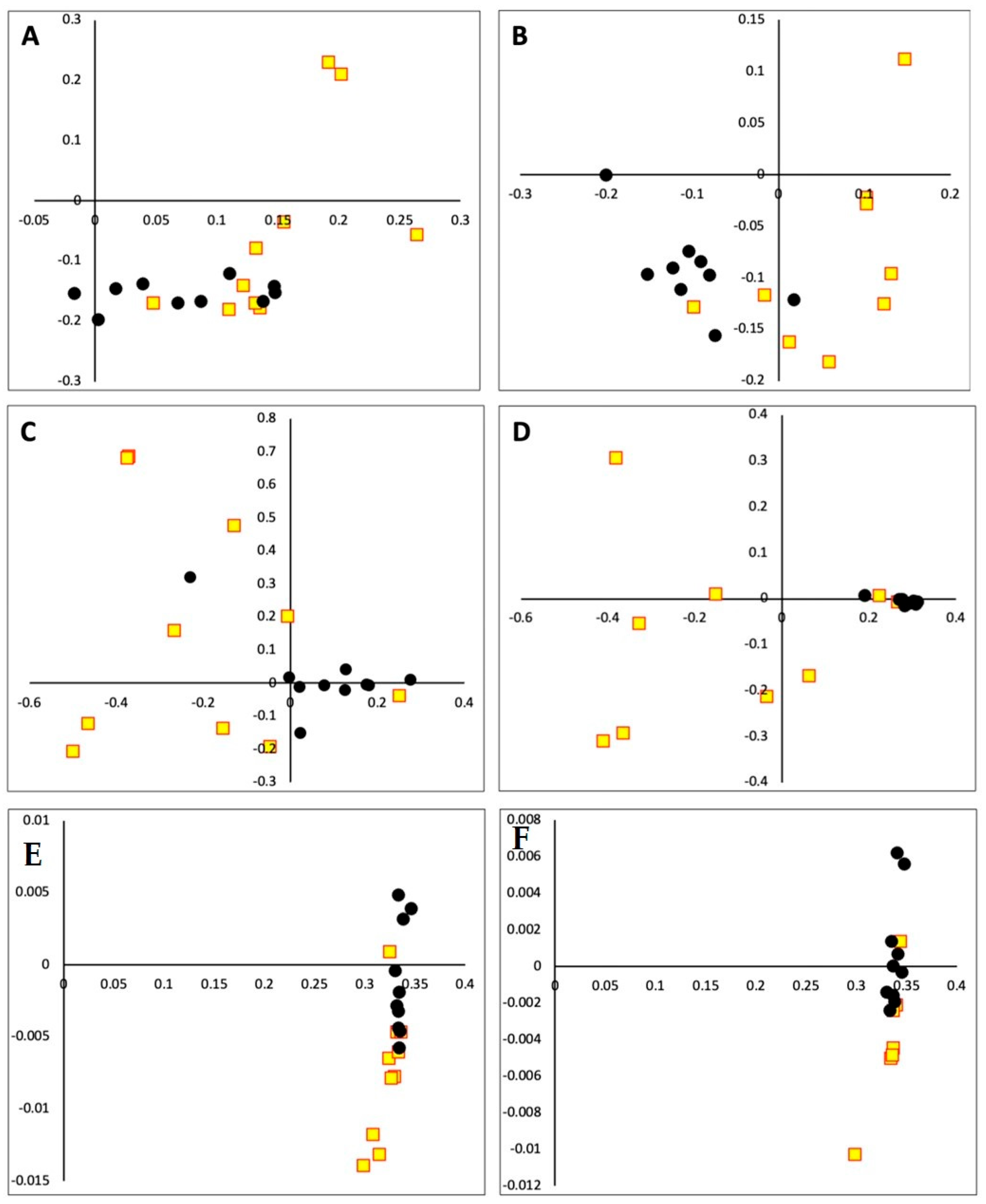
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malmuthuge, N.; Li, M.; Goonewardene, L.A.; Oba, M.; Guan, L.L. Effect of calf starter feeding on gut microbial diversity and expression of genes involved in host immune responses and tight junctions in dairy calves during weaning transition. J. Dairy Sci. 2013, 96, 3189–3200. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Wang, S.; Mora, J.R.; et al. Gut immune maturation depends on colonization with a host-specific microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef] [PubMed]
- Klein-Jobstl, D.; Quijada, N.M.; Dzieciol, M.; Feldbacher, B.; Wagner, M.; Drillich, M.; Schmitz-Esser, S.; Mann, E. Microbiota of newborn calves and their mothers reveals possible transfer routes for newborn calves’ gastrointestinal microbiota. PLoS ONE 2019, 14, e0220554. [Google Scholar] [CrossRef] [PubMed]
- Yeoman, C.J.; Ishaq, S.L.; Bichi, E.; Olivo, S.K.; Lowe, J.; Aldridge, B.M. Biogeographical Differences in the Influence of Maternal Microbial Sources on the Early Successional Development of the Bovine Neonatal Gastrointestinal tract. Sci. Rep. 2018, 8, 3197. [Google Scholar] [CrossRef] [PubMed]
- Klein-Jobstl, D.; Schornsteiner, E.; Mann, E.; Wagner, M.; Drillich, M.; Schmitz-Esser, S. Pyrosequencing reveals diverse fecal microbiota in Simmental calves during early development. Front. Microbiol. 2014, 5, 622. [Google Scholar] [CrossRef] [PubMed]
- Alipour, M.J.; Jalanka, J.; Pessa-Morikawa, T.; Kokkonen, T.; Satokari, R.; Hynonen, U.; Iivanainen, A.; Niku, M. The composition of the perinatal intestinal microbiota in cattle. Sci. Rep. 2018, 8, 10437. [Google Scholar] [CrossRef] [PubMed]
- Oultram, J.; Phipps, E.; Teixeira, A.G.; Foditsch, C.; Bicalho, M.L.; Machado, V.S.; Bicalho, R.C.; Oikonomou, G. Effects of antibiotics (oxytetracycline, florfenicol or tulathromycin) on neonatal calves’ faecal microbial diversity. Vet. Rec. 2015, 177, 598. [Google Scholar] [CrossRef]
- Gomez, D.E.; Galvao, K.N.; Rodriguez-Lecompte, J.C.; Costa, M.C. The Cattle Microbiota and the Immune System: An Evolving Field. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 485–505. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.E.; Arroyo, L.G.; Costa, M.C.; Viel, L.; Weese, J.S. Characterization of the Fecal Bacterial Microbiota of Healthy and Diarrheic Dairy Calves. J. Vet. Intern. Med. 2017, 31, 928–939. [Google Scholar] [CrossRef]
- Xie, G.; Duff, G.C.; Hall, L.W.; Allen, J.D.; Burrows, C.D.; Bernal-Rigoli, J.C.; Dowd, S.E.; Guerriero, V.; Yeoman, C.J. Alteration of digestive tract microbiome in neonatal Holstein bull calves by bacitracin methylene disalicylate treatment and scours. J. Anim. Sci. 2013, 91, 4984–4990. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Dill-McFarland, K.A.; Breaker, J.D.; Suen, G. Microbial succession in the gastrointestinal tract of dairy cows from 2 weeks to first lactation. Sci. Rep. 2017, 7, 40864. [Google Scholar] [CrossRef] [PubMed]
- Vigors, S.; Flores-Villalva, S.; Meade, K.G. The impact of vitamin D3 supplementation on the faecal and oral microbiome of dairy calves indoors or at pasture. Sci. Rep. 2023, 13, 9111. [Google Scholar] [CrossRef] [PubMed]
- Cristobal-Carballo, O.; McCoard, S.A.; Cookson, A.L.; Laven, R.A.; Ganesh, S.; Lewis, S.J.; Muetzel, S. Effect of Divergent Feeding Regimes during Early Life on the Rumen Microbiota in Calves. Front. Microbiol. 2021, 12, 711040. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Chen, X.; Liu, Y.; Ali, F.; Idrees, A.; Ataya, F.S.; Shang, Z.; Li, K. Sodium acetate and sodium butyrate attenuate diarrhea in yak calves by regulating gut microbiota and metabolites. Heliyon 2024, 10, e26564. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Ma, Y.; Chen, X.; Yang, S.; Zhang, S.; Liu, S.; Xiao, J.; Wang, Y.; Wang, W.; Yang, H.; et al. Temporal Changes in Fecal Unabsorbed Carbohydrates Relative to Perturbations in Gut Microbiome of Neonatal Calves: Emerging of Diarrhea Induced by Extended-Spectrum beta-lactamase-Producing Enteroaggregative Escherichia coli. Front. Microbiol. 2022, 13, 883090. [Google Scholar] [CrossRef] [PubMed]
- Cristobal-Carballo, O.; McCoard, S.A.; Cookson, A.L.; Ganesh, S.; Lowe, K.; Laven, R.A.; Muetzel, S. Effect of Methane Inhibitors on Ruminal Microbiota during Early Life and Its Relationship with Ruminal Metabolism and Growth in Calves. Front. Microbiol. 2021, 12, 710914. [Google Scholar] [CrossRef] [PubMed]
- Kido, K.; Tejima, S.; Haramiishi, M.; Uyeno, Y.; Ide, Y.; Kurosu, K.; Kushibiki, S. Provision of beta-glucan prebiotics (cellooligosaccharides and kraft pulp) to calves from pre- to post-weaning period on pasture. Anim. Sci. J. 2019, 90, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Meale, S.J.; Li, S.; Azevedo, P.; Derakhshani, H.; Plaizier, J.C.; Khafipour, E.; Steele, M.A. Development of Ruminal and Fecal Microbiomes Are Affected by Weaning but Not Weaning Strategy in Dairy Calves. Front. Microbiol. 2016, 7, 582. [Google Scholar] [CrossRef]
- Meale, S.J.; Li, S.C.; Azevedo, P.; Derakhshani, H.; DeVries, T.J.; Plaizier, J.C.; Steele, M.A.; Khafipour, E. Weaning age influences the severity of gastrointestinal microbiome shifts in dairy calves. Sci. Rep. 2017, 7, 198. [Google Scholar] [CrossRef]
- Weese, J.S.; Jelinski, M. Assessment of the Fecal Microbiota in Beef Calves. J. Vet. Intern. Med. 2017, 31, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Hitch, T.C.A.; Chen, Y.; Creevey, C.J.; Guan, L.L. Comparative metagenomic and metatranscriptomic analyses reveal the breed effect on the rumen microbiome and its associations with feed efficiency in beef cattle. Microbiome 2019, 7, 6. [Google Scholar] [CrossRef]
- Paz, H.A.; Anderson, C.L.; Muller, M.J.; Kononoff, P.J.; Fernando, S.C. Rumen Bacterial Community Composition in Holstein and Jersey Cows Is Different under Same Dietary Condition and Is Not Affected by Sampling Method. Front. Microbiol. 2016, 7, 1206. [Google Scholar] [CrossRef] [PubMed]
- Roehe, R.; Dewhurst, R.J.; Duthie, C.A.; Rooke, J.A.; McKain, N.; Ross, D.W.; Hyslop, J.J.; Waterhouse, A.; Freeman, T.C.; Watson, M.; et al. Bovine Host Genetic Variation Influences Rumen Microbial Methane Production with Best Selection Criterion for Low Methane Emitting and Efficiently Feed Converting Hosts Based on Metagenomic Gene Abundance. PLoS Genet. 2016, 12, e1005846. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Bian, B.; Teng, L.; Nelson, C.D.; Driver, J.; Elzo, M.A.; Jeong, K.C. Host genetic effects upon the early gut microbiota in a bovine model with graduated spectrum of genetic variation. ISME J. 2020, 14, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S.; MacNeil, M.D.; Carvalho, H.G.; Vaz, R.Z.; Cardoso, F.F. Direct and maternal breed additive and heterosis effects on growth traits of beef cattle raised in southern Brazil. J. Anim. Sci. 2018, 96, 2536–2544. [Google Scholar] [CrossRef]
- Mendonca, F.S.; MacNeil, M.D.; Leal, W.S.; Azambuja, R.C.C.; Rodrigues, P.F.; Cardoso, F.F. Crossbreeding effects on growth and efficiency in beef cow-calf systems: Evaluation of Angus, Caracu, Hereford and Nelore breed direct, maternal and heterosis effects. Transl. Anim. Sci. 2019, 3, 1286–1295. [Google Scholar] [CrossRef]
- Nogueira, G.P. Puberty in South American Bos indicus (Zebu) cattle. Anim. Reprod. Sci. 2004, 82–83, 361–372. [Google Scholar] [CrossRef]
- Cho, I.; Yamanishi, S.; Cox, L.; Methe, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef]
- Oikonomou, G.; Teixeira, A.G.; Foditsch, C.; Bicalho, M.L.; Machado, V.S.; Bicalho, R.C. Fecal microbial diversity in pre-weaned dairy calves as described by pyrosequencing of metagenomic 16S rDNA. Associations of Faecalibacterium species with health and growth. PLoS ONE 2013, 8, e63157. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.A.; Hassan, F.U.; Rehman, M.S.; Huws, S.A.; Cheng, Y.; Din, A.U. Gut microbiome colonization and development in neonatal ruminants: Strategies, prospects, and opportunities. Anim. Nutr. 2021, 7, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Difford, G.F.; Plichta, D.R.; Lovendahl, P.; Lassen, J.; Noel, S.J.; Hojberg, O.; Wright, A.G.; Zhu, Z.; Kristensen, L.; Nielsen, H.B.; et al. Host genetics and the rumen microbiome jointly associate with methane emissions in dairy cows. PLoS Genet. 2018, 14, e1007580. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Recio, O.; Zubiria, I.; Garcia-Rodriguez, A.; Hurtado, A.; Atxaerandio, R. Short communication: Signs of host genetic regulation in the microbiome composition in 2 dairy breeds: Holstein and Brown Swiss. J. Dairy Sci. 2018, 101, 2285–2292. [Google Scholar] [CrossRef] [PubMed]
- Abecia, L.; Jimenez, E.; Martinez-Fernandez, G.; Martin-Garcia, A.I.; Ramos-Morales, E.; Pinloche, E.; Denman, S.E.; Newbold, C.J.; Yanez-Ruiz, D.R. Natural and artificial feeding management before weaning promote different rumen microbial colonization but not differences in gene expression levels at the rumen epithelium of newborn goats. PLoS ONE 2017, 12, e0182235. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Ma, S.; Zhu, Z.; Su, Y.; Zoetendal, E.G.; Mackie, R.; Liu, J.; Mu, C.; Huang, R.; Smidt, H.; et al. Age, introduction of solid feed and weaning are more important determinants of gut bacterial succession in piglets than breed and nursing mother as revealed by a reciprocal cross-fostering model. Environ. Microbiol. 2016, 18, 1566–1577. [Google Scholar] [CrossRef]
- Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Pokorna, A.; Cizek, A.; et al. Contact with adult hen affects development of caecal microbiota in newly hatched chicks. PLoS ONE 2019, 14, e0212446. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Maldonado-Barragan, A.; Moles, L.; Rodriguez-Banos, M.; Campo, R.D.; Fernandez, L.; Rodriguez, J.M.; Jimenez, E. Sharing of bacterial strains between breast milk and infant feces. J. Hum. Lact. 2012, 28, 36–44. [Google Scholar] [CrossRef]
- Li, M.; Wang, Z.; Wang, L.; Xue, B.; Hu, R.; Zou, H.; Liu, S.; Shah, A.M.; Peng, Q. Comparison of changes in fecal microbiota of calves with and without dam. PeerJ 2022, 10, e12826. [Google Scholar] [CrossRef]
- Conneely, M.; Berry, D.P.; Murphy, J.P.; Lorenz, I.; Doherty, M.L.; Kennedy, E. Effect of feeding colostrum at different volumes and subsequent number of transition milk feeds on the serum immunoglobulin G concentration and health status of dairy calves. J. Dairy Sci. 2014, 97, 6991–7000. [Google Scholar] [CrossRef] [PubMed]
- Novo, S.M.F.; Costa, J.; Baccili, C.C.; Sobreira, N.M.; Silva, B.T.; de Oliveira, P.L.; Hurley, D.J.; Gomes, V. Effect of maternal cells transferred with colostrum on the health of neonate calves. Res. Vet. Sci. 2017, 112, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Barden, M.; Richards-Rios, P.; Ganda, E.; Lenzi, L.; Eccles, R.; Neary, J.; Oultram, J.; Oikonomou, G. Maternal influences on oral and faecal microbiota maturation in neonatal calves in beef and dairy production systems. Anim. Microbiome 2020, 2, 31. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.C.; Stampfli, H.R.; Allen-Vercoe, E.; Weese, J.S. Development of the faecal microbiota in foals. Equine Vet. J. 2016, 48, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, G.; Siegel, P.; He, C.; Wang, H.; Zhao, W.; Zhai, Z.; Tian, F.; Zhao, J.; Zhang, H.; et al. Quantitative genetic background of the host influences gut microbiomes in chickens. Sci. Rep. 2013, 3, 1163. [Google Scholar] [CrossRef] [PubMed]
- Melis, C.; Billing, A.M.; Wold, P.A.; Ludington, W.B. Gut microbiome dysbiosis is associated with host genetics in the Norwegian Lundehund. Front. Microbiol. 2023, 14, 1209158. [Google Scholar] [CrossRef] [PubMed]
- Cahana, I.; Iraqi, F.A. Impact of host genetics on gut microbiome: Take-home lessons from human and mouse studies. Animal Model. Exp. Med. 2020, 3, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Org, E.; Parks, B.W.; Joo, J.W.; Emert, B.; Schwartzman, W.; Kang, E.Y.; Mehrabian, M.; Pan, C.; Knight, R.; Gunsalus, R.; et al. Genetic and environmental control of host-gut microbiota interactions. Genome Res. 2015, 25, 1558–1569. [Google Scholar] [CrossRef]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef]
- Hernandez-Sanabria, E.; Goonewardene, L.A.; Wang, Z.; Zhou, M.; Moore, S.S.; Guan, L.L. Influence of sire breed on the interplay among rumen microbial populations inhabiting the rumen liquid of the progeny in beef cattle. PLoS ONE 2013, 8, e58461. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, D.M.; Marcondes, M.I.; Renno, L.N.; de Barros, L.V.; Cabral, C.H.A.; Martins, L.S.; Marquez, D.E.C.; Saldarriaga, F.V.; Villadiego, F.A.C.; Cardozo, M.A.; et al. Estimation of daily milk yield of Nellore cows grazing tropical pastures. Trop. Anim. Health Prod. 2018, 50, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Calegare, L.; Alencar, M.M.; Packer, I.U.; Lanna, D.P. Energy requirements and cow/calf efficiency of Nellore and Continental and British Bos taurus x Nellore crosses. J. Anim. Sci. 2007, 85, 2413–2422. [Google Scholar] [CrossRef] [PubMed]
- Mezzadra, C.; Paciaroni, R.; Vulich, S.; Villarreal, E.; Melucci, L. Estimation of milk consumption curve parameters for different genetic groups of bovine calves. Anim. Sci. 1989, 49, 83–87. [Google Scholar] [CrossRef]
- Calegare, L.; Alencar, M.M.; Packer, I.U.; Leme, P.R.; Ferrell, C.L.; Lanna, D.P. Preweaning performance and body composition of calves from straightbred Nellore and Bos taurus x Nellore crosses. J. Anim. Sci. 2009, 87, 1814–1820. [Google Scholar] [CrossRef]
- Xiao, H.; Kang, S. The Role of the Gut Microbiome in Energy Balance With a Focus on the Gut-Adipose Tissue Axis. Front. Genet. 2020, 11, 297. [Google Scholar] [CrossRef] [PubMed]
- Andres, S.; Gini, C.; Ceciliani, F.; Gutierrez-Exposito, D.; Arteche-Villasol, N.; Martin, A.; Cremonesi, P.; Fare, F.; Ghaffari, M.H.; Giraldez, F.J.; et al. Essential oil supplementation in milk replacers: Short- and long-term impacts on feed efficiency, the faecal microbiota and the plasma metabolome in dairy calves. J. Dev. Orig. Health Dis. 2024, 15, e5. [Google Scholar] [CrossRef]
- Shen, Y.; Li, Y.; Wu, T.; Dong, Q.; Deng, Q.; Liu, L.; Guo, Y.; Cao, Y.; Li, Q.; Shi, J.; et al. Early microbial intervention reshapes phenotypes of newborn Bos taurus through metabolic regulations. Gigascience 2024, 13, giad118. [Google Scholar] [CrossRef] [PubMed]
- Alawneh, J.I.; Ramay, H.; Olchowy, T.; Allavena, R.; Soust, M.; Jassim, R.A. Effect of a Lactobacilli-Based Direct-Fed Microbial Product on Gut Microbiota and Gastrointestinal Morphological Changes. Animals 2024, 14, 693. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Fu, Y.; Sun, T.Y.; Jiang, Z.; Miao, Z.; Shuai, M.; Gou, W.; Ling, C.W.; Yang, J.; Wang, J.; et al. The interplay between host genetics and the gut microbiome reveals common and distinct microbiome features for complex human diseases. Microbiome 2020, 8, 145. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Backhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | All Calves | Nellore | Crossbreed |
|---|---|---|---|
| Number of genera/Median (Q1, Q3) | |||
| 5 d | 44.59 a (33.62, 54.72) | 42.11 a (32.69, 50.70) | 45.70 a (33.24, 55.44) |
| 14 d | 53.66 ab (47.59, 57.11) | 51.26 ab (43.57, 58.12) | 54.81 ab (49.14, 57.04) |
| 28 d | 61.17 bc (55.80, 67.45) | 58.77 bcd (48.50, 74.91) | 61.51 abc (58.26, 66.55) |
| 60 d | 61.72 abc (55.12, 65.44) | 59.45 abc (48.59, 64.88) | 62.43 abc (58.97, 65.80) |
| 90 d | 64.47 cd (61.42, 67.88) | 63.88 bcd (59.50, 66.09) | 65.96 bcd (62.60, 70.14) |
| 180 d | 66.61 cde (63.96, 69.05) | 65.21 cd (57.41, 67.95) | 67.78 cd (66.11, 70.00) |
| 245 d | 71.30 de (67.39, 75.16) | 71.45 de (67.22, 73.48) | 71.00 cd (67.33, 75.50) |
| 260 d | 71.80 de (70.13, 74.65) | 71.80 de 68.50, 77.68) | 72.11 cd (70.34, 74.11) |
| Dam | 82.41 e (77.28, 85.37) | 82.41 e (77.27, 85.37) | 82.41 d (77.28, 85.37) |
| Chao/Median (Q1, Q3) | |||
| 5 d | 54.11 a (43.46, 69.26) | 53.48 ac (42.61, 66.96) | 55.62 a (42.61, 72.24) |
| 14 d | 68.21 ab (57.14, 71.91) | 67.83 abcd (53.74, 71.34) | 68.35 ab (57.43, 72.45) |
| 28 d | 78.76 bcd (67.28, 87.61) | 73.41 bcdef (65.26, 94.56) | 80.15 abcd (72.18, 86.55) |
| 60 d | 74.12 abc (67.89, 80.31) | 72.10 cde (61.73, 77.01) | 76.88 abc (70.97, 81.93) |
| 90 d | 81.28 cde (77.78, 89.15) | 80.17 def (66.69, 82.75) | 89.13 bcde (79.68, 89.76) |
| 180 d | 84.29 cdef (80.91, 90.81) | 83.10 ef (76.80, 92.45) | 84.29 bcde (82.32, 88.05) |
| 245 d | 92.06 def (86.61, 94.08) | 91.73 fg (83.67, 93.46) | 92.42 cde (87.70, 98.16) |
| 260 d | 93.16 ef (88.92, 96.12) | 93.16 fg (88.89, 104.40) | 92.69 de (90.72, 95.75) |
| Dam | 105.67 f (100.19, 111.06) | 105.66 g (100.19, 111.06) | 105.67 e (100.19, 111.06) |
| Inverse Simpson/Median (Q1, Q3) | |||
| 5 d | 3.92 a (2.96, 6.85) | 4.96 (2.66, 7.36) | 3.65 a (2.90, 6.34) |
| 14 d | 6.70 ab (4.93, 8.17) | 7.40 (4.94, 8.79) | 6.64 ab (4.33, 7.05) |
| 28 d | 7.86 bc (5.82, 10.89) | 7.41 (3.84, 9.21) | 9.61 b (5.94, 12.37) |
| 60 d | 7.89 bc (5.77, 9.65) | 7.12 (3.81, 9.34) | 7.89 b (6.80, 10.04) |
| 90 d | 8.15 b (7.19, 9.24) | 8.85 (7.17, 11.37) | 8.09 b (6.95, 8.40) |
| 180 d | 6.34 ac (5.51, 6.94) | 5.97 (5.27, 6.73) | 6.68 ab (6.05, 7.04) |
| 245 d | 6.82 ab (6.17, 7.12) | 7.10 * (6.52, 7.48) | 6.32 ab * (5.65, 7.03) |
| 260 d | 6.94 ab (6.64, 7.41) | 6.81 (6.73, 7.08) | 7.10 ab (6.50, 7.51) |
| Dam | 6.84 ab (6.69, 7.24) | 6.84 (6.69, 7.24) | 6.84 ab (6.69, 7.24) |
| Shannon/Median (Q1, Q3) | |||
| 5 d | 1.95 a (1.58, 2.40) | 1.95 a (1.60, 2.41) | 1.96 a (1.40, 2.41) |
| 14 d | 2.46 ab (2.27, 2.57) | 2.50 ab (2.11, 2.60) | 2.42 ab (2.19, 2.54) |
| 28 d | 2.59 b (2.41, 2.85) | 2.56 ab (1.95, 2.75) | 2.66 b (2.45, 3.01) |
| 60 d | 2.62 ab (2.24, 2.71) | 2.51 ab (1.86, 2.72) | 2.68 b (2.39, 2.74) |
| 90 d | 2.56 b (2.48, 2.76) | 2.75 b (2.46, 2.88) | 2.55 ab (2.47, 2.61) |
| 180 d | 2.49 ab (2.35, 2.56) | 2.38 ab (2.31, 2.51) | 2.52 ab (2.41, 2.59) |
| 245 d | 2.54 ab (2.45, 2.57) | 2.55 ab (2.50, 2.61) | 2.49 ab (2.38, 2.56) |
| 260 d | 2.54 b (2.51, 2.60) | 2.53 ab (2.53, 2.58) | 2.58 b (2.49, 2.61) |
| Dam | 2.55 ab (2.52, 2.57) | 2.55 ab (2.52, 2.57) | 2.55 ab (2.52, 2.57) |
| Age | All Calves | Nellore | Crossbreed | NEL vs. CRO (p-Values) |
|---|---|---|---|---|
| Membership (Jaccard) | ||||
| 5 d | A | A | A | 0.827 |
| 14 d | B | A | B | 0.258 |
| 28 d | C | B | C | 0.261 |
| 60 d | D | B | D | 0.005 |
| 90 d | E | C | E | 0.001 |
| 180 d | F | D | F | 0.411 |
| 245 d | G | E | G | 0.149 |
| 260 d | H | F | H | 0.161 |
| Dam | I | G | I | - |
| Structure (Yue and Clayton) | ||||
| 5 d | A | A | A | 0.933 |
| 14 d | A | A | A | 0.773 |
| 28 d | B | AB | B | 0.209 |
| 60 d | C | AB | C | 0.001 |
| 90 d | D | B | D | 0.001 |
| 180 d | E | CD | E | 0.918 |
| 245 d | F | C | F | 0.013 |
| 260 d | G | C | G | 0.015 |
| Dam | EG | D | G | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bessegatto, J.A.; Lisbôa, J.A.N.; Santos, B.P.; Curti, J.M.; Montemor, C.; Alfieri, A.A.; Mach, N.; Costa, M.C. Fecal Microbial Communities of Nellore and Crossbred Beef Calves Raised at Pasture. Animals 2024, 14, 1447. https://doi.org/10.3390/ani14101447
Bessegatto JA, Lisbôa JAN, Santos BP, Curti JM, Montemor C, Alfieri AA, Mach N, Costa MC. Fecal Microbial Communities of Nellore and Crossbred Beef Calves Raised at Pasture. Animals. 2024; 14(10):1447. https://doi.org/10.3390/ani14101447
Chicago/Turabian StyleBessegatto, José Antônio, Júlio Augusto Naylor Lisbôa, Bruna Parapinski Santos, Juliana Massitel Curti, Carlos Montemor, Amauri Alcindo Alfieri, Núria Mach, and Marcio Carvalho Costa. 2024. "Fecal Microbial Communities of Nellore and Crossbred Beef Calves Raised at Pasture" Animals 14, no. 10: 1447. https://doi.org/10.3390/ani14101447
APA StyleBessegatto, J. A., Lisbôa, J. A. N., Santos, B. P., Curti, J. M., Montemor, C., Alfieri, A. A., Mach, N., & Costa, M. C. (2024). Fecal Microbial Communities of Nellore and Crossbred Beef Calves Raised at Pasture. Animals, 14(10), 1447. https://doi.org/10.3390/ani14101447