
Impact of CDK Inhibitors on TBXT Expression in Chordoma Cell Lines Including the First Stable Cell Line of a High-Grade Chordoma

Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of the Chordoma Cell Line U-CH22
2.2. Screening for Mycoplasma Contamination
2.3. Proliferation Measurement
2.4. Library Preparation and Next-Generation Sequencing
2.5. Short Tandem Repeat (STR) Analysis
2.6. Cell Lines
2.7. CDK Inhibition
2.8. Quantitative Polymerase Chain Reaction (qPCR)
2.9. Western Blot Analysis
2.10. Immunocytochemistry and Immunohistochemistry
2.11. Fluorescence In Situ Hybridisation (FISH)
3. Results
3.1. Features of U-CH22
3.1.1. Case Report
3.1.2. Cell Culture
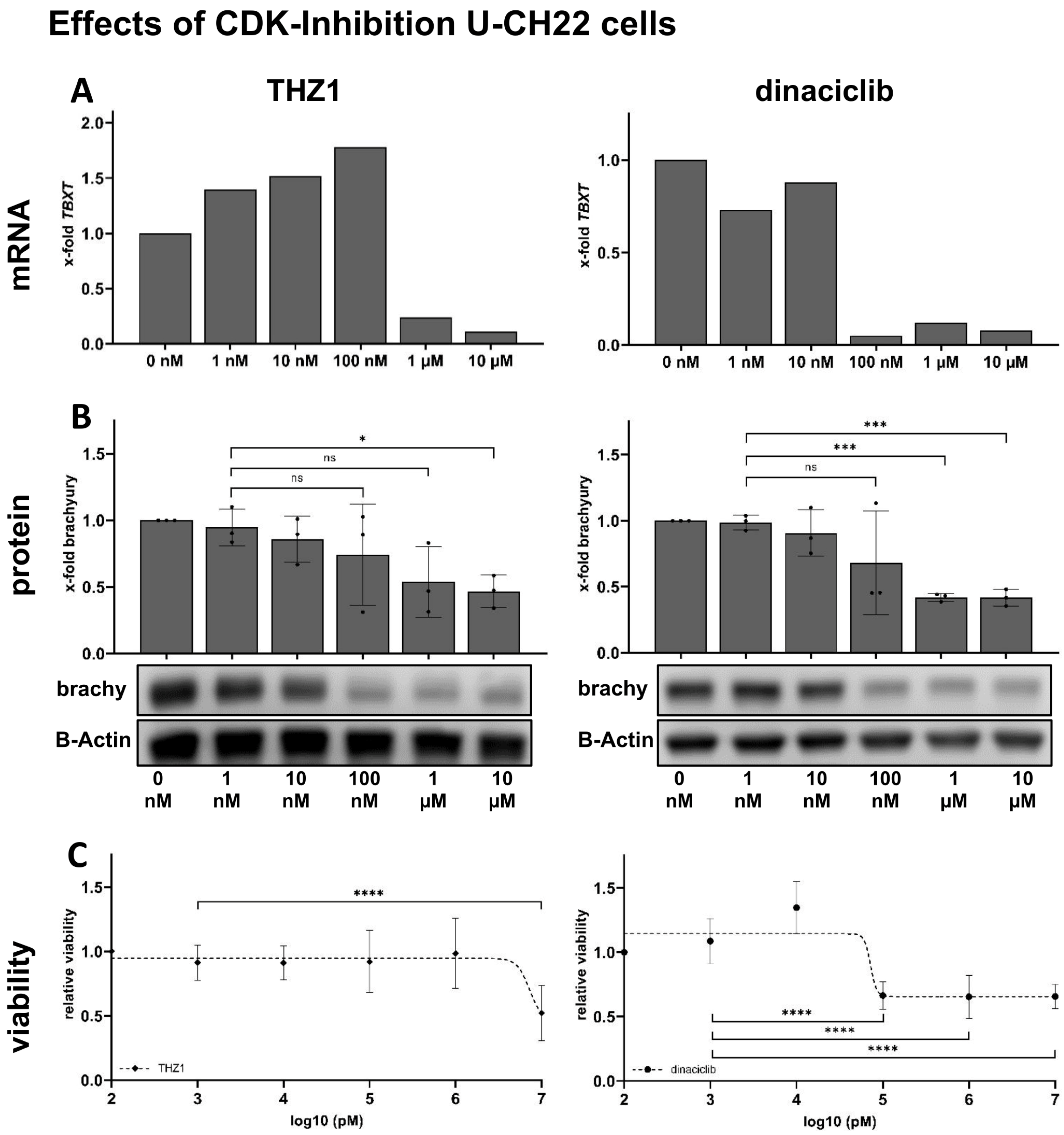
3.2. THZ1 and Dinaciclib Reduce TBXT Expression on mRNA Levels in U-CH22
3.3. THZ1 and Dinaciclib Reduce TBXT Expression on Protein Levels in U-CH22
3.4. THZ1 and Dinaciclib Reduce the Viability of U-CH22
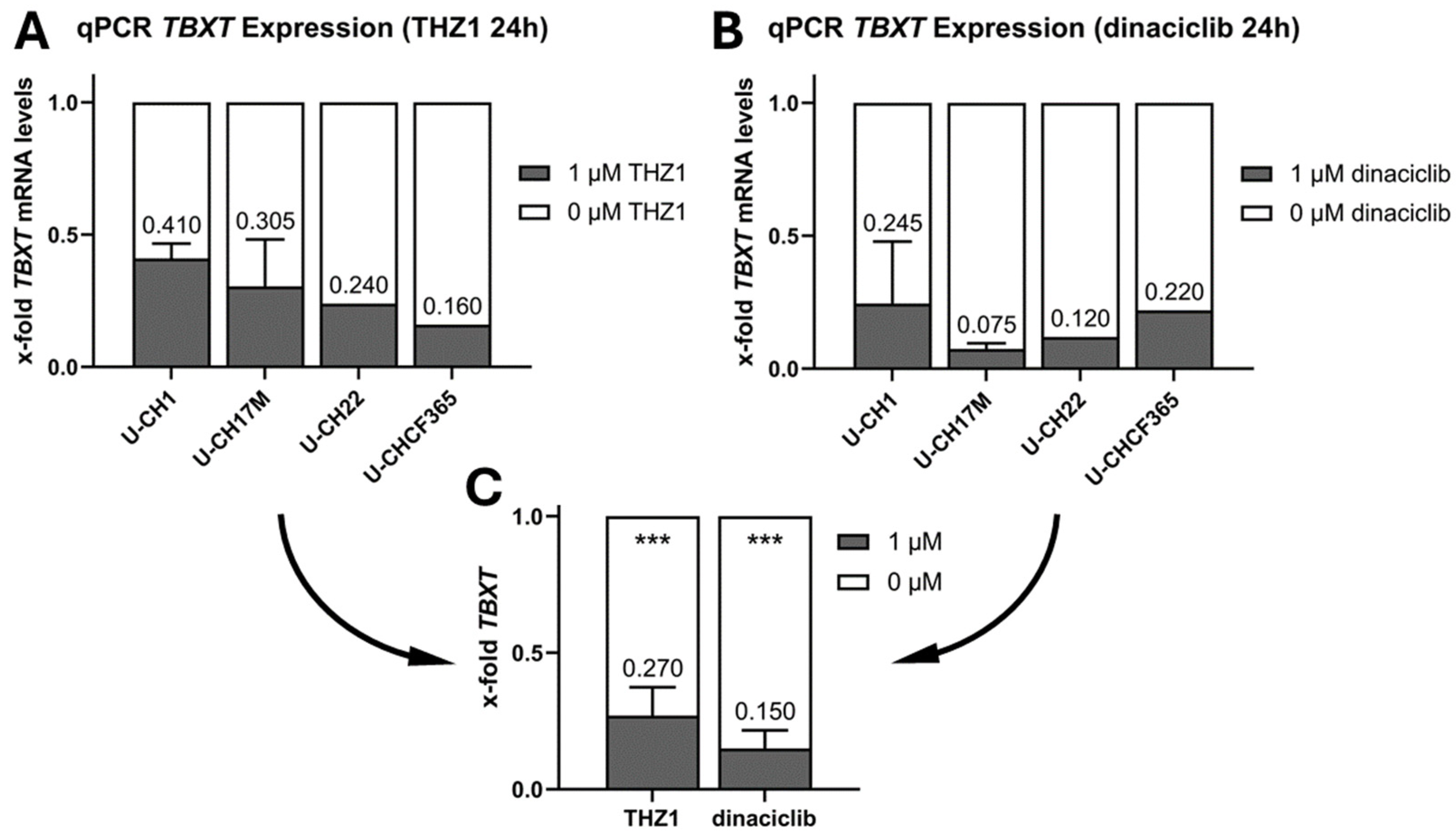
3.5. THZ1 and Dinaciclib Reduce the TBXT Message in Four Cell Culture Models of Progressed Chordoma
3.6. THZ1 and Dinaciclib Reduce Brachyury Levels in Nine Chordoma Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, K.; Yang, H.; Kandimalla, Y.; Liu, J.; Wang, G. Review of Current Treatment of Sacral Chordoma. Orthop. Surg. 2009, 1, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Bakker, S.H.; Jacobs, W.C.H.; Pondaag, W.; Gelderblom, H.; Nout, R.A.; Dijkstra, P.D.S.; Peul, W.C.; Vleggeert-Lankamp, C.L.A. Chordoma: A Systematic Review of the Epidemiology and Clinical Prognostic Factors Predicting Progression-Free and Overall Survival. Eur. Spine J. 2018, 27, 3043–3058. [Google Scholar] [CrossRef] [PubMed]
- Ridenour, R.V.; Ahrens, W.A.; Folpe, A.L.; Miller, D.V. Clinical and Histopathologic Features of Chordomas in Children and Young Adults. Pediatr. Dev. Pathol. 2010, 13, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Barth, T.F.E.; Von Witzleben, A.; Möller, P.; Scheil-Bertram, S. Notochordale Tumoren: Benigne notochordale Tumoren und Chordome. Pathologe 2018, 39, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Coindre, J.-M.; Rivel, J.; Trojani, M.; De Mascarel, I.; De Mascarel, A. Immunohistological Study in Chordomas. J. Pathol. 1986, 150, 61–63. [Google Scholar] [CrossRef] [PubMed]
- Heikinheimo, K.; Persson, S.; Kindblom, L.-G.; Morgan, P.R.; Virtanen, I. Expression of Different Cytokeratin Subclasses in Human Chordoma. J. Pathol. 1991, 164, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, C.D.M.; Bridge, J.M.; Hogendoorn, P.C.W.; Mertens, F. WHO Classification of Tumours of Soft Tissue and Bone. In WHO Classification of Tumours of Soft Tissue and Bone; IARC: Lyon, France, 2013; Volume 5, pp. 328–329. ISBN 978-92-832-2434-1. [Google Scholar]
- Yang, L.; Liu, Y.; Tang, Y.; Wei, J.; Fan, L.; Wang, Y.; Zhang, X.; Cheng, H. Poorly Differentiated Chordoma. Hum. Pathol. Rep. 2022, 28, 300624. [Google Scholar] [CrossRef]
- Park, L.; DeLaney, T.F.; Liebsch, N.J.; Hornicek, F.J.; Goldberg, S.; Mankin, H.; Rosenberg, A.E.; Rosenthal, D.I.; Suit, H.D. Sacral Chordomas: Impact of High-Dose Proton/Photon-Beam Radiation Therapy Combined with or without Surgery for Primary versus Recurrent Tumor. Int. J. Radiat. Oncol.*Biol.*Phys. 2006, 65, 1514–1521. [Google Scholar] [CrossRef]
- Stacchiotti, S.; Longhi, A.; Ferraresi, V.; Grignani, G.; Comandone, A.; Stupp, R.; Bertuzzi, A.; Tamborini, E.; Pilotti, S.; Messina, A.; et al. Phase II Study of Imatinib in Advanced Chordoma. JCO 2012, 30, 914–920. [Google Scholar] [CrossRef]
- Stacchiotti, S.; Sommer, J. Building a Global Consensus Approach to Chordoma: A Position Paper from the Medical and Patient Community. Lancet Oncol. 2015, 16, e71–e83. [Google Scholar] [CrossRef] [PubMed]
- Casali, P.G.; Messina, A.; Stacchiotti, S.; Tamborini, E.; Crippa, F.; Gronchi, A.; Orlandi, R.; Ripamonti, C.; Spreafico, C.; Bertieri, R.; et al. Imatinib Mesylate in Chordoma. Cancer 2004, 101, 2086–2097. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, M.; Wang, Z.; Lasota, J.; Heery, C.; Schlom, J.; Palena, C. Nuclear Brachyury Expression Is Consistent in Chordoma, Common in Germ Cell Tumors and Small Cell Carcinomas, and Rare in Other Carcinomas and Sarcomas: An Immunohistochemical Study of 5229 Cases. Am. J. Surg. Pathol. 2015, 39, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Sharifnia, T.; Wawer, M.J.; Chen, T.; Huang, Q.-Y.; Weir, B.A.; Sizemore, A.; Lawlor, M.A.; Goodale, A.; Cowley, G.S.; Vazquez, F.; et al. Small-Molecule Targeting of Brachyury Transcription Factor Addiction in Chordoma. Nat. Med. 2019, 25, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, H.E.; Dall’Agnese, A.; Park, W.D.; Shamim, M.H.; Dubrulle, J.; Johnson, H.L.; Stossi, F.; Cogswell, P.; Sommer, J.; Levy, J.; et al. Targeted Brachyury Degradation Disrupts a Highly Specific Autoregulatory Program Controlling Chordoma Cell Identity. Cell Rep. Med. 2021, 2, 100188. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Shi, J.; Zhang, Y.; Li, C.; Xiong, Y.; Koka, H.; Wang, D.; Zhang, T.; Song, L.; Luo, W.; et al. Gene Expression Profiling Identifies Two Chordoma Subtypes Associated with Distinct Molecular Mechanisms and Clinical Outcomes. Clin. Cancer Res. 2023, 29, 261–270. [Google Scholar] [CrossRef] [PubMed]
- OMS. Soft Tissue and Bone Tumours, 5th ed.; Organisation Mondiale de la Santé, Centre International de Recherche sur le Cancer, Eds.; World Health Organization Classification of Tumours; OMS: Geneva, Switzerland, 2020; ISBN 978-92-832-4502-5. [Google Scholar]
- Mobley, B.C.; McKenney, J.K.; Bangs, C.D.; Callahan, K.; Yeom, K.W.; Schneppenheim, R.; Hayden, M.G.; Cherry, A.M.; Gokden, M.; Edwards, M.S.B.; et al. Loss of SMARCB1/INI1 Expression in Poorly Differentiated Chordomas. Acta Neuropathol. 2010, 120, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Disease Models. Available online: https://www.chordomafoundation.org/researchers/disease-models/ (accessed on 19 September 2023).
- Lovén, J.; Hoke, H.A.; Lin, C.Y.; Lau, A.; Orlando, D.A.; Vakoc, C.R.; Bradner, J.E.; Lee, T.I.; Young, R.A. Selective Inhibition of Tumor Oncogenes by Disruption of Super-Enhancers. Cell 2013, 153, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, N.; Zhang, T.; Rahl, P.B.; Abraham, B.J.; Reddy, J.; Ficarro, S.B.; Dastur, A.; Amzallag, A.; Ramaswamy, S.; Tesar, B.; et al. Targeting Transcription Regulation in Cancer with a Covalent CDK7 Inhibitor. Nature 2014, 511, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Gnant, M.; Dueck, A.C.; Frantal, S.; Martin, M.; Burstein, H.J.; Greil, R.; Fox, P.; Wolff, A.C.; Chan, A.; Winer, E.P.; et al. Adjuvant Palbociclib for Early Breast Cancer: The PALLAS Trial Results (ABCSG-42/AFT-05/BIG-14-03). J. Clin. Oncol. 2022, 40, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Von Witzleben, A.; Goerttler, L.T.; Marienfeld, R.; Barth, H.; Lechel, A.; Mellert, K.; Böhm, M.; Kornmann, M.; Mayer-Steinacker, R.; Von Baer, A.; et al. Preclinical Characterization of Novel Chordoma Cell Systems and Their Targeting by Pharmocological Inhibitors of the CDK4/6 Cell-Cycle Pathway. Cancer Res. 2015, 75, 3823–3831. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Sharma, S.S.; Ma, L.; Chu, B.; Bui, M.M.; Reed, D.; Pledger, W.J. Apoptosis of Osteosarcoma Cultures by the Combination of the Cyclin-Dependent Kinase Inhibitor SCH727965 and a Heat Shock Protein 90 Inhibitor. Cell Death Dis. 2013, 4, e566. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.K.; LaPlant, B.; Chng, W.J.; Zonder, J.; Callander, N.; Fonseca, R.; Fruth, B.; Roy, V.; Erlichman, C.; Stewart, A.K. Dinaciclib, a Novel CDK Inhibitor, Demonstrates Encouraging Single-Agent Activity in Patients with Relapsed Multiple Myeloma. Blood 2015, 125, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Mita, M.M.; Joy, A.A.; Mita, A.; Sankhala, K.; Jou, Y.-M.; Zhang, D.; Statkevich, P.; Zhu, Y.; Yao, S.-L.; Small, K.; et al. Randomized Phase II Trial of the Cyclin-Dependent Kinase Inhibitor Dinaciclib (MK-7965) versus Capecitabine in Patients with Advanced Breast Cancer. Clin. Breast Cancer 2014, 14, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Marineau, J.J.; Rajagopal, N.; Hamman, K.B.; Choi, Y.J.; Schmidt, D.R.; Ke, N.; Johannessen, L.; Bradley, M.J.; Orlando, D.A.; et al. Discovery and Characterization of SY-1365, a Selective, Covalent Inhibitor of CDK7. Cancer Res. 2019, 79, 3479–3491. [Google Scholar] [CrossRef] [PubMed]
- Sava, G.P.; Fan, H.; Coombes, R.C.; Buluwela, L.; Ali, S. CDK7 Inhibitors as Anticancer Drugs. Cancer Metastasis Rev. 2020, 39, 805–823. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Kretz, A.-L.; Schneider, S.; Knippschild, U.; Henne-Bruns, D.; Kornmann, M.; Lemke, J.; Traub, B. Evaluation of CDK9 Inhibition by Dinaciclib in Combination with Apoptosis Modulating izTRAIL for the Treatment of Colorectal Cancer. Biomedicines 2023, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Ghia, P.; Scarfò, L.; Perez, S.; Pathiraja, K.; Derosier, M.; Sisk, C.M.; Patton, N. Efficacy and Safety of Dinaciclib versus Ofatumumab in Patients with Relapsed/Refractory Chronic Lymphocytic Leukemia. J. Am. Soc. Hematol. 2017, 129, 1876–1878. [Google Scholar]
- Scheipl, S.; Barnard, M.; Cottone, L.; Jorgensen, M.; Drewry, D.H.; Zuercher, W.J.; Turlais, F.; Ye, H.; Leite, A.P.; Smith, J.A.; et al. EGFR Inhibitors Identified as a Potential Treatment for Chordoma in a Focused Compound Screen. J. Pathol. 2016, 239, 320–334. [Google Scholar] [CrossRef]
- Beisaw, A.; Tsaytler, P.; Koch, F.; Schmitz, S.U.; Melissari, M.; Senft, A.D.; Wittler, L.; Pennimpede, T.; Macura, K.; Herrmann, B.G.; et al. BRACHYURY Directs Histone Acetylation to Target Loci during Mesoderm Development. EMBO Rep. 2018, 19, 118–134. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Species | Tumour | Localisation | Status | Patient Age | Origin |
|---|---|---|---|---|---|---|
| U-CH1 | Human | Chordoma | sacral | recurrence | 56 | University Hospital Ulm (Germany) |
| U-CH2 | Human | Chordoma | sacral | recurrence | 72 | University Hospital Ulm (Germany) |
| U-CH17PII | Human | Chordoma | sacral | primary | 38 | University Hospital Ulm (Germany) |
| U-CH17M | Human | Chordoma | sacral | metastasis | 38 | University Hospital Ulm (Germany) |
| U-CH22 | Human | Chordoma | pleural effusion | recurrence + pan-metastasised | 78 | University Hospital Ulm (Germany) |
| U-CHCF365 | Human | Chordoma | clival | Xenograft model of metastasis | <20 | University Hospital Ulm (Germany) |
| UM-Chor1 | Human | Chordoma | clival | primary | 66 | Michigan University (USA) |
| MUG-Chor1 | Human | Chordoma | sacral | recurrence | 57 | Graz University (Austria) |
| MUG-CC1 | Human | Chordoma | clival | primary | 72 | Graz University (Austria) |
| Antibody Name | Manufacturer (Reference No.:) | Species | Dilution | Pretreatment |
|---|---|---|---|---|
| brachyury | Abcam (ab209665) | rabbit | 1:4000 | Steamer; in EDTA pH 8 |
| CDK4 | Zytomed (A03-61096) | mouse | 1:50 | Steamer; in EDTA pH 8 |
| CDK6 | LS Bio (LS-B5388) | mouse | 1:100 | Pressure cooker |
| EMA (Clone E29) | Dako (M0613) | mouse | 1:500 | Pronase |
| INI-1 (Clone 25/BAF47) | BD Bioscience (612111) | mouse | 1:50 | Pressure cooker |
| KI-67 (Clone MIB-1) | Dako (M7240) | mouse | 1:200 | Pressure cooker |
| p53 (Clone DO-7) | Dako (M7001) | mouse | 1:400 | Microwave; in citric acid pH 6.1 |
| pan-cytokeratin (Clone CK AE1/AE3) | Dako (M3515) | mouse | 1:100 | Pronase |
| S100 (Clone Polyclonal) | Dako (GA504) | rabbit | 1:1000 | Pronase |
| vimentin (Clone Vim 3B4) | Dako (M7020) | mouse | 1:300 | Microwave; in citric acid pH 6.1 |
| Molarity of CDK Inhibitors to Reach the Half-Maximal Amount of Brachyury Protein | ||
|---|---|---|
| THZ1 [nM] | Dinaciclib [nM] | |
| U-CH1 | 979 | 10 |
| U-CH2 | 272 | 8 |
| U-CH17M | 333 | 11 |
| U-CH17PII | 918 | 10 |
| U-CH22 | 132 | 83 |
| U-CHCF365 | 11 | 11 |
| UM-Chor1 | 308 | 11 |
| MUG-Chor1 | 326 | 10 |
| MUG-CC1 | 22 | 87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bette, S.; Haase, L.; Nell, J.; Grieser, T.; von Baer, A.; Schultheiss, M.; Marienfeld, R.; Möller, P.; Barth, T.F.E.; Mellert, K. Impact of CDK Inhibitors on TBXT Expression in Chordoma Cell Lines Including the First Stable Cell Line of a High-Grade Chordoma. Diagnostics 2024, 14, 1028. https://doi.org/10.3390/diagnostics14101028
Bette S, Haase L, Nell J, Grieser T, von Baer A, Schultheiss M, Marienfeld R, Möller P, Barth TFE, Mellert K. Impact of CDK Inhibitors on TBXT Expression in Chordoma Cell Lines Including the First Stable Cell Line of a High-Grade Chordoma. Diagnostics. 2024; 14(10):1028. https://doi.org/10.3390/diagnostics14101028
Chicago/Turabian StyleBette, Sarah, Luisa Haase, Juliane Nell, Thomas Grieser, Alexandra von Baer, Markus Schultheiss, Ralf Marienfeld, Peter Möller, Thomas F. E. Barth, and Kevin Mellert. 2024. "Impact of CDK Inhibitors on TBXT Expression in Chordoma Cell Lines Including the First Stable Cell Line of a High-Grade Chordoma" Diagnostics 14, no. 10: 1028. https://doi.org/10.3390/diagnostics14101028