
Modeling of Water, Heat, and Nitrogen Dynamics in Summer Maize under Broad Furrow Irrigation and the Mechanism of Enzyme Activity Response
Abstract
1. Introduction
2. Materials and Methods
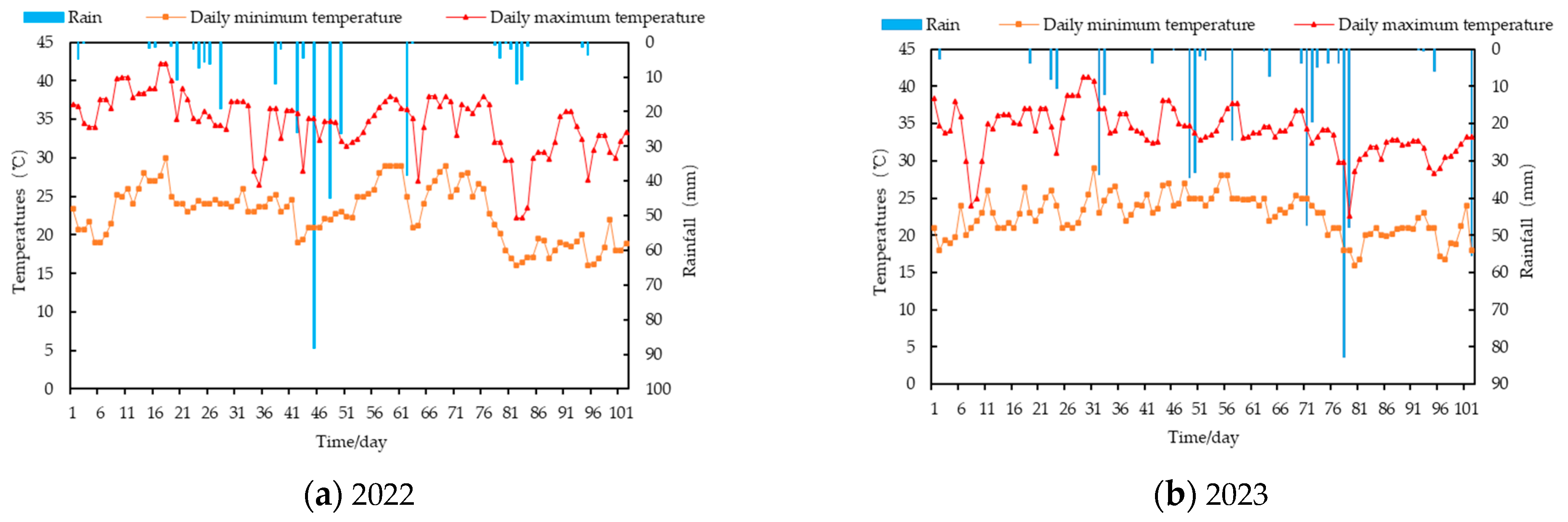
2.1. Overview and Soil Characteristics of the Study Area
2.2. Experimental Design
2.3. Measurement Items and Methods
2.3.1. Soil Water Content
2.3.2. Soil Temperature
2.3.3. Soil Ammonium Nitrogen and Nitrate Nitrogen Contents
2.3.4. Soil Enzyme Activities
2.3.5. Summer Maize Yield and Components
2.4. Model Construction
2.4.1. Governing Equation
2.4.2. Model Parameter
2.4.3. Initial and Boundary Conditions
- Initial conditions
- 2.
- Boundary conditions
3. Results
3.1. Model Validation
3.2. Distribution Patterns and Modeling of Hydrothermal Nitrogen in Soil Profiles
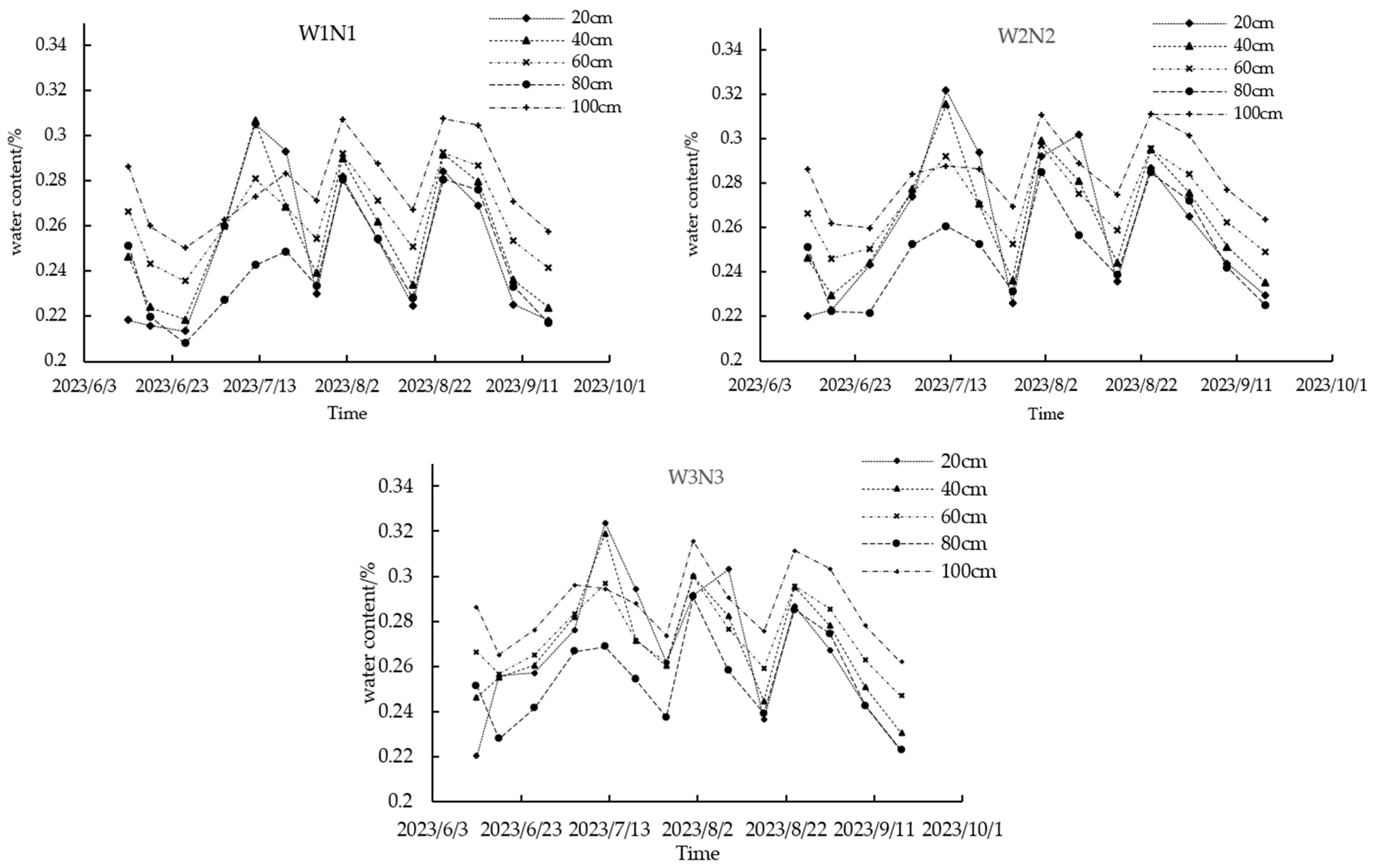
3.2.1. Soil Profile’s Water Distribution Pattern
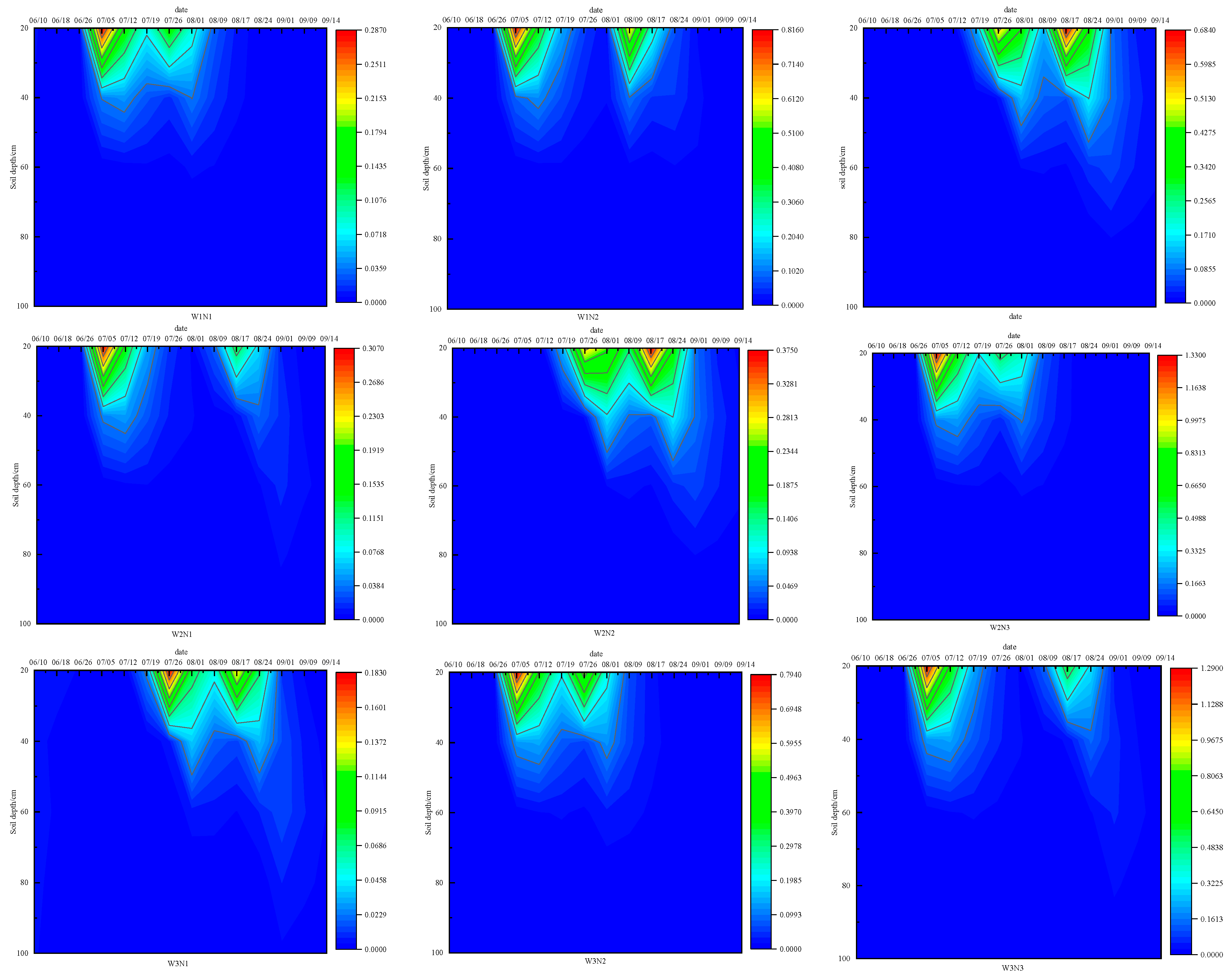
3.2.2. Characteristics of Ammonium Nitrogen Distribution in Soil Profiles
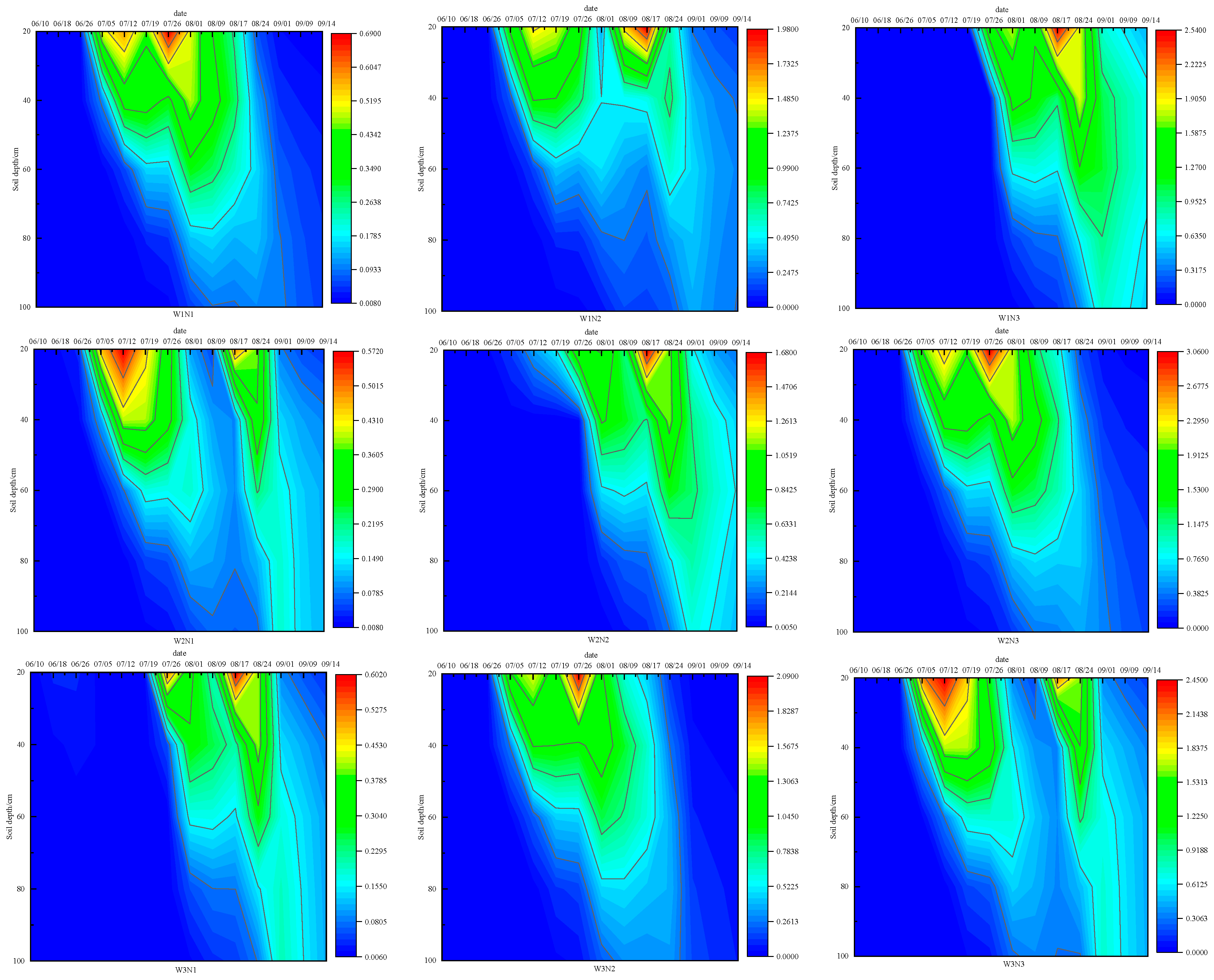
3.2.3. Characteristics of Nitrate Nitrogen Distribution in Soil Profiles
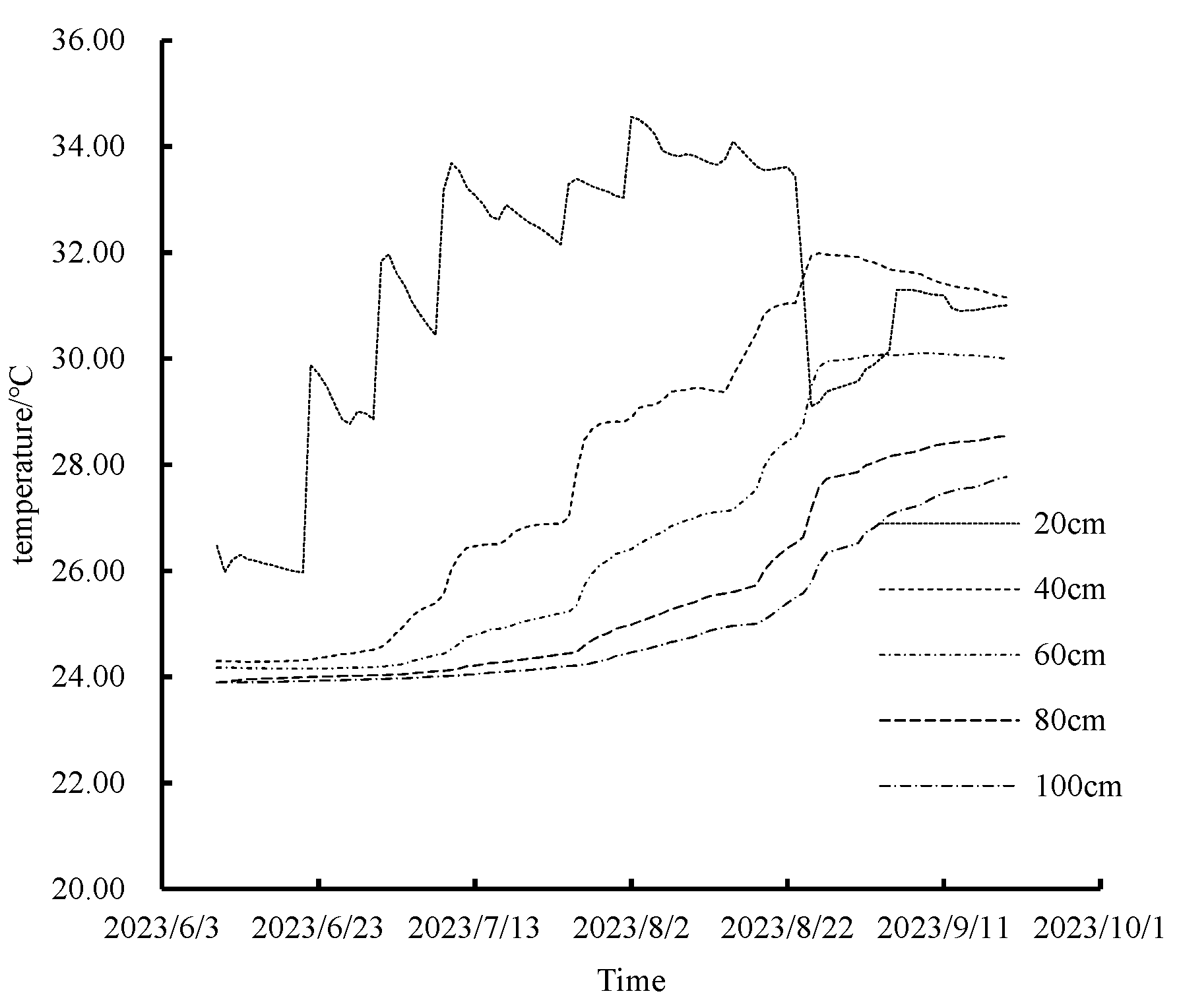
3.2.4. Analysis and Modeling of Temperature Dynamic Patterns in Soil Profiles
3.3. Enzyme Activity Response Mechanisms under Different Water and Nitrogen Application Regimes
3.4. Effect of Water–Nitrogen Coupling on Yield and Components of Summer Maize
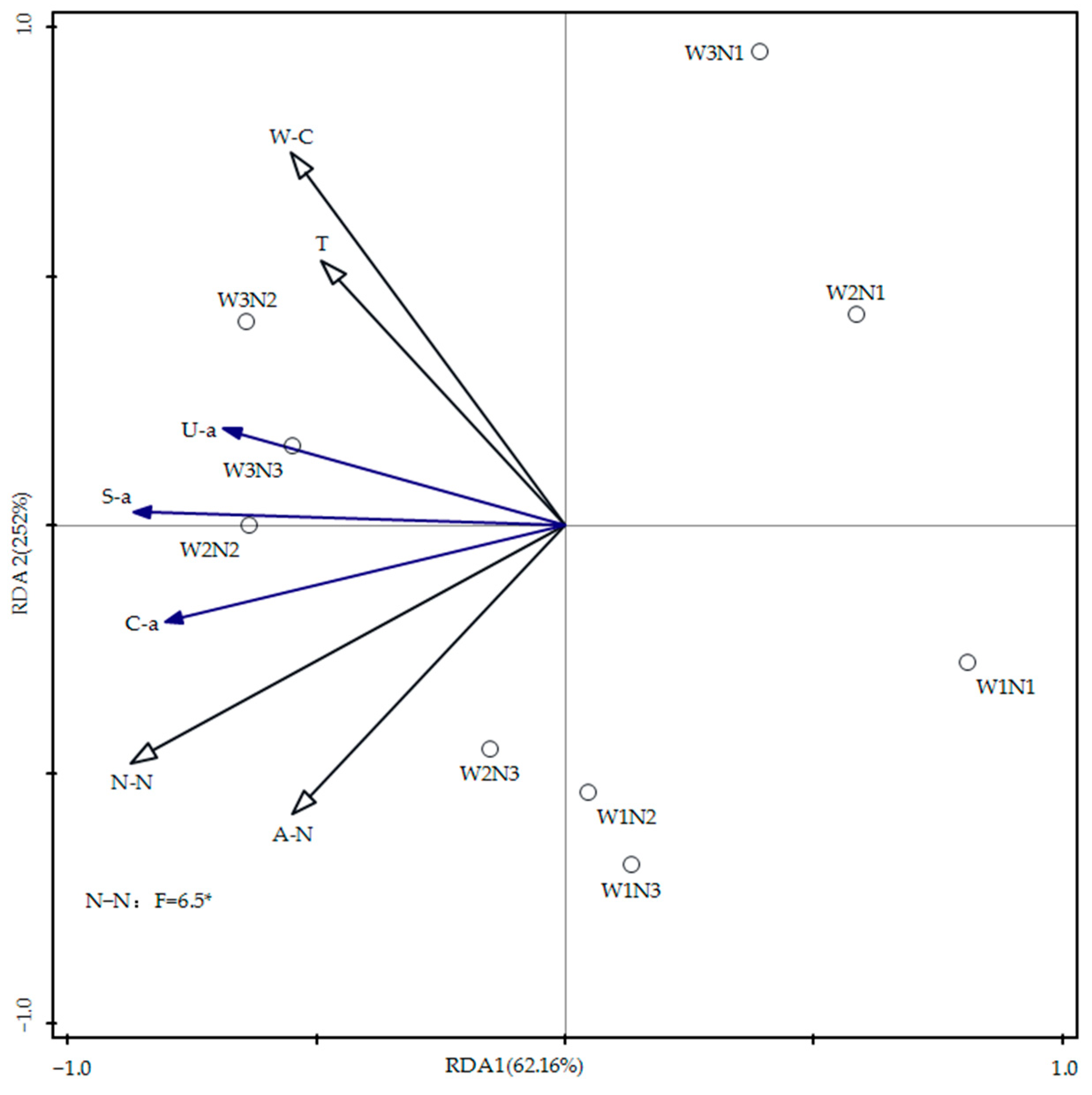
3.5. Redundancy Analysis of Soil Moisture, Nitrogen, Temperature, and Enzyme Activity
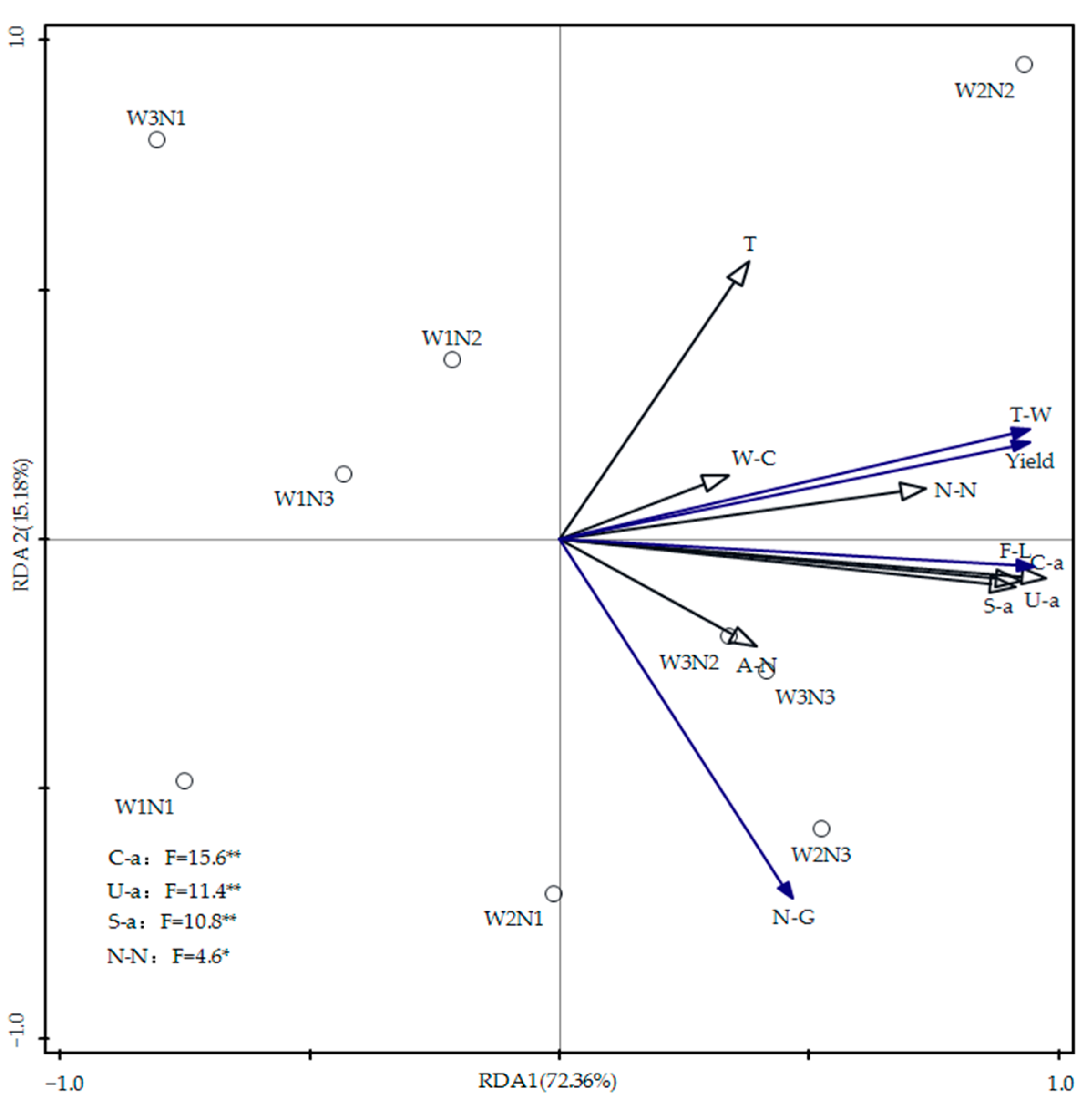
3.6. Redundancy Analysis of Soil Moisture, Nitrogen, Temperature, Enzyme Activity, and Summer Maize Yield
4. Discussion
5. Conclusions
- (1)
- The rate-optimized HYDRUS model effectively simulated soil water, heat, and nitrogen dynamics, with all R2 not falling below 0.7. The model proved to be suitable for the simulation of water movement, nitrogen transport transformation, and heat transport.
- (2)
- This study identified a positive correlation between soil hydrothermal nitrogen dynamics and enzyme activities, with increased water–nitrogen application enhancing soil enzyme activities. Soil urease, catalase, sucrase activities, and nitrate nitrogen concentration emerged as the primary factors influencing variations in summer maize yields and constituent elements, demonstrating significant positive correlations. Based on the analysis of the effects of water and nitrogen application on summer maize yield and soil properties, the W2N2 regime was determined to be the optimal water and nitrogen application system.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, Y.; Liu, Y.; Yin, J.; Chang, Z.; Wang, Y.; Liu, L.; Tian, Y.; Chen, W.; Wang, L.; Hao, L. Effects of moisture and CO2 on photosynthetic performance and water utilisation in maize. J. Agric. Eng. 2023, 39, 71–81. [Google Scholar]
- Teng, Y.; Kong, F.; Chen, Y.; Chen, S.; Xiong, H.; Zhu, K.; Yang, Z. Meta-analysis of the effects of combined application of organic and chemical fertilizers on soil nitrogen leaching. Trans. Chin. Soc. Agric. Eng. 2022, 38, 81–88. [Google Scholar]
- Xu, R.; Shi, J.; Hao, D.; Ding, Y.; Gao, J. Research on temporal and spatial differentiation and impact paths of agricultural grey water footprints in the Yellow River Basin. Water 2022, 14, 2759. [Google Scholar] [CrossRef]
- Qing, Y.; Zhao, B.; Wen, C. The Coupling and Coordination of Agricultural Carbon Emissions Efficiency and Economic Growth in the Yellow River Basin, China. Sustainability 2023, 15, 971. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, Z.; Sun, D.; Song, J.; Zhang, Z.; Li, T. Effect of water and nitrogen coupling on rice yield and nitrogen absorption and utilization in black soil. Trans. Chin. Soc. Agric. Mach. 2021, 52, 324–335, 357. [Google Scholar]
- Gu, X.; Li, Y.; Huang, P.; Du, Y.; Fang, H.; Chen, P. Effects of planting patterns and nitrogen application rates on yield, water and nitrogen use efficiencies of winter oilseed rape. Trans. Chin. Soc. Agric. Eng. 2018, 34, 113–123. [Google Scholar]
- Gao, S.; Liu, T.; Wang, S.; Li, Y.; Ding, J.; Liu, Y.; Wang, D.; Li, H. Optimizing Fertilizer Management Practices in Summer Maize Fields in the Yellow River Basin. Agronomy 2023, 13, 2236. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, W.; Song, X.; Liu, P.; Sun, R.; Yue, X. Analysis of the dynamic evolution of soil quality in reclaimed abandoned salt fields in the Yellow River Delta. J. Appl. Basic Eng. Sci. 2021, 29, 545–561. [Google Scholar]
- Liu, S.; Xia, X.; Chen, G.; Mou, D.; Che, S.; Li, Y. Research progress of soil enzymes. Chin. Agron. Bull. 2011, 27, 1–7. [Google Scholar]
- Li, Y.; Niu, W.; Dyck, M.; Wang, J.; Zou, X. Yields and Nutritional of Greenhouse Tomato in Response to Different Soil Aeration Volume at two depths of Subsurface drip irrigation. Sci. Rep. 2016, 6, 39307. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Shouliang, Y.; Yang, X.; Li, J.; Hai, Y.; Zi, L.; Fan, K.; Pan, G. The Richness and Diversity of Catalases in Bacteria. Front. Microbiol. 2021, 12, 645477. [Google Scholar]
- Li, T.F.; Liu, C.Y.; Jin, X.M.; Cao, X. Effects of different cultivation strategies on soil nutrients and bacterial diversity in kiwifruit orchards. Eur. J. Hortic. Sci. 2022, 87, 1–8. [Google Scholar]
- Kang, N.; Siegfried, A.P. Soil Moisture and Temperature Effects on Granule Dissolution and Urease Activity of Urea with and without Inhibitors—An Incubation Study. Agriculture 2022, 12, 2037. [Google Scholar] [CrossRef]
- Ren, J.; Liu, X.; Han, F.; Kong, F.; Li, J.; Peng, H.; Li, Q. Effects of nitrogen application levels on soil enzyme activities and fruit quality in dry loess sand-covered apple orchards. J. Agric. Eng. 2019, 35, 206–213. [Google Scholar]
- Mi, G.; Yuan, L.; Gong, Y.; Zhang., F.; Ren., H. Effects of different water and nitrogen supply on soil enzyme activity and biological environment of tomato in solar greenhouse. J. Agric. Eng. 2005, 21, 124–127. [Google Scholar]
- Xiao, X.; Zhu, W.; Xiao, L.; Deng, Y.; Zhao, Y.; Wang, J. Improvement of soil enzyme activity and soil microbiota carbon and nitrogen in rice-based agricultural fields by appropriate water and nitrogen treatments. J. Agric. Eng. 2013, 29, 91–98. [Google Scholar]
- Wang, Y.; Mao, X.; Chen, S.; Bao, L. Experimentation and simulation of soil hydrothermal salinity dynamics and growth of oil sunflower in farmland irrigated by brackish water border. J. Agric. Eng. 2021, 37, 76–86. [Google Scholar]
- Su, Y.; Guo, X.; Hu, F.; Sun, X.; Ma, J.; Lei, T. Numerical simulation of soil water-heat-oxygen distribution in orchards under pit irrigation. J. Irrig. Drain. 2023, 42, 67–74. [Google Scholar]
- Wang, G.; Shi, H.; Li, X.; Yan, J.; Miao, Q.; Chen, N.; Wang, W. Simulation and assessment of water and salt transport in desert oasis based on HYDRUS-1D model. J. Agric. Eng. 2021, 37, 87–98. [Google Scholar]
- Šimůnek, J.I.R.K.A.; Van Genuchten, M.T.; Šejna, M. The HYDRUS Software Package for Simulating Two- and Three Dimensional Movement of Water, Heat, and Multiple Solutes in Variably-Saturated Porous Media; Technical Manual, Version 3.0; PC Progress: Prague, Czech Republic, 2020. [Google Scholar]
- Tamás, M.; Zsolt, F.; Erika, B.; János, T.; Attila, N. Modeling of soil moisture and water fluxes in a maize field for the optimization of irrigation. Comput. Electron. Agric. 2023, 213, 108159. [Google Scholar]
- Balkhi, A.; Ebrahimian, H.; Ghameshlou, A.N.; Amin, M. Modeling of nitrate and ammonium leaching and crop uptake under wastewater application considering nitrogen cycle in the soil. Model. Earth Syst. Environ. 2022, 9, 901–911. [Google Scholar] [CrossRef]
- Farshad, K.; Javad, B.; Vahid, R.; Nasrin, A. Field evaluation and numerical simulation of water and nitrate transport in subsurface drip irrigation of corn using HYDRUS-2D. Irrig. Sci. 2023, 42, 327–352. [Google Scholar]
- Nasrin, A.; Javad, B.; Vahid, R.; Habib, K.; Sally, E.; Dirk, M.; Tiago, B.; He, H. CNN deep learning performance in estimating nitrate uptake by maize and root zone losses under surface drip irrigation. J. Hydrol. 2023, 625, 130148. [Google Scholar]
- Guan, S. Soil Enzymes and Their Research Methods; Agricultural Press: Beijing, China, 1986. [Google Scholar]
- Schaap, M.G.; Van Genuchten, M.T. A Modified Mualem–van Genuchten Formulation for Improved Description of the Hydraulic Conductivity Near Saturation. Vadose Zone J. 2006, 5, 27–34. [Google Scholar] [CrossRef]
- Wang, S.; Liu, T.; Yang, J.; Wu, C.; Zhang, H. Simulation Effect of Water and Nitrogen Transport under Wide Ridge and Furrow Irrigation in Winter Wheat Using HYDRUS-2D. Agronomy 2023, 13, 457. [Google Scholar] [CrossRef]
- Feddes, R.A.; Kowalik, P.J.; Zaradny, H. Simulation of Field Water Use and Crop Yield; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1978. [Google Scholar]
- Monteith, J.L. Principles of environmental physics. Plant Growth Regul. 1991, 10, 177–178. [Google Scholar] [CrossRef]
- Sophocleous, L. Analysis of water and heat flow in unsaturated-saturated porous media. Water Resour. Res. 1979, 15, 1195–1206. [Google Scholar] [CrossRef]
- Chung, O.; Horton, R. Soil heat and water flow with a partial surface mulch. Water Resour. 1987, 23, 2175–2186. [Google Scholar] [CrossRef]
- Chen, S.; Huang, D.; Zhang, E.; Zhu, J.; Guo, X. Effects of straw compartmentalisation and water-nitrogen management on soil inorganic nitrogen and enzyme activities. J. Irrig. Drain. 2022, 41, 45–54. [Google Scholar]
- Wang, Q.; Wang, W.; Wang, Z.; Zhang, J.; Li, Y. Determination of technique parameters for saline-alkali soil through drip irrigation under film. Trans. Chin. Soc. Agric. Eng. 2001, 17, 47–50. [Google Scholar]
- Shi, X.; Wang, Q.; Su, Y. Effects of slight saltwater quality on the characteristics of soil water and salt transference. Arid Land Geogr. 2005, 28, 100–104. [Google Scholar]
- Lei, C.D.; Yang, S.X.; Xie, S.C. Soil Hydrodynamics; Tsinghua University Press: Beijing, China, 1988. [Google Scholar]
- Jin, Y.; Li, X.; Cai, Y.; Hu, H.; Liu, Y.; Fu, S.; Zhang, B. Effects of straw return with fertiliser on enzyme activity and microbial community structure in rice-oil rotation soil. Environ. Sci. 2021, 42, 3985–3996. [Google Scholar]
- Wei, W.; Dai, F.; Zhang, F.; Zhang, S.; Shi, R.; Liu, Y.; Shen, P. Agronomic optimisation of dryland maize in full-film twomonade furrow based on numerical simulation of hydrothermal effect. J. China Agric. Univ. 2020, 25, 19–28. [Google Scholar]
- Song, C.; Wang, Y. Characteristics of soil temperature in wetland ecosystems in response to air temperature and its effect on CO2 emission. J. Appl. Ecol. 2006, 17, 4625–4629. [Google Scholar]
- Li, M.; Hu, T.; Cui, X.; Luo, L.; Lu, J. Comprehensive effects of irrigation water and nitrogen levels for controlled release fertilizer with different release periods on winter wheat yield. Trans. Chin. Soc. Agric. Eng. 2020, 36, 153–161. [Google Scholar]
- Wang, H.; Su, W.; Zhao, X.; Aimulaguli, K.; Sun, S.; Xue, L.; Zhang, J. Effect of different irrigation and nitrogen application on yield, water and nitrogen use efficiency of spring wheat sown in winter. J. Triticeae Crops 2022, 42, 1381–1390. [Google Scholar]
- Mi, X.; Shi, L.; He, G.; Wang, Z. Fertilizer reduction potential and economic benefits of crop production for smallholder farmers in Shaanxi Province. Sci. Agric. Sin. 2021, 54, 4370–4384. [Google Scholar]
- Zhai, Y.Q.; Ma, K.; Jia, B.; Wei, X.; Yun, B.; Ma, J.; Zhang, H.; Ji, L.; Li, J. Distribution of nitrate nitrogen in drip-irrigated maize soils with different precipitation years, loss of nitrogen, and nitrogen uptake and utilisation. Chin. J. Ecol. Agric. 2023, 31, 765–775. [Google Scholar]
- Xu, G.; Lu, D.; Sun, H.; Wang, H.; Li, Y. Effects of coupled dry and wet alternate irrigation and nitrogen application on the inter-root environment of rice. J. Agric. Eng. 2017, 33, 186–194. [Google Scholar]
- Shi, X.; De, Y.; Ding, M.; Liu, X.; Wang, Z.; Liu, Y.; Xiang, P.; Yang, Q. Characteristics of soil carbon and nitrogen content, enzyme activity and yield of Panax pseudoginseng under water and fertiliser control. Agric. Res. Arid Reg. 2024, 42, 76–86. [Google Scholar]
- Treseder, K.K. Nitrogen additions and microbial biomass: Ameta analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Wang, K.Y.; Wang, H.R.; Zhao, M.D.; Zhou, Y.K. Nutrients and ecological stoichiometry characteristics of typical wetland soils in the lower Yellow River. Environ. Sci. 2024, 45, 1674–1683. [Google Scholar]
- Yu, Y.H.; Li, Y.T.; Wu, Y.G.; Wang, X.J.; Song, Y.P.; Deng, X.H. Effects of soil and leaf traits on peel quality of Zanthoxylum planispinum ‘dintanensis’. J. For. Environ. 2023, 43, 8–16. [Google Scholar]
- Zhai, C.; Feng, X.; Liu, C.; Li, Y.; Fan, J.; Zhang, J.; Meng, Q. Linkages of Enzymatic Activity and Stoichiometry with Soil Physical-Chemical Properties under Long-Term Manure Application to Saline-Sodic Soil on the Songnen Plain. Agronomy 2023, 13, 2972. [Google Scholar] [CrossRef]
- Li, Y.; Han, T.; Liu, K.; Huang, T.; Han, D.; Xiao, G.; Zhang, W. Response of soil enzyme activity and rice yield to tilling of winter green manure in a red soil paddy field. J. Plant Nutr. Fertil. 2023, 29, 1313–1322. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Depth (cm) | Characteristic Parameters of the Soil | Particle Size Composition (%) | Physicochemical Parameters of the Tested Soil | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Dry Bulk Weight of the Soil (g/cm3) | Soil Field Capacity (V/V%) | Soil Organic Matter (m/m%) | Average Content of Total Nitrogen (%) | Clay | Silt | Sand | Ammonium Nitrogen (mg·kg−1) | Nitrate Nitrogen (mg·kg−1) | |
| 0–20 | 1.46 | 33.3 | 0.83 | 0.04 | 7 | 42 | 51 | 9.52 | 7.95 |
| 20–40 | 1.47 | 34.2 | 0.86 | 0.05 | 7 | 43 | 50 | 7.85 | 7.64 |
| 40–60 | 1.47 | 31.6 | 0.82 | 0.03 | 6 | 44 | 49 | 8.46 | 6.35 |
| 60–80 | 1.46 | 32.5 | 0.58 | 0.02 | 6 | 46 | 48 | 6.89 | 6.12 |
| 80–100 | 1.48 | 32.4 | 0.52 | 0.01 | 2 | 15 | 83 | 6.31 | 6.48 |
| Treatment | Irrigation Scheme | Number of Topdressing (Times) | Base Fertilizer | Nitrogen Application Rate (kg·ha−1) | ||||
|---|---|---|---|---|---|---|---|---|
| Lower Limit of Irrigation | Irrigation Norm (mm) | Jointing Stage | Trumpet Stage | Flowering Stage | Footing | |||
| W1N1 | 60% of θs | 30 | 2 | 55 | 32.5 | 32.5 | / | 120 |
| W1N2 | 60% of θs | 82.5 | / | 82.5 | 120 | |||
| W1N3 | 60% of θs | / | 132.5 | 132.5 | 120 | |||
| W2N1 | 70%of θs | 32.5 | / | 32.5 | 220 | |||
| W2N2 | 70%of θs | / | 82.5 | 82.5 | 220 | |||
| W2N3 | 70%of θs | 132.5 | 132.5 | / | 220 | |||
| W3N1 | 80%of θs | / | 32.5 | 32.5 | 320 | |||
| W3N2 | 80%of θs | 82.5 | 82.5 | / | 320 | |||
| W3N3 | 80%of θs | 132.5 | / | 132.5 | 320 | |||
| Phenological Period | 2022 | 2023 | Characteristics |
|---|---|---|---|
| Emergence | 6/8–6/17 | 6/10–6/20 | The germinal sheath is exposed to the ground. |
| First leaf | 6/15–6/26 | 6/19–6/30 | First leaf fully expanded. |
| Third leaf | 6/23–7/5 | 6/27–7/8 | The third leaf is fully expanded, at which point the growing point of the corn is still underground. |
| Sixth leaf | 7/3–7/15 | 7/4–7/17 | The sixth leaf is fully expanded, and the male spike cone begins to elongate. |
| Twelfth leaf | 7/14–8/3 | 7/15–8/1 | |
| Tasseling | 7/29–8/10 | 7/26–8/12 | The last branch of the male spike is visible before silking. |
| Silking | 8/5–8/13 | 8/6–8/15 | Filaments of female spikes begin to show bracts. |
| Blister stage | 8/11–8/25 | 8/12–8/27 | The volume of the kernel in the middle of the cob is basically built up and the endosperm is clear and pulpy. |
| Milk stage | 8/23–9/10 | 8/24–9/11 | Maize kernels turn yellow, and the endosperm is milky then mushy. |
| Physiological maturity | 9/7–9/17 | 9/8–9/19 | Plants with dry, hard kernels; black layer at the base of the kernel; disappearance of the milkline. |
| Soil Depth (cm) | θr (cm3·cm−3) | θs (cm3·cm−3) | α (cm−1) | n | Ks (cm·day−1) |
|---|---|---|---|---|---|
| 0–20 | 0.031 | 0.3832 | 0.0143 | 1.3531 | 37.7 |
| 20–40 | 0.3895 | 0.0134 | 1.2786 | 29.8 | |
| 40–60 | 0.4032 | 0.0139 | 1.4957 | 32.1 | |
| 60–80 | 0.3837 | 0.0147 | 1.3854 | 33.9 | |
| 80–100 | 0.3843 | 0.0139 | 1.5295 | 42.2 |
| Soil Depth (cm) | (d) | Kd (cm3·mg−1) | (d) | (d) | (d) | (d) | (day−1) | (mg·cm−3·d−1) |
|---|---|---|---|---|---|---|---|---|
| 20 | 0.56 | 0.0032 | 0.02 | 0.2 | 0.2 | 0.04 | 0.04 | 3 × 10−5 |
| 40 | 0.55 | 0.0035 | 0.025 | 0.3 | 0.3 | 0.03 | 0.03 | 7 × 10−5 |
| 60 | 0.54 | 0.0035 | 0.03 | 0.26 | 0.26 | 0.03 | 0.03 | 5 × 10−6 |
| 80 | 0.58 | 0.0032 | 0.028 | 0.27 | 0.27 | 0.04 | 0.04 | 4 × 10−6 |
| 100 | 0.57 | 0.0037 | 0.37 | 0.36 | 0.36 | 0.05 | 0.05 | 3 × 10−6 |
| Soil Depth (cm) | Solid (%) | b1 | b2 | b3 | Cn (J∙s−1∙°C−1) | C0 (J∙s−1∙°C−1) | Cw (J∙s−1∙°C−1) |
|---|---|---|---|---|---|---|---|
| 20 | 0.6168 | 7.26 × 1010 | 1.17 × 1011 | 4.58 × 1011 | 3.98 × 1010 | 5.2 × 1010 | 8.67 × 1010 |
| 40 | 0.6105 | 5.43 × 1010 | 8.4 × 1010 | 3.53 × 1011 | 3.44 × 1010 | 4.8 × 1010 | 7.57 × 1010 |
| 60 | 0.5968 | 4.32 × 1010 | 7.7 × 1010 | 3.47 × 1011 | 2.56 × 1010 | 4.21 × 1010 | 7.37 × 1010 |
| 80 | 0.6163 | 8.45 × 1010 | 1.21 × 1011 | 3.65 × 1011 | 2.47 × 1010 | 3.95 × 1010 | 6.27 × 1010 |
| 100 | 0.6157 | 1.13 × 1011 | 1.27 × 1011 | 2.54 × 1011 | 1.95 × 1010 | 3.64 × 1010 | 7.27 × 1010 |
| Treatment | Classification | MAE | RMSE | R2 |
|---|---|---|---|---|
| W1N1 | Water content | 0.0114 | 0.0115 | 0.7844 |
| Ammonium nitrogen | 0.0096 | 0.0028 | 0.7176 | |
| Nitrate nitrogen | 0.0424 | 0.0853 | 0.792 | |
| Temperature | 1.4699 | 1.6338 | 0.7344 | |
| W2N2 | Water content | 0.0099 | 0.0097 | 0.8045 |
| Ammonium nitrogen | 0.0079 | 0.0272 | 0.7332 | |
| Nitrate nitrogen | 0.0809 | 0.1628 | 0.8026 | |
| Temperature | 1.5161 | 1.6446 | 0.7185 | |
| W3N3 | Water content | 0.0093 | 0.0102 | 0.8179 |
| Ammonium nitrogen | 0.035 | 0.0911 | 0.7619 | |
| Nitrate nitrogen | 0.1714 | 0.2842 | 0.7928 | |
| Temperature | 1.2735 | 1.5606 | 0.7163 |
| Treatment | Seeding Stage | Nodulation Stage | Trumpet Stage | Silking Stage | Maturity Stage | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Urease | Catalase | Sucrase | Urease | Catalase | Sucrase | Urease | Catalase | Sucrase | Urease | Catalase | Sucrase | Urease | Catalase | Sucrase | |
| W1N1 | 0.84 bc | 0.88 b | 21.67 | 1.05 de | 1.26 ef | 24.56 bc | 0.86 d | 2.40 d | 23.29 d | 0.77 e | 2.14 e | 15.49 fg | 0.66 d | 1.74 f | 13.39 e |
| W1N2 | 0.85 bc | 0.94 ab | 21.31 | 1.18 c | 1.49 cd | 22.06 c | 0.92 d | 2.86 c | 24.04 d | 0.95 d | 3.02 bc | 19.53 e | 0.68 d | 2.06 d | 19.06 c |
| W1N3 | 0.81 cd | 0.95 ab | 21.87 | 0.92 f | 1.13 f | 14.41 d | 0.86 d | 0.98 f | 17.78 f | 0.67 e | 0.88 f | 17.49 ef | 0.52 e | 0.92 h | 10.05 f |
| W2N1 | 0.86 bc | 0.96 ab | 22.78 | 1.32 b | 1.39 de | 24.60 bc | 1.65 b | 3.02 c | 28.48 c | 1.41 b | 3.22 bc | 25.65 c | 0.93 bc | 2.92 b | 23.64 b |
| W2N2 | 0.84 bcd | 0.85 b | 22.89 | 1.43 a | 1.99 a | 32.06 a | 1.82 a | 4.02 a | 36.61 a | 1.72 a | 4.06 a | 35.17 a | 1.00 a | 3.27 a | 29.58 a |
| W2N3 | 0.95 a | 0.88 b | 22.49 | 1.40 ab | 1.79 b | 27.39 b | 1.81 a | 3.27 b | 30.98 bf | 1.68 a | 3.21 bc | 22.92 d | 0.96 ab | 3.05 b | 23.46 b |
| W3N1 | 0.85 bc | 0.91 ab | 21.51 | 0.93 f | 1.31 e | 17.17 d | 0.85 d | 2.56 d | 19.81 e | 0.98 cd | 2.55 d | 24.58 cd | 0.89 c | 1.87 ef | 16.83 d |
| W3N2 | 0.83 bcd | 1.06 a | 21.64 | 0.97 ef | 1.55 c | 26.34 b | 1.28 c | 3.25 b | 27.71 c | 0.92 d | 3.33 b | 33.54 a | 0.95 bc | 2.65 c | 30.59 a |
| W3N3 | 0.89 ab | 0.92 ab | 22.46 | 1.15 cd | 1.81 b | 25.41 b | 1.64 b | 3.13 b | 26.37 c | 1.08 c | 2.95 c | 27.78 b | 0.97 ab | 2.02 de | 31.08 a |
| CK | 0.76 d | 0.86 b | 21.24 | 0.93 f | 1.12 f | 21.85 c | 0.75 e | 2.09 e | 20.26 e | 0.67 e | 1.87 e | 13.54 g | 0.57 e | 1.50 g | 11.51 ef |
| W1 | 0.83 | 0.92 | 21.61 b | 1.05 b | 1.29 c | 20.34 c | 0.88 c | 2.08 c | 21.70 c | 0.80 c | 2.01 c | 17.50 c | 0.62 b | 1.57 c | 14.16 b |
| W2 | 0.88 | 0.89 | 22.72 a | 1.38 a | 1.72 a | 28.02 a | 1.76 a | 3.44 a | 30.25 a | 1.60 a | 3.49 a | 27.91 b | 0.97 a | 3.08 a | 25.56 a |
| W3 | 0.86 | 0.96 | 21.87 ab | 1.01 b | 1.56 b | 22.97 b | 1.26 b | 3.04 b | 24.63 b | 0.99 b | 2.94 b | 28.63 a | 0.94 a | 2.18 b | 26.17 a |
| N1 | 0.85 | 0.91 | 21.99 | 1.10 b | 1.32 c | 22.11 b | 1.12 c | 2.66 b | 23.86 b | 1.05 b | 2.64 b | 21.91 b | 0.83 ab | 2.17 b | 17.96 c |
| N2 | 0.84 | 0.95 | 21.94 | 1.19 a | 1.68 a | 26.82 a | 1.34 b | 3.38 a | 29.45 a | 1.19 a | 3.47 a | 29.41 a | 0.87 a | 2.66 a | 26.41 a |
| N3 | 0.88 | 0.91 | 22.27 | 1.16 ab | 1.57 b | 22.40 b | 1.44 a | 2.52 c | 25.04 b | 1.14 ab | 2.34 c | 22.73 b | 0.81 b | 2.00 c | 21.53 b |
| F | |||||||||||||||
| Lower limit of irrigation | 3.494 | 1.987 | 4.386 * | 91.434 ** | 70.431 ** | 53.419 ** | 529.87 ** | 274.601 ** | 204.572 ** | 452.612 ** | 182.397 ** | 238.379 ** | 346.866 ** | 512.757 ** | 256.071 ** |
| Nitrogen application | 2.331 | 0.565 | 0.425 | 5.014 * | 50.295 ** | 24.464 ** | 73.395 ** | 118.89 ** | 62.798 ** | 13.332 ** | 111.234 ** | 104.124 ** | 9.653 ** | 105.594 ** | 100.91 ** |
| Lower limit of irrigation × nitrogen application | 3.031 | 2.753 | 0.645 | 10.308 ** | 21.848 ** | 27.339 ** | 40.378 ** | 73.71 ** | 25.163 ** | 16.461 ** | 28.868 ** | 15.6 ** | 11.872 ** | 29.278 ** | 41.333 ** |
| Treatment | Fruit Length | Number of Grains | Thousand Grain Weight | Yield |
|---|---|---|---|---|
| W1N1 | 16.46 e | 552 bc | 359.66 fg | 7453.35 f |
| W1N2 | 16.80 e | 567 ab | 386.50 e | 8215.30 e |
| W1N3 | 17.33 d | 530 cde | 366.00 f | 7285.37 f |
| W2N1 | 18.43 c | 573 ab | 413.66 d | 8891.25 d |
| W2N2 | 20.13 a | 540 cd | 539.50 a | 10,928.52 a |
| W2N3 | 18.40 c | 589 a | 462.00 b | 10,211.46 b |
| W3N1 | 15.90 f | 526 de | 364.00 fg | 7188.12 f |
| W3N2 | 18.86 c | 516 e | 430.66 c | 8335.14 e |
| W3N3 | 19.43 b | 571 ab | 428.16 cd | 9172.30 c |
| CK | 14.93 g | 493 f | 348.66 g | 6450.45 g |
| W1 | 16.87 c | 550.11 b | 370.72 c | 7651.34 c |
| W2 | 18.99 a | 567.78 a | 471.72 a | 10,010.41 a |
| W3 | 18.07 b | 538.11 b | 407.61 b | 8231.86 b |
| N1 | 16.93 b | 550.89 ab | 379.11 c | 7844.24 c |
| N2 | 18.60 a | 541.22 b | 452.22 a | 9159.65 a |
| N3 | 18.39 a | 563.89 a | 418.72 b | 8889.71 b |
| F | ||||
| Lower limit of irrigation | 118.048 ** | 13.041 ** | 278.807 ** | 512.007 ** |
| Nitrogen application | 85.847 ** | 7.575 ** | 142.970 ** | 163.574 ** |
| Lower limit of irrigation × nitrogen application | 40.036 ** | 13.984 ** | 29.102 ** | 48.440 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Wang, S.; Yang, M. Modeling of Water, Heat, and Nitrogen Dynamics in Summer Maize under Broad Furrow Irrigation and the Mechanism of Enzyme Activity Response. Agronomy 2024, 14, 1044. https://doi.org/10.3390/agronomy14051044
Liu T, Wang S, Yang M. Modeling of Water, Heat, and Nitrogen Dynamics in Summer Maize under Broad Furrow Irrigation and the Mechanism of Enzyme Activity Response. Agronomy. 2024; 14(5):1044. https://doi.org/10.3390/agronomy14051044
Chicago/Turabian StyleLiu, Tengfei, Shunsheng Wang, and Mingwei Yang. 2024. "Modeling of Water, Heat, and Nitrogen Dynamics in Summer Maize under Broad Furrow Irrigation and the Mechanism of Enzyme Activity Response" Agronomy 14, no. 5: 1044. https://doi.org/10.3390/agronomy14051044
APA StyleLiu, T., Wang, S., & Yang, M. (2024). Modeling of Water, Heat, and Nitrogen Dynamics in Summer Maize under Broad Furrow Irrigation and the Mechanism of Enzyme Activity Response. Agronomy, 14(5), 1044. https://doi.org/10.3390/agronomy14051044