
Identifying Key Regulatory Genes in Drug Resistance Acquisition: Modeling Pseudotime Trajectories of Breast Cancer Single-Cell Transcriptome

Institute for Protein Research, Osaka University, Suita 565-0871, Osaka, Japan
*
Author to whom correspondence should be addressed.
Cancers 2024, 16(10), 1884; https://doi.org/10.3390/cancers16101884
Submission received: 28 April 2024
/
Revised: 11 May 2024
/
Accepted: 15 May 2024
/
Published: 15 May 2024
(This article belongs to the Special Issue Adaptive Resistance to Targeted Cancer Therapies and Rational Development of Combination Therapies)
Abstract
:Simple Summary
Despite recent advancements in experimental technology for genome-wide molecular profiling, our understanding of the dynamic mechanism underlying cancer drug resistance remains limited. In this study, we present an approach that combines mathematical modeling with the pseudotime analysis of single-cell time-series transcriptome data of drug-treated breast cancer cells. Our method identifies approximately 600 genes out of 6000 exhibiting multistable expression states, including RPS6KB1, a predictor of poor prognosis, cell survival, and growth in estrogen-receptor-positive breast cancers. The bifurcation analysis elucidates the regulatory mechanisms of the key regulatory genes, which can also be mapped into a molecular network based on cell survival and metastasis-related pathways, providing a comprehensive understanding of the interplay between signaling pathways and regulatory genes. Our method serves as a powerful tool for deciphering the complexities of drug resistance mechanisms in human diseases.
Abstract
Single-cell RNA-sequencing (scRNA-seq) technology has provided significant insights into cancer drug resistance at the single-cell level. However, understanding dynamic cell transitions at the molecular systems level remains limited, requiring a systems biology approach. We present an approach that combines mathematical modeling with a pseudotime analysis using time-series scRNA-seq data obtained from the breast cancer cell line MCF-7 treated with tamoxifen. Our single-cell analysis identified five distinct subpopulations, including tamoxifen-sensitive and -resistant groups. Using a single-gene mathematical model, we discovered approximately 560–680 genes out of 6000 exhibiting multistable expression states in each subpopulation, including key estrogen-receptor-positive breast cancer cell survival genes, such as RPS6KB1. A bifurcation analysis elucidated their regulatory mechanisms, and we mapped these genes into a molecular network associated with cell survival and metastasis-related pathways. Our modeling approach comprehensively identifies key regulatory genes for drug resistance acquisition, enhancing our understanding of potential drug targets in breast cancer.
1. Introduction
Breast cancer has been the most prevalent cancer among women and a leading cause of death since the late 20th century [1,2,3]. In the United States, approximately 75% of breast cancers are diagnosed as luminal A subtype [4], indicating their dependency on the estrogen receptor (ER) for growth. Tamoxifen, a widely used endocrine therapy for ER-positive breast cancer, has resulted in improvements in overall survival [5,6]. However, a significant proportion of patients treated with tamoxifen experience a relapse within 5–15 years, often resulting in metastasis and mortality [5,6]. Despite accumulating knowledge on the genetic and nongenetic molecular mechanisms of endocrine resistance, such as ESR1 mutation, transcriptomic and epigenetic changes, and the activation of signaling pathways [6,7,8], our understanding of the dynamic processes remains limited, requiring a comprehensive and systemic approach.
Recently, single-cell RNA-sequencing (scRNA-seq) analysis using MCF-7 cells, a luminal A subtype breast cancer model, has provided insights into drug resistance acquisition, revealing cellular heterogeneity and dynamic cell transitions. Notably, a “pre-adapted” subpopulation exhibiting the plasticity between drug tolerance and resistance has been identified in MCF-7 cells [9,10]. These studies investigated the activation of cell survival signals, such as the NF-κB pathway. Furthermore, Magi et al. reported altered metabolic regulation, cell adhesion, and histone modification based on the analysis of time-series scRNA-seq data under tamoxifen treatment [11]. While these studies have uncovered important genes and pathways associated with drug resistance acquisition, gaps still exist in our understanding of regulatory mechanisms at the systems level. Hence, a further exploration for potential drug targets is warranted.
To address these issues, we propose an approach that combines mathematical modeling with a pseudotime analysis using time-series transcriptome data, which automatically identifies key regulatory genes. We first identified potential drug-sensitive and -resistant cells by analyzing the time-series scRNA-seq data of tamoxifen-treated MCF-7 cells. Next, we performed a pseudotime analysis, from which we observed a transcriptional relay involved in cAMP, PI3K-AKT-mTOR, RHO-GTP, NF-κB, and NOTCH pathways. We identified genes exhibiting multistable expression states by modeling the individual gene expression patterns and conducting a steady-state analysis. Finally, we propose a transcriptional relay network model that includes cell survival, focal adhesion, and epithelial–mesenchymal transition (EMT), along with the key regulatory genes. Our results suggest that the modeling approach through a pseudotime analysis of the single-cell transcriptome serves as a powerful tool for understanding the drug-induced cellular transition.
2. Results
2.1. Overview of the Analysis Workflow
To theoretically address the challenge of identifying effective targets for regulating complex molecular systems during drug resistance acquisition (Figure 1a), we developed an approach combining mathematical modeling and a pseudotime analysis using time-series transcriptome data, which automatically identifies key regulatory genes (Figure 1b). Previous research on cell differentiation revealed that transcription factor networks exhibiting multistable states simulate dynamic cell transitions of progenitor cells into different terminal states [12,13,14]. This suggests that genes with multistable expression states could serve as potential indicators of cell differentiation, potentially regulated by switch-like responding pathways. To identify these key regulatory genes, we modeled each gene expression using an ordinary differential equation, determining whether the expression state in the long-time limit is mono- or multistable. Although multistability was investigated in the steady state, we considered these “multistable genes” key regulators, assuming the cell fate is determined within the long-time limit. Subsequently, as discussed later, we constructed a transcriptional relay network model for cell survival, focal adhesion, and EMT.
2.2. Dissecting the Heterogeneity and Defining Pseudotime for MCF-7 Cells
To understand transcriptional regulation dynamics underlying drug resistance acquisition, we analyzed a published dataset containing time-series scRNA-seq data of MCF-7 cells subjected to continuous tamoxifen treatment [11]. The data were collected at weeks 0 (before tamoxifen treatment), 3, 6, and 9, referred to as W0, W3, W6, and W9, respectively (Figure 2a). The original authors reported that cell growth was significantly inhibited by the fifth week, after which two resistant subpopulations emerged: one displaying altered metabolic regulation and the other exhibiting cell adhesion and histone modification. To provide a deeper insight into these findings, we first analyzed the W0 data to identify potential drug-sensitive and -resistant cells present before tamoxifen treatment. Subsequently, we analyzed the W0–W9 data, allowing us to derive the pseudotime gene expression profiles.
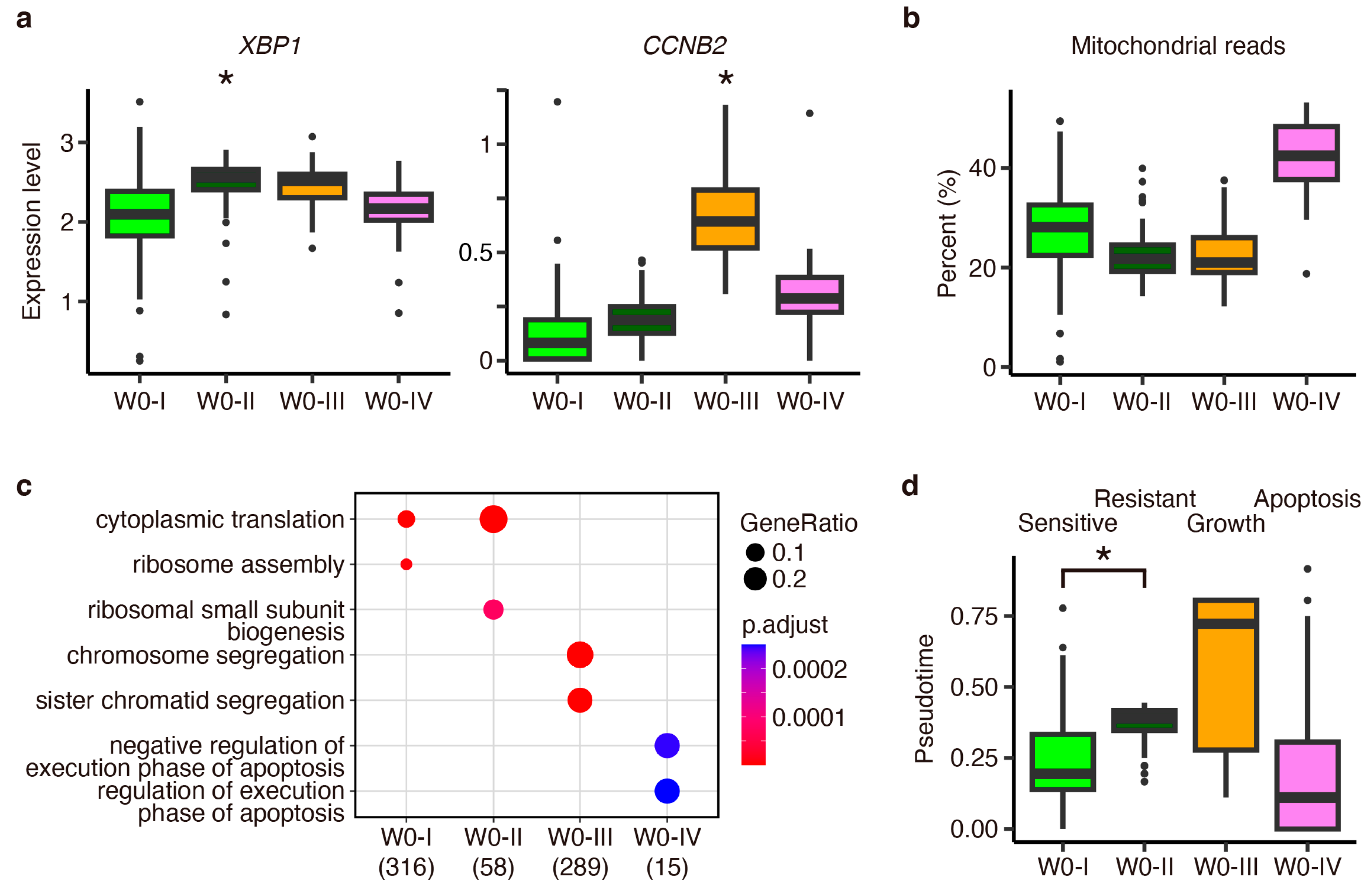
After normalizing the W0–W9 data (Appendix A and Appendix B), we initially focused on W0. Using Seurat (v4.4.0) [15], a standard software for scRNA-seq data analysis, we clustered the cells into four distinct subpopulations (W0-I, -II, -III, and -IV) and inferred the cell cycle phases (Figure 2b). We examined differentially expressed genes to assess potential drug-resistant states (Figure 2c). We observed the significant upregulation of IGFBP5 and JUN in the W0-I subpopulation. Previous studies have implicated IGFBP5 in cancer inhibition by temporally inactivating the insulin-like growth factor receptor [16], and JUN has been linked to increased tamoxifen sensitivity through protein kinase C activation [17,18]. Conversely, a positive cell cycle regulator CCND1 [19] and apoptosis inhibitor XBP1 [20] were significantly upregulated in W0-II, while other positive cell cycle regulators CCNA2 [21] and CCNB1/2 [22] were significantly upregulated in W0-III (Figure A1a). A notable increase in mitochondrial read fractions [23] and results from a Gene Ontology enrichment analysis [24] suggested that cells within W0-IV undergo apoptosis (Figure A1b,c).
Subsequently, we analyzed the W0–W9 data using Seurat and conducted a principal component analysis (PCA) based on 5000 highly variable genes, followed by a nonlinear dimensionality reduction using a diffusion map (Figure 2d, left). Thereafter, using MERLoT (v0.2.2) [25], a pseudotime analysis software, we smoothed the gene expression profiles and clustered the cells into five classes, C1–C5 (Figure 2d, right). We observed that cells in W0, W3–W6, and W9 predominantly belong to C1, C2–C3, and C4, respectively (Figure 2e, left). We noticed that the population ratio in the G2/M phase drastically decreased from C1 to C2 but gradually increased from C2 to C4 (Figure 2e, right). Since tamoxifen treatment induces G1 arrest, followed by gradual cell growth recovery [11], we assumed that the C1–C2–C3–C4 axis approximately corresponds to the progression of tamoxifen resistance acquisition, whereas C5 predominantly consists of cells in the S and G2/M phases.
Then, we defined pseudotime t for all cells, where we set t = 0 and t = 1 at the putative start and end points in C1 and C4, respectively (Figure 2f and Appendix C). Consistent with our interpretation of pseudotime as the progression of tamoxifen resistance acquisition, we annotated W0-I and W0-II as the potential tamoxifen-sensitive and resistant populations, respectively (Figure A1d). According to the pseudotime gene expression profiles (Figure 2g), we observed that ESR1 was downregulated for t > 0.5, suggesting that ESR1-independent mechanisms occur after tamoxifen treatment, as previously indicated [11]. We also noticed that key regulators in the cAMP (such as GREB1, XBP1, and CREB1), PI3K-AKT (RPS6KB1), RHOA-GTP (RHOA), NF-κB (NFKB1), and NOTCH (NOTCH2) pathways were sequentially upregulated, suggesting the existence of transcriptional relay.
Finally, we investigated the presence of a common transcriptional relay mechanism among different cell lines using external data. In a previous study, preadapted (PA) subpopulations were identified in both MCF-7 and T47D cells [9], a luminal A subtype breast cancer model distinguished from MCF-7 by TP53 mutation. PA cells were characterized by a high CD44 expression and transcriptomic signatures associated with cell survival and EMT in response to estradiol (E2) deprivation. Analyzing the scRNA-seq data of T47D- and T47D-derived long-term estrogen-deprived cells, we discovered a potential common transcriptional relay mechanism underlying the responses to tamoxifen treatment in MCF-7 cells and E2 depletion in T47D cells (Appendix D).
2.3. Modeling Pseudotime Gene Expression Patterns
As extensively reviewed, several computational tools have been developed to construct gene regulatory networks from scRNA-seq data [26]. One common approach is transcription factor networks based on the assumption that mRNA levels correlate well with protein levels [27,28]. However, questions arise from the intricate processes, including environmental changes (e.g., drug concentration [29]), the interconnection of signaling pathways, chemical modification, and the nuclear translocation of transcription factors, as well as co-operation with coactivators and corepressors [7]. These processes can significantly influence target gene expressions. To address such complexity, we modeled individual gene expressions using an autoregulatory gene circuit widely used for modeling various biochemical processes (Figure 3a) [30,31], which is exemplified by an ER-related signaling pathway (Figure 3b) [32,33,34,35]. This model operates under the assumptions that all genes are controlled by positive and negative feedback mechanisms, and the strength of this feedback control is determined by the mRNA levels and interactions among intracellular signals. The model is described by the following nonautonomous ordinary differential equation:
where I is the number of genes, is a normalized expression level () for gene , and are the magnitudes of positive and negative feedback regulation, and are the transcription and degradation rates, , , , and are the half-saturation constants, and , , , and are Hill coefficients. All the parameter values are non-negative real numbers.
We separately estimated the model parameters for 6082 genes, which passed a filter removing low-expressed genes—for the C1, C3, and C4 subpopulations—wherein we neglected C2 and C5 due to the lack of sufficient pseudotime lengths (Appendix E and Appendix F). Numerical data fitting in mechanistic models with tens to hundreds of unknown parameters remains challenging [36]. Nevertheless, we addressed this problem using a simplified model (Equation (1)). Here, we used a multi-starts method [37], one of the successful approaches to avoid a suboptimal estimation using different initial guesses, followed by a method using smooth and match estimators [38,39], a fast optimization method fitting a curve to a given batch of data (Figure 3c). Furthermore, we applied our original l2-norm penalty method—a parameter set minimizing is likely to be selected—by assuming that these parameters represent the energy costs of activation and inhibition for gene expressions. This restriction improved the robustness of the estimation.
We observed that fitting errors were generally small (<0.1) for most genes, indicating a good fit of our model to the pseudotime gene expression data (Figure 3d). However, suboptimal parameter sets, derived from different initial guesses during local optimization, also yielded small fitting errors but resulted in different predictions (Figure A2). This suggests the complexity of our model, which may involve an excessive number of parameters compared with the pseudotime data and a possibility of overfitting. Thus, we evaluated the robustness of parameter optimization against small differences in gene expression profiles, controlling the number of representative pseudotime points, denoted as (i.e., the number of “nodes” in Figure 2d, right), which impacts the pseudotime gene expression profiles [25].
To test whether gene-wide median parameter values were consistent across different values of , we independently conducted a nonparametric one-way ANOVA for C1, C3, and C4 (Figure 3e). Notably, all 12 parameters for C1 showed no significant differences between , whereas parameters , , and exhibited significant differences between (Figure 3e, left), which may be attributed to fluctuations in parameter values observed for certain genes (Figure 3e, right). Robustness was also observed for C3 and C4 (Figure A3 and Appendix G). Moreover, we verified that all 12 parameters were essential for our model, confirming that every principal component computed from all the estimated parameter values has an almost equal number of contributions (Figure A4). Consequently, we demonstrated the statistically accurate and robust parameter estimation, offering a reliable method for predicting cellular transition states.
2.4. Transient and Bifurcation Analysis for Key Regulatory Genes
We conducted a steady-state analysis to identify genes exhibiting multistable expression states at equilibrium. We counted the number of stable fixed points by deriving the steady-state equation from Equation (1) and computing its zero points with a negative differential coefficient. Our analysis revealed that 664, 558, and 680 genes exhibited multistability in the C1, C3, and C4 subpopulations, respectively (Figure 4a). Notably, RPS6KB1, a predictor of poor prognosis, cell survival, and growth in ER-positive breast cancers [35], exhibited bistability in both C1 and C4, implying that its low and high expression levels are regulated by a binary switch governing distinct cell fates.
To further investigate the transient gene expression dynamics dependent on the initial conditions, we computed a potential landscape analogous to Waddington’s landscape [40]. This landscape visually represents how gene expression states evolve over time, analogous to marbles rolling on hills and valleys. Derived from Equation (1), the potential function is given by the following gradient form of the landscape:
where 2F1 is the Gauss hypergeometric function [41]. Note that the direct differentiation of Equation (4) using the formula leads to Equation (2). Our observations indicated that the long-time behavior of RPS6KB1 expression is predominantly determined early in the pseudotime in C1, regardless of initial perturbations (Figure 4b, left). Furthermore, the potential landscape drove most initial states toward higher expression levels in both C1 and C4 (Figure 4b, right), suggesting challenges in controlling RPS6KB1 expression post cell fate determination.
To interpret the association of RPS6KB1 with tamoxifen sensitivity and resistance, we partitioned C1 based on normalized expression levels <1 or ≥1. Tamoxifen-sensitive cells in the W0-I subpopulation predominantly belonged to the RPS6KB1-low population, while tamoxifen-resistant and growing cells in W0-II and W0-III primarily belonged to the RPS6KB1-high population (Figure 4c). Thus, the RPS6KB1 expression serves as a potential indicator of tamoxifen-resistant states regulated by binary switching pathways.
In a bifurcation analysis, we investigated the relationship between equilibrium expression states and parameters and , which represent the inhibitory effects on positive and negative regulation, respectively. The bistable region for RPS6KB1 was quite asymmetric with respect to and : Changes in were more efficient than those in for driving the bistable state into a low-expression state (Figure 4d). This suggests that positive regulation inhibitors effectively control RPS6KB1 expression, offering potential targets for drug resistance interventions.
2.5. Targeting Key Regulatory Genes in Survival and Metastasis-Related Pathways
To comprehensively identify key regulatory genes in the C1 subpopulation, we computed bifurcation diagrams for the genes exhibiting mono- and multistable expression states involved in the cAMP, PI3K-AKT-mTOR, MAPK, NOTCH, and several other pathways, with and serving as parameters controlling the bifurcation (Figure 5a). Notably, we observed significant asymmetry in the bistable regions; for genes such as RPS6KB1, NRAS, ITGB1BP1, and ARID1A, exhibited a higher efficacy for inducing bifurcation, whereas, for TBL1XR1, exhibited a higher efficacy. Conversely, for JUND, controlling both and proved efficient for inducing bifurcation. On the other hand, for MYC, neither nor induced multistability.
Additionally, we integrated the genes exhibiting multistable expression states into a schematic representation of cell survival and metastasis-related pathways, focusing on those involved in focal adhesion and EMT (Figure 5b and Figure A6), which was manually constructed based on the results of the pseudotime analysis and previous reports (Appendix H). This comprehensive mapping provides a method for further understanding the complex interplay between signaling pathways and genes for therapeutic intervention. Furthermore, the observed peaks in RPS6KB1, NFKB1, RHOA, and NOTCH2 expressions (Figure 2g) suggest cell survival, focal adhesion, and EMT processes in C2 (~3 weeks), C3 (~3–6 weeks), and C4 (~3–9 weeks), respectively, adding further information to the dynamic processes of drug resistance acquisition.
3. Discussion
We proposed an approach that combines mathematical modeling with a pseudotime analysis using single-cell time-series transcriptome data to identify key regulatory genes involved in drug resistance acquisition in breast cancer. In contrast to previous systems biology models that rely on predefined kinetic networks [42,43,44,45], our approach offers a widely applicable and computationally efficient method for modeling each gene expression. We demonstrated statistically accurate and robust parameter estimation for approximately 6000 genes and identified 560–680 genes exhibiting multistable expression states, including RPS6KB1, a predictor of poor prognosis, cell survival, and growth in ER-positive breast cancers [35]. Furthermore, our analysis revealed that the long-time behavior of the RPS6KB1 expression is predominantly determined early in the pseudotime and can be effectively modulated by inhibitors for positive regulation. Nevertheless, experimental validation is important and potentially involves targeting the drug design, focusing on the key genes.
Despite yielding small fitting errors, suboptimal parameter sets produced divergent gene expression dynamics, indicative of the sensitivity of our model, which may involve an excessive number of parameters. Thus, future work should focus on model parameter reduction, possibly by sharing parameters among specific genes and mapping the original and reduced parameter spaces. A further model reduction will also mitigate the risk of overfitting. Furthermore, cell-to-cell heterogeneity should not be disregarded. While some genes, such as NFKB1 and NOTCH2, exhibited similar pseudotime gene expression patterns (Figure 2g), the Pearson correlation coefficient across cells was <0.05. This low correlation coefficient indicates a considerable variability in the gene expression among cells. Oral cancer cells exhibit heterogeneous responses to transforming growth factor β stimuli via multiple EMT pathways [46]. This emphasizes the critical role of heterogeneous signaling states in affecting the drug response, which should be incorporated into future modeling approaches.
Finally, several limitations are worth noting. We identified potential drug-sensitive and -resistance subpopulations in the tamoxifen-untreated cells and found that genes exhibiting bistability were concentrated in the MAPK-related pathways associated with cell survival (Figure 5b), suggesting that cell survival is regulated by switch-like responding pathways. Additionally, a “pre-adapted” MCF-7 subpopulation exhibiting drug tolerance has been identified [9,10]. Although these findings are useful, our analysis is currently limited to only the published scRNA-seq data [11]. This does not ensure its generalizability to other cancer datasets. Therefore, a further analysis using multiple datasets such as time-series transcriptome [9,10] and proteome data [47] is necessary to improve our data-driven model and thoroughly understand cancer drug resistance acquisition.
4. Conclusions
Our modeling approach, combined with a single-cell pseudotime analysis, revealed genes with multistable expression states and elucidated their regulatory mechanisms through a bifurcation analysis. Mapping the key regulatory genes into a molecular network considering cell survival and metastasis-related pathways provided a comprehensive understanding of the interplay between the signaling pathways and genes, serving as a powerful tool for deciphering the complexities of drug resistance mechanisms in breast cancer. Future analyses should aim to uncover additional insights into the mechanisms of cancer drug resistance by leveraging larger datasets.
Author Contributions
Conceptualization, K.I.; methodology, K.I.; data analysis, K.I.; writing—original draft preparation, K.I.; writing—review and editing, K.I. and M.O.; project administration, M.O.; funding acquisition, K.I. and M.O. All authors have read and agreed to the published version of the manuscript.
Funding
K.I. was supported by JST Moonshot R&D [JPMJMS2021]. M.O. was supported by JST CREST [JPMJCR21N3], JSPS KAKENHI [18H04031], and the Uehara Memorial Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The scRNA-seq data of MCF-7 cells treated with tamoxifen are available in the Gene Expression Omnibus with accession code DRA009126. The scRNA-seq data of T47D- and T47D-derived long-term estrogen-deprived cells were obtained from the Gene Expression Omnibus with accession code GSE122743. The codes for analyzing data in this article are available on Github (https://github.com/keita-iida/PSEUDOTIMEABC, accessed on 27 April 2024).
Acknowledgments
We thank Takeya Kasukawa and Masato Tsutsui for their assistance in identifying mathematical errors and improving the study design. We also thank Masahiro Inoue and Sho Soeda for improving the figures.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A. Removing Low-Quality Genes and Cells from Single-Cell RNA-Sequencing Data
Low-quality genes and cells were removed from all single-cell RNA-sequencing (scRNA-seq) datasets through two steps: (i) removing genes for which the number of non-zero expressing cells is less than 10; (ii) removing cells whose read counts , number of genes expressed with non-zero read counts , and percent reads mapped to mitochondrial genes is within 0–55. The remaining read count table contained 14,476 genes and 1000 cells.
Appendix B. Identifying Potential Drug-Sensitive and -Resistant Cells in Week 0 Data
The W0–W9 data were normalized using the R package RECODE (v1.0.0) [48]. Then, the W0 data were selected and normalized by Seurat NormalizeData() (v4.4.0) [49]. Thereafter, highly variable genes were selected using FindVariableFeatures() based on a variance-stabilizing transformation with 800 genes. Dimensionality reduction was conducted using ScaleData() and RunPCA() with features = Seurat::VariableFeatures(). The principal components that explained 50% of the total variability were used for FindNeighbors() computations. Cells were clustered into four distinct groups (W0-I, -II, -III, and -IV) by FindClusters() at a resolution of 0.5. Cell cycle phases were inferred by CellCycleScoring() with s.features = Seurat::cc.genes$s.genes and g2m.features = Seurat::cc.genes$g2m.genes. Differentially expressed genes were detected by FindAllMarkers() based on adjusted p-values < 0.001 (Figure 2c and Figure A1a) and were subjected to a subsequent Gene Ontology enrichment analysis. We performed bitr() inputting org.Hs.eg.db (v3.15.0) using clusterProfiler (v4.4.4) [24], followed by compareCluster() with fun = “enrichGO”, ont = “BP”, pAdjustMethod = “BH”, and pvalueCutoff = 0.001 (Figure A1c).
Appendix C. Defining Pseudotime in Weeks 0–9 Data
The W0–W9 data were normalized using the R packages bayNorm (v1.14.0) [50] with Condition = [sample labels, W0, W3, W6, and W9], Prior_type = “LL”, and mode_version = TRUE, followed by Seurat NormalizeData(). Afterwards, highly variable genes were selected using FindVariableFeatures() based on a variance-stabilizing transformation with 5000 genes. Dimensionality reduction was conducted using ScaleData() and RunPCA() with features = Seurat::VariableFeatures(), followed by a further dimensionality reduction using DiffusionMap() in the destiny package (v3.13.0) [51]. Cell cycle phases were inferred by CellCycleScoring() with s.features = Seurat::cc.genes$s.genes and g2m.features = Seurat::cc.genes$g2m.genes. Based on the first four components of the diffusion map co-ordinates, cells were clustered into five distinct groups (C1, C2, C3, C4, and C5) using MERLoT (v0.2.2) [25] as follows: CalculateScaffoldTree() with NEndpoints = 4 and random_seed = 1, CalculateElasticTree() with N_yk = 60, and GenesSpaceEmbedding() with NCores = 3, which was subjected to a subsequent pseudotime analysis. Pseudotime (t) was computed using CalculatePseudotimes() and scaled to fall within the range of 0 to 1, setting t = 0 and t = 1 at the putative start and end points of branches C1 and C4, respectively. The pseudotime distribution was significantly lower in W0-I compared with that in W0-II, which were annotated as the tamoxifen-sensitive and -resistant subpopulations, respectively (Figure A1d).
Figure A1.
Differentially expressed gene (DEG) analysis and annotation of MCF-7 cells. (a) Box plots for W0 data showing the normalized DEG expression levels. Adjusted p-values for the clusters marked with asterisks indicate statistical significance (* p < 0.001, Mann–Whitney U test by Seurat). (b) Percentage of reads mapped to mitochondrial genes across subpopulations W0-I, -II, -III, and -IV. (c) Gene Ontology enrichment analysis based on the DEGs, highlighting the top two enriched terms. (d) Box plots for W0, showing pseudotimes across cells. An asterisk indicates statistical significance (* p < , Mann–Whitney U test by R stats). Subpopulations W0-I, -II, -III, and -IV are manually annotated as tamoxifen-sensitive, tamoxifen-resistant, cell growth, and apoptosis, respectively, based on the DEG analysis and the interpretation of pseudotime distribution, which reflects the progression of tamoxifen resistance acquisition (see also the main text).
Figure A1.
Differentially expressed gene (DEG) analysis and annotation of MCF-7 cells. (a) Box plots for W0 data showing the normalized DEG expression levels. Adjusted p-values for the clusters marked with asterisks indicate statistical significance (* p < 0.001, Mann–Whitney U test by Seurat). (b) Percentage of reads mapped to mitochondrial genes across subpopulations W0-I, -II, -III, and -IV. (c) Gene Ontology enrichment analysis based on the DEGs, highlighting the top two enriched terms. (d) Box plots for W0, showing pseudotimes across cells. An asterisk indicates statistical significance (* p < , Mann–Whitney U test by R stats). Subpopulations W0-I, -II, -III, and -IV are manually annotated as tamoxifen-sensitive, tamoxifen-resistant, cell growth, and apoptosis, respectively, based on the DEG analysis and the interpretation of pseudotime distribution, which reflects the progression of tamoxifen resistance acquisition (see also the main text).
Appendix D. Investigation of Estradiol-Depletion-Responding Genes Using External Data
The previous study identified preadapted (PA) subpopulations in both MCF-7 and T47D cells [9], a luminal A subtype breast cancer model distinguished from MCF-7 by TP53 mutation. These subpopulations were characterized under experimental conditions of short-term (2 days) and long-term (1 year) estrogen deprivation, which mimics clinical treatment with aromatase inhibitors. PA cells were characterized by a high CD44 expression and transcriptomic signatures associated with cell survival and EMT in response to estradiol (E2) deprivation. Here, we explored the presence of a common transcriptional relay mechanism in tamoxifen-treated MCF-7 and estrogen-deprived T47D cells.
The scRNA-seq data of T47D- and T47D-derived long-term estrogen-deprived (LTED) cells, either in the presence or absence of E2, were obtained from the Gene Expression Omnibus with accession code GSE122743 as published by Hong et al. [9]. The dataset includes T47D (control sample), T47D_RM_CD44H (CD44-positive T47D cells with E2), T47D_WM2d_CD44H (CD44-positive T47D cells without E2 collected on the second day), and T47D_LTED (unsorted LTED cells), labeled as TD1, TD2, TD3, and TD4, respectively (Figure A2a).
Figure A2.
Statistical analysis of single-cell RNA-seq data of T47D- and T47D-derived long-term estrogen-deprived (LTED) cells, either in the presence or absence of estradiol (E2). (a) Overview of the dataset. (b) Uniform manifold approximation and projection (UMAP) plots showing the sample labels. (c) Gene expression profiles of key regulators shown in Figure 2g. Adjusted p-values marked with asterisks indicate statistical significance (** p < , Mann–Whitney U test by Seurat).
Figure A2.
Statistical analysis of single-cell RNA-seq data of T47D- and T47D-derived long-term estrogen-deprived (LTED) cells, either in the presence or absence of estradiol (E2). (a) Overview of the dataset. (b) Uniform manifold approximation and projection (UMAP) plots showing the sample labels. (c) Gene expression profiles of key regulators shown in Figure 2g. Adjusted p-values marked with asterisks indicate statistical significance (** p < , Mann–Whitney U test by Seurat).

Normalization of the TD1–TD4 data was performed using the R packages RECODE (v1.0.0) [48] and Seurat NormalizeData() (v4.4.0) [49] under default settings. Subsequently, highly variable genes were selected using FindVariableFeatures() based on a variance-stabilizing transformation with 3161 genes. Dimensionality reduction was then conducted using ScaleData() and RunPCA() with features = Seurat::VariableFeatures(). A visualization of the results in a uniform manifold approximation and projection (UMAP) space confirmed the almost complete separation of T47D and LTED cells (Figure A2b), consistent with previous reports [9]. Finally, the expression profiles against tamoxifen treatment and E2 deprivation were compared (Figure A2c and Figure 2g). Notably, similar patterns in the gene expression changes were observed, such as decreased expressions of GREB1 and XBP1 along the TD2–TD4 axis and increased expressions of NFKB1 and NOTCH2 along the TD2–TD3 axis. These findings suggest a potential common transcriptional relay mechanism underlying the responses to tamoxifen treatment in MCF-7 cells and E2 depletion in T47D cells.
Appendix E. Processing Pseudotime Gene Expression Data
We excluded low-expressed genes with maximal expression levels along all pseudotime trajectories that were less than 0.1. Subsequently, 8448 genes out of 14,476 were excluded, leaving 6028 genes. The pseudotime ranges for C1, C2, C3, C4, and C5 were approximately [0, 0.5], [0.5, 0.61], [0.61, 0.94], [0.61, 1], and [0.5, 0.81], respectively. In this study, we neglected C2 and C5 due to insufficient pseudotime length. The pseudotime data computed by MERLoT contained small negative expression values, with a minimum value of approximately −0.024 across all genes. Since the pseudotime gene expression data will be used in our mathematical model analysis, we set the negative expression values to by adding the required values.
Appendix F. Parameter Estimation of the Mathematical Model
Let us denote the pseudotime gene expression profile of the ith gene by and the parameter set by:
where is the number of genes. We separately estimated for the C1, C3, and C4 subpopulations. For the parameter estimation, we set multiple initial guesses as:
where , and . To constrain the parameter values to be positive, we rewrote
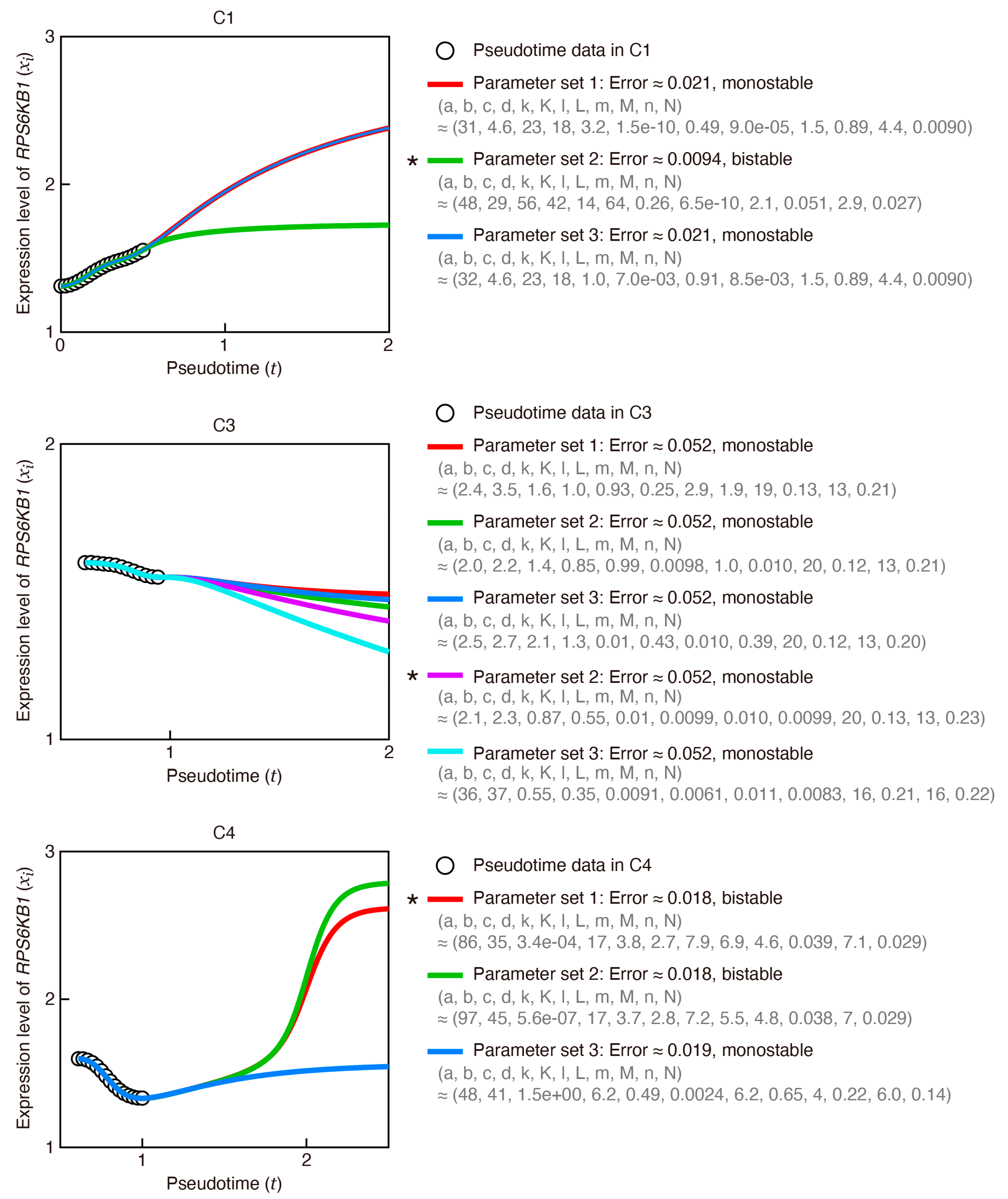
and fitted the curve to the pseudotime data using Gnuplot software (v6.0) for estimating , and , which implements the Levenberg–Marquardt algorithm, a nonlinear least-squares method. Our parameter estimation method provided multiple candidates of the optimal parameter set for each gene, along with the fitting error and l2-norm . We then selected the optimal parameter sets as follows: when the absolute difference between two parameter candidates is less than 0.01, we selected the parameter set minimizing as the optimal one; otherwise, we selected the parameter set with the minimal fitting error. Although this restriction improved the robustness of the estimation, suboptimal parameter sets, derived from different initial guesses during local optimization, also yielded small fitting errors but resulted in different predictions (Figure A3).
Figure A3.
Model-based prediction of pseudotime gene expressions with the optimal and suboptimal parameter sets, exemplified by the results for RPS6KB1 in C1, C3, and C4 subpopulations. Trajectories computed using the optimal parameter set are marked with asterisks. Information regarding the estimated parameter values, fitting errors, and stability (indicating whether the expression levels at equilibrium are mono- or bistable) is provided on the right.
Figure A3.
Model-based prediction of pseudotime gene expressions with the optimal and suboptimal parameter sets, exemplified by the results for RPS6KB1 in C1, C3, and C4 subpopulations. Trajectories computed using the optimal parameter set are marked with asterisks. Information regarding the estimated parameter values, fitting errors, and stability (indicating whether the expression levels at equilibrium are mono- or bistable) is provided on the right.
Appendix G. Evaluation of Robustness in Parameter Estimation
To evaluate the robustness of parameter optimization against small differences in gene expression profiles, we controlled the number of representative pseudotime points, denoted as (i.e., the number of “nodes” in Figure 2d, right), which impacts the pseudotime gene expression profiles [25]. We independently tested whether gene-wide median parameter values were consistent across different values of for C1, C3, and C4 using a nonparametric one-way ANOVA (Figure 3e and Figure A4). Additionally, to verify the necessity of all 12 parameters in our model, we performed PCA on the scaled gene-by-parameter matrix. This matrix comprises all estimated parameter values for C1, C3, and C4, resulting in a size of 17,394 () by 12. Our analysis confirmed that each principal component contributes almost equally (Figure A5).
Figure A4.
Evaluation of robustness in parameter estimation. (a) Gene-wide median parameter values for C3 (left), along with individual parameter values and the number of stable fixed points for RPS6KB1 (top right) and MAPK14 (bottom right) across different numbers of representative pseudotime points (). In this study, was used (indicated by vertical broken lines). According to the Kruskal–Wallis test, all 12 parameters showed no significant differences between (p-value > ), whereas , , , , , , and exhibited significant differences between : * p-values < . (b) Similar to (a), all 12 parameters showed no significant differences between (p-value > ), whereas , , , , and exhibited significant differences between : * p-values < .
Figure A4.
Evaluation of robustness in parameter estimation. (a) Gene-wide median parameter values for C3 (left), along with individual parameter values and the number of stable fixed points for RPS6KB1 (top right) and MAPK14 (bottom right) across different numbers of representative pseudotime points (). In this study, was used (indicated by vertical broken lines). According to the Kruskal–Wallis test, all 12 parameters showed no significant differences between (p-value > ), whereas , , , , , , and exhibited significant differences between : * p-values < . (b) Similar to (a), all 12 parameters showed no significant differences between (p-value > ), whereas , , , , and exhibited significant differences between : * p-values < .
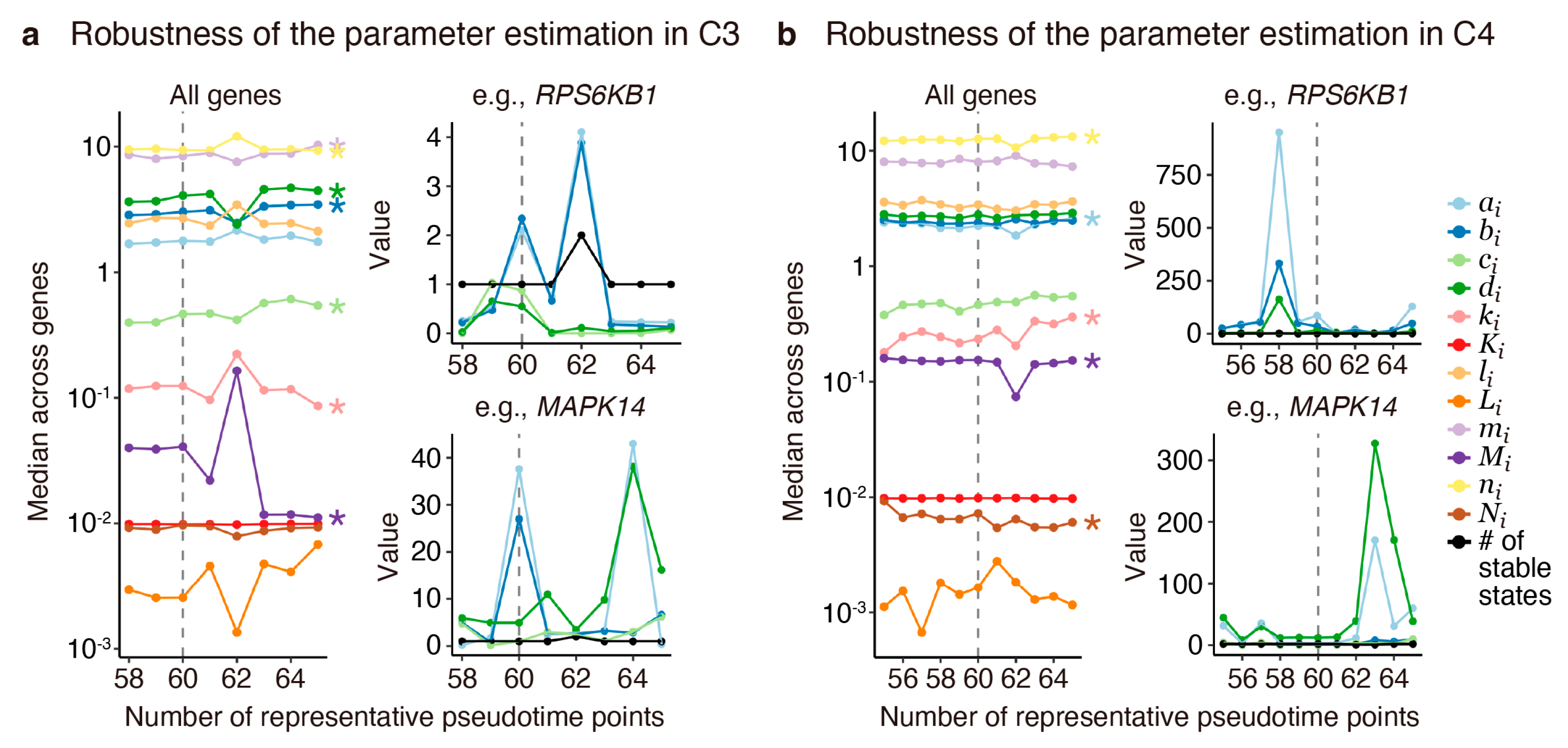
Figure A5.
Principal component analysis (PCA) of the estimated parameter values for 5798 genes. (a) PCA plots for C1, C3, and C4 subpopulations, with 6, 4, and 13 outlier genes, respectively, removed. (b) Cumulative variance across the number of principal components, demonstrating approximately equal contributions from each component.
Figure A5.
Principal component analysis (PCA) of the estimated parameter values for 5798 genes. (a) PCA plots for C1, C3, and C4 subpopulations, with 6, 4, and 13 outlier genes, respectively, removed. (b) Cumulative variance across the number of principal components, demonstrating approximately equal contributions from each component.

Appendix H. Construction of a Transcriptional Relay Network Model
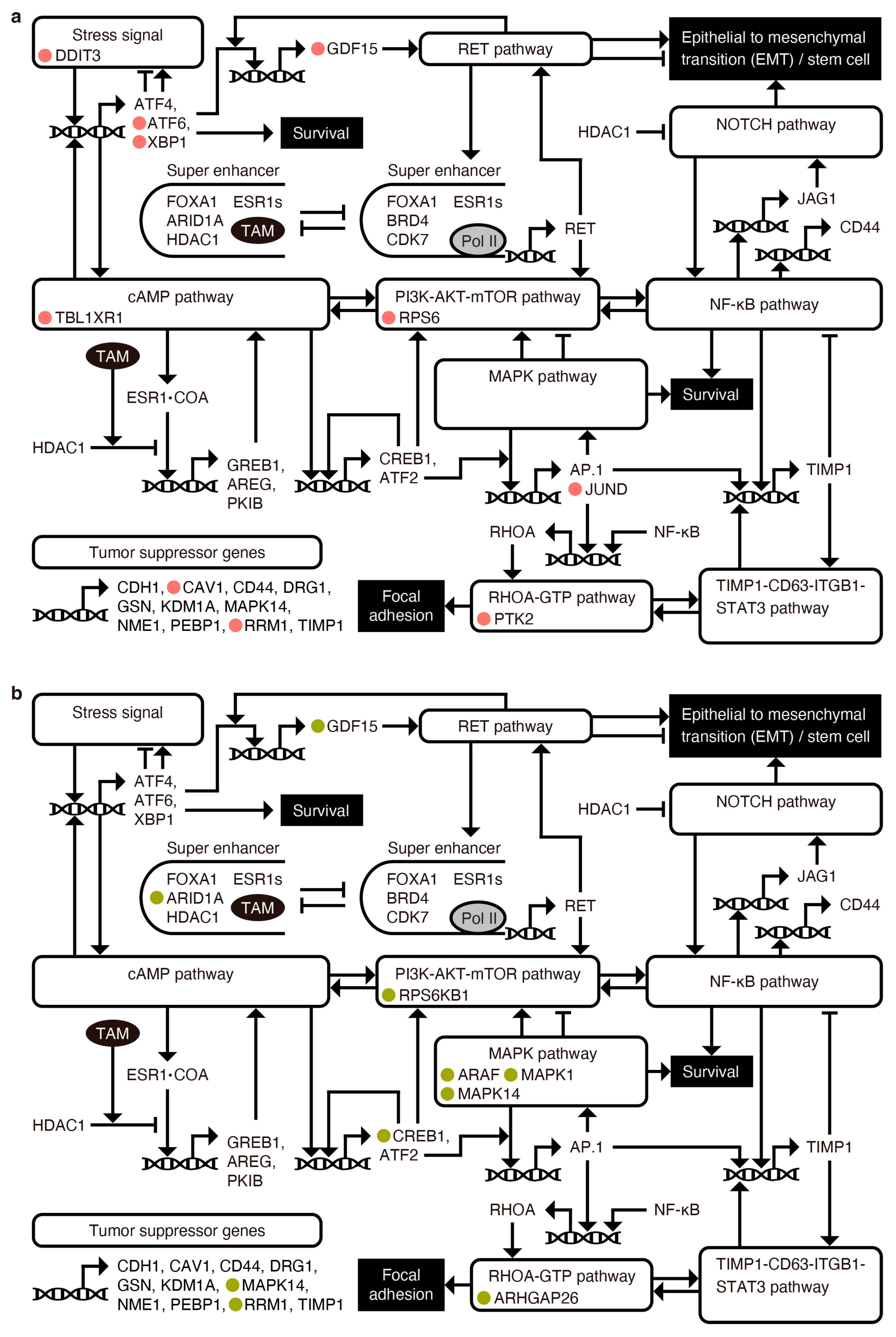
We manually constructed a transcription relay network model based on the result of the pseudotime analysis, genes exhibiting multistable expression states, and bibliographic survey for cell survival, focal adhesion, and epithelial–mesenchymal transition (Table A1).
Figure A6.
Comprehensive mapping of the key regulatory genes exhibiting multistability into cell survival and metastasis-related pathways. These genes were identified in (a) C3 and (b) C4 populations, marked by red- and yellow-filled circles, respectively. Arrow and blunt arrow represent activation and inhibition, respectively. TAM, tamoxifen.
Figure A6.
Comprehensive mapping of the key regulatory genes exhibiting multistability into cell survival and metastasis-related pathways. These genes were identified in (a) C3 and (b) C4 populations, marked by red- and yellow-filled circles, respectively. Arrow and blunt arrow represent activation and inhibition, respectively. TAM, tamoxifen.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
List of genes associated with cell survival, focal adhesion, and epithelial–mesenchymal transition-related pathways.
Table A1.
List of genes associated with cell survival, focal adhesion, and epithelial–mesenchymal transition-related pathways.
| Pathway Name | Related Gene | Reference |
|---|---|---|
| cAMP | HDAC1, HDAC2, NCOR1, TBL1X, TBL1XR1, CREBBP, TRAM1, GREB1, AREG, PKIB, CREB1, ATF2 | [6,7,52,53,54,55,56,57,58,59] |
| Cellular stress | ATF2, ATF4, ATF6, XBP1, DDIT3, EIF2A | [60,61,62] |
| PI3K-AKT-mTOR | PIK3R1, PTEN, PIK3C2A, AKT1, AKTIP, MTOR, RPS6KB1, RPS6 | [56,63,64] |
| MAPK | NRAS, MAP3K2, MAP3K7, TAB2, MAP3K13, MAP3K20, RAF1, ARAF, MAP2K2, MAPK3, MAPK1, MAPK14, MAPK9, MAPKAP1 | [55,56,57] |
| Survival | ESR1, MYC | |
| AP.1 | JUN, JUNB, JUND, FOS, FOSB | [57,65,66] |
| NF-κB | RELA, NFKB1, NFKBIA | [66,67,68,69,70,71,72,73] |
| TIMP-1-CD63-ITGB1-STAT3 | TIMP1, CD63, ITGB1, ITGB1BP1, STAT3 | [65,67,68,73,74,75] |
| RHO-GTP | RHOA, PTK2, ARHGAP26, ROCK1, ROCK2, TRIOBP | [74,75] |
| RET | GDF15, RET, YY1, FOXA1, ARID1A, BRD4, CDK7 | [63,76,77,78,79,80,81,82,83] |
| NOTCH | JAG1, NOTCH2, RBPJ | [70,71,72,84,85] |
References
- Silverberg, E. Cancer statistics, 1984. CA Cancer J. Clin. 1984, 34, 7–23. [Google Scholar] [PubMed]
- Jemal, A.; Tiwari, R.C.; Murray, T.; Ghafoor, A.; Samuels, A.; Ward, E.; Feuer, E.J.; Thun, M.J.; American Cancer, S. Cancer statistics, 2004. CA Cancer J. Clin. 2004, 54, 8–29. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Society, A.C. Breast Cancer Facts & Figures 2019–2020; American Cancer Society, Inc.: Atlanta, GA, USA, 2019. [Google Scholar]
- Pan, H.; Gray, R.; Braybrooke, J.; Davies, C.; Taylor, C.; McGale, P.; Peto, R.; Pritchard, K.I.; Bergh, J.; Dowsett, M.; et al. 20-Year Risks of Breast-Cancer Recurrence after Stopping Endocrine Therapy at 5 Years. N. Engl. J. Med. 2017, 377, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Saatci, O.; Huynh-Dam, K.T.; Sahin, O. Endocrine resistance in breast cancer: From molecular mechanisms to therapeutic strategies. J. Mol. Med. 2021, 99, 1691–1710. [Google Scholar] [CrossRef] [PubMed]
- Ring, A.; Dowsett, M. Mechanisms of tamoxifen resistance. Endocr. Relat. Cancer 2004, 11, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Hanker, A.B.; Sudhan, D.R.; Arteaga, C.L. Overcoming Endocrine Resistance in Breast Cancer. Cancer Cell 2020, 37, 496–513. [Google Scholar] [CrossRef]
- Hong, S.P.; Chan, T.E.; Lombardo, Y.; Corleone, G.; Rotmensz, N.; Bravaccini, S.; Rocca, A.; Pruneri, G.; McEwen, K.R.; Coombes, R.C.; et al. Single-cell transcriptomics reveals multi-step adaptations to endocrine therapy. Nat. Commun. 2019, 10, 3840. [Google Scholar] [CrossRef]
- Semina, S.E.; Pal, P.; Kansara, N.S.; Huggins, R.J.; Alarid, E.T.; Greene, G.L.; Frasor, J. Selective pressure of endocrine therapy activates the integrated stress response through NF-κB signaling in a subpopulation of ER positive breast cancer cells. Breast Cancer Res. 2022, 24, 19. [Google Scholar] [CrossRef]
- Magi, S.; Ki, S.; Ukai, M.; Dominguez-Huttinger, E.; Naito, A.T.; Suzuki, Y.; Okada, M. A combination approach of pseudotime analysis and mathematical modeling for understanding drug-resistant mechanisms. Sci. Rep. 2021, 11, 18511. [Google Scholar] [CrossRef]
- Huang, S.; Guo, Y.P.; May, G.; Enver, T. Bifurcation dynamics in lineage-commitment in bipotent progenitor cells. Dev. Biol. 2007, 305, 695–713. [Google Scholar] [CrossRef] [PubMed]
- Ferrell, J.E., Jr. Bistability, bifurcations, and Waddington’s epigenetic landscape. Curr. Biol. 2012, 22, R458–R466. [Google Scholar] [CrossRef]
- Marco, E.; Karp, R.L.; Guo, G.; Robson, P.; Hart, A.H.; Trippa, L.; Yuan, G.C. Bifurcation analysis of single-cell gene expression data reveals epigenetic landscape. Proc. Natl. Acad. Sci. USA 2014, 111, E5643–E5650. [Google Scholar] [CrossRef] [PubMed]
- Satija, R.; Farrell, J.A.; Gennert, D.; Schier, A.F.; Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 2015, 33, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, J. Biological effects and regulation of IGFBP5 in breast cancer. Front. Endocrinol. 2022, 13, 983793. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhi, H.; Lepp, A.; Wang, P.; Huang, J.; Basir, Z.; Chitambar, C.R.; Myers, C.R.; Chen, G. p38γ mitogen-activated protein kinase (MAPK) confers breast cancer hormone sensitivity by switching estrogen receptor (ER) signaling from classical to nonclassical pathway via stimulating ER phosphorylation and c-Jun transcription. J. Biol. Chem. 2012, 287, 14681–14691. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zou, S.T.; Zhu, R.; Li, W.; Gu, C.W.; Wei, S.H.; Xie, J.M.; Wu, H.R. Inhibition of proliferation of estrogen receptor-positive MCF-7 human breast cancer cells by tamoxifen through c-Jun transcription factors. Mol. Med. Rep. 2013, 7, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Marks, B.A.; Pipia, I.M.; Mukai, C.; Horibata, S.; Rice, E.J.; Danko, C.G.; Coonrod, S.A. GDNF-RET signaling and EGR1 form a positive feedback loop that promotes tamoxifen resistance via cyclin D1. BMC Cancer 2023, 23, 138. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Chen, Z.; Li, L.; Liu, H.; Zhang, R.; Cheng, Q.; Xu, D.; Wu, L. Unravel the molecular mechanism of XBP1 in regulating the biology of cancer cells. J. Cancer 2019, 10, 2035–2046. [Google Scholar] [CrossRef]
- Gao, T.; Han, Y.; Yu, L.; Ao, S.; Li, Z.; Ji, J. CCNA2 is a prognostic biomarker for ER+ breast cancer and tamoxifen resistance. PLoS ONE 2014, 9, e91771. [Google Scholar] [CrossRef]
- Meng, C.; Zou, Y.; Hong, W.; Bao, C.; Jia, X. Estrogen-regulated PTTG1 promotes breast cancer progression by regulating cyclin kinase expression. Mol. Med. 2020, 26, 33. [Google Scholar] [CrossRef]
- Luecken, M.D.; Theis, F.J. Current best practices in single-cell RNA-seq analysis: A tutorial. Mol. Syst. Biol. 2019, 15, e8746. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Parra, R.G.; Papadopoulos, N.; Ahumada-Arranz, L.; Kholtei, J.E.; Mottelson, N.; Horokhovsky, Y.; Treutlein, B.; Soeding, J. Reconstructing complex lineage trees from scRNA-seq data using MERLoT. Nucleic Acids Res. 2019, 47, 8961–8974. [Google Scholar] [CrossRef]
- Fiers, M.; Minnoye, L.; Aibar, S.; Bravo Gonzalez-Blas, C.; Kalender Atak, Z.; Aerts, S. Mapping gene regulatory networks from single-cell omics data. Brief. Funct. Genom. 2018, 17, 246–254. [Google Scholar] [CrossRef]
- Ocone, A.; Haghverdi, L.; Mueller, N.S.; Theis, F.J. Reconstructing gene regulatory dynamics from high-dimensional single-cell snapshot data. Bioinformatics 2015, 31, i89–i96. [Google Scholar] [CrossRef]
- Matsumoto, H.; Kiryu, H.; Furusawa, C.; Ko, M.S.H.; Ko, S.B.H.; Gouda, N.; Hayashi, T.; Nikaido, I. SCODE: An efficient regulatory network inference algorithm from single-cell RNA-Seq during differentiation. Bioinformatics 2017, 33, 2314–2321. [Google Scholar] [CrossRef]
- Bachmann, N.; Barberi-Heyob, M.; Bour, C.; Parache, R.M.; Guillemin, F.; Batt, A.M.; Merlin, J.L. Intracellular distribution of tamoxifen in resistant human breast adenocarcinoma cells using tamoxifen-eosin association. Cell Biol. Toxicol. 1998, 14, 429–435. [Google Scholar] [CrossRef]
- Becskei, A.; Serrano, L. Engineering stability in gene networks by autoregulation. Nature 2000, 405, 590–593. [Google Scholar] [CrossRef]
- Jia, C.; Wang, L.Y.; Yin, G.G.; Zhang, M.Q. Single-cell stochastic gene expression kinetics with coupled positive-plus-negative feedback. Phys. Rev. E 2019, 100, 052406. [Google Scholar] [CrossRef]
- Giuliano, M.; Schifp, R.; Osborne, C.K.; Trivedi, M.V. Biological mechanisms and clinical implications of endocrine resistance in breast cancer. Breast 2011, 20 (Suppl. 3), S42–S49. [Google Scholar] [CrossRef]
- Citro, S.; Miccolo, C.; Meloni, L.; Chiocca, S. PI3K/mTOR mediate mitogen-dependent HDAC1 phosphorylation in breast cancer: A novel regulation of estrogen receptor expression. J. Mol. Cell Biol. 2015, 7, 132–142. [Google Scholar] [CrossRef]
- Wu, X.; Xie, W.; Xie, W.; Wei, W.; Guo, J. Beyond controlling cell size: Functional analyses of S6K in tumorigenesis. Cell Death Dis. 2022, 13, 646. [Google Scholar] [CrossRef]
- Mayrovitz, H.N. (Ed.) Breast Cancer; Exon Publications: Brisbane, Australia, 2022. [Google Scholar]
- Villaverde, A.F.; Frohlich, F.; Weindl, D.; Hasenauer, J.; Banga, J.R. Benchmarking optimization methods for parameter estimation in large kinetic models. Bioinformatics 2019, 35, 830–838. [Google Scholar] [CrossRef]
- Frohlich, F.; Kaltenbacher, B.; Theis, F.J.; Hasenauer, J. Scalable Parameter Estimation for Genome-Scale Biochemical Reaction Networks. PLoS Comput. Biol. 2017, 13, e1005331. [Google Scholar] [CrossRef]
- Richard Bellman, R.S.R. The use of splines with unknown end points in the identification of systems. J. Math. Anal. Appl. 1971, 34, 26–33. [Google Scholar] [CrossRef]
- Gugushvili, S.; Klaassen, C.A.J. √n-consistent parameter estimation for systems of ordinary differential equations: Bypassing numerical integration via smoothing. Bernoulli 2012, 18, 1061–1098. [Google Scholar] [CrossRef]
- Waddington, C.H. The Strategy of the Genes; Routledge Library Editions: 20th Century Science; Routledge, Taylor and Francis Group: London, UK; New York, NY, USA, 2014. [Google Scholar]
- Abramowitz, M.; Stegun, I.A. Handbook of Mathematical Functions with Formulas, Graphs, and Mathematical Tables; U.S. Government Printing Office: Washington, DC, USA, 1964; p. xiv. 1046p.
- Nakakuki, T.; Birtwistle, M.R.; Saeki, Y.; Yumoto, N.; Ide, K.; Nagashima, T.; Brusch, L.; Ogunnaike, B.A.; Okada-Hatakeyama, M.; Kholodenko, B.N. Ligand-specific c-Fos expression emerges from the spatiotemporal control of ErbB network dynamics. Cell 2010, 141, 884–896. [Google Scholar] [CrossRef]
- Tian, T.; Song, J. Mathematical modelling of the MAP kinase pathway using proteomic datasets. PLoS ONE 2012, 7, e42230. [Google Scholar] [CrossRef]
- Imoto, H.; Zhang, S.; Okada, M. A Computational Framework for Prediction and Analysis of Cancer Signaling Dynamics from RNA Sequencing Data-Application to the ErbB Receptor Signaling Pathway. Cancers 2020, 12, 2878. [Google Scholar] [CrossRef]
- Imoto, H.; Yamashiro, S.; Okada, M. A text-based computational framework for patient -specific modeling for classification of cancers. iScience 2022, 25, 103944. [Google Scholar] [CrossRef]
- Takahashi, K.; Podyma-Inoue, K.A.; Saito, M.; Sakakitani, S.; Sugauchi, A.; Iida, K.; Iwabuchi, S.; Koinuma, D.; Kurioka, K.; Konishi, T.; et al. TGF-beta generates a population of cancer cells residing in G1 phase with high motility and metastatic potential via KRTAP2-3. Cell Rep. 2022, 40, 111411. [Google Scholar] [CrossRef]
- Madden, S.F.; Cremona, M.; Farrelly, A.M.; Low, W.H.; McBryan, J. Proteomic time course of breast cancer cells highlights enhanced sensitivity to Stat3 and Src inhibitors prior to endocrine resistance development. Cancer Gene Ther. 2023, 30, 324–334. [Google Scholar] [CrossRef]
- Imoto, Y.; Nakamura, T.; Escolar, E.G.; Yoshiwaki, M.; Kojima, Y.; Yabuta, Y.; Katou, Y.; Yamamoto, T.; Hiraoka, Y.; Saitou, M. Resolution of the curse of dimensionality in single-cell RNA sequencing data analysis. Life Sci. Alliance 2022, 5, e202201591. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M., 3rd; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated analysis of multimodal single-cell data. Cell 2021, 184, 3573–3587.e29. [Google Scholar] [CrossRef]
- Tang, W.; Bertaux, F.; Thomas, P.; Stefanelli, C.; Saint, M.; Marguerat, S.; Shahrezaei, V. bayNorm: Bayesian gene expression recovery, imputation and normalization for single-cell RNA-sequencing data. Bioinformatics 2020, 36, 1174–1181. [Google Scholar] [CrossRef]
- Angerer, P.; Haghverdi, L.; Buttner, M.; Theis, F.J.; Marr, C.; Buettner, F. destiny: Diffusion maps for large-scale single-cell data in R. Bioinformatics 2016, 32, 1241–1243. [Google Scholar] [CrossRef]
- Bischoff, P.; Kornhuber, M.; Dunst, S.; Zell, J.; Fauler, B.; Mielke, T.; Taubenberger, A.V.; Guck, J.; Oelgeschlager, M.; Schonfelder, G. Estrogens Determine Adherens Junction Organization and E-Cadherin Clustering in Breast Cancer Cells via Amphiregulin. iScience 2020, 23, 101683. [Google Scholar] [CrossRef]
- Mohammed, H.; D’Santos, C.; Serandour, A.A.; Ali, H.R.; Brown, G.D.; Atkins, A.; Rueda, O.M.; Holmes, K.A.; Theodorou, V.; Robinson, J.L.; et al. Endogenous purification reveals GREB1 as a key estrogen receptor regulatory factor. Cell Rep. 2013, 3, 342–349. [Google Scholar] [CrossRef]
- Ahmed, M.B.; Alghamdi, A.A.A.; Islam, S.U.; Lee, J.S.; Lee, Y.S. cAMP Signaling in Cancer: A PKA-CREB and EPAC-Centric Approach. Cells 2022, 11, 2020. [Google Scholar] [CrossRef]
- Gazon, H.; Barbeau, B.; Mesnard, J.M.; Peloponese, J.M., Jr. Hijacking of the AP-1 Signaling Pathway during Development of ATL. Front. Microbiol. 2017, 8, 2686. [Google Scholar] [CrossRef]
- Ye, N.; Ding, Y.; Wild, C.; Shen, Q.; Zhou, J. Small molecule inhibitors targeting activator protein 1 (AP-1). J. Med. Chem. 2014, 57, 6930–6948. [Google Scholar] [CrossRef]
- Eferl, R.; Wagner, E.F. AP-1: A double-edged sword in tumorigenesis. Nat. Rev. Cancer 2003, 3, 859–868. [Google Scholar] [CrossRef]
- Wang, Y.W.; Chen, X.; Ma, R.; Gao, P. Understanding the CREB1-miRNA feedback loop in human malignancies. Tumour Biol. 2016, 37, 8487–8502. [Google Scholar] [CrossRef]
- Zhang, H.; Kong, Q.; Wang, J.; Jiang, Y.; Hua, H. Complex roles of cAMP-PKA-CREB signaling in cancer. Exp. Hematol. Oncol. 2020, 9, 32. [Google Scholar] [CrossRef]
- B’Chir, W.; Maurin, A.C.; Carraro, V.; Averous, J.; Jousse, C.; Muranishi, Y.; Parry, L.; Stepien, G.; Fafournoux, P.; Bruhat, A. The eIF2alpha/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res. 2013, 41, 7683–7699. [Google Scholar] [CrossRef]
- Wang, D.; Day, E.A.; Townsend, L.K.; Djordjevic, D.; Jorgensen, S.B.; Steinberg, G.R. GDF15: Emerging biology and therapeutic applications for obesity and cardiometabolic disease. Nat. Rev. Endocrinol. 2021, 17, 592–607. [Google Scholar] [CrossRef]
- Barua, D.; Gupta, A.; Gupta, S. Targeting the IRE1-XBP1 axis to overcome endocrine resistance in breast cancer: Opportunities and challenges. Cancer Lett. 2020, 486, 29–37. [Google Scholar] [CrossRef]
- Morandi, A.; Plaza-Menacho, I.; Isacke, C.M. RET in breast cancer: Functional and therapeutic implications. Trends Mol. Med. 2011, 17, 149–157. [Google Scholar] [CrossRef]
- Rozengurt, E.; Soares, H.P.; Sinnet-Smith, J. Suppression of feedback loops mediated by PI3K/mTOR induces multiple overactivation of compensatory pathways: An unintended consequence leading to drug resistance. Mol. Cancer Ther. 2014, 13, 2477–2488. [Google Scholar] [CrossRef]
- Kwak, H.J.; Park, M.J.; Cho, H.; Park, C.M.; Moon, S.I.; Lee, H.C.; Park, I.C.; Kim, M.S.; Rhee, C.H.; Hong, S.I. Transforming growth factor-beta1 induces tissue inhibitor of metalloproteinase-1 expression via activation of extracellular signal-regulated kinase and Sp1 in human fibrosarcoma cells. Mol. Cancer Res. 2006, 4, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Islam, R.; Cho, J.Y.; Jeong, H.; Cap, K.C.; Park, Y.; Hossain, A.J.; Park, J.B. Regulation of RhoA GTPase and various transcription factors in the RhoA pathway. J. Cell. Physiol. 2018, 233, 6381–6392. [Google Scholar] [CrossRef] [PubMed]
- Ries, C. Cytokine functions of TIMP-1. Cell. Mol. Life Sci. 2014, 71, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Friedmann-Morvinski, D.; Narasimamurthy, R.; Xia, Y.; Myskiw, C.; Soda, Y.; Verma, I.M. Targeting NF-κB in glioblastoma: A therapeutic approach. Sci. Adv. 2016, 2, e1501292. [Google Scholar] [CrossRef] [PubMed]
- Tannous, B.A.; Badr, C.E. A TNF-NF-κB-STAT3 loop triggers resistance of glioma-stem-like cells to Smac mimetics while sensitizing to EZH2 inhibitors. Cell Death Dis. 2019, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Emuss, V.; Lagos, D.; Pizzey, A.; Gratrix, F.; Henderson, S.R.; Boshoff, C. KSHV manipulates Notch signaling by DLL4 and JAG1 to alter cell cycle genes in lymphatic endothelia. PLoS Pathog. 2009, 5, e1000616. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Taguchi, Y.; Ito-Kureha, T.; Semba, K.; Yamaguchi, N.; Inoue, J. NF-κB non-cell-autonomously regulates cancer stem cell populations in the basal-like breast cancer subtype. Nat. Commun. 2013, 4, 2299. [Google Scholar] [CrossRef] [PubMed]
- Hossain, F.; Sorrentino, C.; Ucar, D.A.; Peng, Y.; Matossian, M.; Wyczechowska, D.; Crabtree, J.; Zabaleta, J.; Morello, S.; Del Valle, L.; et al. Notch Signaling Regulates Mitochondrial Metabolism and NF-κB Activity in Triple-Negative Breast Cancer Cells via IKKalpha-Dependent Non-canonical Pathways. Front. Oncol. 2018, 8, 575. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, A.; Dittmer, J. A CAF-Fueled TIMP-1/CD63/ITGB1/STAT3 Feedback Loop Promotes Migration and Growth of Breast Cancer Cells. Cancers 2022, 14, 4983. [Google Scholar] [CrossRef]
- Ando, T.; Charindra, D.; Shrestha, M.; Umehara, H.; Ogawa, I.; Miyauchi, M.; Takata, T. Tissue inhibitor of metalloproteinase-1 promotes cell proliferation through YAP/TAZ activation in cancer. Oncogene 2018, 37, 263–270. [Google Scholar] [CrossRef]
- Justo, B.L.; Jasiulionis, M.G. Characteristics of TIMP1, CD63, and beta1-Integrin and the Functional Impact of Their Interaction in Cancer. Int. J. Mol. Sci. 2021, 22, 9319. [Google Scholar] [CrossRef]
- Wang, S.F.; Chang, Y.L.; Fang, W.L.; Li, A.F.; Chen, C.F.; Yeh, T.S.; Hung, G.Y.; Huang, K.H.; Lee, H.C. Growth differentiation factor 15 induces cisplatin resistance through upregulation of xCT expression and glutathione synthesis in gastric cancer. Cancer Sci. 2023, 114, 3301–3317. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, D.J. GFRA1: A Novel Molecular Target for the Prevention of Osteosarcoma Chemoresistance. Int. J. Mol. Sci. 2018, 19M, 1078. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.; Rao, S.V.; Sutton, J.; Cheeseman, D.; Dunn, S.; Papachristou, E.K.; Prada, J.G.; Couturier, D.L.; Kumar, S.; Kishore, K.; et al. ARID1A influences HDAC1/BRD4 activity, intrinsic proliferative capacity and breast cancer treatment response. Nat. Genet. 2020, 52, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Seachrist, D.D.; Anstine, L.J.; Keri, R.A. FOXA1: A Pioneer of Nuclear Receptor Action in Breast Cancer. Cancers 2021, 13, 5205. [Google Scholar] [CrossRef]
- Zheng, Z.Z.; Xia, L.; Hu, G.S.; Liu, J.Y.; Hu, Y.H.; Chen, Y.J.; Peng, J.Y.; Zhang, W.J.; Liu, W. Super-enhancer-controlled positive feedback loop BRD4/ERalpha-RET-ERalpha promotes ERalpha-positive breast cancer. Nucleic Acids Res. 2022, 50, 10230–10248. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.H.; Qu, Q.; Teng, X.Q.; Dai, Y.H.; Qu, J. Superenhancers as master gene regulators and novel therapeutic targets in brain tumors. Exp. Mol. Med. 2023, 55, 290–303. [Google Scholar] [CrossRef]
- Johansson, J.; Tabor, V.; Wikell, A.; Jalkanen, S.; Fuxe, J. TGF-beta1-Induced Epithelial-Mesenchymal Transition Promotes Monocyte/Macrophage Properties in Breast Cancer Cells. Front. Oncol. 2015, 5, 3. [Google Scholar] [CrossRef]
- Siaw, J.T.; Gabre, J.L.; Uckun, E.; Vigny, M.; Zhang, W.; Van den Eynden, J.; Hallberg, B.; Palmer, R.H.; Guan, J. Loss of RET Promotes Mesenchymal Identity in Neuroblastoma Cells. Cancers 2021, 13, 1909. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch signaling pathway: Architecture, disease, and therapeutics. Signal Transduct. Target. Ther. 2022, 7, 95. [Google Scholar] [CrossRef]
- Bai, J.W.; Wei, M.; Li, J.W.; Zhang, G.J. Notch Signaling Pathway and Endocrine Resistance in Breast Cancer. Front. Pharmacol. 2020, 11, 924. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Research question and overview of our modeling approach. (a) Research question: How to identify the effective targets regulating complex molecular systems upon drug resistance acquisition. (b) Modeling approach: (i) Smoothing of single-cell data; (ii) pseudotime analysis, which outputs pseudotime gene expression data; (iii) mathematical modeling of each gene expression using an ordinary differential equation; (iv) parameter estimation by fitting the model (left) and steady-state analysis (right); (v) screening of key regulatory genes exhibiting multistable expression levels at equilibrium; and (vi) mapping of key regulatory genes into a molecular network associated with drug resistance acquisition and identifying potential regulatory points.
Figure 1.
Research question and overview of our modeling approach. (a) Research question: How to identify the effective targets regulating complex molecular systems upon drug resistance acquisition. (b) Modeling approach: (i) Smoothing of single-cell data; (ii) pseudotime analysis, which outputs pseudotime gene expression data; (iii) mathematical modeling of each gene expression using an ordinary differential equation; (iv) parameter estimation by fitting the model (left) and steady-state analysis (right); (v) screening of key regulatory genes exhibiting multistable expression levels at equilibrium; and (vi) mapping of key regulatory genes into a molecular network associated with drug resistance acquisition and identifying potential regulatory points.
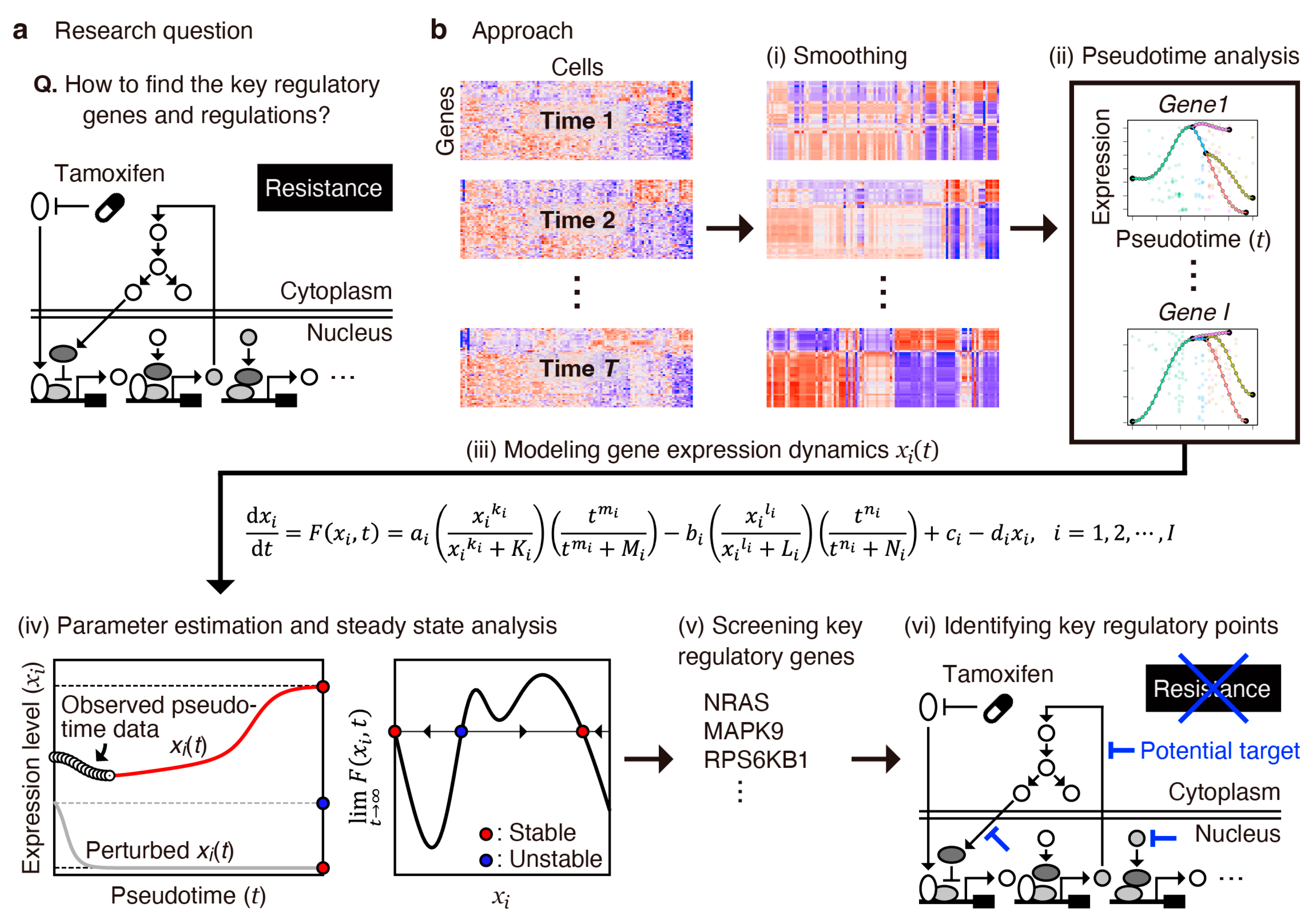
Figure 2.
Statistical and pseudotime analysis of time-series single-cell RNA-seq data of MCF-7 cells under continuous tamoxifen treatment. (a) Overview of the dataset. (b) Uniform manifold approximation and projection (UMAP) plots for the control sample (W0), showing the clustering results (left) and cell cycle phases (right) computed using Seurat. (c) Box plots for W0, showing the normalized expression levels of differentially expressed genes. Adjusted p-values for the clusters marked with asterisks indicate statistical significance (* p < 0.001, Mann–Whitney U test by Seurat). (d) Diffusion map plots for all samples (W0–W9), showing the sample labels (left) and clustering labels (right) computed using MERLoT. (e) Population ratios across clustering versus sample labels (left) and clustering versus cell cycle phases (right). (f) Diffusion map plot showing pseudotime t, where we set t = 0 and t = 1 at the putative start and end points of branches C1 and C4, respectively. (g) Pseudotime gene expression profiles of key regulators in estrogen receptor pathway (such as ESR1), cAMP pathway (GREB1, XBP1, and CREB1), PI3K-AKT (RPS6KB1), RHOA-GTP (RHOA), NF-κB (NFKB1), and NOTCH (NOTCH2) pathways, which are sequentially upregulated from top left to bottom right, suggesting the existence of transcriptional relay.
Figure 2.
Statistical and pseudotime analysis of time-series single-cell RNA-seq data of MCF-7 cells under continuous tamoxifen treatment. (a) Overview of the dataset. (b) Uniform manifold approximation and projection (UMAP) plots for the control sample (W0), showing the clustering results (left) and cell cycle phases (right) computed using Seurat. (c) Box plots for W0, showing the normalized expression levels of differentially expressed genes. Adjusted p-values for the clusters marked with asterisks indicate statistical significance (* p < 0.001, Mann–Whitney U test by Seurat). (d) Diffusion map plots for all samples (W0–W9), showing the sample labels (left) and clustering labels (right) computed using MERLoT. (e) Population ratios across clustering versus sample labels (left) and clustering versus cell cycle phases (right). (f) Diffusion map plot showing pseudotime t, where we set t = 0 and t = 1 at the putative start and end points of branches C1 and C4, respectively. (g) Pseudotime gene expression profiles of key regulators in estrogen receptor pathway (such as ESR1), cAMP pathway (GREB1, XBP1, and CREB1), PI3K-AKT (RPS6KB1), RHOA-GTP (RHOA), NF-κB (NFKB1), and NOTCH (NOTCH2) pathways, which are sequentially upregulated from top left to bottom right, suggesting the existence of transcriptional relay.

Figure 3.
Mathematical modeling, parameter estimation, and evaluation of accuracy and robustness. (a) Schematic representation of the mathematical model in which intracellular tamoxifen levels affect gene regulation via extrinsic pathways. The arrows and blunt arrows represent positive and negative regulation, respectively. Particularly, the and parameters represent inhibitory effects on positive and negative regulation, respectively. (b) Schematic representation of RPS6KB1 regulation, an example of estrogen-receptor-related signaling. (c) Demonstration of parameter estimation using NOTCH2 pseudotime trajectories. Parameters were independently optimized for subpopulations C1, C3, and C4, whereas C2 and C5 were neglected due to insufficient pseudotime length. (d) Histograms of the fitting errors showing that the errors are generally small (≤0.1) for most genes (5798 genes out of 6082), as exemplified by the result for MTCO1P12 in C1 at the right bottom. (e) Gene-wide median parameter values for C1 (left), along with individual parameter values and the number of stable fixed points for RPS6KB1 (top right) and MAPK14 (bottom right) across different numbers of representative pseudotime points (). In this study, was used (indicated by vertical broken lines). According to the Kruskal–Wallis test, all 12 parameters showed no significant differences between (p-value > ), whereas , , and exhibited significant differences between : * p-values < .
Figure 3.
Mathematical modeling, parameter estimation, and evaluation of accuracy and robustness. (a) Schematic representation of the mathematical model in which intracellular tamoxifen levels affect gene regulation via extrinsic pathways. The arrows and blunt arrows represent positive and negative regulation, respectively. Particularly, the and parameters represent inhibitory effects on positive and negative regulation, respectively. (b) Schematic representation of RPS6KB1 regulation, an example of estrogen-receptor-related signaling. (c) Demonstration of parameter estimation using NOTCH2 pseudotime trajectories. Parameters were independently optimized for subpopulations C1, C3, and C4, whereas C2 and C5 were neglected due to insufficient pseudotime length. (d) Histograms of the fitting errors showing that the errors are generally small (≤0.1) for most genes (5798 genes out of 6082), as exemplified by the result for MTCO1P12 in C1 at the right bottom. (e) Gene-wide median parameter values for C1 (left), along with individual parameter values and the number of stable fixed points for RPS6KB1 (top right) and MAPK14 (bottom right) across different numbers of representative pseudotime points (). In this study, was used (indicated by vertical broken lines). According to the Kruskal–Wallis test, all 12 parameters showed no significant differences between (p-value > ), whereas , , and exhibited significant differences between : * p-values < .
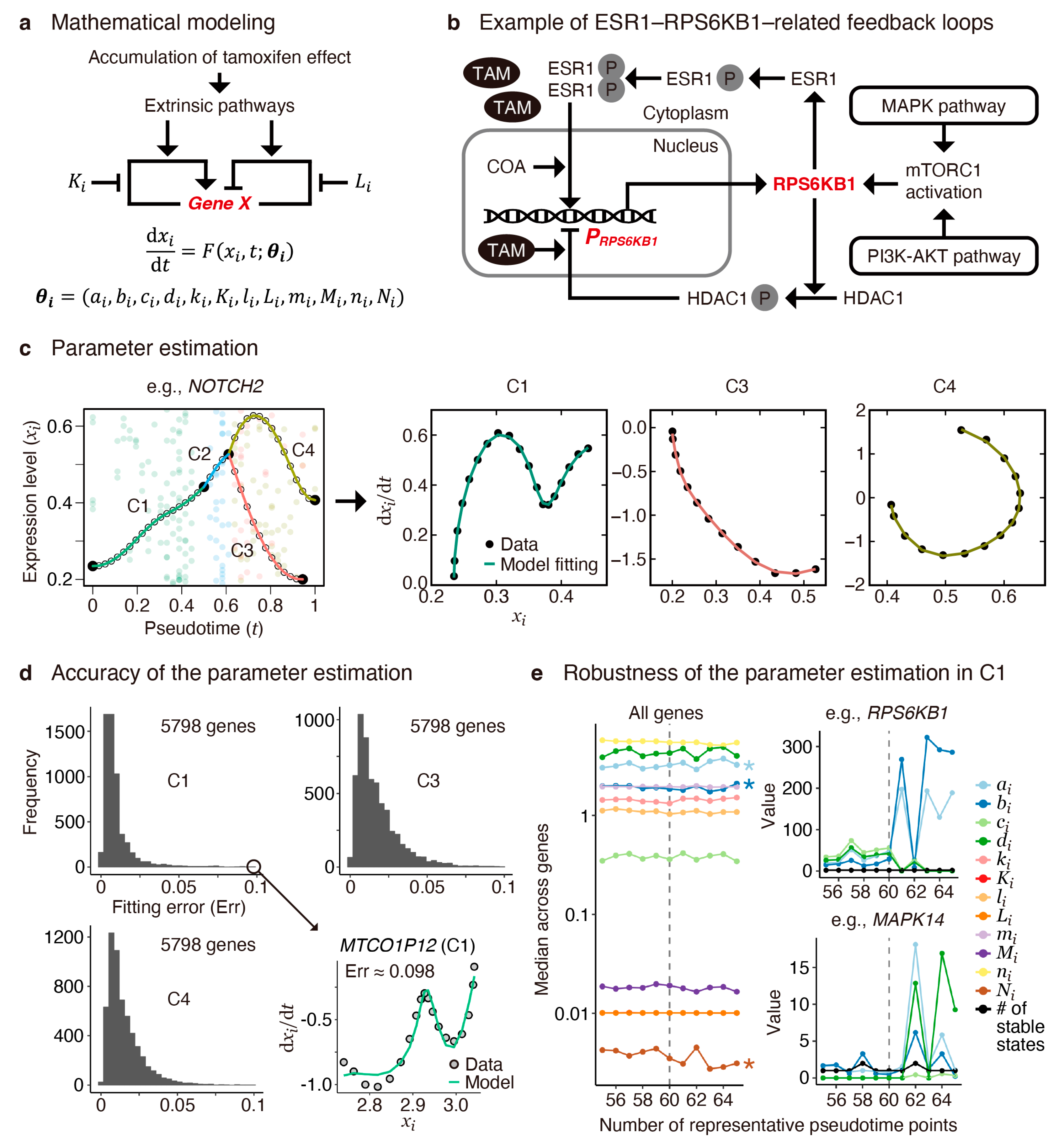
Figure 4.
Transient and bifurcation analysis for “multistable genes” exhibiting multistable expression levels at equilibrium. (a) The number of multistable genes identified in C1, C3, and C4 subpopulations and those whose pseudotime gene expression profiles could not be well fitted by the model. (b) Potential RPS6KB1 expression landscape in C1 (left) and C4 (right), providing a visual representation of how gene expression states evolve over pseudotime. Colored, filled circles are snapshots of inferred expression dynamics given several initial values: red circles correspond to the pseudotime data, while yellow, orange, magenta, and violet circles correspond to initial perturbations with 0.6-, 0.8-, 1.2-, and 1.4-fold changes, respectively. (c) Population ratios across W0-I (tamoxifen-sensitive cells), W0-II (tamoxifen-resistant cells), W0-III (growing cells), and W0-IV (apoptotic cells) versus RPS6KB1-low and -high populations. (d) Bifurcation diagrams showing stable (red) and unstable (blue) expression states dependent on parameters and , which represent inhibitory effects on positive and negative regulation, respectively. The vertical black lines represent the parameter set estimated from the pseudotime data.
Figure 4.
Transient and bifurcation analysis for “multistable genes” exhibiting multistable expression levels at equilibrium. (a) The number of multistable genes identified in C1, C3, and C4 subpopulations and those whose pseudotime gene expression profiles could not be well fitted by the model. (b) Potential RPS6KB1 expression landscape in C1 (left) and C4 (right), providing a visual representation of how gene expression states evolve over pseudotime. Colored, filled circles are snapshots of inferred expression dynamics given several initial values: red circles correspond to the pseudotime data, while yellow, orange, magenta, and violet circles correspond to initial perturbations with 0.6-, 0.8-, 1.2-, and 1.4-fold changes, respectively. (c) Population ratios across W0-I (tamoxifen-sensitive cells), W0-II (tamoxifen-resistant cells), W0-III (growing cells), and W0-IV (apoptotic cells) versus RPS6KB1-low and -high populations. (d) Bifurcation diagrams showing stable (red) and unstable (blue) expression states dependent on parameters and , which represent inhibitory effects on positive and negative regulation, respectively. The vertical black lines represent the parameter set estimated from the pseudotime data.
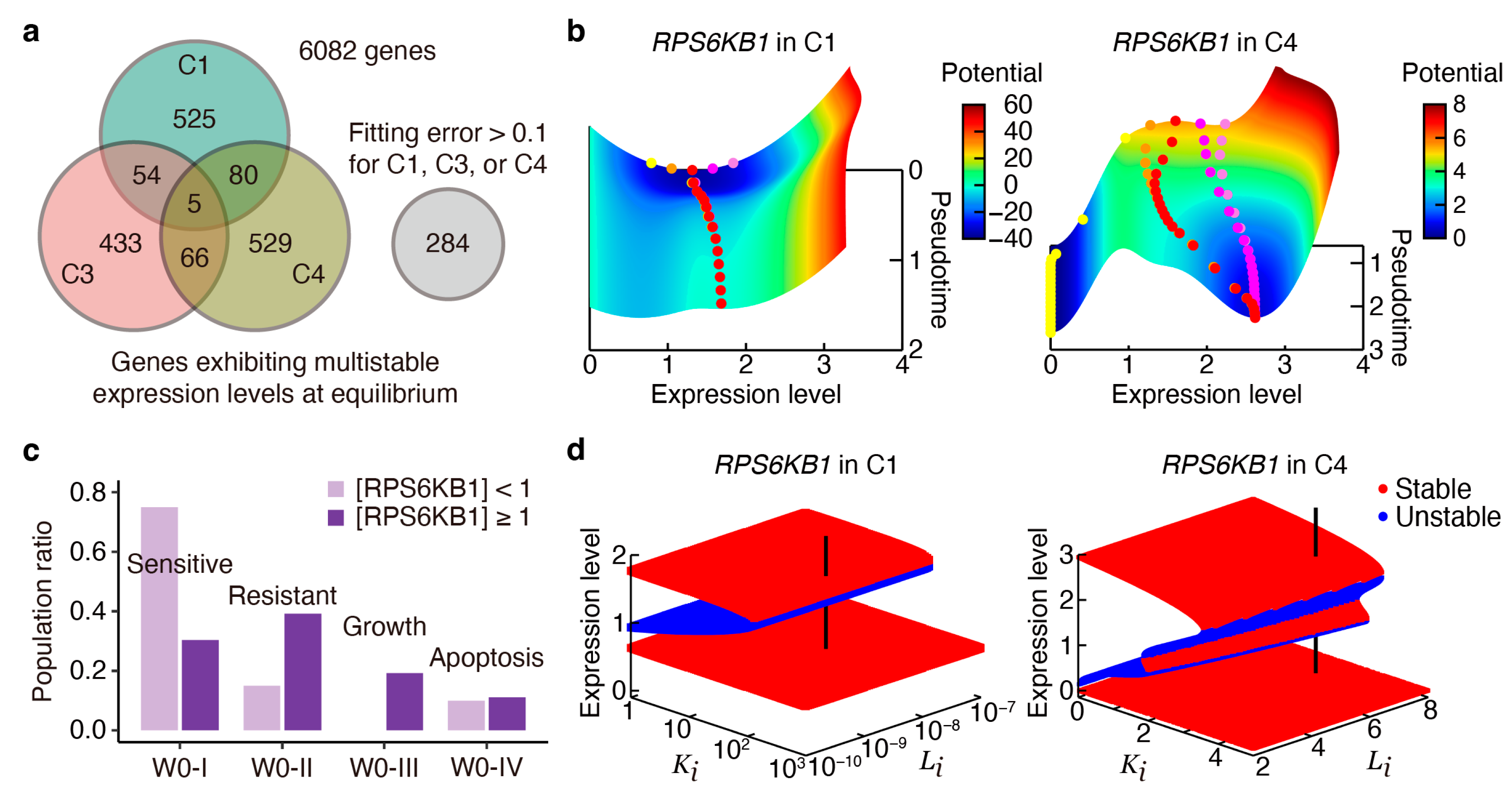
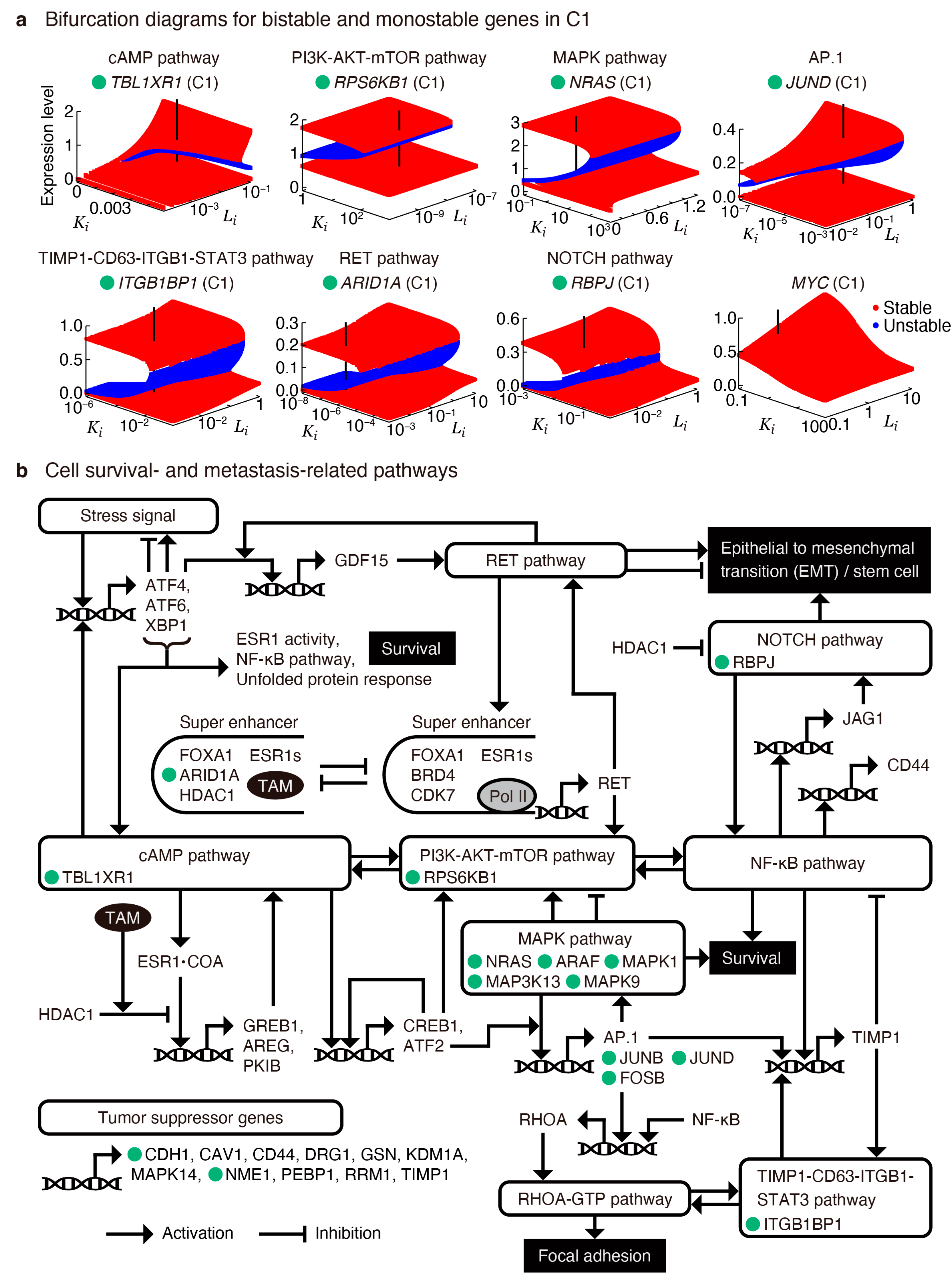
Figure 5.
Integration of key regulatory genes into cell survival and metastasis-related pathways involved in focal adhesion and epithelial–mesenchymal transition (EMT). (a) Bifurcation diagrams for genes exhibiting bistable (TBL1XR1, RPS6KB1, NRAS, JUND, ITGB1BP1, ARID1A, and RBPJ) and monostable (MYC) expressions in C1 population that are involved in cAMP, PI3K-AKT-mTOR, MAPK, AP.1, TIMP-1-CD63-ITGB1-STAT3, RET, and NOTCH pathways. (b) Comprehensive mapping of the key regulatory genes, marked by green-filled circles, into a schematic representation of cell survival and metastasis-related pathways, focusing on those involved in focal adhesion and EMT. The green-labeled genes are shown in (a). TAM, tamoxifen.
Figure 5.
Integration of key regulatory genes into cell survival and metastasis-related pathways involved in focal adhesion and epithelial–mesenchymal transition (EMT). (a) Bifurcation diagrams for genes exhibiting bistable (TBL1XR1, RPS6KB1, NRAS, JUND, ITGB1BP1, ARID1A, and RBPJ) and monostable (MYC) expressions in C1 population that are involved in cAMP, PI3K-AKT-mTOR, MAPK, AP.1, TIMP-1-CD63-ITGB1-STAT3, RET, and NOTCH pathways. (b) Comprehensive mapping of the key regulatory genes, marked by green-filled circles, into a schematic representation of cell survival and metastasis-related pathways, focusing on those involved in focal adhesion and EMT. The green-labeled genes are shown in (a). TAM, tamoxifen.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Iida, K.; Okada, M. Identifying Key Regulatory Genes in Drug Resistance Acquisition: Modeling Pseudotime Trajectories of Breast Cancer Single-Cell Transcriptome. Cancers 2024, 16, 1884. https://doi.org/10.3390/cancers16101884
AMA Style
Iida K, Okada M. Identifying Key Regulatory Genes in Drug Resistance Acquisition: Modeling Pseudotime Trajectories of Breast Cancer Single-Cell Transcriptome. Cancers. 2024; 16(10):1884. https://doi.org/10.3390/cancers16101884
Chicago/Turabian StyleIida, Keita, and Mariko Okada. 2024. "Identifying Key Regulatory Genes in Drug Resistance Acquisition: Modeling Pseudotime Trajectories of Breast Cancer Single-Cell Transcriptome" Cancers 16, no. 10: 1884. https://doi.org/10.3390/cancers16101884
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.