
Dysregulation of lncRNA MALAT1 Contributes to Lung Cancer in African Americans by Modulating the Tumor Immune Microenvironment
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Specimens
2.2. Droplet Digital PCR (ddPCR)
2.3. Cell Culture
2.4. Cell Transfection
2.5. MTT Assay (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide)
2.6. Transwell Assays
2.7. Matrigel Invasion Assay
2.8. Dual Luciferase Reporter Assay
2.9. RNA Pull-Down Assay
2.10. Western Blot Analysis
2.11. Luminex® Multiplex Assays
2.12. Human Lung Cancer Subcutaneous Xenografts in Mice
2.13. Immunohistochemical Analysis of Tumor Tissues
2.14. Bioinformatics Analysis for Gene Interaction
2.15. Statistical Analysis
3. Results
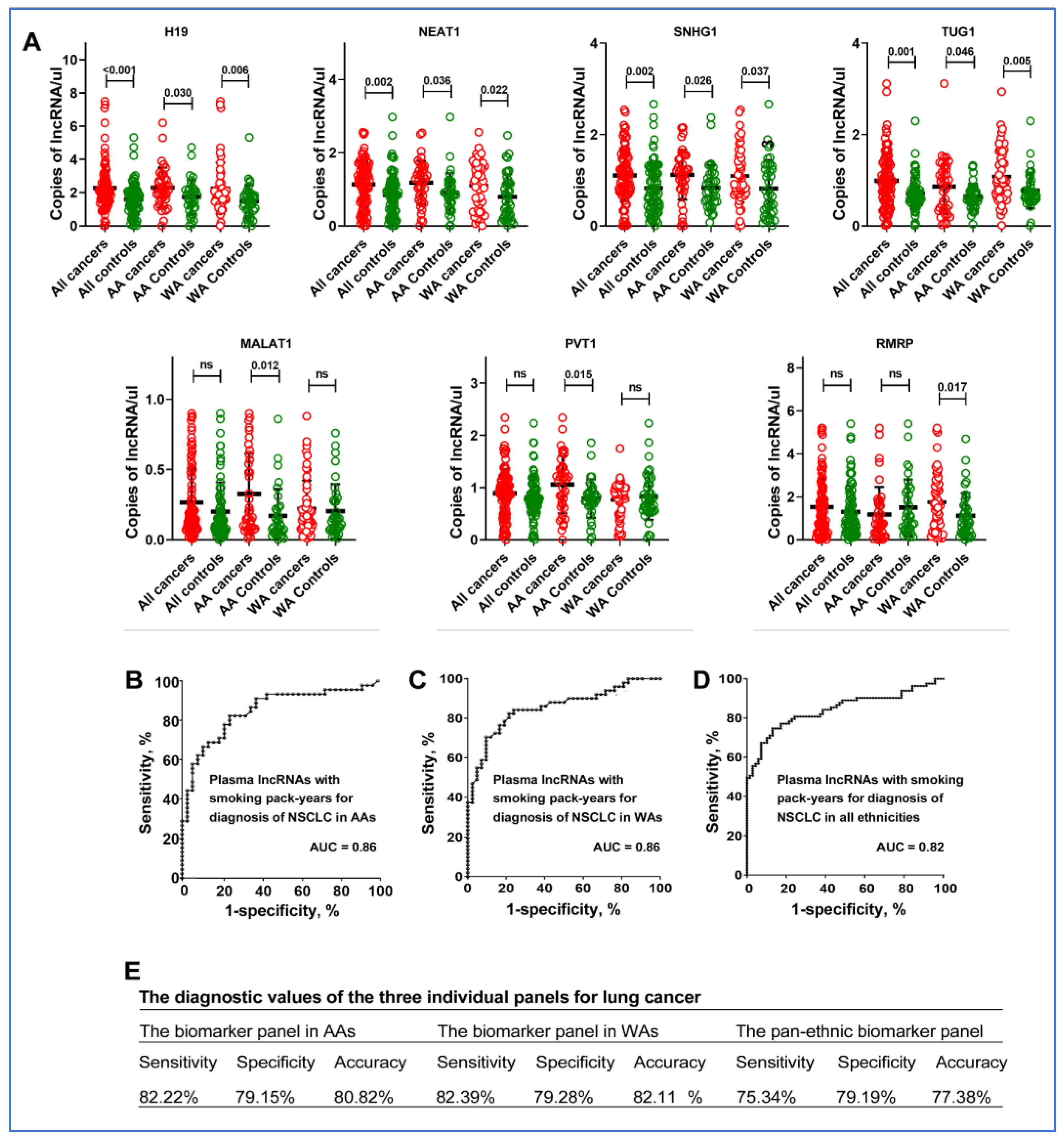
3.1. Differences in Plasma lncRNA Profiles across Ethnic Groups
3.2. The Diagnostic Utility of Plasma lncRNAs for Lung Cancer in AAs
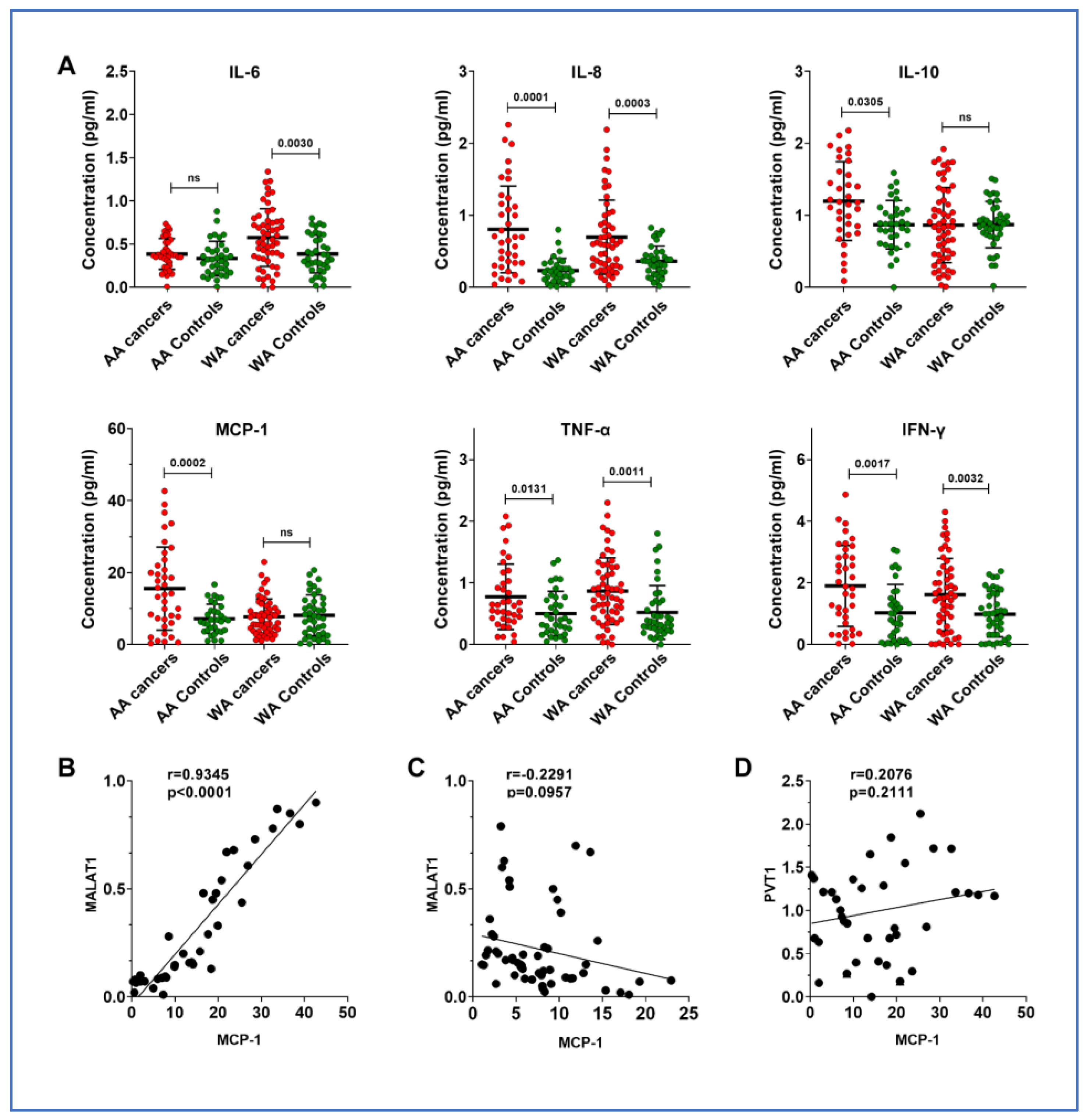
3.3. Overexpression of MALAT1 Is Associated with Monocyte Chemoattractant Protein-1 (MCP-1), a Chemokine, in AA Patients with Lung Cancer
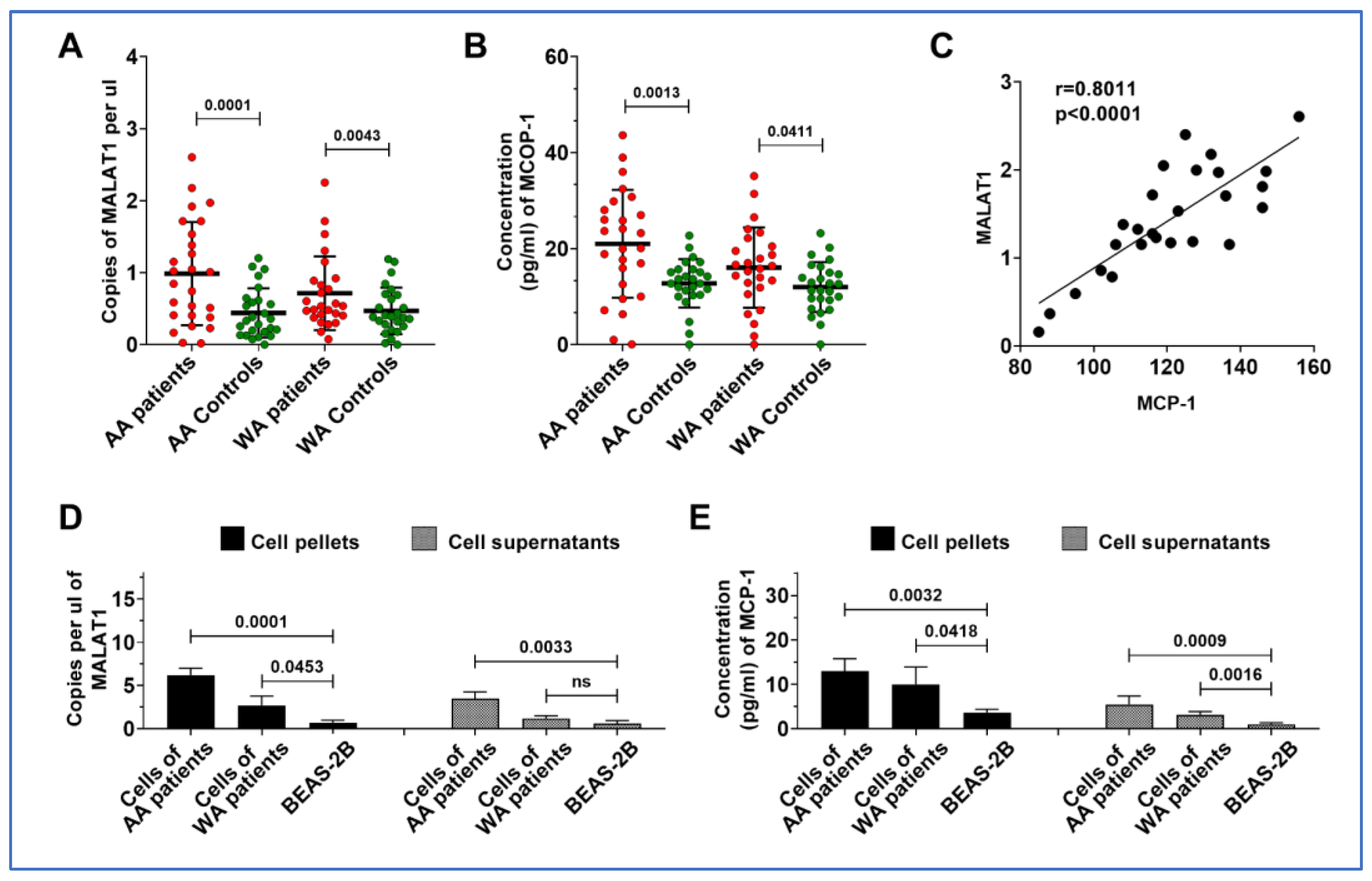
3.4. MALAT1 and MCP-1 Are Both Highly Expressed in Lung Tumor Tissues and Cancer Cell Lines from AA Patients
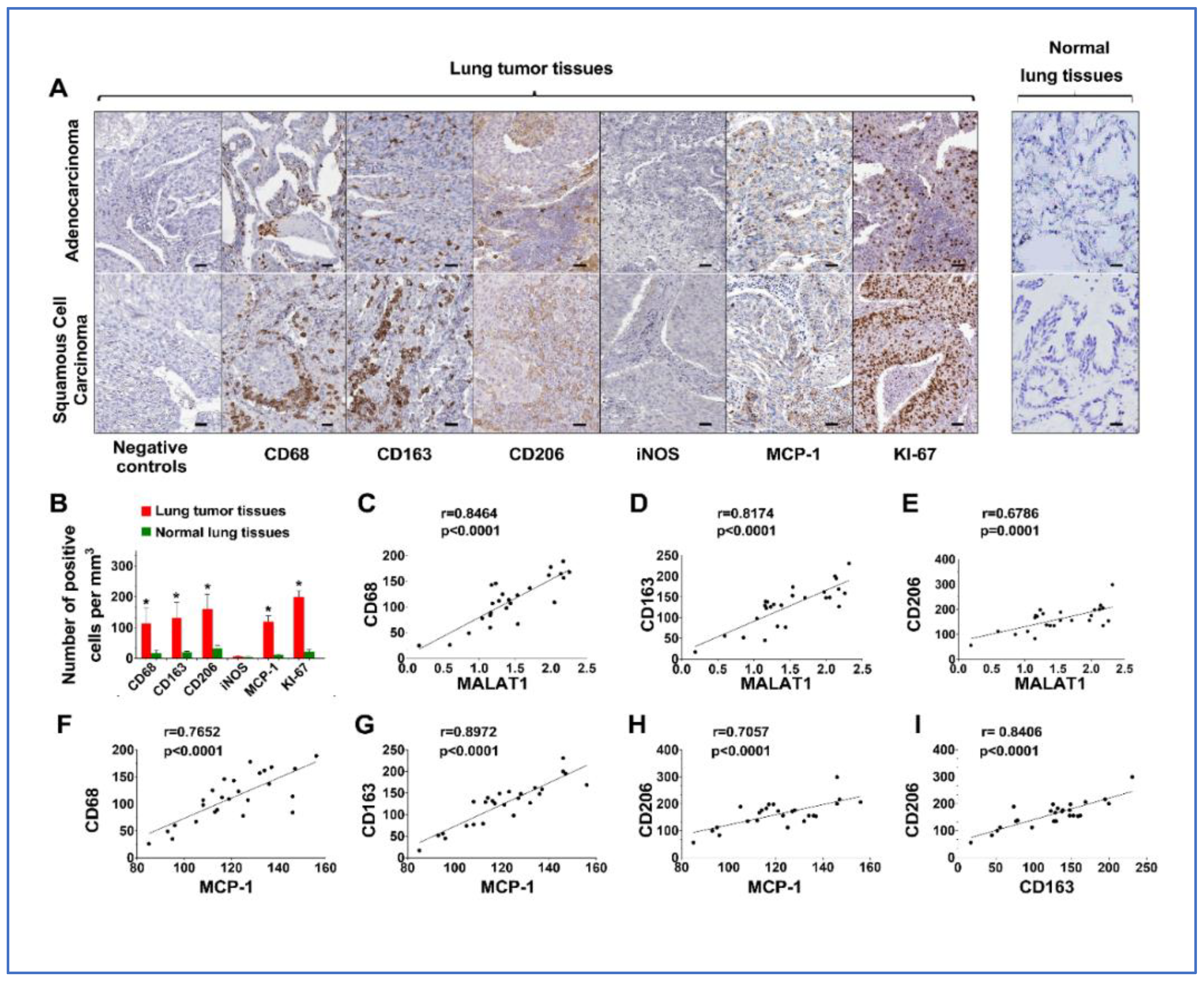
3.5. Upregulation of MALAT1 Enhances Tumor-Associated Macrophage Presence and Alters the Dynamics of the Tumor Immune Microenvironment (TIME)
3.6. Upregulation of MALAT1 Promotes the Tumorigenicity of Lung Cancer by Activating MCP-1 in NSCLC Cells
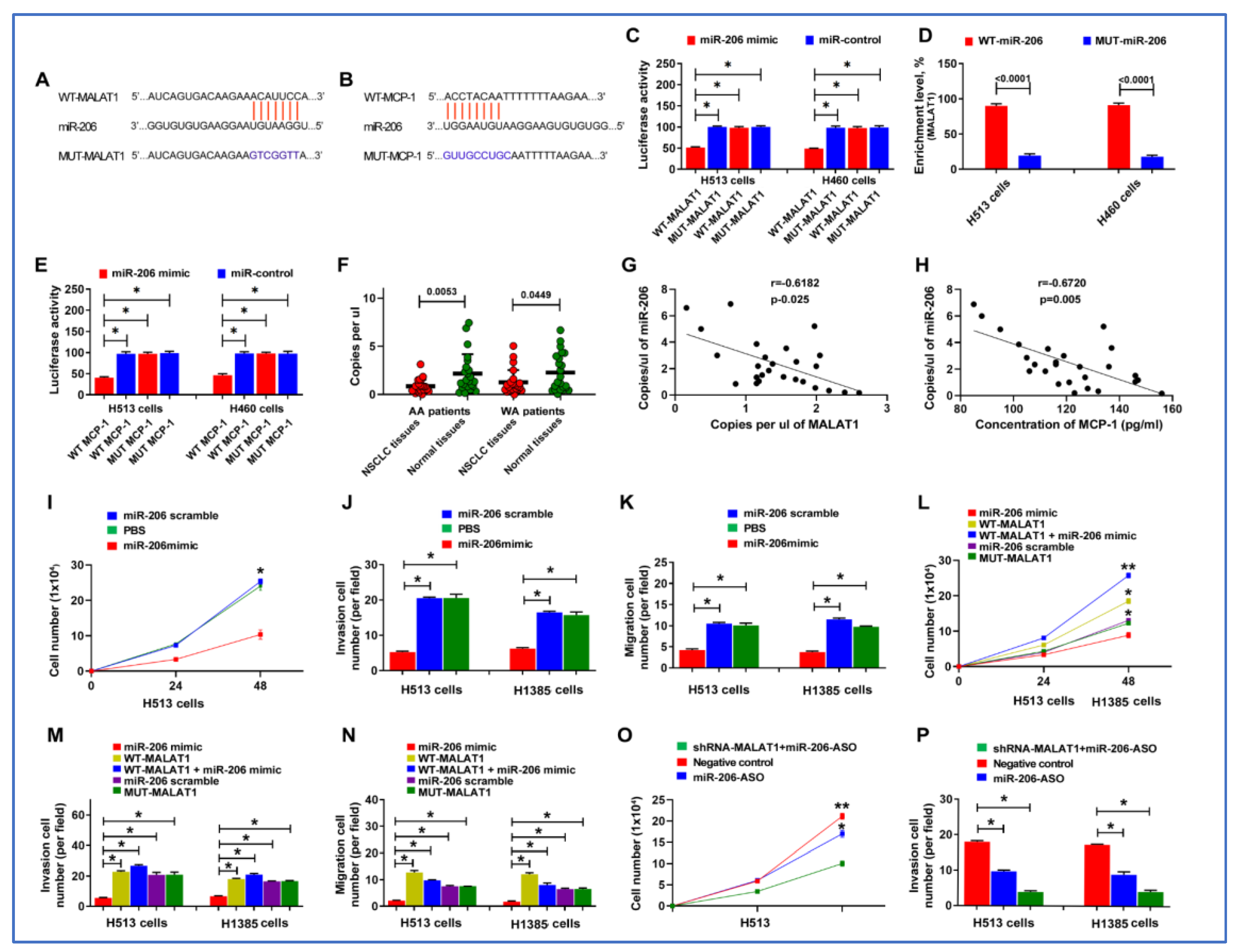
3.7. miR-206 Acts as a Direct Downstream Target of MALAT1
3.8. MALAT1 Enhances Malignant Behaviors of Lung Cancer Cells by Targeting miR-206
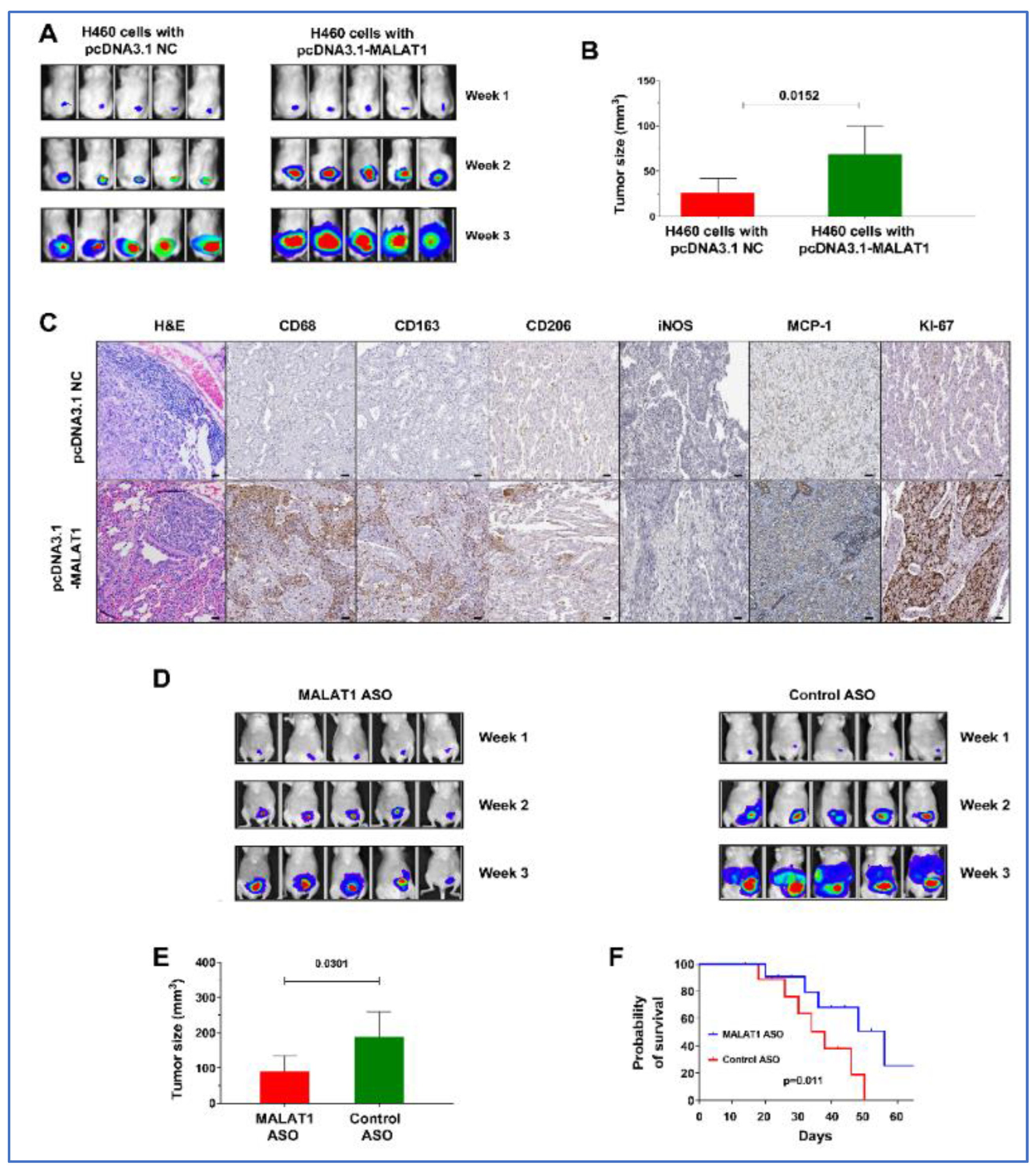
3.9. MALAT1 Promotes Growth of Lung Tumor in Xenograft Animal Models
3.10. Effective Antitumor Effects of MALAT1-Targeting ASOs In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aldrich, M.C.; Mercaldo, S.F.; Sandler, K.L.; Blot, W.J.; Grogan, E.L.; Blume, J.D. Evaluation of USPSTF Lung Cancer Screening Guidelines among African American Adult Smokers. JAMA Oncol. 2019, 5, 1318–1324. [Google Scholar] [CrossRef]
- Zanetti, K.A.; Wang, Z.; Aldrich, M.; Amos, C.I.; Blot, W.J.; Bowman, E.D.; Burdette, L.; Cai, Q.; Caporaso, N.; Chung, C.C.; et al. Genome-wide association study confirms lung cancer susceptibility loci on chromosomes 5p15 and 15q25 in an African-American population. Lung Cancer 2016, 98, 33–42. [Google Scholar] [CrossRef]
- Gujrati, H.; Ha, S.; Waseem, M.; Wang, B.D. Downregulation of miR-99b-5p and Upregulation of Nuclear mTOR Cooperatively Promotes the Tumor Aggressiveness and Drug Resistance in African American Prostate Cancer. Int. J. Mol. Sci. 2022, 23, 9643. [Google Scholar] [CrossRef]
- Leidner, R.S.; Fu, P.; Clifford, B.; Hamdan, A.; Jin, C.; Eisenberg, R.; Boggon, T.J.; Skokan, M.; Franklin, W.A.; Cappuzzo, F.; et al. Genetic abnormalities of the EGFR pathway in African American Patients with non-small-cell lung cancer. J. Clin. Oncol. 2009, 27, 5620–5626. [Google Scholar] [CrossRef]
- Lusk, C.M.; Watza, D.; Dyson, G.; Craig, D.; Ratliff, V.; Wenzlaff, A.S.; Lonardo, F.; Bollig-Fischer, A.; Bepler, G.; Purrington, K.; et al. Profiling the Mutational Landscape in Known Driver Genes and Novel Genes in African American Non-Small Cell Lung Cancer Patients. Clin. Cancer Res. 2019, 25, 4300–4308. [Google Scholar] [CrossRef]
- Enewold, L.; Mechanic, L.E.; Bowman, E.D.; Zheng, Y.L.; Yu, Z.; Trivers, G.; Alberg, A.J.; Harris, C.C. Serum concentrations of cytokines and lung cancer survival in African Americans and Caucasians. Cancer Epidemiol. Biomark. Prev. 2009, 18, 215–222. [Google Scholar] [CrossRef]
- Leng, Q.; Dhilipkannah, P.; Jiang, F. Cytokine Signatures for Lung Cancer Diagnosis in African American Populations. J. Pers. Med. 2024, 14, 117. [Google Scholar] [CrossRef]
- Bhat, A.A.; Afzal, O.; Afzal, M.; Gupta, G.; Thapa, R.; Ali, H.; Hassan Almalki, W.; Kazmi, I.; Alzarea, S.I.; Saleem, S.; et al. MALAT1: A key regulator in lung cancer pathogenesis and therapeutic targeting. Pathol. Res. Pract. 2024, 253, 154991. [Google Scholar] [CrossRef]
- Chen, Z.; Lei, T.; Chen, X.; Gu, J.; Huang, J.; Lu, B.; Wang, Z. Long non-coding RNA in lung cancer. Clin. Chim. Acta 2020, 504, 190–200. [Google Scholar] [CrossRef]
- Ishola, A.A.; La’ah, A.S.; Le, H.D.; Nguyen, V.Q.; Yang, Y.P.; Chou, S.J.; Tai, H.Y.; Chien, C.S.; Wang, M.L. Non-coding RNA and lung cancer progression. J. Chin. Med. Assoc. 2020, 83, 8–14. [Google Scholar] [CrossRef]
- Gao, X.; Wang, N.; Wu, S.; Cui, H.; An, X.; Yang, Y. Long non-coding RNA FER1L4 inhibits cell proliferation and metastasis through regulation of the PI3K/AKT signaling pathway in lung cancer cells. Mol. Med. Rep. 2019, 20, 182–190. [Google Scholar] [CrossRef]
- Niu, Y.; Ma, F.; Huang, W.; Fang, S.; Li, M.; Wei, T.; Guo, L. Long non-coding RNA TUG1 is involved in cell growth and chemoresistance of small cell lung cancer by regulating LIMK2b via EZH2. Mol. Cancer 2017, 16, 5. [Google Scholar] [CrossRef]
- Lin, P.C.; Huang, H.D.; Chang, C.C.; Chang, Y.S.; Yen, J.C.; Lee, C.C.; Chang, W.H.; Liu, T.C.; Chang, J.G. Long noncoding RNA TUG1 is downregulated in non-small cell lung cancer and can regulate CELF1 on binding to PRC2. BMC Cancer 2016, 16, 583. [Google Scholar] [CrossRef]
- Lin, Y.; Leng, Q.; Zhan, M.; Jiang, F. A Plasma Long Noncoding RNA Signature for Early Detection of Lung Cancer. Transl. Oncol. 2018, 11, 1225–1231. [Google Scholar] [CrossRef]
- Lin, Y.; Holden, V.; Dhilipkannah, P.; Deepak, J.; Todd, N.W.; Jiang, F. A Non-Coding RNA Landscape of Bronchial Epitheliums of Lung Cancer Patients. Biomedicines 2020, 8, 88. [Google Scholar] [CrossRef]
- Beebe-Dimmer, J.L.; Ruterbusch, J.J.; Harper, F.W.K.; Baird, T.M.; Finlay, D.G.; Rundle, A.G.; Pandolfi, S.S.; Hastert, T.A.; Schwartz, K.L.; Bepler, G.; et al. Physical activity and quality of life in African American cancer survivors: The Detroit Research on Cancer Survivors study. Cancer 2020, 126, 1987–1994. [Google Scholar] [CrossRef]
- Gong, X.; Zhu, Y.; Chang, H.; Li, Y.; Ma, F. Long noncoding RNA MALAT1 promotes cardiomyocyte apoptosis after myocardial infarction via targeting miR-144-3p. Biosci. Rep. 2019, 39, BSR20191103. [Google Scholar] [CrossRef]
- Wang, Y.; Tai, Q.; Zhang, J.; Kang, J.; Gao, F.; Zhong, F.; Cai, L.; Fang, F.; Gao, Y. MiRNA-206 inhibits hepatocellular carcinoma cell proliferation and migration but promotes apoptosis by modulating cMET expression. Acta Biochim. Biophys. Sin. 2019, 51, 243–253. [Google Scholar] [CrossRef]
- Powe, B.D.; Ross, L.; Cooper, D.L. Attitudes and beliefs about smoking among African-American college students at historically black colleges and universities. J. Natl. Med. Assoc. 2007, 99, 338–344. [Google Scholar]
- Sale, M.M.; Smith, S.G.; Mychaleckyj, J.C.; Keene, K.L.; Langefeld, C.D.; Leak, T.S.; Hicks, P.J.; Bowden, D.W.; Rich, S.S.; Freedman, B.I. Variants of the transcription factor 7-like 2 (TCF7L2) gene are associated with type 2 diabetes in an African-American population enriched for nephropathy. Diabetes 2007, 56, 2638–2642. [Google Scholar] [CrossRef]
- Dupont, N.C.; Wang, K.; Wadhwa, P.D.; Culhane, J.F.; Nelson, E.L. Validation and comparison of luminex multiplex cytokine analysis kits with ELISA: Determinations of a panel of nine cytokines in clinical sample culture supernatants. J. Reprod. Immunol. 2005, 66, 175–191. [Google Scholar] [CrossRef]
- Power, D.; Lagunes, D.R. Adenocarcinoma of unknown primary in a 20-year-old African American male. Clin. Genitourin. Cancer 2009, 7, E45–E48. [Google Scholar] [CrossRef]
- Rakaee, M.; Busund, L.R.; Jamaly, S.; Paulsen, E.E.; Richardsen, E.; Andersen, S.; Al-Saad, S.; Bremnes, R.M.; Donnem, T.; Kilvaer, T.K. Prognostic Value of Macrophage Phenotypes in Resectable Non-Small Cell Lung Cancer Assessed by Multiplex Immunohistochemistry. Neoplasia 2019, 21, 282–293. [Google Scholar] [CrossRef]
- Mina, N.; Soubani, A.O.; Cote, M.L.; Suwan, T.; Wenzlaff, A.S.; Jhajhria, S.; Samarah, H.; Schwartz, A.G. The relationship between chronic obstructive pulmonary disease and lung cancer in African American patients. Clin. Lung Cancer 2012, 13, 149–156. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Gandhi, K.K.; Foulds, J.; Steinberg, M.B.; Lu, S.E.; Williams, J.M. Lower quit rates among African American and Latino menthol cigarette smokers at a tobacco treatment clinic. Int. J. Clin. Pract. 2009, 63, 360–367. [Google Scholar] [CrossRef]
- Mak, A.C.Y.; Sajuthi, S.; Joo, J.; Xiao, S.; Sleiman, P.M.; White, M.J.; Lee, E.Y.; Saef, B.; Hu, D.; Gui, H.; et al. Lung Function in African American Children with Asthma Is Associated with Novel Regulatory Variants of the KIT Ligand KITLG/SCF and Gene-By-Air-Pollution Interaction. Genetics 2020, 215, 869–886. [Google Scholar] [CrossRef]
- Neergheen, V.L.; Topel, M.; Van Dyke, M.E.; Sullivan, S.; Pemu, P.E.; Gibbons, G.H.; Vaccarino, V.; Quyyumi, A.A.; Lewis, T.T. Neighborhood social cohesion is associated with lower levels of interleukin-6 in African American women. Brain Behav. Immun. 2019, 76, 28–36. [Google Scholar] [CrossRef]
- Panch, S.R.; Yau, Y.Y.; Fitzhugh, C.D.; Hsieh, M.M.; Tisdale, J.F.; Leitman, S.F. Hematopoietic progenitor cell mobilization is more robust in healthy African American compared to Caucasian donors and is not affected by the presence of sickle cell trait. Transfusion 2016, 56, 1058–1065. [Google Scholar] [CrossRef]
- Demerath, E.W.; Guan, W.; Grove, M.L.; Aslibekyan, S.; Mendelson, M.; Zhou, Y.H.; Hedman, A.K.; Sandling, J.K.; Li, L.A.; Irvin, M.R.; et al. Epigenome-wide association study (EWAS) of BMI, BMI change and waist circumference in African American adults identifies multiple replicated loci. Hum. Mol. Genet. 2015, 24, 4464–4479. [Google Scholar] [CrossRef]
- Pichardo, C.M.; Ezeani, A.; Pichardo, M.S.; Agurs-Collins, T.; Powell-Wiley, T.M.; Ryan, B.; Minas, T.Z.; Bailey-Whyte, M.; Tang, W.; Dorsey, T.H.; et al. Association of neighborhood gentrification with prostate cancer and immune markers in African American and European American men. Cancer Med. 2023, 13, e6828. [Google Scholar] [CrossRef]
- Roh, J.; Kim, B.; Im, M.; Jang, W.; Chae, Y.; Kang, J.; Youn, B.; Kim, W. MALAT1-regulated gene expression profiling in lung cancer cell lines. BMC Cancer 2023, 23, 818. [Google Scholar] [CrossRef]
- Xin, R.; Hu, B.; Qu, D.; Chen, D. Withdrawn: Oncogenic lncRNA MALAT-1 recruits E2F1 to upregulate RAD51 expression and thus promotes cell autophagy and tumor growth in non-small cell lung cancer. Pulm. Pharmacol. Ther. 2023, 102199. [Google Scholar] [CrossRef]
- Tiansheng, G.; Junming, H.; Xiaoyun, W.; Peixi, C.; Shaoshan, D.; Qianping, C. lncRNA Metastasis-Associated Lung Adenocarcinoma Transcript 1 Promotes Proliferation and Invasion of Non-Small Cell Lung Cancer Cells via Down-Regulating miR-202 Expression. Cell J. 2020, 22, 375–385. [Google Scholar] [CrossRef]
- Wei, S.; Wang, K.; Huang, X.; Zhao, Z. LncRNA MALAT1 contributes to non-small cell lung cancer progression via modulating miR-200a-3p/programmed death-ligand 1 axis. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419859699. [Google Scholar] [CrossRef]
- Feng, C.; Zhao, Y.; Li, Y.; Zhang, T.; Ma, Y.; Liu, Y. LncRNA MALAT1 Promotes Lung Cancer Proliferation and Gefitinib Resistance by Acting as a miR-200a Sponge. Arch. Bronconeumol. 2019, 55, 627–633. [Google Scholar] [CrossRef]
- Kalinski, P.; Talmadge, J.E. Tumor Immuno-Environment in Cancer Progression and Therapy. Adv. Exp. Med. Biol. 2017, 1036, 1–18. [Google Scholar] [CrossRef]
- McClellan, J.L.; Davis, J.M.; Steiner, J.L.; Enos, R.T.; Jung, S.H.; Carson, J.A.; Pena, M.M.; Carnevale, K.A.; Berger, F.G.; Murphy, E.A. Linking tumor-associated macrophages, inflammation, and intestinal tumorigenesis: Role of MCP-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1087–G1095. [Google Scholar] [CrossRef]
- Lanctot, J.Q.; Klesges, R.C.; Stockton, M.B.; Klesges, L.M. Prevalence and characteristics of energy underreporting in African-American girls. Obesity 2008, 16, 1407–1412. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasma Specimens from Patients with NSCLC and Cancer-Free Smokers Were Analyzed | Tissue Specimens Were Analyzed from Forty-Six Patients with NSCLC | ||
|---|---|---|---|
| NSCLC cases (n = 92) | Controls (n = 76) | ||
| Age | 69.13 (SD 11.37) | 67.85 (SD 10.34) | 68.3 ± 7.5 |
| Sex | |||
| Female | 24 | 20 | 29 |
| Male | 68 | 56 | 17 |
| Race | |||
| African Americans | 38 | 35 | 23 |
| White Americans | 54 | 41 | 23 |
| Smoking pack-years (median) | 32.9 | 26.5 | 39 |
| Stage | |||
| Stage I | 23 | 24 | |
| Stage II | 35 | 22 | |
| Stage III–IV | 34 | 8 | |
| Histological type | |||
| Adenocarcinoma | 52 | 26 | |
| Squamous cell carcinoma | 40 | 20 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Dhilipkannah, P.; Holden, V.K.; Sachdeva, A.; Todd, N.W.; Jiang, F. Dysregulation of lncRNA MALAT1 Contributes to Lung Cancer in African Americans by Modulating the Tumor Immune Microenvironment. Cancers 2024, 16, 1876. https://doi.org/10.3390/cancers16101876
Li J, Dhilipkannah P, Holden VK, Sachdeva A, Todd NW, Jiang F. Dysregulation of lncRNA MALAT1 Contributes to Lung Cancer in African Americans by Modulating the Tumor Immune Microenvironment. Cancers. 2024; 16(10):1876. https://doi.org/10.3390/cancers16101876
Chicago/Turabian StyleLi, Jin, Pushpa Dhilipkannah, Van K. Holden, Ashutosh Sachdeva, Nevins W. Todd, and Feng Jiang. 2024. "Dysregulation of lncRNA MALAT1 Contributes to Lung Cancer in African Americans by Modulating the Tumor Immune Microenvironment" Cancers 16, no. 10: 1876. https://doi.org/10.3390/cancers16101876
APA StyleLi, J., Dhilipkannah, P., Holden, V. K., Sachdeva, A., Todd, N. W., & Jiang, F. (2024). Dysregulation of lncRNA MALAT1 Contributes to Lung Cancer in African Americans by Modulating the Tumor Immune Microenvironment. Cancers, 16(10), 1876. https://doi.org/10.3390/cancers16101876