
Study on the Effect and Enhancement of Near-Natural Integrated Plant Positioning Configuration in the Hilly Gully Region, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Research Methodology
2.2.1. Plot Layout and Vegetation Survey
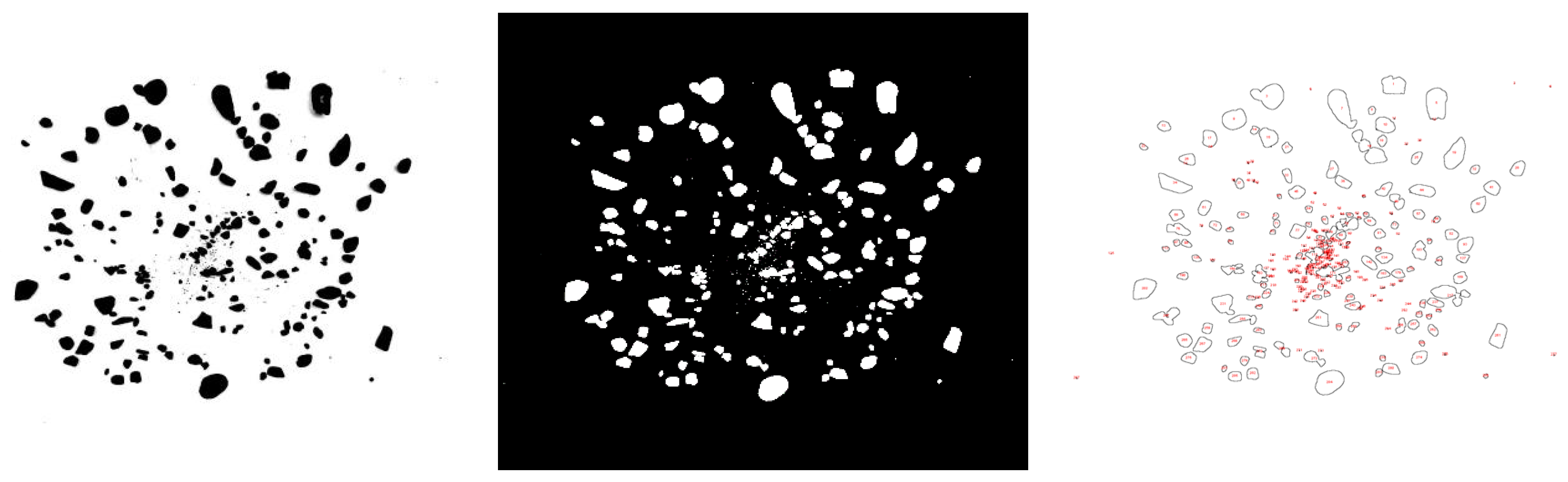
2.2.2. Determination of the Surface Gravel Coverage
2.2.3. M. Godron’s Method for Determining Community Structural Stability
2.2.4. Investigation of the Water Retention Function of Litterfall
Measurement of Litterfall Water-Holding Capacity
Evaluation of Water Retention Function of Litterfall
2.2.5. Saturated Hydraulic Conductivity of Soil
2.2.6. Calculation of Species Diversity
2.2.7. Data Processing
3. Results and Analysis
3.1. Characteristics of Plant Communities in the Study Area
Analysis of Plant Diversity Indices under Different Configuration Modes
3.2. Protective Forest Configuration Effects at the Slope Top
3.2.1. Analysis of Surface Gravel Coverage under the Canopy
3.2.2. Comparison of Growth Conditions between the Two Configuration Modes
3.3. Protective Forest Configuration Mode Effect on the Slope Middle
3.3.1. Analysis of Forest Stand Community Structural Stability
3.3.2. Hydrological Benefits of Understory Litter under Different Configuration Modes
3.4. Protective Effect of Slope Bottom Configuration Modes
3.4.1. Analysis of Ks
3.4.2. Variation Coefficient of Ks
4. Discussion
4.1. Diversity of Different Forest Stand Structures
4.2. Community Structure Stability
4.2.1. Relationship between Plant Diversity and Community Structure Stability
4.2.2. Community Stability Research
4.3. Factors Affecting the Water-Holding and Retention Abilities of Litter
4.4. Factors Influencing the Ks
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Grima, N.; Edwards, D.; Edwards, F.; Petley, D.; Fisher, B. Landslides in the Andes: Forests can provide cost-effective landslide regulation services. Sci. Total. Environ. 2020, 745, 141128. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tao, J.; Wu, G.H. Analysis of Ecological Slope Protection Technology in River Regulation. In Proceedings of the 2019 International Conference on Management, Finance and Social Sciences Research (MFSSR 2019), Lyon, France, 2–4 August 2019; pp. 19–22. [Google Scholar] [CrossRef]
- Zhai, J.; Wang, L.; Liu, Y.; Wang, C. Assessing the effects of China’s Three-North Shelter Forest Program over 40 years. Sci. Total Environ. 2022, 857, 159354. [Google Scholar] [CrossRef]
- Gyssels, G.; Poesen, J. The importance of plant root characteristics in controlling concentrated flow erosion rates. Earth Surf. Process. Landf. 2003, 28, 371–384. [Google Scholar] [CrossRef]
- Rey, F. Effectiveness of vegetation barriers for marly sediment trapping. Earth Surf. Process. Landf. 2004, 29, 1161–1169. [Google Scholar] [CrossRef]
- Wang, Y.; Shangguan, Z. Formation mechanisms and remediation techniques for low-efficiency artificial shelter forests on the Chinese Loess Plateau. J. Arid. Land 2022, 14, 837–848. [Google Scholar] [CrossRef]
- Wang, S.; Fu, B.; Gao, G.; Liu, Y.; Zhou, J. Responses of soil moisture in different land cover types to rainfall events in a re-vegetation catchment area of the Loess Plateau, China. CATENA 2013, 101, 122–128. [Google Scholar] [CrossRef]
- Feng, X.M.; Wang, Y.F.; Chen, L.D.; Fu, B.; Bai, G. Modeling soil erosion and its response to land-use change in hilly catchments of the Chinese Loess Plateau. Geomorphology 2010, 118, 239–248. [Google Scholar] [CrossRef]
- Peng, T.; Wang, S.J. Effects of land use, land cover and soil loss on karts slopes in southwest China. and rainfall regimes on the surface runoff. CATENA 2012, 90, 53–62. [Google Scholar] [CrossRef]
- Wei, W.; Chen, L.; Fu, B.; Huang, Z.; Wu, D.; Gui, L. The effect of land uses andrainfall regimes on runoff and soil erosion in the semi-arid loess hilly area, China. J. Hydrol. 2007, 335, 247–258. [Google Scholar] [CrossRef]
- Andres, P.; Jorba, M. Mitigation strategies in some motorway embankments (Catalonia, Spain). Restor. Ecol. 2000, 8, 268–275. [Google Scholar] [CrossRef]
- Yang, F.; Zhou, Y. Quantifying spatial scale of positive and negative terrains pattern at watershed-scale: Case in soil and water conservation region on Loess Plateau. J. Mt. Sci. 2017, 14, 1642–1654. [Google Scholar] [CrossRef]
- Li, L.; Li, X.-Y.; Zhang, S.-Y.; Jiang, Z.-Y.; Zheng, X.-R.; Hu, X.; Huang, Y.-M. Stemflow and its controlling factors in the subshrub Artemisia ordosica during two contrasting growth stages in the Mu Us sandy land of northern China. Hydrol. Res. 2016, 47, 409–418. [Google Scholar] [CrossRef]
- Shen, E.; Liu, G.; Xia, X.; Dan, C.; Zheng, F.; Zhang, Q.; Zhang, Y.; Guo, Z. Resistance to sheet flow induced by raindrop impact on rough surfaces. CATENA 2023, 231, 5514. [Google Scholar] [CrossRef]
- Hu, Y.; Li, H.; Wu, D.; Chen, W.; Zhao, X.; Hou, M.; Li, A.; Zhu, Y. LAI-indicated vegetation dynamic in ecologically fragile region: A case study in the Three-North Shelter Forest program region of China. Ecol. Indic. 2021, 120, 106932. [Google Scholar] [CrossRef]
- Wen, B.; Duan, G.; Lu, J.; Zhou, R.; Ren, H.; Wen, Z. Response relationship between vegetation structure and runoff-sediment yield in the hilly and gully area of the Loess Plateau, China. CATENA 2023, 227, 107107. [Google Scholar] [CrossRef]
- Chen, Z.; Guo, M.; Wang, W.; Wang, W.; Feng, L. Response of soil erodibility of permanent gully heads to revegetation along a vegetation zone gradient in the loess-table and gully region of the Chinese Loess Plateau. Sci. Total. Environ. 2023, 892, 164833. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Feng, S.; Wang, S.; Zhang, B.; Ning, Z.; Wang, R.; Chen, X.; Yu, L.; Zhao, H.; Lan, D.; et al. Patterns and driving mechanism of soil organic carbon, nitrogen, and phosphorus stoichiometry across northern China’s desert-grassland transition zone. CATENA 2023, 220, 106695. [Google Scholar] [CrossRef]
- Lu, J.; Zhao, X.; Wang, S.; Feng, S.; Ning, Z.; Wang, R.; Chen, X.; Zhao, H.; Chen, M. Untangling the influence of abiotic and biotic factors on leaf C, N, and P stoichiometry along a desert-grassland transition zone in northern China. Sci. Total. Environ. 2023, 884, 163902. [Google Scholar] [CrossRef]
- Katra, I.; Lavee, H.; Sarah, P. The effect of rock fragment size and position on topsoil moisture on arid and semr arid hillslopes. CATENA 2008, 72, 49–55. [Google Scholar] [CrossRef]
- Yang, Z.; van Ruijven, J.; Du, G. The effects of long-term fertilization on the teporal stability ofalpine meadow communities. Plant Soil 2011, 345, 315–324. [Google Scholar] [CrossRef]
- Xie, P.; Liu, T.; Chen, H.; Su, Z. Community Structure and Soil Mineral Concentration in Relation to Plant Invasion in a Subtropical Urban and Rural Ecotone. Forests 2021, 12, 185. [Google Scholar] [CrossRef]
- Lamit, L.J.; Wojtowicz, T.; Kovacs, Z.; Wooley, S.C.; Zinkgraf, M.; Whitham, T.G.; Lindroth, R.L.; Gehring, C.A. Hybridization among foundation tree species influences the structure of associated understory plant communities. Botany 2011, 89, 165–174. [Google Scholar] [CrossRef]
- Wong, N.K.; Morgan, J.W. Experimental changes in disturbance type do not induce short-term shifts in plant community structure in three semi-arid grasslands of the Victorian Riverine Plain managed for nature conservation. Ecol. Manag. Restor. 2012, 13, 175–182. [Google Scholar] [CrossRef]
- Fu, T.; Chen, H.; Zhang, W.; Nie, Y.; Wang, K. Vertical distribution of soil saturated hydraulic conductivity and its influencing factors in a small karst catchment in Southwest China. Environ. Monit. Assess. 2015, 187, 92. [Google Scholar] [CrossRef]
- Pereira, L.C.; Balbinot, L.; Lima, M.T.; Bramorski, J.; Tonello, K.C. Aspects of forest restoration and hydrology: The hydrological function of litter. J. For. Res. 2021, 33, 543–552. [Google Scholar] [CrossRef]
- Morbidelli, R.; Saltalippi, C.; Flammini, A.; Cifrodelli, M.; Picciafuoco, T.; Corradini, C.; Govindaraju, R. In situ measurements of soil saturated hydraulic conductivity: Assessment of reliability through rainfall–runoff experiments. Hydrol. Process. 2017, 31, 3084–3094. [Google Scholar] [CrossRef]
- Aşkın, T. Soil Saturated Hydraulic Conductivity: A Study on Path Analysis in Clayey Soils. Res. Agric. Sci. 2005, 36, 23–25. [Google Scholar]
- Chand, H.; Verma, K.S.; Bhardwaj, S.K.; Reddy, M.C. Assessment of floral diversity in north western Himalayas—A case study from Kol Dam watershed. Ecol. Environ. Conserv. 2014, 20, 823–828. [Google Scholar]
- Chen, Y.; Cao, Y. Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China. Ecol. Eng. 2014, 73, 238–245. [Google Scholar] [CrossRef]
- Steur, G.; Verburg, R.W.; Wassen, M.J.; Teunissen, P.A.; Verweij, P.A. Exploring relationships between abundance of non-timber forest product species and tropical forest plant diversity. Ecol. Indic. 2020, 121, 107202. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, X.; Li, G.; Zhu, H.; Mao, X.; Yuan, X. Time effect of young shrub roots on slope protection of loess area in Northeast Qinghai-Tibetan plateau. Editor. Off. Trans. Chin. Soc. Agric. Eng. 2012, 55, 136–141. [Google Scholar]
- Hao, G.; Yang, N.; Dong, K.; Xu, Y.; Ding, X.; Shi, X.; Chen, L.; Wang, J.; Zhao, N.; Gao, Y. Shrub-encroached grassland as an alternative stable state in semiarid steppe regions: Evidence from community stability and assembly. Land Degrad. Dev. 2021, 32, 3142–3153. [Google Scholar] [CrossRef]
- Celentano, D.; Zahawi, R.A.; Finegan, B.; Casanoves, F.; Ostertag, R.; Cole, R.J.; Holl, K.D. Tropical forest restoration in Costa Rica: The effect of several strategies on litter production, accumulation and decomposition. Rev. Biol. Trop. 2011, 59, 1323–1336. [Google Scholar]
- Zuo, Y.; He, K. Evaluation and Development of Pedo-Transfer Functions for Predicting Soil Saturated Hydraulic Conductivity in the Alpine Frigid Hilly Region of Qinghai Province. Agronomy 2021, 11, 1581. [Google Scholar] [CrossRef]
- Wan, H.; Bai, Y.; Schönbach, P.; Gierus, M.; Taube, F. Effects of grazing management system on plant community structure and functioning in a semiarid steppe: Scaling from species to community. Plant Soil 2011, 340, 215–226. [Google Scholar] [CrossRef]
- Faith, D.P.; Walker, P.A. The role of trade-offs in biodiversity conservation planning: Linking local management, regional planning and global conservation efforts. J. Biosci. 2002, 27, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Alemayhu, A. Role of Participatory Forest Management in Woody Species Diversity and Forest Conservation: The Case of Gimbo Woreda in Keffa Zone South West Ethiopia. J. Environ. Earth Sci. 2019, 9, 1–12. [Google Scholar]
- Zhang, W.; Huang, D.; Wang, R.; Liu, J.; Du, N. Altitudinal Patterns of Species Diversity and Phylogenetic Diversity across Temperate Mountain Forests of Northern China. PLoS ONE 2016, 11, e0159995. [Google Scholar] [CrossRef]
- Van Do, T.; Kozan, O.; Tuan, T.M. Altitudinal Changes in Species Diversity and Stand Structure of Tropical Forest, Vietnam. Annu. Res. Rev. Biol. 2014, 6, 156–165. [Google Scholar] [CrossRef]
- Zelikova, T.J.; Blumenthal, D.M.; Williams, D.G.; Souza, L.; LeCain, D.R.; Morgan, J.; Pendall, E. Long-term exposure to elevated CO2 enhances plant community stability by suppressing dominant plant species in a mixed-grass prairie. Proc. Natl. Acad. Sci. USA 2014, 111, 15456–15461. [Google Scholar] [CrossRef]
- Fakhry, A.M.; khazzan, M.M.; Aljedaani, G.S. Impact of disturbance on species diversity and composition of Cyperus conglomeratus plant community in southern Jeddah, Saudi Arabia. J. King Saud Univ.-Sci. 2020, 32, 600–605. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, W. Grassland degradation alters the effect of nitrogen enrichment on the multidimensional stability of plant community productivity. Journal of Applied Ecology. J. Appl. Ecol. 2023, 60, 2437–2448. [Google Scholar] [CrossRef]
- De Keersmaecker, W.; Lhermitte, S.; Honnay, O.; Farifteh, J.; Somers, B.; Coppin, P. How to measure ecosystem stability? An evaluation of the reliability of stability metrics based on remote sensing time series across the major global ecosystems. Glob. Change Biol. 2014, 20, 2149–2161. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Yu, Q.; Grogan, P. Cryptogam plant community stability: Warming weakens influences of species richness but enhances effects of evenness. Ecology 2022, 104, e3842. [Google Scholar] [CrossRef] [PubMed]
- Birmele, J.; Kopp, G.; Brodbeck, F.; Konold, W.; Sauter, U.H. Successional changes of phytodiversity on a short rotation coppice plantation in Oberschwaben, Germany. Front. Plant Sci. 2015, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Liu, H.; Yin, Y.; Song, Z. The Role of Legumes in Plant Community Succession of Degraded Grasslands in Northern China. Land Degrad. Dev. 2016, 27, 366–372. [Google Scholar] [CrossRef]
- Wright, A.J.; Ebeling, A.; de Kroon, H.; Roscher, C.; Weigelt, A.; Buchmann, N.; Buchmann, T.; Fischer, C.; Hacker, N.; Hildebrandt, A.; et al. Flooding disturbances increase resource availability and productivity but reduce stability in diverse plant communities. Nat. Commun. 2015, 6, 6092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tan, L.; Cai, Q.; Ye, L. Environmental factors indirectly reduce phytoplankton community stability via functional diversity. Front. Ecol. Evol. 2022, 10, 990835. [Google Scholar] [CrossRef]
- Song, X.; Yan, C.; Xie, J.; Li, S. Assessment of changes in the area of the water conservation forest in the Qilian Mountains of China’s Gansu province, and the effects on water conservation. Environ. Earth Sci. 2012, 66, 2441–2448. [Google Scholar] [CrossRef]
- Yao, Z.; Zhang, X.; Wang, X.; Shu, Q.; Liu, X.; Wu, H.; Gao, S. Functional Diversity of Soil Microorganisms and Influencing Factors in Three Typical Water-Conservation Forests in Danjiangkou Reservoir Area. Forests 2022, 14, 67. [Google Scholar] [CrossRef]
- Cao, C.; Jiang, S.; Ying, Z.; Zhang, F.; Han, X. Spatial variability of soil nutrients and microbiological properties after the establishment of leguminous shrub Caragana microphylla Lam. plantation on sand dune in the Horqin Sandy Land of Northeast China. Ecol. Eng. 2011, 37, 1467–1475. [Google Scholar] [CrossRef]
- Horn, R.; Mordhorst, A.; Fleige, H.; Zimmermann, I.; Burbaum, B.; Filipinski, M.; Cordsen, E. Soil type and land use effects on tensorial properties of saturated hydraulic conductivity in northern Germany. Eur. J. Soil Sci. 2020, 71, 179–189. [Google Scholar] [CrossRef]
- Hao, M.; Zhang, J.; Meng, M.; Chen, H.Y.H.; Guo, X.; Liu, S.; Ye, L. Impacts of changes in vegetation on saturated hydraulic conductivity of soil in subtropical forests. Sci. Rep. 2019, 9, 8372. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, W.; Liao, X.; Tan, X.; Yue, J.; Zhang, W.; Wu, J.; Willison, J.H.M.; Tian, Q.; Liu, Y. Soil nutrients, enzyme activities, and bacterial communities in varied plant communities in karst rocky desertification regions in Wushan County, Southwest China. Front. Microbiol. 2023, 14, 1180562. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xie, Y.; Liu, Z.; Shi, L.; Liu, X.; Liang, M.; Yu, S. Plant species identity and mycorrhizal type explain the root-associated fungal pathogen community assembly of seedlings based on functional traits in a subtropical forest. Front. Plant Sci. 2023, 14, 1251934. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xu, G.; Ma, T.; Chen, L.; Cheng, Y.; Li, P.; Li, Z.; Zhang, Y. Effects of vegetation restoration on soil aggregates, organic carbon, and nitrogen in the Loess Plateau of China. CATENA 2023, 231, 107340. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pattern | Altitude/m | Slope/° | Aspect of Slope | Different Slope Position Configuration Structure | ||
|---|---|---|---|---|---|---|
| Slope Top | Slope Middle | Slope Bottom | ||||
| 1 | 1439 | 15 | NW | Grass | arbor + Shrub + Grass | Grass |
| 2 | 1418 | 13 | NW | arbor + Grass | arbor + Shrub + Grass | Shrub + Grass |
| 3 | 1416 | 16 | NW | arbor + Grass | Shrub + Grass | arbor + Grass |
| 4 | 1426 | 14 | NW | arbor + Grass | arbor + Shrub + Grass | arbor + Shrub + Grass |
| 5 | 1420 | 14 | NW | Grass | Grass | Grass |
| 6 | 1431 | 13 | NW | Grass | Shrub + Grass | Shrub + Grass |
| 7 | 1425 | 15 | NW | Shrub + Grass | Grass | Shrub + Grass |
| 8 | 1427 | 15 | NW | Shrub + Grass | Grass | Grass |
| Pattern | Position | Species of Arbors | Configuration Structure (Plant Space × Row Space) | Quantity (Strain/hm2) |
|---|---|---|---|---|
| P-1 | Slope top | Herbal | —— | —— |
| Slope middle | P. tabuliformis Caragana korshinskii | 5 m × 6 m 6 m × 1 m | 333 1667 | |
| Slope bottom | Herbal | —— | —— | |
| P-2 | Slope top | C. korshinskii | 1 m × 5 m | 2000 |
| Slope middle | P. tabuliformis C. korshinskii | 5 m × 3 m 1 m × 5 m | 667 2000 | |
| Slope bottom | C. korshinskii | 1 m × 5 m | 2000 | |
| P-3 | Slope top | P. tabuliformis | 3 m × 3 m | 1111 |
| Slope middle | C. korshinskii | 1 m × 3 m | 3333 | |
| Slope bottom | P. tabuliformis | 3 m × 3 m | 1111 | |
| P-4 | Slope top | P. tabuliformis | 5 m × 5 m | 400 |
| Slope middle | P. tabuliformis C. korshinskii | 5 m × 10 m 1 m × 10 m | 200 1000 | |
| Slope bottom | P. tabuliformis C. korshinskii | 5 m × 5 m 1 m × 5 m | 400 2000 |
| Pattern | Fit Line | R2 | Intersection Coordinates | Euclidean Square Distance | Judgment Results |
|---|---|---|---|---|---|
| P-1 | y = −0.0055x2 + 0.9421x − 58.8068 | 0.97 | (22.65, 77.35) | 14.05 | Stabilize |
| P-2 | y = −0.00943x2 + 1.3931x − 50.5464 | 0.93 | (22.69, 77.31) | 14.52 | Stabilize |
| P-3 | y = −0.00922x2 + 1.3884x − 49.7916 | 0.94 | (24.08, 75.92) | 25.94 | Instabilize |
| P-4 | y = −0.0102x2 + 1.5875x − 40.7730 | 0.99 | (25.44, 74.56) | 59.21 | Instability |
| Pattern | Thickness | Max. Water Retention | Effective Retention Rate | Standing Volume | Max. Water Capacity | Effective Storage Capacity | Evaluate | Sort |
|---|---|---|---|---|---|---|---|---|
| P-1 | 0.00 | 0.02 | 0.14 | 0.00 | 0.00 | 0.00 | 0.16 | 1 |
| P-2 | 0.17 | 0.00 | 0.00 | 0.33 | 0.25 | 0.07 | 0.82 | 2 |
| P-3 | 0.25 | 0.02 | 0.01 | 0.28 | 0.35 | 0.25 | 1.16 | 3 |
| P-4 | 0.21 | 0.15 | 0.14 | 0.23 | 0.21 | 0.17 | 1.11 | 4· |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Feng, S.; Li, W.; Gao, Y. Study on the Effect and Enhancement of Near-Natural Integrated Plant Positioning Configuration in the Hilly Gully Region, China. Forests 2024, 15, 841. https://doi.org/10.3390/f15050841
Zhao H, Feng S, Li W, Gao Y. Study on the Effect and Enhancement of Near-Natural Integrated Plant Positioning Configuration in the Hilly Gully Region, China. Forests. 2024; 15(5):841. https://doi.org/10.3390/f15050841
Chicago/Turabian StyleZhao, Hongsheng, Shuang Feng, Wanjiao Li, and Yong Gao. 2024. "Study on the Effect and Enhancement of Near-Natural Integrated Plant Positioning Configuration in the Hilly Gully Region, China" Forests 15, no. 5: 841. https://doi.org/10.3390/f15050841
APA StyleZhao, H., Feng, S., Li, W., & Gao, Y. (2024). Study on the Effect and Enhancement of Near-Natural Integrated Plant Positioning Configuration in the Hilly Gully Region, China. Forests, 15(5), 841. https://doi.org/10.3390/f15050841