Abstract
Transcription factors (TFs) regulate gene expression by binding to specific sequences on DNA through their DNA-binding domain (DBD), a universal process. This update conveys information about the diverse roles of TFs, focusing on the NACs (NAM-ATAF-CUC), in regulating target-gene expression and influencing various aspects of plant biology. NAC TFs appeared before the emergence of land plants. The NAC family constitutes a diverse group of plant-specific TFs found in mosses, conifers, monocots, and eudicots. This update discusses the evolutionary origins of plant NAC genes/proteins from green algae to their crucial roles in plant development and stress response across various plant species. From mosses and lycophytes to various angiosperms, the number of NAC proteins increases significantly, suggesting a gradual evolution from basal streptophytic green algae. NAC TFs play a critical role in enhancing abiotic stress tolerance, with their function conserved in angiosperms. Furthermore, the modular organization of NACs, their dimeric function, and their localization within cellular compartments contribute to their functional versatility and complexity. While most NAC TFs are nuclear-localized and active, a subset is found in other cellular compartments, indicating inactive forms until specific cues trigger their translocation to the nucleus. Additionally, it highlights their involvement in endoplasmic reticulum (ER) stress-induced programmed cell death (PCD) by activating the vacuolar processing enzyme (VPE) gene. Moreover, this update provides a comprehensive overview of the diverse roles of NAC TFs in plants, including their participation in ER stress responses, leaf senescence (LS), and growth and development. Notably, NACs exhibit correlations with various phytohormones (i.e., ABA, GAs, CK, IAA, JA, and SA), and several NAC genes are inducible by them, influencing a broad spectrum of biological processes. The study of the spatiotemporal expression patterns provides insights into when and where specific NAC genes are active, shedding light on their metabolic contributions. Likewise, this review emphasizes the significance of NAC TFs in transcriptional modules, seed reserve accumulation, and regulation of seed dormancy and germination. Overall, it effectively communicates the intricate and essential functions of NAC TFs in plant biology. Finally, from an evolutionary standpoint, a phylogenetic analysis suggests that it is highly probable that the WRKY family is evolutionarily older than the NAC family.
1. Introduction
The regulation of gene expression is a pivotal mechanism that enables plants to systematically organize their growth and development. This intricate process is governed by two essential components: trans-acting factors and cis-acting elements [1,2]. Thus, the cis-acting element in the promoter region regulates the precise initiation of gene transcription and transcriptional efficiency through the binding of transcription factors (TFs), likely determining the core region of transcriptional activation [3,4,5]. The transcriptional regulation of gene expression is directed by the action of specific proteins known as TFs, which interact with the cis-acting elements of key target genes to modulate determined signaling pathways [6]. TFs are fundamental controllers of cellular regulation, functioning in a complex and combinatorial manner, often grouping into modules. TFs bind DNA by recognizing specific sequence motifs, known as TF binding sites (TFBS), located in the promoters of target genes, thereby differentially regulating their effectiveness [7]. Consequently, the identification of TFBS holds immense importance in unraveling gene regulation during the growth and development of a specific species. TFs play a crucial role in regulating gene expression in both animals and plants, directly influencing almost all biological processes [8]. These proteins direct temporal and spatial expressions necessary for normal development and an adequate response to physiological or environmental stimuli. For a protein to be considered a typical TF, it must possess three main properties: nuclear localization, transactivation activity, and DNA-binding activity, enabling transcriptional modification. In summary, transcriptional regulation is the most extensively studied mechanism of gene regulation [8,9].
In addition to yeast one- (Y1H) and two-hybrid (Y2H) techniques, chromatin immunoprecipitation (ChIP) is a robust methodology for evaluating in vivo interactions of proteins with specific regions of genomic DNA under physiological conditions and estimating the density of TF binding at specific loci [10,11,12]. In addition to the previously mentioned techniques, the RNA-sequencing (RNA-Seq) platform [13] is widely employed to measure the transcript levels of TFs and obtain information about genome-wide binding sites. The outcome of DNA-TF binding is the activation or repression of target gene transcription. The status of chromatin is crucial for the function of TFs. Therefore, the use of ATAC-seq, a method for assessing chromatin accessibility across the genome, is recommended. Interestingly, despite TFs representing only a small fraction of plant transcriptomes (e.g., approximately 10% in Arabidopsis), they exert control over the global gene expression and regulation of a specific organism. Approximately 2296 TFs were cataloged in the PlantTFDB database and categorized into 58 families [14]. Notable among these families are DOF (DNA binding with one finger) [15], WRKY (named for its WRKYGQK sequence at their N-terminus) [16], MYB/MYC (bHLH) [17], NAC (NAM-ATAF-CUC), bZIP (basic region/leucine zipper) [18], HSF (heat shock factors) [19], DREB (dehydration-responsive element binding) [20], and AP2-EREBP (APETALA 2/ethylene (ET) response element binding protein) [21]. Other families are classified based on DNA structural domains.
NAC TFs are present in higher plants, as well as in a few species of algae. The NAC family, discovered by Souer et al. (1996) [22], stands out as one of the largest plant TF families. Land plants evolved from an ancestral charophycean alga, inheriting developmental, biochemical, and cell biological traits [23]. The NAC family is present in both aquatic green algae and higher terrestrial plants. In other words, NACs evolved from streptophyte green algae to higher plants, coinciding with the transition from aquatic to terrestrial living [24]. The moss Physcomitrella patens (a bryophyte) possesses 35 genes encoding putative NAC TFs [25] and 38 WRKYs [26]. Notably, the NAC family is absent in unicellular green algae and sparsely exists in mosses, such as P. patens. Most NAC genes have been reported to function as positive stress response TF. The abbreviation NAC is derived from the first three reported genes of this family: (i) NO APICAL MERISTEM (NAM), the first NAC gene discovered in 1996 from Petunia hybrida; (ii) TRANSCRIPTION ACTIVATION FACTOR 1 and 2 (ATAF1/ATAF2) from A. thaliana; and (iii) CUP-SHAPED COTYLEDON 2 (CUC2) from A. thaliana, where the NAC domain was first reported [27,28]. CUP-SHAPED COTYLEDON 2 (CUC2) was demonstrated to exhibit sequence homology and functional similarity to NAM in Petunia hybrida and displayed partial functional redundancy with CUC1. These three aforementioned genes encode protein sequences with a highly conserved amino acid sequence at the N-terminus. ATAF1/2, on the other hand, was found to be involved in stress responses and senescence regulation in plants [29]. NAM, ATAF1/ATAF2, and CUC2 possess a similar DNA-binding domain. In Arabidopsis, it has been reported that the NAC binding site contains the consensus DNA sequence (CGT[GA]), to which other relatively distant NAC TFs also bind [30]. However, the consensus DNA sequence in soybean is C[AC]C[GA][TC][GA]CC [31].
This update compiles recent data on the significant role of the NAC family in crucial processes such as stress, leaf senescence (LS), and seed life, across model species (e.g., Arabidopsis) and agronomically important species (e.g., rice). Particularly noteworthy is the intervention of NAC TFs in seed dormancy, germination, and reserve accumulation (e.g., starch). Additionally, considerable attention is devoted to exploring the hormonal regulation of the spatial and temporal expression of these TFs, given their modular nature and complexity. Furthermore, to provide an evolutionary framework for such diversity, based on already published sequences, this review presents a detailed phylogenetic study of the NAC and WRKY families, revealing a possible evolutionary origin for the NAC gene family. Finally, the study synthesizes the most significant conclusions from a series of investigations conducted in the last three years.
2. Unraveling the Complex Molecular NAC Structure
NAC proteins are abundant in terrestrial plants, exhibiting wide distribution. Notably, NAC TFs play a role in the transition of both aquatic-to-terrestrial plants and vegetative-to-reproductive growth [4,24]. This underscores the clear involvement of NACs in plant evolution. Mosses and lycophytes, representing early-diverged land plants, are predicted to have fewer NAC proteins (≤30), indicating that the expansion of NAC proteins occurred after the evolution of vascular plants. Vascular plants, in contrast, possess over 100 copies in various angiosperm species [32]. From an evolutionary perspective, NACs transitioned from algae to land organisms and subsequently expanded throughout land plants. The number of proteins increased from 20–30 in mosses and lycophytes to over 100 copies in various higher plant species [33]. This expansion is evident even in several groups of streptophytic green algae, considered to be the sister group to land plants. It has been hypothesized that the NAC family dates back more than 400 million years [24,32]. In summary, the NAC family exhibits a gradual increase in size from basal streptophytic green algae to higher angiosperms. Additionally, the plant TF database includes a total of 19,997 NAC TFs from 150 species. Among these, 328 are distributed in rice, 280 in tobacco, 138 in A. thaliana, and 101 in tomato (http://planttfdb.gao-lab.org; [34]). The abundance of NAC proteins in these species underscores the vital role of NACs in plant evolution [35]. Furthermore, NAC proteins exhibit a modular organization [36]. Most NACs contain a highly conserved N-terminal DNA-binding domain (the NAC domain responsible for oligomerization, typically consisting of about 150–160 amino acid residues) and a variable C-terminal domain for regulating gene expression (a relatively divergent transcriptional regulatory domain; TR) [29,37,38]. Over 10% of the NAC TFs in Arabidopsis contain an α-helical transmembrane motif within their C-terminal domain. This domain typically functions as a transcriptional activator or repressor [39,40]. Notably, NAC18 possesses both activator and repressor domains [41]. Liang et al. (2014) conducted a comprehensive analysis of the transcriptional activator OsNAP, determining that its C-terminal region (amino acids 181–392) exhibits high transcriptional activation activity. In contrast, the N-terminal region of the NAC domain (amino acids 1–190) showed no activity [42]. TaNAC019-A1 possesses a very short C-terminal region, comprising fewer than 40 amino acids, and functions as a transcriptional repressor [43]. In contrast, ZmNAC128 and ZmNAC130 in maize have a long C-terminal region associated with transcriptional activation activities [44]. The multiple localizations and translocation traits of NAC TFs implicate various biological functions and regulatory mechanisms [45,46]. Notably, the C-terminal domain of AtNAC2 exhibits transcriptional activation activity, whereas its N-terminal domain does not [47]. In summary, the NAC N-terminal region harbors the repression domain, while the C-terminal region largely functions as the transactivation region [48]. These properties enable NACs to exhibit multiple regulatory patterns at the transcriptional, post-transcriptional, and translational levels, serving as both activators and repressors in response to developmental and environmental cues. Studies have shown that interactions between TFs and cofactors can alter DNA-binding affinity and convert activators into repressors [49]. Similarly, TFs are enriched with intrinsically disordered regions that collaborate in DNA binding specificity. Interestingly, these regions regulate transcription through either an ABA-dependent or ABA-independent pathway, modulating stress-related gene expression in plants [50,51].
The NAC domain can be subdivided into A, B, C, D, and E subdomains (Figure 1). Subdomain A is implicated in both homo- and heterodimerization. For instance, the detection of yellow fluorescence resulting from the interaction between ONAC127-YFPN and ONAC129-YFPC confirms the formation of a heterodimer by ONAC127 and ONAC129 in the nucleus [52]. Subdomains C and D, characterized by positive charge and high conservation, constitute the DNA-binding sites. Lysine residues within subdomain D play indispensable roles in nuclear shuttling [29,39]. Divergent subdomains B and E contribute to the functional diversity of NAC proteins [29,53]. As a result, the NAC domain is responsible for binding properties in the N-terminus, playing pivotal roles in plant–pathogen interaction, stress resistance [54], and various other key physiological functions. Concurrently, NACs often function in a dimeric form, with the dimerization site located within the NAC domain. Dimerization is essential for stable DNA binding [55] and is established by amino acid residues Leu14-Thr23 and Glu26-Tyr31. For instance, AtNAC2 functions as a dimer, predominantly expressed in roots and moderately in flowers [47]. Intriguingly, the first structure determined by X-ray diffraction of a NAC domain protein was the crystal structure of the NAC domain from A. thaliana (i.e., ANAC019) [36]. The crystal structure of the NAC rice domain has also been reported [56]. The NAC domain monomer consists of a twisted antiparallel β-sheet surrounded by two helices, with this β-sheet playing a crucial role in sequence-specific DNA recognition [39].
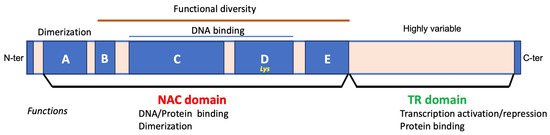
Figure 1.
TFs exhibit characteristic domains responsible for various functions, including DNA binding, oligomerization/protein–protein interactions, transcriptional regulation, and nuclear localization. The accompanying figure illustrates the structure of a NAC-TF. The NAC domain, enclosed by a red ellipse, comprises nearly 150 amino acid residues and often includes a nuclear localization signal, enabling protein binding. Subdomain A facilitates protein dimerization, subdomains B and E contribute to functional diversity, while subdomains C and D, which are positively charged and highly conserved, are responsible for DNA binding. The C-terminal transcriptional regulatory (TR) region, surrounded by a green ellipse, functions as a transcriptional activator or repressor and may possess protein binding activity, interacting with other TFs. Adapted from Singh [33] and Diao [57].
Most NAC TFs are localized in the nucleus, representing their active forms, where they bind DNA and carry out specific transcriptional roles [4,48,58,59,60,61]. However, a subset of NACs is found in other cellular compartments such as the plasma membrane (PM), cytoplasm, or endoplasmic reticulum (ER), indicating inactive forms (i.e., dormant state) (Figure 2). Membrane-bound NAC TFs can be identified by the presence of a distinctive transmembrane domain, directing their association with the ER [45,62,63,64,65] or PM [30,63,66,67,68,69]. A considerable number of NAC TFs in Arabidopsis possess transmembrane domains, anchoring them to membranes [45,64]. For instance, it has been demonstrated that the full-length ANAC060 protein, containing a transmembrane domain, associates with the nuclear membrane. Conversely, its truncated form lacking the transmembrane domain is localized in the nucleus [70]. In contrast, full-length ANAC040 and ANAC089 proteins are predominantly detected on PM or ER membranes, while their truncated forms lacking the transmembrane domain are found in the nucleus [66,67,71,72]. Notably, NAC103 does not have a predicted transmembrane domain [66]. After exposure to specific developmental or environmental cues, membrane-bound TFs undergo proteolytic processing and are transported to the nucleus, where they exert their regulatory function [73]. In other words, membrane-tethered proteins exhibit extranuclear localization, requiring detachment after specific signals reach the nucleus and regulate gene expression. A recent study focused on an attractive ER membrane-tethered NAC TF (MTTF) from oilseed rape (Brassica napus L.) [71]. This update highlights the mechanism of MTTF nuclear import as well as its functions in regulating plant growth and stress response. Significantly, the absence of the membrane-anchoring domain in the Cvi FSQ6/ANAC089 protein resulted in nuclear localization [74]. Notably, nuclear localization signals (NLSs) have been experimentally identified in NAC proteins. The NLSs, rich in positively charged amino acids, are predominantly located toward the N-terminal end in the majority of NAC proteins [75]. This work represents the first report on the presence of unique NLSs in both the N- and C-terminal regions of NAC TFs. To summarize, NLSs are mainly situated toward the N-terminal end in the majority of NAC proteins. Collectively, at least 8.60% of NAC TFs possess a transmembrane domain at both the N- and C-terminal ends. In most cases, the transmembrane domain is located toward the C-terminal end. Current knowledge also suggests that NAC TFs play a vital role in stress resistance in ER.
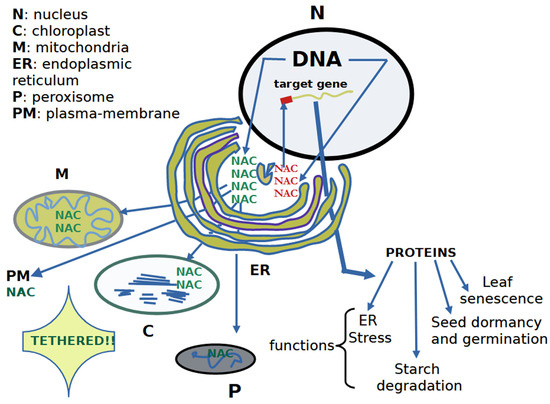
Figure 2.
The cellular synthesis of NACs generates a population of these TFs in the ER. The nuclear genes responsible are affected by phytohormones. A notable portion of the NAC population is sent to the nucleus to alter the transcription of target genes, producing the corresponding proteins. These proteins alter a series of physiological processes such as LS, seed reserve degradation (e.g., starch), ER stress, and seed dormancy and germination, among others. The remaining endoplasmic population of NACs (membrane-tethered NAC TFs) is sent to cellular compartments (i.e., chloroplasts, peroxisomes, and mitochondria) and plasma membranes. The membrane-tethered subset is a small family specific to plants, which lose their transmembrane domain and are then sent to the nucleus to exert their physiological role once bound to the corresponding target genes. The exit of membrane-tethered NAC TF from the corresponding cellular compartment occurs in response to environmental and developmental changes. For further information, see [71,76,77].
Through comprehensive analysis using relevant databases and software tools, a recent detailed study on Dendrobium nobile revealed the localization of DnoNACs as follows: 70 members in the nucleus, 6 in the chloroplast (DnoNAC11, DnoNAC12, DnoNAC23, DnoNAC30, DnoNAC69, and DnoNAC80), 3 in the cytoplasm (DnoNAC26, DnoNAC74, and DnoNAC78), 3 in the mitochondria (DnoNAC55, DnoNAC76, and DnoNAC83), and 3 in the peroxisome (DnoNAC16, DnoNAC17, and DnoNAC43) [78]. These locations align with those found for the Passiflora edulis NAC TF family [69]. Notably, ONAC127 and ONAC129 proteins were observed in the nucleus, cytoplasm, and membrane of rice seeds [52]. In conclusion, research on NAC TFs has underscored the significance of this protein family in plant biology. Moreover, the number of NAC TFs per genome, along with their structural and functional properties, tends to increase with the complexity of the organism. In essence, the evolution of NAC TFs is strongly linked to the adaptation of plant life from aquatic to terrestrial forms. Recent studies have unveiled that B. napus, with 410 encoded NAC TFs, possesses the highest number, while Klebsormidium flaccidum algae, with 3 encoded NAC TFs, has the lowest [75]. Genetic studies suggest that NAC TFs in all species underwent duplication, with no loss of any of these TFs. This observation suggests that NAC TFs evolved from common ancestors through orthology and experienced numerous duplication events during divergence and speciation, indicating paralogy.
3. Relevant Physiological Functions of NACs
NAC proteins constitute one of the largest plant-specific TF families, renowned for their pivotal roles in various metabolic pathways during plant growth, development, and stress processes [33,34,76]. Among their diverse functions, NAC proteins are key players in processes such as LS and programmed cell death (PCD) across several studied species, including Arabidopsis, rice, tomato, wheat, oilseed rape, and cabbage [33,42,79,80,81,82,83,84,85,86,87,88,89]. They are also involved in cell wall (CW) metabolism [33,48,90,91,92,93,94,95,96], reactive oxygen species (ROS) production [33,77,96,97,98], nutrient remobilization [44,99], xylem/phloem differentiation and formation [88,100,101,102], regulation of plant immunity [54], and hormone signal transduction (e.g., salicylic acid (SA), jasmonic acid (JA), ethylene (ET), and abscisic acid (ABA)) [33,34]. Furthermore, NAC proteins respond to signals such as ROS, contributing to plants’ resistance against pathogens [60,87,103]. Their involvement extends to seed life (to be further discussed) and the regulation of biotic and abiotic stresses, including drought, heat stress, low-O2 (anoxia), and cold tolerance [33,52,57,61,104,105,106,107,108]. As demonstrated in this update, NAC TFs emerge as dominant regulators in response to abiotic stresses. Surprisingly, the role of NAC TFs in regulating plant functions under non-stress conditions is less explored. Plants frequently encounter combined stresses rather than single stressors, leading to more severe disruptions. However, the underlying mechanisms of these combined stresses remain unknown. Notably, IbNAC3 from sweet potato (Ipomoea batatas) has been found to confer tolerance to both single and combined salt and drought stresses in Arabidopsis [61]. Interestingly, ANAC102 is essential for tolerance to stress induced by low-O2 concentrations (i.e., 0.1% O2 stimulates ANAC102 expression) during the seed stage but not in early adult plants. In other words, although ANAC102 is not crucial for adult tolerance to low-O2, it plays a significant role in tolerating low-O2 levels during germination [104]. In the complicated regulatory network governing the CW, NAC29 and NAC31 play crucial roles by influencing downstream cellulose synthase (CESA) activity. These two TFs exert their regulatory effects by activating the MYB61 TF. The activation of MYB61, in turn, serves as a crucial mechanism for controlling the synthesis of secondary CW [109].
3.1. NAC TFs Play Crucial Roles in Both ER Stress Responses and Leaf Senescence
The accumulation of unfolded or misfolded proteins can disrupt ER protein homeostasis, resulting in ER stress and compromising cellular function. In response, the unfolded protein response pathway is activated to mitigate ER stress. Additionally, ER stress is known to directly generate ROS signaling and modify the redox status. NAC TFs have been identified as key players in the response to ER stress. Under these conditions, the Arabidopsis NAC103 transcriptional activator [66], along with the membrane-associated NAC089 and NAC062, are all upregulated. NAC103 forms homodimers in both yeast and plants and is induced by bZIP60 [110]. It plays a crucial role in the expression of genes related to DNA damage response, contributing to stabilization under unfolded protein stress [110,111]. The overexpression of NAC103 leads to a significant increase in the transcription of ER stress-responsive genes [110]. Meanwhile, AtNAC062, a plasmalemma-associated TF, plays a vital role in promoting cell survival under ER stress conditions. NAC062 undergoes relocation from the plasmalemma to the nucleus, where it regulates the expression of ER stress-responsive genes [71,72]. On the other hand, NAC089 relocates from the ER membrane to the nucleus, inducing programmed cell death (PCD) in response to ER stress treatment [71,72]. Recently, AtNAC091 was identified as a crucial unfolded protein response mediator, and its expression was demonstrated to be induced by ER stress. This induction is primarily dependent on the ER stress regulators bZIP60 and bZIP28 [112]. In contrast, the Glycine max transcriptional repressor GmNAC81 is positively regulated during LS. Notably, the GmNAC81-mediated response and senescence-induced response share similar mechanisms. As a consequence, overexpression of GmNAC81 in the early vegetative stage accelerates LS and increases sensitivity to drought [113]. Furthermore, GmNAC30 and GmNAC81 (formerly GmNAC6) have been demonstrated to participate in ER stress [71,79,113]. Both TFs collaborate to activate the caspase-1-like vacuolar processing enzyme (VPE) gene, playing a crucial role in the stress-induced PCD response in plants [114]. Interestingly, GmNAC30 forms a heterodimeric interaction with GmNAC81 in the nucleus of plant transfected cells, suggesting the potential biological significance of this interaction. Mendes’s group has demonstrated several roles for both GmNAC81 and GmNAC30: (i) both are induced by ER stress with similar kinetics; (ii) they are upregulated by the cell death-inducer cycloheximide and promote cell death when transiently expressed in soybean protoplasts; (iii) both TFs bind in vivo to common target promoters and regulate their expression in a coordinated manner; (iv) the VPE promoter contains a GmNAC81/GmNAC30 binding site; and (v) GmNAC81 and GmNAC30 specifically bind to the core consensus sequence TGTGTT [115]. In summary, GmNAC30 and GmNAC81 work together to regulate VPE expression, a plant-specific executor of cell death. VPE is likely responsible for the execution of the cell death program induced by ER and osmotic stress.
The LS is a genetically and epigenetically programmed process that can be induced by internal and adverse environmental factors and marks the final stage of leaf development [42]. In Glycine max, approximately 40% of the NAC genes are upregulated in LS, and only GmNAC030, GmNAC065, and GmNAC081, have been functionally characterized. In Arabidopsis, several NAC TFs have been identified as specific regulators of LS. Notably, ATNAP, ANAC029, ANAC092 (ORE1), ANAC059 (ORS1), ANACO42 (JUB1), and ANACO83 (VN12) play prominent roles in the regulatory network governing LS. The overexpression of AtNAP, ANACO59, and ANACO92 induces precocious senescence, while blocking the function of these three TFs significantly delays LS [42]. ANACO92 (ORE1), identified as a positive regulator of LS, plays a crucial role in controlling the aging process in A. thaliana. The overexpression of ORE1 leads to early senescence, whereas its inhibition retards LS [116,117]. Indeed, the ore1-2 and ore1-6 lines exhibit delayed LS. The interaction between ET and ORE1 in LS has been demonstrated in several manuscripts [118,119]. The increased expression of EIN2 directly upregulates the expression of ORE1 [119]. In contrast, the expression of ORE1 is negatively regulated at the post-transcriptional level [117]. These authors have provided evidence of ORE1 binding in vivo to the promoters of two other senescence-associated genes (SAG29/SWEET15 and SINA1). Additional manuscripts have highlighted the direct binding of ORE1 to PRR9, SAG29, and BFN1, among other senescence-associated genes, to regulate its expression [83,118,119]. In a recent study, a mechanism modulating ore1 in G. max L. has been proposed [120]. This mutant demonstrates high tolerance to oxidative stress and significantly delays LS in A. thaliana [115]. It has been proposed that upon introducing At-ore1 into G. max, At-ore1 exhibits an inverse role in LS depending on the ABA/IAA balance. This effect is possibly mediated through an increase in GmNAC081- or GmNAC065-mediated H2O2 regulation [121]. Recently, it was found that GmNAC81 overexpression in the early vegetative stage accelerated LS and increased sensitivity to drought. In addition, it was suggested that GmNAC81 may negatively control drought tolerance not only via VPE activation but also through the suppression of ABA signaling, and that GmNAC81 regulated the expression of six target genes (e.g., KIT1 and STI were highly repressed by GmNAC81 overexpression) involved in LS [113]. Collectively, data from [113] propose that GmNAC81 directly modulates drought tolerance by controlling critical regulators of ABA signaling. In other words, GmNAC81 may negatively regulate drought tolerance not only through VPE activation but also through the suppression of ABA signaling. In summary, current knowledge suggests that NAC TFs serve as positive or negative regulators of LS by either activating or repressing RBOH-mediated ROS production. For instance, BrNAC055 has been shown to activate the in vivo transcription of BrRbohB, BrRbohC-like, BrNYC1, and BrNYE1, indicating its role in regulating LS through ROS-related mechanisms. These findings provide evidence to support that BrNAC055 works as an activator of BrRbohB, BrRbohC-like, BrNYC1, and BrNYE1 transcription through direct binding to their promoters.
3.2. Functions of OsNAC2 and Other Members of OsNAC Family in Rice
Rice, being a staple food consumed by half of the human population, warrants comprehensive study at various molecular levels. The rice genome is predicted to contain 151 NAC genes. Among the transcription factors, OsNAC2, considered the ortholog of AtORE1 (Arabidopsis NAC092), stands out as one of the most extensively studied in rice. OsNAC2 plays a crucial role in modulating plant height and architecture by participating in GAs signaling pathways [122]. Additionally, it contributes to LS through ABA biosynthesis [123], enhances abiotic stress tolerance [11], is involved in PCD [124], and has recently been implicated in the release of seed dormancy and germination [11]. Overexpressing OsNAC2 in plants enhances stress resistance to cold, drought, and salt, while transgenic rice with inhibited OsNAC2 expression shows reduced salt tolerance [125]. Moreover, OsNAC2 has been demonstrated to play a role in regulating root development by integrating auxin and cytokinin (CK) signaling pathways [126]. Additionally, OsNAC45 is implicated in ABA response and contributes to salt tolerance [95]. Rice plants overexpressing OsNAC2 exhibit increased transcription of ABA biosynthesis genes, namely OsNCED3 and OsZEP1, with OsNAC2 binding to their promoters [123]. Interestingly, OsNAC2 is upregulated in response to low ABA levels and downregulated under high ABA levels, suggesting a feedback repression mechanism for OsNAC2. In other words, (i) OsNAC2 functions as a repressor of yield, and flowering time in rice [123]. Additionally, OsNAC2 functions at the intersection of ABA and abiotic stress pathways; (ii) the role of OsNAC2 contrasts with other reported NAC members such as OsNAC5, OsNAC6, and OsNAC10, where their overexpression has been shown to enhance drought stress resistance and grain yield [125]; (iii) overexpression of OsNAC2 suppresses salt resistance in transgenic rice plants, increasing sensitivity to high salt stress. Additionally, alterations in OsNAC2 expression impact drought sensitivity during the vegetative state; (iv) OsNAC2 has the ability to directly bind to the promoter of LATE-EMBRYOGENESIS-ABUNDANT-3 (OsLEA3) and downregulate its expression; and (v) OsNAC2 exhibits elevated expression in OsEIN2 overexpressing lines, whereas its expression is downregulated in ein2 mutants. OsLEA3, a well-known stress marker gene, has been shown to confer stress resistance when overexpressed [125]. OsNAC2 expression is significantly induced by ABA and osmotic stress. Therefore, OsNAC2 functions through the ABA-dependent pathway and may downregulate numerous ABA-responsive stress marker genes. It is plausible that OsNAC2 regulates both abiotic stress responses and ABA-mediated responses, influencing the intersection between the ABA and abiotic stress pathways. Additionally, the root-specific overexpression of SNAC1, OsNAC5, OsNAC6, OsNAC9, and OsNAC10 remarkably enhances drought resistance in rice plants at the reproductive stage. In particular, OsNAC9 modifies root architecture, extending aerenchyma and stele, contributing to drought tolerance and enhanced grain yield under field conditions [127]. Similarly, OsNAC10 enhances stele, cortex, and epidermis size, improving drought tolerance and grain yield in field drought conditions [128]. Lastly, OsNAC011 serves as a promoter of LS, as plants overexpressing OsNAC011 exhibit a precocious senescence phenotype, while knockdown plants display reduced LS with a high accumulation of chlorophyll [119,123].
3.3. Additional Roles of NACs Genes in Other Eudicot Species
Recent findings related to BnaNAC60 in B. napus indicate its involvement in various processes associated with LS and stress responses. BnaNAC60 expression is induced during the progression of LS in B. napus, suggesting its involvement in the regulation of senescence-related processes [98]. BnaNAC60 is a membrane-tethered protein, indicating its initial association with cellular membranes. However, it undergoes translocation to the nucleus in response to both ER stress and oxidative stress treatments [98]. BnaNAC60 acts as an upstream regulator of cell death, LS, and ROS accumulation. It likely influences these processes by activating the promoter activity of genes involved in ROS generation (such as BnaRbohD) and ROS signaling (such as BnaZAT12) [77]. These findings suggest that BnaNAC60 is a multifaceted regulator involved in coordinating responses to stress conditions and senescence in B. napus. The activation of genes related to ROS suggests a potential link between BnaNAC60 and redox signaling, which is often crucial in stress responses and senescence. Recently, it was demonstrated that the CmNACP1-mRNA of Cucumis melo moves through the phloem, indicating its capacity for long-distance transport. and the little-characterized AtNAC087 of A. thaliana is expressed in the phloem, apex, and developing flowers. In contrast to CmNACP1-mRNA, AtNAC087-mRNA is not mobile, suggesting a cell-autonomous function affecting the cells where it is expressed without being transported over long distances. Given that CmNACP1 and AtNAC087 are orthologs (i.e., a common evolutionary origin), this feature was used to investigate the cellular role of the AtNAC087 gene [129]. The overexpression of AtNAC087 leads to the accelerated LS of aerial rosettes, confirming its function as a positive regulator of LS in a tissue-dependent manner. Specifically, AtNAC087 orchestrates post-mortem chromatin degradation specifically in roots via BFN1 nuclease, a senescence-associated type I nuclease. Interestingly, the tissue-dependent nature of its regulatory functions implies that AtNAC087 may have distinct roles in different plant tissues, contributing to the complexity of senescence-related processes. Ectopic expression of AtNAC087 correlates with the emergence of aerial rosettes, likely resulting from the induction of HUA2, a gene involved in flower induction [129]. Additionally, the gaseous hormone ET regulates various aspects of plant growth and development, with fruit ripening being the most well-known, along with responses to biotic and abiotic factors [130,131]. Numerous studies have indicated that NAC TFs play a significant regulatory role in fruit development and ripening, particularly in relation to ET signaling [132,133,134,135]. The recent identification of two NAC TFs, PpNAC1 and PpNAC5, in Prunus represents a significant advance in understanding their role in fruit development and ripening. Orthologs PpNAC1 and PpNAC5 activate the transcription of genes associated with CW degradation, cell elongation, and ET biosynthesis. Interestingly, the authors suggest a critical regulatory role for both genes in the ripening of peach fruit-producing plants but not in fruitless ones [136]. On the other hand, Peng et al. (2023) [137] obtained these attractive results in C. maxima (pumpkin): (i) CmoNCED6 is identified as the key gene for ABA synthesis, and CmoNAC1 is most likely an important TF regulating H2O2 and ABA signaling in pumpkin rootstock under salt stress; (ii) CmoNAC1 influences the production of H2O2 and ABA by regulating the expressions of CsaRBOHD and CsaNCED3 in grafted cucumber leaves, as well as CmoRBOHD1 and CmoNCED6 in roots. Similar results were also obtained in tomatoes [23,47], cowpeas [35], soybeans [48], and Arabidopsis [37]; (iii) CmoNAC1 binds to the promoters of CmoRBOHD1, CmoNCED6, CmoAKT1;2, and CoHKT1;1 in response to salt stress in grafted cucumbers. In summary, CmoNAC1 is not only a key TF positively regulating salt tolerance in grafted cucumbers but also promotes the synthesis of H2O2 and ABA signals in roots, contributing to an increased K+/Na+ ratio. This suggests a role for CmoNAC1 in maintaining ion homeostasis under salt stress conditions [137].
3.4. Recent Novelties in Stress Response by NACs Genes
As evidenced throughout this review, NAC TFs play essential roles in stress responses. However, there is limited information available on stress-related NAC genes in maize, despite the identification of a total of 190 predicted NAC members. With respect to salt tolerance, recent research in Zea mays demonstrated the involvement of the ZmNAC89 gene in stress caused by Na2CO3 [138]. Here are some key properties of ZmNAC89: (i) it possesses transcriptional activation activity and is a nuclear protein; (ii) it exhibits outstanding function against salt-alkali tolerance, with overexpression of ZmNAC89 improving both stresses in A. thaliana; and (iii) its expression is induced by ABA, being upregulated under ABA stress [138]. These findings collectively highlight ZmNAC89 as a stress-responsive TF with promising attributes for enhancing salt and alkali tolerance in plants. Previously, functions of other maize NAC genes have been identified. For instance, ZmSNAC1, ZmNAC33, and ZmNAC55 significantly enhance drought tolerance in transgenic Arabidopsis, while ZmNAC111 is associated with drought resistance [139,140,141]. Similarly, ectopic expression of CmNAC1 enhances the tolerance of transgenic A. thaliana to cold, salt, and drought stresses [142]. The overexpression of VvNAC17 from grape in Arabidopsis, leading to increased sensitivity to ABA and enhanced tolerance to salt, freezing, and drought, underscores the multifaceted roles that NAC TFs can play in plant responses to environmental stresses [143]. It is noteworthy that elevating ZmNAC111 expression in transgenic Z. mays enhances water use efficiency at the seedling stage [144]. On the other hand, ZmNAC33 is strongly induced by ABA and acts as a positive regulator in drought tolerance [141]. This suggests that ZmNAC33 may be part of the ABA-mediated signaling pathway, contributing to the plant’s ability to cope with drought stress. Interestingly, several stress-related cis-acting elements exist in the promoter region of the ZmNAC55 gene, which is induced by ABA, high salinity, and cold stresses. Similarly, overexpression of ZmNAC55 in Arabidopsis led to hypersensitivity to ABA during seed germination [140]. In summary, ZmNAC55 appears to be a stress-responsive NAC gene with a role in ABA signaling. Its induction under various stress conditions and its impact on seed germination highlight its potential importance in the plant’s adaptation to environmental challenges, particularly those related to ABA-mediated responses. The results obtained with ZmNAC55 open the possibility of feasible investigations in the transgenic breeding of drought-tolerant crops. However, ZmSNAC1 was strongly induced by low temperature, NaCl, drought stress, and exogenous ABA, while being downregulated by SA. Overexpression of ZmSNAC1 in A. thaliana led to hypersensitivity to ABA and osmotic stress, conferring tolerance to dehydration without any pleiotropic effects [139]. Finally, it is noteworthy that overexpression of ZmNAC84 in tobacco improved drought tolerance, while ZmNAC88 played a crucial role in ABA-induced antioxidant defense against drought stress [145]. Given its remarkable impact, the study of the ZmNAC84 function has recently advanced. Today, it is known that ZmNAC84 physically interacts with ZmCCaMK (Ca2+/calmodulin-dependent protein kinase) both in vitro and in vivo. Functional analysis has revealed that ZmNAC84 is essential for ABA-induced antioxidant defense in a ZmCCaMK-dependent manner. In other words, ABA-produced H2O2 first induces the expression of ZmCCaMK and ZmNAC84, then activates ZmCCaMK, which phosphorylates ZmNAC84 at Ser-113, thereby initiating antioxidant defense by activating downstream genes [145]. Much more recently, it was demonstrated in Z. mays that phosphorylated ZmNAC84 enhances drought tolerance by directly modulating the expression of ZmSOD2, a key player in the plant’s antioxidant defense against ROS generated during drought stress [146].
Three novel nuclear NAC genes from the important crop Triticum aestivum, TaNAC2, TaNAC29, and TaNAC67, were characterized for their enhanced multi-abiotic stress tolerance in transgenic plants. TaNAC29 expression is stimulated by H2O2, salt, and ABA and exhibits high levels in LS, indicating its involvement in the senescence process and response to salt and drought stresses [147]. Interestingly, introducing the rice stress-responsive NAC gene into wheat resulted in a transgenic plant with significantly enhanced tolerance to multiple abiotic stresses [148]. On the other hand, transgenic lines overexpressing TaNAC69 produce more biomass in the shoot and root under stress-inducing conditions, demonstrating increased resistance to salinity and drought stresses through enhanced scavenging of ROS [149]. Taken together, all the results discussed in Section 3, and others not included in it, demonstrate that a large number of NAC genes play a key role in enhancing abiotic stress tolerance, and their function is likely conserved in angiosperms. Moreover, these findings suggest that the use of transgenic plants appears to be an adequate approach for addressing stress tolerance in the near future through biotechnological methods [150]. Recently, 104 NAC genes were identified in Camellia sinensis, the tea plant [150]. Some of these genes possess one or two transmembrane domains at the C-terminus, while others have these domains at the N-terminus. The expression of most of them was induced by exogenous ABA and drought stress (i.e., PEG treatment). Interestingly, overexpression of CsNAC28 in Arabidopsis increased sensitivity to ABA and resulted in additional upregulation of drought-responsive genes. In summary, [150] provides demonstrative data that CsABF2 could bind to the ABRE cis-acting element in the promoter of CsNAC28 and activate CsNAC28 expression. Comparatively, CsNAC28 transgenic plants, when compared to the WT, exhibited significantly reduced ROS content and less oxidative damage under drought stress. In other words, the improved antioxidant capability of CsNAC28 transgenic plants enhanced drought resistance at the cellular level in transgenic Arabidopsis.
In gymnosperms, such as conifers, the molecular mechanisms that regulate multiple abiotic stress responses remain largely unnoticed. Recently, data have been obtained that point to a notable role of NAC TFs. Thus, it was demonstrated in Pinus tabuliformis under abiotic stresses that PtNAC3 is nuclear, and the activation of PtNAC3—a stress-related NAC gene—is mediated by ET rather than ABA [60]. In short, PtNAC3 and AtNAC3 possess identical amino acid N-terminal NAC domains and are highly upregulated by ET, exhibiting co-expression. PtNAC3 protein may directly target PtZFP30, a zinc finger protein involved in abiotic stresses, activating it. PtZFP30 is upregulated by ET and ACC, and is likely one of the downstream genes of PtNAC3 [60]. This recent work provides intriguing insights into the hub nodes of the abiotic stress response network in conifers. On the other hand, two NAC genes, namely PpNAC2 and PpNAC3 (which cluster with Arabidopsis ATAF1 and ATAF2), were characterized from Pinus pinaster (maritime pine). Their expression was strongly and rapidly induced by MeJA and high salinity. PpNAC2 and PpNAC3 promoters contain cis-elements involved in biotic and abiotic stress. Similarly, NbbHLH2 proteins can mediate the JA induction of the PpNAC3 promoter, suggesting the conservation of the JA signaling pathway between angiosperms and gymnosperms. In summary, the knowledge about NAC TFs in terrestrial plants leaves no doubt that their role has been strengthened throughout evolution, becoming particularly evident in the regulation of stress processes.
Finally, the development of bioinformatics and modern molecular biology techniques, coupled with the recognition that NAC genes play pivotal roles in various plant processes, has promoted extensive studies on the NAC gene family across numerous species (Table 1). This is the case with A. thaliana [151], rice [152], wheat [153], soybean [154], peanut [46], maize [155], potato [156], cassava [157], Chinese cabbage [158], pepper [159], melon [160], apple [161], sunflower [162], Saccharum spontaneum [163], Tartary buckwheat [141], Zanthoxylum bungeanum [164], Asparagus officinalis [165], Brassica juncea var. Tumida [166], Liriodendrum [4], Hibiscus hamabo Sieb [167], and other species (see Introduction from [168]). Likewise, accumulated studies have shown that there are currently 117 NAC genes in Arabidopsis [169], 163 in poplar [170], 74 in Vitis vinifera [171], 85 in Liriodendrum [4], 151 in rice [54], 110 in potato [156], 152 in tobacco [172], 152 in soybean [154], 97 in Medicago truncatula [173], 104 in tomato [53], 204 in Chinese cabbage [174], 152 in maize [155], 85 in Dendrobium nobile [78], 150 in Helianthus annus [46], 105 in passion fruit (Passiflora edulis) [69], and 123 in Pinus tabuliformis [60]. Similarly, the genome-wide identification of NAC genes has been extended to numerous plant species (see Introduction and Discussion in [175]).

Table 1.
Selected species in which NAC TFs have been compiled in this study.
4. Phytohormonal Requirements for NACs Function in Vegetative Organs and Leaf Senescence
As previously stated, NAC TFs play important roles in various biological processes and responses to abiotic stresses. Many of these tasks are regulated by hormones, and several NAC genes play critical roles in these processes [37,47,135]. This is so much so that various NAC genes are hormone-inducible [192,193,194]. To date, only a few NAC genes have been specifically shown to regulate LS. For example, ANAC029, ANAC042, ANAC059 (ORS1), ANAC083, and ANAC092 (ORE1) play significant regulatory roles in this aging process [129]. Overexpression of AtNAP, ORS1, and ORE1 triggers acceleration of LS, and blocking the function of these TFs significantly delays LS. Therefore, it has been suggested that AtNAP, ORE1, and ORS1 act as non-redundant positive regulators of LS in Arabidopsis [195]. The prematurely senile 1 (ps1-D), a dominant precocious LS mutant, was previously identified [42]. PS1 (formerly called OsNAP) encodes a plant-specific NAC and is considered an adequate marker for the onset of the senescence process in rice. One notable finding is the connection between OsNAP and ABA. A feedback repression mechanism has been identified where OsNAP negatively regulates ABA biosynthesis. This is evidenced by a significant reduction in ABA content in ps1-D mutants, which are associated with OsNAP. This feedback loop implies that OsNAP not only participates in the regulation of LS and endosperm maturation but also influences ABA levels, showing its multifaceted role in plant physiology. Accordingly, the transcription level of OsNAP is directly linked to the nutrient remobilization associated with senescence [42].
4.1. The Relationship of ABA and ET with NAC
The RD26 gene was the first NAC identified as a regulator of both ABA and JA signaling during stress responses in Arabidopsis [103]. On the other hand, it is well-known that ABA synthesis and signaling upregulate LS, a stress process in which several NAC TFs are involved. NAC/ATAF1 directly regulates the ABA biosynthetic gene NCED3 in A. thaliana [196], whereas OsNAP (NAC-LIKE, ACTIVATED BY AP3/PI) confers abiotic stress response exclusively through the ABA pathway in rice [182]. These findings indicate the intricate crosstalk between ABA and JA signaling pathways, with NAC TFs like RD26, NAC/ATAF1, and OsNAP playing roles in mediating stress responses and senescence through these pathways. The transcription of OsNAP increased rapidly, up to 102-fold after 12 h of ABA treatment, and sharply reduced in aba1 and aba2 mutants [42]. In summary, OsNAP appears to act in rice as a key regulator linking the ABA signaling and LS processes. No hormone other than ABA can regulate the OsNAP-mRNA level [42]. Understanding the specific regulatory interactions involving OsNAP and ABA highlights its importance in mediating stress responses and senescence in rice, particularly in response to ABA signaling. Likewise, overexpression of OsNAP also led not only to an increased expression of JA biosynthesis genes (i.e., OsLOX2 and OsAOC) but also to endogenous JA in transgenic plants [183]. On the other hand, OsNAC6 has been reported to be induced by exogenous JA [176,184], whereas ANAC019 and ANAC055 participate in the crosstalk between ABA and methyl jasmonate in plant defense response [176]. In tomato, SlNAP2 has a central role in controlling LS and fruit yield [84]. That is, both SlNAP1- and SlNAP2-mRNAs increased in the leaves during tomato dark-induced senescence. The AtNAC2 expression pattern following ABA treatment was similar to the salt stress treatment. ABA and salinity treatments resulted in an approximately threefold induction of AtNAC2 expression. However, the level of AtNAC1-mRNA was not influenced by salt stress, ABA, or ACC (i.e., the immediate precursor of ET) treatments, suggesting that NAC1 is not linked to stress in Arabidopsis [45]. In addition, in the ET-insensitive ein2-1 mutant, the salt induction of AtNAC2 was completely abolished, indicating that EIN2 is absolutely required for the signaling pathway that leads to the AtNAC2 induction under salt stress. However, the salt induction of the AtNAC2 gene was not affected by EIN3 (i.e., TF downstream EIN2) and was not dependent upon ABI2, ABI3, or ABI4 and the ABA signaling pathway. On the other hand, AtNAC2 promoted lateral root formation, and salt-induced AtNAC2 expression was dependent upon the ET and auxin signaling pathways [45]. As in Arabidopsis, rice, and cotton, expression of SlNAP2 is rapidly ABA-induced, demonstrating conservation of the upstream regulatory pathways that control the ABA-mediated induction of NAP genes across monocot and eudicot species [42]. In addition, it should be stated that the NAC TFs possessing the C-terminal transmembrane domain (TMD) are located in the ER.
Recently, by upregulating ABA signaling, it was reported in rice that the membrane-bound ONAC054 is essential for ABA-induced LS [197]. However, the cleavage of TMD allows ONAC054 (i.e., ONAC054α) to relocate to the nucleus. LOF onac054 mutants exhibited a functional stay-green phenotype. Interestingly, ONA054α has an alternative splice variant (i.e., nuclear ONAC054β). Furthermore, the transcript level of ONAC054β increases at a much faster rate than that of ONAC054α [198]. In [197], it was demonstrated that the level of two splice variants is low in the absence of exogenous ABA, and the ONA054α,β level rises in the nucleus upon ABA signal perception. Probably, ABA removes TMD from ONAC054α by currently unidentified proteases. The nuclear pool of ONA054α, β promotes LS through direct activation of both OsABI5 and NYC1 (Chlb reductase compounds NON-YELLOW COLORING1) transcription [197,199,200]. More recently, it was proved that (i) dehydration stress controls the cleavage of TMD from ONAC054, as observed previously in the ABA treatment [197,200]; (ii) ONAC054 expression is upregulated by several ABRE binding factors (ABF); and (iii) ONAC054 overexpression improved the grain yield of rice without causing any growth defect [200]. Collectively, all of the above data demonstrate that the activity of ONAC054, which is important for ABA-induced LS in rice, is minutely controlled by multilayered regulatory processes. Interestingly, nuclear SNAC3 protein confers heat and drought tolerance at the rice vegetative stage through modulation of downstream ROS genes [96]. SNAC3 was shown to be induced by exogenous ABA. Overexpressing SNAC3 not only enhanced heat and oxidative tolerance via enhancing the cell membrane stability but also improved drought resistance by reducing water loss. Likewise, SNAC3 likely regulates not only ROS scavenging but also ROS metabolism. Consistently, silencing SNAC3 by RNAi caused increased sensitivity to drought, high temperature, salinity, and ABA treatments [12,96]. Data from ABA content, SNAC3 expression, and response in mutants suggest that SNAC3 may function mainly in an ABA-independent manner [96], in contrast to some previously reported NAC genes. Finally, last year, two SNAC3-OX lines and two SNAC3-RNAi lines were created and subjected to arsenic stress treatments [201]. Interestingly, SNAC3 overexpression significantly intensified rice tolerance to arsenic stress and boosted grain yield, while the expression elimination generated a contrary response in both parameters. For further clarification, SNAC3 overexpression induced the enzymatic antioxidant levels of transgenic rice lines which in turn scavenge ROS, causing decreased oxidative stress and enhanced arsenic stress tolerance [201].
On the other hand, MYC2, MYC3, and MYC4 are targets of JAZ2 [202], a gene expressed primarily in the stomata guard cells that directly regulate the expression of ANAC19, ANAC55, and ANAC72, modulating the degree of stomata opening. JAZ2 degradation and the subsequent activation of MYC2, MYC3, and MYC4, in turn, activates ANAC19, ANAC55, and ANAC72. MYC2 and MYC3 bind to the promoter of ANAC genes. When a pathogen is perceived, Arabidopsis plants without JAZ2 are alerted to the stomatal apparatus closure (i.e., stomatal immunity). This research also supports that JAZ2 is a repressor of ANAC gene expression mediated by MYCs [202,203]. Altogether, (i) NAC072, NAC019, and NAC055 may act individually in different stress responses. NAC019 and NAC055 are involved in JA and/or ET signaling pathways, whereas NAC072 has been reported to be associated with the ABA-dependent stress response and to be strongly induced by ABA [195,204]; (ii) the nac019nac055nac072 triple mutant was slightly more sensitive to ABA or dehydration than single or double mutants [205]; (iii) NAC072, NAC019, and NAC055 genes contain ABRE motifs, and so rapidly respond to exogenous ABA [206]; and (iv) the NAC072 gene is upregulated by overexpression of ABF3, and the LOF of NAC072 turned out in ABA insensitivity. Meanwhile, the LOF of both NAC072 and ABF3 further improves ABA insensitivity. In other words, it is interesting to observe that NAC072 displays a dual function in ABF3-mediated ABA signaling [207].
The interplay between the ET and auxin signaling pathways with NAC TFs is highly intricate and multifaceted. In a recent study, a mechanism has been elucidated wherein the master regulator of mitochondrial disturbance, ANAC017, activates auxin-related genes along with genes associated with the ET pathway. This mechanism plays a crucial role in the control of mitochondrial stress signaling [208]. Significantly, ANAC017 exerts direct control over components in both the auxin and ET pathways. This dual regulatory role indicates the versatility of ANAC017 in coordinating responses to mitochondrial stress and modulating the signaling pathways associated with auxin and ET. The integration of stress signaling with hormonal pathways is a common feature in plant responses to environmental cues, and ANAC017 appears to be a key player in this regulatory network. The binding of ANAC017 to the promoters of MKK9 and ACS2 indicates that enhanced ET biosynthesis, as indicated by the activation of ACS2, is associated with an early response to mitochondrial stress. Intriguingly, the manuscript discusses a model illustrating the roles of ET and auxin in governing the mitochondrial retrograde response (MRR) through the direct activation by ANAC017 [208]. In summary, MKK9 and EIN3 collaborate to induce the mitochondrial retrograde response (MRR) as a protective mechanism against mitochondrial disturbance. Concurrently, the ET-induced auxin biosynthesis pathway leads to an increase in IAA levels, which, in turn, suppresses the MRR. This establishes an intricate feedback loop between ET and auxin, allowing for a finely tuned response to mitigate mitochondrial disorganization. Additionally, it has been demonstrated that the nuclear-localized AtNAC017 positively interacts with xyloglucan endo-transglucosylase/hydrolase (XTH31), regulating its expression by directly binding to the XTH31 promoter region. This interaction further emphasizes the multifaceted roles of AtNAC017 in coordinating cellular responses, including interactions with CW-related processes through XTH31 regulation. Likewise, a groundbreaking discovery indicates that, for the first time, ANAC017 functions upstream of the target gene XTH31 to govern aluminum (Al) tolerance and retention in Arabidopsis [48]. In summary, this study demonstrates that (i) anac017 mutants exhibit reduced Al accumulation in root CW hemicellulose, specifically xyloglucan; (ii) considering the significance of XTH31, XTH15, and XTH17 in hemicellulose modulation, the authors propose the hypothesis that ANAC017 may also regulate the function of these three proteins; (iii) the expression of ANAC017 and the accumulation of ANAC017 protein significantly decreased under Al stress; and (iv) it has been established that ANAC017 functions as a transcriptional activator of XTH31. In a broader context, the overarching conclusion is that ANAC017 operates upstream of XTH31, thereby regulating Al tolerance in Arabidopsis [48]. This sheds light on the specific involvement of ANAC017 in the cellular responses to Al stress, potentially contributing to the understanding of plant mechanisms in coping with environmental challenges, particularly those related to metal toxicity.
In tomato, a typical climacteric fruit, both SNAC4 and SNAC9 are stimulated by a brief exposure to ET, playing a positive regulatory role in ET synthesis and contributing to the ripening process of tomatoes [134]. The reduced expression of ET signaling genes observed in fruits with silenced SNAC4/SNAC9 confirms the regulatory role of both TFs in ET production. Notably, the silencing of SNAC9 specifically hampers the expression of LeACS2, LeACS4, and LeACO1, which are crucial components of the ET biosynthesis pathway. This feature emphasizes the significance of SNAC9 in modulating specific genes involved in ET synthesis. In summary, SNAC4/9 has the ability to bind to the promoter regions of ET biosynthesis genes, exerting a positive influence on the ripening process [134].
4.2. The Complexity of NAC TFs In Vivo Is Marked by the Existence of Modules
The concept of transcriptional modules suggests that the TFs can form interconnected and functionally related groups. These modules may involve multiple NAC TFs cooperating or sharing regulatory pathways to collectively influence the expression of specific target genes. The formation of transcriptional modules adds complexity to the regulatory network governed by the NAC TF family, highlighting the intricate nature of their interactions and the potential for coordinated control over various biological processes. Understanding these modules can provide valuable insights into the precise regulatory mechanisms orchestrated by NAC TFs in different cellular contexts. However, the study of in vivo complexes remains to be addressed. Chromatin immunoprecipitation (ChIP) followed by high-throughput sequencing (ChIP-Seq) is a powerful technique that significantly advances our understanding of these transcriptional modules. It allows researchers to identify and map the genomic locations where specific TFs bind, providing critical insights into the regulatory elements that control gene expression and contribute to the complexity of transcriptional networks. This emphasizes the crucial role of the AtNAC017, AtNAC082, and AtNAC090 modules in modulating the timing of LS in Arabidopsis [83]. The regulatory landscape of LS involves multiple players, and AtNAC042 has been identified as a key negative regulator that modulates cellular H2O2 levels. It is noteworthy that while AtNAC017 mediates responses to H2O2 in plants, it is not induced by H2O2 itself. Intriguingly, the loss of ANAC017 function leads to increased ROS levels, contributing to the acceleration of LS. This underscores the complexity of regulatory interactions within the plant senescence process, where different NAC TFs play distinct roles in modulating cellular responses to oxidative stress [83]. Despite AtNAC090 not being induced by SA treatment, both AtNAC090 and AtNAC017 assume predominant roles in regulating SA and ROS responses within the AtNAC017, AtNAC082, and AtNAC090 module [83]. The distinct roles of AtNAC090 and AtNAC017 within the AtNAC017, AtNAC082, and AtNAC090 module, particularly in regulating SA and ROS responses, further highlight the functional diversity and specificity of individual NAC TFs. Unfortunately, despite a considerable amount of data on the AtNAC017-AtNAC082-AtNAC090 module, it is unknown how it performs its role once bound to DNA in vivo. Understanding modules at this level is a significant challenge for the future.
The role of TgNAP in Tulipa gesneriana contrasts with its counterpart AtNAP in Arabidopsis. While AtNAP induces precocious LS, TgNAP promotes petal senescence in Tulipa gesneriana. TgNAP achieves this by activating the expression of genes involved in SA biosynthesis, such as TgPAL1 and TgICS1. Additionally, TgNAP enhances pathways related to the detoxification of ROS. This illustrates the versatility of NAC TFs in different plant species and highlights their capacity to regulate diverse biological processes, including senescence, in a context-dependent manner. This illustrates the diverse functions of NAP-like TFs in regulating senescence processes in different plant tissues and species [209]. It effectively highlights the dual and multifaceted roles of NAC TFs in the regulation of LS, acting as both positive and negative regulators and influencing the process through the modulation of SA and ROS pathways. The intricacies of these regulatory networks demonstrate the versatility of NAC TFs in orchestrating the complex processes associated with plant senescence. In reference [209], it was revealed that TgNAP serves as a positive regulator in H2O2-induced LS, and SA induces the expression of TgNAP. This implies that TgNAP plays a role in both H2O2-induced LS and SA biosynthesis in tulip, emphasizing its involvement in the complex regulatory network governing senescence processes.
Returning to the functional module formed by AtNAC017, AtNAC082, and AtNAC090, it is noteworthy that these three components interact with and directly regulate the promoters of target genes [83]. This study reveals variations in regulatory importance among the components of the module. Specifically, AtNAC090 emerges as the primary regulator in downregulating LS by SA and ROS responses. In contrast, AtNAC017 takes on a prominent role in ROS eradication. This differential contribution emphasizes the nuanced and specialized functions of each component within the regulatory module.
As previously mentioned, JAZ2 primarily functions within guard cells. In contrast, the roles of ANAC19, ANAC55, and ANAC72 are not limited to guard cells; they extend to mesophyll cells where these proteins act to repress SA defenses. This highlights the diverse cellular functions and regulatory activities exhibited by different components within the plant defense and signaling pathways [210]. The findings highlight the presence of a signaling module, consisting of COI1-JAZ2-MYC2,3,4-ANAC19,55,72, responsible for the regulation of stomatal aperture. Notably, CORONATINE-INSENSITIVE-1 (COI1) interacts with JAZ proteins, facilitating JAZ ubiquitination and degradation through the 26S proteasome pathway. In summary, bacterial infection facilitated by CORONATINE induces the degradation of JAZ2, leading to the subsequent activation of MYC2, MYC3, and MYC4. These activated MYC proteins, in turn, transcriptionally induce the expression of ANAC19, ANAC55, and ANAC72, contributing to the overall regulatory response in plants. In Nelumbo nucifera Geartn, specific NAC TFs, namely NnNAC45, NnNAC003, NnNAC016, NnNAC043, NnNAC060, and NnNAC070, demonstrate significant induction under NaCl treatment. Notably, their functions in response to salt stress are contingent upon ABA signaling, suggesting that ABA plays a regulatory role in the functions of these NAC TFs in the context of salt stress. The induction of these NAC genes underlines their potential involvement in the plant’s adaptive mechanisms to cope with saline conditions [59]. The ability of NAC016, NAC025, and NAC070 proteins to form complexes with other proteins potentially indicates a collaborative role in plant processes [59]. Knockout of OsNAC041 alters plant hormone homeostasis, possibly causing the salt-sensitive phenotype [130]. In apple, the MdNAC047 gene was isolated and functionally characterized for its involvement in ET regulation of salt tolerance. MdNAC047 overexpression facilitated the release of ET and increased the expression of ET-responsive genes [211].
Studying SNAC5, SNAC6, SNAC9, and SNAC10 genes and utilizing RNA-Seq and ChIP-Seq, Chung et al. (2018) identified 391 direct target genes. Among these, 239 loci were bound by SNAC5, SNAC6, SNAC9, and SNAC01, leading to the upregulation of 65, 91, 115, and 186 genes, respectively [212]. On the other hand, AaNAC1 from Artemisia annua (sweet wormwood; Chinese wormwood) was induced by SA and MJ, indicating its potential roles in abiotic or biotic stresses in A. annua [213]. Specifically, SA induces AaNAC1 expression about 30-fold, while MJ induces a 40-fold increase. Under the treatment of SA + MJ, the expression level of AaNAC1 can reach up to 60-fold. The findings regarding AaNAC1 suggest its significance in improving artemisinin content, which is an antimalarial drug localized in trichomes, as well as enhancing plant defense. The study’s conclusions propose that AaNAC1 has potential applications in transgenic breeding to enhance both artemisinin content and drought tolerance in Artemisia annua. This highlights the potential of NAC TFs, such as AaNAC1, in molecular breeding strategies for the development of crops with improved pharmaceutical and stress tolerance traits [214]. Interestingly, a current study provides the first indication of the molecular basis of artemisinin biosynthesis regulation through YABBY–WRKY interactions, regulated by AaJAZ8. Additionally, it highlights the potential of AaYABBY5 overexpression plants as a valuable genetic resource for artemisinin biosynthesis [215].
Several studies showed that plant-specific NAC103 TF has a variety of biological functions in various stress responses. Recently, NAC103 was reported to be involved in the expression of genes related to DNA damage response [111]. Genetic experiments demonstrated that NAC103 is a positive regulator in the ABA pathway and also regulates several ABA-responsive downstream genes. NAC103 is upregulated by ABA treatment at both the transcriptional and post-translational levels and plays an important role in ABA responses, especially during seed germination. Overexpression of NAC103 inhibited seedling growth when plants were treated with ABA.
5. Spatial Expression of NAC Genes
Studying the spatiotemporal expression patterns provides insights into when and where specific NAC genes are active, shedding light on their contributions to plant growth, adaptation to environmental challenges, and other important biological processes i.e., [4,69,216,217]. Let us take a look at some outstanding works. The observation that the ANAC017-mRNA level is higher in flowers and siliques (i.e., seed formation) and relatively lower expression in roots, leaves, and stems suggests that this NAC TF may play specific roles in reproductive organs [48]. The study by Li et al. (2021) [181] that analyzed 80 HaNAC genes in Helianthus annuus (sunflower) provides valuable information about the expression patterns of these genes in various plant organs. Notably, the expression analysis across different organs, including pistil, stamen, mature leaves, roots, and seeds, sheds light on the tissue-specific roles of HaNAC genes. The observation that, except for HaNAC23 and HaNAC105, the remaining 78 genes were lowly expressed in seeds suggests that these particular NAC families may not play a prominent role during seed development or maturation [181]. The tissue-specific expression pattern of ONAC127 and ONAC129 in rice, with predominant expression in the pericarp and weaker expression in the starch endosperm and aleurone layer, proposes a role in seed development, particularly during the early and middle stages [52]. Understanding the specific functions of ONAC127 and ONAC129 in the pericarp and their potential interaction as a heterodimer could provide insights into the regulatory mechanisms of seed development. The findings regarding ClNAC68 provide valuable insights into its role in seed development and germination in watermelon (Citrullus lanatus) [177]. The knockout of ClNAC68 is associated with delayed seed maturation and inhibited germination. The observed decrease in IAA and ABA contents in seeds of clnac68 mutants during development suggests the involvement of ClNAC68 in regulating phytohormone levels. Specifically, this study indicates that ClNAC68 was highly expressed in sweet flesh and acts as a transcriptional repressor that positively regulates seed germination. This positive regulation is achieved by increasing the free-IAA content through the direct repression of the IAA deactivator ClGH3.6. Additionally, ClNAC68 is implicated in the positive regulation of ABI5 expression, enhancing ABA sensitivity and influencing seed germination. These findings highlight the complex regulatory mechanisms involving ClNAC68 in the control of phytohormone levels and the expression of key genes related to seed germination and maturation in watermelon [177,178]. The high expression of OsNAC24 in the immature endosperm of rice grains may indicate its involvement in the control of processes such as nutrient allocation, storage compound biosynthesis, and overall seed development. The specific expression pattern of OsNAC24 in the immature endosperm points to its likely involvement in the molecular pathways associated with endosperm maturation, nutrient storage, and possibly other aspects of seed development [218]. Interestingly, ChIP-PCR assays revealed that the OsNAC24 gene directly targets the promoters of six starch-synthesis enzyme-coding genes and regulates their expression. The OsNAP gene interacts with OsNAC24 to coactivate the expression of its target genes and facilitates the localization of OsNAC24 to the nucleus. In other words, OsNAP and OsNAC24 likely form a protein–protein interaction that enhances their ability to regulate specific target genes [218].
A detailed study on the spatial expression of NAC genes was recently published in an article about jojoba (Simmondsia chinensis) [219]. It is interesting to note that a total of 57 NAC genes have been identified. Among these, seven are highly expressed in seeds, and specifically, ScNAC34, ScNAC37, and ScNAC14 show higher expression in the embryonic axis compared to other genes, suggesting potential key roles in seed development. Additionally, ScNAC34, ScNAC37, and ScNAC39 are highly expressed in both cotyledon and embryonic axis tissues. Indeed, the positive or negative correlation between the expression of NAC genes and seed development suggests that ScNAC genes likely play a regulatory role in the growth and development of Simmondsia chinensis seeds. Moreover, as previously determined for the ClNAC68 gene (210), the presence of auxin response elements in 30 ScNAC genes indicates their potential involvement in auxin-mediated processes during the development of jojoba seeds [219].
In relation to Triticum aestivum, quantitative PCR (qPCR) analysis revealed higher expression levels of TaNAC100 in developing seeds, particularly at 5 DAF (days after flowering). This suggests that TaNAC100 may play a specific role during the early stages of Triticum aestivum seed development. The increased expression of TaNAC100 in developing seeds, as opposed to other tissues such as stems, leaves, roots, and young spikes, indicates its potential involvement in processes related to seed formation and maturation [185]. The observation that TaNAC100 demonstrates an appropriate spatiotemporal expression pattern and co-localizes with GLU-1 (a gene associated with glutenin synthesis in wheat) suggests the possibility of functional interactions or shared regulatory pathways between these two genes. This feature could potentially contribute to the overall quality and characteristics of wheat seeds. To gain deeper insights, it would be beneficial to investigate the specific functions of TaNAC100 and GLU-1, explore their regulatory networks, and determine whether they directly interact or influence each other’s expression. The findings from Li et al. (2021) indicate that TaNAC100 functions as a transcriptional repressor in modulating GLU-1 expression [185]. Notably, similar to OsNAC2 [123], overexpression of TaNAC100 also leads to a significant reduction in plant height and a delay in flowering time, suggesting a conserved function between TaNAC100 and OsNAC2. Further investigations into the specific mechanisms through which TaNAC100 and OsNAC2 regulate plant height and flowering time would be intriguing. Additionally, exploring their common downstream targets or pathways could provide valuable insights into their shared regulatory functions.
In soybean, a recent study has discovered two NAC TFs exclusively expressed in nodules and endosperm, along with one that is spatiotemporally expressed. Firstly, SNAP1-4 NAC TFs, referred to as Soybean Nitrogen Associated NAPs (SNAPs), primarily impact the N-responsive transcriptome in mature nodules and rank among the most highly connected hub TFs. Secondly, NAP clade TFs exhibit evolutionary conservation. The authors of this study identified 388 genes directly regulated by SNAP1-4 that responded to nitrogen (N) (223 activated and 165 repressed) [180]. The nodules of snap1-4 quadruple mutants exhibit reduced sensitivity to the high N inhibition of nitrogenase activity, leading to an acceleration of senescence. Additionally, RT-qPCR analysis confirmed a significant reduction in mRNA levels for all target genes in the nodules of these mutants. Integrative analysis reveals that SNAP1-4 significantly impacts the transcriptional response to high N through the direct regulation of a subnetwork comprising senescence-associated genes and transcriptional regulators. It is proposed that the transcriptional network mediated by SNAP1-4 may trigger nodule senescence in response to high N [180]. Intriguingly, SNAP1-4 genes play pivotal roles in regulating nodule responses to high N, and the mature nodules of snap1-4 quadruple mutants demonstrate a notable tolerance to high N exposure.
Secondly, an important function of the NAC family has recently been proposed in endospermic seeds [220]. Previously, it was demonstrated that the endosperm-specific ZOU (bHLH-type TF) promotes the weakening of endosperm CWs and facilitates invasive embryo growth [188]. The weakening of CWs likely occurs through the activation of CW-modifying enzymes [221]. Consequently, in ZOU mutants, the elimination of endosperm is delayed, leading to significant damage to embryo growth. Intriguingly, the role of ZOU in activating PCD seems to be evolutionarily conserved [222]. The gradual elimination of the endosperm in WT is initiated by a PCD program, likely regulated by multiple NACs. The specific NACs involved in this process remain unidentified [223]. The work by Doll demonstrates that both ZOU-controlled CW weakening and the PCD promoted by both ZOU and NACs are essential. These processes work together to eliminate the mechanical resistance of the endosperm, facilitating the rapid and effective invasive growth of the A. thaliana embryo. In essence, (i) ZOU is necessary for initiating PCD, and (ii) there are two parallel pathways: ZOU regulates endosperm softening, while PCD regulates endosperm loosening. As a result, the combined action of CW weakening and PCD actively facilitates the breakthrough of the embryo through the active auto-elimination of endosperm cells [220].
Thirdly, the NAC103 protein is a substrate of the 26S proteasome and is maintained at a low concentration under normal conditions [110]. Nuclear NAC103 can form hetero-complexes with various other NAC TFs. NAC103 is expressed in two sites (i.e., seeds and seedlings) and plays an important role in ABA responses, especially during Arabidopsis seed germination. Its expression is significantly upregulated by ABA, with the encoded protein being stabilized by exogenous ABA [110,224]. Under ABA treatment, overexpression of NAC103 inhibits seed germination rates and seedling growth, while nac103 mutants display increased seed germination rates. Previous reports indicate interactions between ABA-responsive NAC proteins and RING-H2, suggesting that this interaction regulates the expression of downstream genes [224]. All of these results suggest that the structure of NAC103 in the N-terminus may play a role in regulating ABA ABA downstream genes. These results suggest the possibility that the overaccumulation of NAC103-mRNA is responsible for root growth inhibition under conditions of excess boron in the rpt5a-6 mutant. Certainly, the mutation in NAC103 (rpt5a-6) alleviates DNA damage in an A. thaliana mutant sensitive to excess boron [179].
6. Involvement of NACs in Seed Development, Dormancy, and Germination
As previously indicated, NAC TFs play notable roles during plant growth and development, exerting significant influence on stress processes. However, the specific role of NACs in the first phases of the seed cycle has not yet been adequately addressed. Nevertheless, NARS1 and NARS2 regulate embryogenesis in A. thaliana by controlling ovule integument development and degeneration, seed morphogenesis, and silique senescence [225]. These authors provide evidence that (i) both TFs regulated the development of cells in the outer integument; (ii) the development of endosperm is delayed in nars1 nars2 mutant; (iii) NARS1 and NARS2 are not expressed in the torpedo-shaped embryo stage; (iv) the absence of both genes in the embryo does not affect embryogenesis; and (v) NARS1 and NARS2 redundantly regulate not only seed morphogenesis but also silique-specific processes [225].
6.1. Involvement of NAC TFs in Seed Reserve Accumulation
The function(s) of NAC TFs in the endosperm, constituting approximately 90% of kernel weight, have not been fully elucidated. Therefore, the molecular mechanism underlying nutrient uptake and the biosynthesis of storage reserves involved in endospermic seed filling, particularly in well-studied species of grasses such as rice and maize, has been recently updated [226]. The size and weight of seeds are critical characteristics in agronomically exploitable species, playing pivotal roles in determining their agricultural value and overall utility. In rice, the conserved genes OsNAC020, OsNAC023, and OsNAC026 are primarily highly expressed during seed development, and are specifically associated with grain (caryopsis) size and weight [227]. OsNAC020 and OsNAC023 have the capability to form dimers with OsNAC026. Additionally, these dimers are reported to localize in the nucleus. Possibly, through their dimerization with OsNAC026, OsNAC020 and OsNAC023 are involved in the regulation of genes associated with seed size and weight [132,227]. On the other hand, ONAC127 and ONAC129 are expressed in the caryopsis and are involved in the regulation of starch accumulation and carbohydrate transport [52].
Nearly 30 genes participate in starch synthesis in maize [97]. However, the mechanism by which starch biosynthesis is regulated remains unclear, and specific information about the direct involvement of NAC TFs in their regulation is limited [44]. Both ZmNAC36 and ZmNAC34 are implicated in directly or indirectly regulating starch synthesis [58,189,190]. The ZmNAC34 gene, a classic TF with nuclear localization and transactivation activity, is specifically expressed during maize endosperm development, exclusively at 15 and 20 DAP. Its pivotal role in endosperm function is well-established [58]. ZmNAC34’s impact extends beyond maize, as evidenced by studies in rice. Overexpression of ZmNAC34 in rice resulted in decreased soluble sugar content and total starch accumulation. This effect was attributed to the downregulation of starch biosynthesis-related genes, leading to alterations in starch spatial structure [58]. In other words, ZmNAC34 is regarded as a negative regulator of starch biosynthesis, inhibiting the process. On the other hand, ZmaNAC36, encoding a predicted 38.40 kDa protein, exhibits strong expression during maize endosperm development, peaking at 15 DAP, compared to roots, stems, embryos, and flowers. ZmaNAC36 is co-expressed with most starch synthetic genes. Transient overexpression of ZmaNAC36 led to the upregulation of numerous starch synthesis genes in the endosperm, highlighting its regulatory role [189]. In conclusion, the close phylogenetic relationship between ZmNAC34 and ZmNAC36, both involved in the regulation of starch biosynthesis genes [189], raises the question of whether the function of ZmNAC34 in rice mirrors its function in maize, a topic that remains to be elucidated. In addition to ZmNAC36, the redundant genes ZmNAC128 and ZmNAC130, whose proteins do not interact, have been identified as positive regulatory factors for starch biosynthesis-related genes in rice and maize [44,228].
On the other hand, two functionally redundant TFs, named ZmNAC128 and ZmNAC130, not only act as activators of zeins in Zea mays but are also specifically and strongly expressed during the filling stage [44]. Deficient expression of ZmNAC128 and ZmNAC130 leads to a decrease in starch and protein accumulation. These genes directly regulate the expression of 16-kD γ-zein and AGPS2 (Bt2) by binding to a specific cis-element (ACGCAA) in their promoters [44]. Therefore, ZmNAC128 and ZmNAC130 are essential for maize endosperm filling, and both genes cooperate with the bZIP TF OPAQUE-2 (O2) promoter [191]. However, the specific contribution of each of the ZmNAC128 and ZmNAC130 genes to endosperm filling remains largely unknown. Recent results from Chen et al. (2023) [191] clearly indicate that ZmNAC128 and ZmNAC130 play crucial roles in the biosynthesis of zeins and starch, redundantly promoting endosperm filling. Their simultaneous LOF severely impaired the accumulation of storage reserves. In addition to 16-KD, the genes 27-kD and 50-kD γ-zein are also direct targets of ZmNAC128 and ZmNAC130. Interestingly, ZmNAC128 and ZmNAC130 also recognize cis-elements beyond the ACGCAA motif [191]. Knocking down the expression of ZmNAC128 and ZmNAC130 through RNA interference (RNAi) resulted in a shrunken kernel phenotype with a significant reduction in starch and protein content [44]. The absence of ZmNAC128 and ZmNAC130 had a pleiotropic effect on the utilization of carbohydrates and amino acids [44,191]. ZmNAC130 interacts with ZmNAC128, but they do not dimerize with each other. Both genes exhibit functional redundancy as they regulate the expression of the same downstream genes, such as 16-KD γ-zein and Bt2 [44,191]. In conclusion, the endosperm-specific ZmNAC128 and ZmNAC130 genes coordinate the accumulation of starch and proteins by regulating the expression of key starch biosynthetic enzymes and major seed proteins, including AGPS2 (Bt2) and the 16-kDa γ-zein (a type of prolamin). In other words, both genes oversee the rate-limiting step of starch synthesis [44].
In addition, the gene NAC-A18, predominately expressed in developing grains, regulates both starch and storage protein synthesis. This gene has been identified to directly bind to the cis-element ACGCAA in the promoters of Triticum aestivum TaLMW-D6 and TaLMW-D1 [41]. Ectopic expression of wheat NAC-A18 in rice significantly decreased starch accumulation and increased seed storage proteins (SSP) accumulation. In wheat, the nucleus-localized transcriptional repressor TaNAC019-A1, homologous to HvNAC019 [153], plays a crucial role in endosperm starch synthesis and negatively regulates kernel size and weight. Overexpression of TaNAC019-A1 inhibits starch synthesis in both wheat and rice kernels, directly binding to the promoters of TaAGPS1-A1 and TaAGPS1-B1 genes, thereby suppressing their expression. This repression of starch synthesis is achieved by downregulating the expression of multiple genes involved in starch synthesis, including TaAGPS1a and TaAGPS1b [43]. However, there is conflicting evidence, as Gao et al. (2021) [186] demonstrated that TaNAC019 acts as a promoter of starch synthesis and enhances the expression of SSP genes. Similarly, Li et al. (2021) [46] found that the TaNAC100 gene has the highest expression among all organs in developing seeds, peaking at 10 DAP, while TaNAC020 peaks at 15 DAP. TaNAC020s are predominantly expressed in developing grains and play a positive role in starch synthesis and accumulation. They are key regulators of seed size and number. Finishing the update of the notable T. aestivum NAC TFs, TaNAC100 functions as a transcriptional repressor, modulating GLU-1 expression by binding to its conserved cis-regulatory module (CCRM). Additionally, TaNAC100 is involved in the crosstalk between protein and starch synthesis in T. aestivum seeds [46].
The synthesis of SSPs and starch is spatiotemporally regulated at the transcriptional level by synergistic interactions between TFs and cis-elements distributed in SSP gene promoters. Recently, the TaSPR-A, TaSPR-B, and TaSPR-D NAC genes were found to be preferentially expressed in developing endosperms and were reported to suppress the transcription of SSP genes in T. aestivum, which has a complex genome [187]. The novel TuSPR gene was also found to be preferentially expressed in the developing endosperm during T. urartu filling, a species with a simpler genome. Both TuSPR and TaSPR, initially characterized as transcriptional repressors of SSP synthesis in T. aestivum, directly regulate SSP genes. Interestingly, in TuSPR-overexpressing common wheat, both the transcription of SSP genes and the accumulation of SSPs were impeded. TuSPR regulates SSP genes directly and binds to SSP gene promoters. TuSPR interacted with no TaSPR-A, TaSPR-B, or TaSPR-D genes [187].
Four NAC proteins—ZmNAC128 and ZmNAC130 from maize, and OsNAC20 and OsNAC26 from rice—sharing a high sequence similarity, were found to bind to a common sequence, ACGCAA, coordinating starch and protein synthesis [44,229]. OsNAC20 and OsNAC26 directly influence starch and storage protein accumulation by regulating the expression of genes related to their synthesis. In a study conducted by Wang et al. [229], it was demonstrated that OsNAC20 and OsNAC26 directly bound to the promoters of GluA1, GluB4, a-globulin, and 16 kD prolamine, regulating storage protein synthesis. Additionally, OsNAC20 physically interacts with OsNAC26 in plant cell nuclei. The NAC domains of OsNAC20 and OsNAC26 bind to the same cis-elements, activating similar starch and protein synthase-related genes. Similar to maize, the reduced starch content in the osnac20/26-1 mutant is not associated with the low expression of starch synthesis-related genes AGPS2b and AGPL2. In summary, OsNAC20 and OsNAC26 play an essential and redundant role in regulating starch and storage protein synthesis in rice [229]. Loss of function (LOF) of OsNAP led to altered expression in all tested starch and protein synthesis-related genes and reduced starch content. As mentioned earlier, CACG constitutes the core NAC-binding motif [30,204]. Jin’s research revealed that in addition to CACG, other motifs like TTGACAA, AGAAGA, and ACAAGA are present in the promoter regions of each of the OsNAC24 target genes [218].
In watermelon, the biological function of ClNAC68 has been investigated, revealing that this transcriptional repressor plays a critical role in sugar accumulation and seed development [178]. ClNAC68 positively regulates the accumulation of sugars, particularly sucrose, fructose, and glucose, by influencing the contents of free-IAA and -ABA. These findings align with the lower sucrose content observed in clnac68 mutant lines, where three SWEETs (sugar transporter family) were significantly downregulated. Additionally, ClNAC68 regulates sugar content by repressing invertase activity in watermelon [179]. In an interesting parallel, two recent studies, unrelated to the NAC family, demonstrated that ZmABI19 (ortholog of FUS3) and ZmbZIP29 are highly expressed in the early stage of seed development, playing initial roles in seed filling, and their expression is induced by ABA [87,175]. Overexpression of ZmABI19 and ZmbZIP29 resulted in enhanced O2 expression and increased seed weight and the contents of starch and zein proteins. As mentioned earlier, the endosperm-specific O2 is considered a central regulator of protein and starch synthesis. As ABA contents substantially increase at 8 DAP, endosperm filling is initiated. Thus, the elevated ABA levels promote O2 expression during endosperm filling. In other words, ZmbZIP29 is an ABA-inducible TF and directly regulates O2 expression genetically in the endosperm. The level of ZmbZIP29-mRNA is generally lower in the endosperm than in the embryo. ZmbZIP29 interacts with ZmABI19, exhibiting an additive transactivation pattern. Notably, the phosphorylation of ZmABI19, ZmbZIP29, and O2 by SnRK2.2 is essential for their full transactivation capacity. Cumulatively, mutations in zmbzip29 and zmabi19 significantly influence seed development and storage protein synthesis [87,175]. Taken together, ZmNAC128, ZmNAC130, ZmABI19, and ZmbZIP29 cooperate with O2 to facilitate endosperm filling and the synthesis of zeins and starch in the starchy endosperm [87,175,191]. That is, the ABA-mediated phosphorylation of two central coordinators, ZmABI19 and ZmbZIP29, by SnRK2.2 enhances gene transactivation for endosperm filling.
6.2. The Participation of NAC TFs in Seed Dormancy and Germination Processes
Seed dormancy is an omnipresent phenomenon in plants, induced and controlled by ABA [230,231,232]. Vivipary, characterized by low levels of dormancy, can lead to preharvest sprouting and substantial economic losses [233]. Upon the disappearance of dormancy under suitable conditions, germination ensues. Seed germination marks the initiation of the second cycle in a plant’s life, constituting a plant-specific developmental process wherein an embryonic plant grows to form a seedling. While the majority of prior studies have focused on NAC-mediated regulation of stress and LS, only a few have delved into the involvement of NACs in seed dormancy and germination.
As discussed earlier, OsNAC2 plays a crucial role in rice development and abiotic responses by binding to various downstream targets [125]. It inhibits seed germination and coleoptile growth, with OsACO or OsABA8ox1 operating downstream of OsNAC2. In essence, OsNAC2 acts as a regulatory element in ET, GAs, and ABA signaling, serving as a metabolic component to modulate rice seedling growth. The inhibition of seed germination by OsNAC2 is likely through the ABA pathway, targeting the promoters of OsNCED3, OsABA8ox1, and OsZEP1 [234]. The promotive effect of ET on seed germination may be counteracted by the excessive accumulation of ABA. Recent findings underscore the role of OsNAC2 in regulating seed dormancy release and the germination process [11]. OsNAC2, acting downstream of ABA signaling and upstream of GA signaling, exhibits high expression during seed development and germination compared to other plant tissues. OsNAC2 overexpression induces (i) inhibition of ABA degradation, leading to increased ABA content during early germination; (ii) enhanced seed dormancy and suppression of germination; (iii) the necessity of OsNAC2 for maintaining primary seed dormancy; and (iv) direct binding of OsNAC2 to the promoters of ABA metabolism genes (i.e., OsABA8ox1, OsABA8ox2, and OsABA8ox3), inhibiting their transcription during seed germination [11]. Notably, OsNAC2 is expressed in dry seeds, with its mRNA markedly diminishing during germination. In summary, experiments in [11] provide evidence supporting the hypothesis that OsNAC2 is involved in the regulation of ABA metabolism to maintain primary seed dormancy.
Recently [227], it was conjectured that OsNAC20 and OsNAC26, whose mRNAs are transcribed in the very early stages of rice zygotic embryogenesis, might function in controlling seed dormancy, as both genes are exclusively detected in the developing endosperm. Alongside OsNAC20 and OsNAC26, eight other NAC genes were identified in O. sativa seeds. Notably, OsNAC25, OsNAC41, OsNAC128, and OsNAC129 were found to be downregulated in the endosperm of the osnac20/26 mutant. Among these, OsNAC25, OsNAC128, and OsNAC129 are endosperm-specific genes, with OsNAC25 exhibiting a similar expression pattern to that of OsNAC20 and OsNAC26 [227]. Interestingly, OsNAC127 and OsNAC129 were identified as regulators of the seed-filling process, forming heterodimers. Both overexpression and knockdown of these genes resulted in incomplete grain filling and shrunken grains. OsEATB (AP2/ERF factor), OsMSR2 (calmodulin-like protein), OsSWEET4 (sugar transporter), and OsMST6 (monosaccharide transporter) were identified as direct targets of OsNAC127 and OsNAC129 proteins during rice grain filling [52]. In summary, data from Ren et al. (2021) suggest that OsNAC127 and OsNAC129 might play critical roles in the translocation and mobilization of starch to the developing rice endosperm [52].
Rice OsNAC041, induced by salinity, plays a role in affecting seed germination under salt stress [130]. Additionally, (i) several hormone-associated pathways contribute to OsNAC041-mediated salt tolerance; (ii) OsNAC041 influences ABC transporters, thereby regulating growth and development; and (iii) OsNAC041 is associated with the ROS system and membrane protection, modulating salt sensitivity [130]. On a different note, OsNAC3 positively regulates seed germination by involving the ABA pathway and the cell expansion gene OsEXP4 [12]. Consequently, OsABA8ox1 and OsEXP4 are considered putative target genes of OsNAC3. In other words, OsNAC3 directly binds to the promoters of OsABA8ox1 and OsEXP4, inducing their expression during seed germination. This implies that OsNAC3 is actively involved in cell expansion during seed germination through ABA signaling. Notably, OsNAC3 expression is not detected in dry rice seeds; it, along with OsEXP4, peaks sharply at 12 h of imbibition, declining thereafter [12]. Acting as a nuclear protein, OsNAC3 directly influences embryo cell elongation by regulating OsEXP4 expression. In conclusion, the expressions of OsABA8ox1 and OsEXP4 are activated during rice seed germination [12].
A T-DNA insertion in the ANAC060 gene results in increased seed dormancy compared with the WT [235]. Recently, Song et al. (2022) conducted a comprehensive study of ANAC040, ANAC060, and ANAC089 [236]. ANAC040 is likely more ancestral than ANAC060 and ANAC089. These genes, located in the same synteny cluster, indicate a common genomic origin shared with other eudicots but lacking in monocots. This was not expressed in dry seeds. Mutant anac060-1 and anac060-2 seeds exhibited deeper primary dormancy. The dormancy of anac060-2 anac040 did not significantly differ from anac060, suggesting that ANAC060 is epistatic over ANAC040 in dormancy regulation. ANAC040 is epistatic over ANAC060 in regulating salt sensitivity, as anac040 and anac060-1 anac040 were resistant to salinity, whereas Col-0 and anac060-1 were not. However, functional redundancy exists between ANAC060 and ANAC040, but not between ANAC060 and ANAC089 [236]. ANAC060 regulates seed dormancy and sugar (glucose and fructose) sensitivity [234]. Notably, ANAC060 expression levels are high in dry seeds but significantly reduced during seed imbibition. Additionally, ANAC060 expression is highly induced by cold stratification. ANAC060 and ANAC040 have opposite functions compared to DOG1, with DOG1 being crucial for inducing seed dormancy [237], whereas ANAC040 and ANAC060 inhibit seed dormancy.
Regarding ANAC089, its expression occurs during seed maturation, peaking in dry seeds and decreasing during imbibition [238]. The gap1-2 mutant reduces the transcript abundance of ANAC089, and the GA3ox2 gene responsible for active GA biosynthesis is induced in the gap1-2 mutant, suggesting increased levels of bioactive GAs to promote germination. Alterations in the cell redox status translocate ANAC089 into the nucleus and promote ANAC089 protein accumulation. ABA can induce the expression of ANAC089 and protein accumulation in seeds and seedlings. The authors conclude that ANAC089 regulates NO levels and cell redox, represses ABA synthesis and signaling, and binds specifically to genes controlling seed germination and abiotic stress [238]. Remarkably, redox-sensitive ANAC089 integrates ABA and NO responses during seed germination, conferring key functions to this central regulator in the successful growth of the plant under abiotic stress.
In a groundbreaking study, Kim et al. (2008) demonstrated that NAC with TRANSMEMBRANE motif 1-LIKE 8 (NTL8 or ANAC040), an Arabidopsis membrane-bound NAC TF, accumulates in response to growth and temperature, specifically induced by high salinity [239]. Recent advancements in our understanding of plant cold response, as outlined in [57], have shed light on the evolving roles of NAC TFs. NTL8 is significantly induced during imbibition and further elevated by cold treatment, suggesting that NTL8 expression is closely associated with the early stages of seed germination. Recent studies have prominently highlighted the central role of NTL8 as a cold-specific transcriptional activator in orchestrating plant cold response. This key regulatory function underscores NTL8’s pivotal role in governing molecular processes that contribute to plants’ ability to adapt and thrive under cold conditions [57]. NTL8 undergoes controlled proteolytic release from its membrane-bound form, and this release is specifically activated by high salinity. This release allows NTL8 to function as a transcriptional activator in the nucleus, where it can regulate the expression of target genes involved in various processes, including seed germination. Moreover, in the cold-resistant mangrove species Kandelia obovata, a recent study investigated the expression patterns of KoNAC6, KoNAC15, KoNAC20, KoNAC38, and KoNAC51 genes under cold treatment. The findings strongly underscore the vital role played by KoNAC genes in the adaptation and response of Kandelia obovata to chilling conditions [168]. This insight enhances our understanding of the molecular mechanisms underlying cold resistance in plants. NTL8 acts by sharply repressing GA3ox1, thereby implicating itself in the salt regulation of seed germination through the GAs pathway. Essentially, NTL8 reduces seed germination under salinity by modulating the GA signaling pathway, which typically promotes germination under favorable growth conditions [239]. In other words, GAs repress both NTL8 transcription and NTL8 processing. Moreover, NTL8 may function as a molecular link that incorporates environmental signals into the GA-mediated signaling pathway in seed germination. The ntl8-1 mutant, on the other hand, functions as a positive regulator of seed germination under high salt conditions. Importantly, NTL8-mediated salt signaling is found to be independent of ABA [239]. Further insights into NTL8’s role come from Tian et al. (2017), who identified TRIPTYCHON (TRY) and TRICHOMELESS1 (TCL1) as direct targets of NTL8 in Arabidopsis. NTL8 negatively regulates trichome formation by directly activating the expression of TRY and TCL1, two inhibitors of trichome development [240].
Recent studies across various plant species underscore the crucial role of auxins in orchestrating the coordination of seed life [241]. Building on this, a connection was established between the NAC family and auxins in the regulation of seed germination. Specifically, NTM2 (ANAC069), identified as a plasmalemma-bound NAC TF, integrates both auxin and salt signals to regulate seed germination in Arabidopsis [242]. NTM2 is induced by high salinity. Notably, the ntm2-1 mutant, unresponsive to high salinity, exhibits resistance to salt-induced germination inhibition, suggesting a role for NTM2 in the salt regulation of seed germination [242]. Intriguingly, NTM2’s function in salt signaling is independent of ABA and GAs. However, auxin influences the germination of ntm2-1 seeds under salinity. Simultaneously, NTM2 modulates auxin signaling through the INDOLE-3-ACETIC ACID 30 (IAA30) gene, linking salt signals to auxin signaling. In conditions of high salinity, auxins and the induction of the IAA30 gene, seen in the WT but not in the ntm2-1 mutant, contribute to delayed germination. In conclusion, NTM2 negatively regulates seed germination under salt stress by upregulating IAA30 expression [242]. Additionally, the NTL8 protein undergoes proteolytic activation in the presence of high salt, leading to the repression of the GA3 oxidase 1 (GA3ox1) gene in an ABA-independent manner [239]. Within the intricate network involving auxin, GAs, and salt signaling, a lingering question remains: how does the positive regulatory role of auxin on radicle emergence become compromised under high salinity conditions [242,243]?
7. Molecular Evolutionary Analysis of NAC TFs: A Derived Origen from WRKY
The transition to sessile life and terrestrial colonization of plants has been accompanied by significant genetic, developmental, and biochemical innovations in plant physiology. Consequently, the terrestrial colonization event led to a notable divergence and subsequent diversification of lineages adapted to either aquatic or terrestrial environments. One such example is the WRKY TF family, a group of proteins now recognized as crucial regulators of plant growth, development, and response to environmental stresses. The first WRKY protein, SWEET POTATO FACTOR1 (IbSPF1), was discovered in Ipomoea batatas [244]. A single copy of the WRKY gene, encoding two WRKY domains, was identified in the unicellular protist Giardia lamblia, a primitive unicellular eukaryote that diverged approximately ~1500 million years ago, as well as in the green alga Chlamydomonas reinhardtii [245]. It appears that the ancestral WRKY gene underwent numerous duplications throughout the evolution of plants, leading to the emergence of a large gene family in angiosperms [246]. Indeed, evolutionary analyses suggest that the C-terminal domain of the two-WRKY-domain encoding gene acts as the precursor to the single-WRKY-domain encoding genes. This observation indicates a possible evolutionary pathway where gene duplication events lead to the formation of new genes with modified structures and functions [247]. In the kingdom Plantae (Archaeplastida), the presence of WRKY proteins has been identified across the entire range of lineages studied from Rhodophyta to angiosperms [245]. These findings underscore the evolutionary significance and complex organization of WRKY proteins [248].
Currently, the evolutionary relationship between WRKY and NAC TFs is a topic of broad debate, mainly because no phylogenetic analysis encompassing both families has yet been achieved. The phylogenetic tree from this update (Figure 3) shows that members of both TF families are present in Charales, Zygnematales, and liverworts. However, Chlorophytes, including the model algae Chlamidomonas reihardii, which diverged earlier than the three aforementioned families, possess members of WRKY but not NAC. This observation supports the view that the WRKY family is evolutionarily older than NAC. Nevertheless, it is worth considering the possibility that the NAC family may have disappeared in Chlorophytes. Interestingly, the W-box elements for NAC and WRKY TFs exhibit high homology. Yet, some researchers do not support the idea of a close phylogenetic relationship between WRKY and NAC. Whereas the WRKY family is found in plants, fungi, and protists, suggesting an ancient evolutionary origin, the NAC family is present in plants and some algae, with its origin likely occurring later in evolutionary history. NAC transcription factors appeared before the emergence of land plants [24]. Evolutionary analysis indicated that segmental duplication and tandem duplication were the main mechanisms contributing to the expansion of the NAC gene family [249]. Structurally speaking, both WRKY and NAC proteins, along with the three more TFs families, belong to the GCM domain factors class [7]. WRKY and NAC share a common feature: a core β-sheet structure, with the edge of this β-sheet playing a crucial role in DNA recognition in a sequence-specific manner. This edge is a main part of the “WRKYGQK” motifs in WRKY and “WKATGTD” motifs in NAC. However, there are differences between WRKY and NAC: (i) WRKY possessed a Zinc-binding motif (not found in NAC); (ii) NAC proteins often have a dimerization arm, a structural motif not typically found in WRKY; (iii) WRKY often has two tandem repeated WRKY domains, whereas only a limited number of NAC exhibit such domain repetition; and (iv) in terms of DNA recognition, WRKY primarily utilizes one edge of the β-sheet (characterized by the ‘‘WRKYGQK” motifs) to interact with the major groove of DNA. In contrast, NAC employs an additional loop, positioned before the “WKATGTD” motifs, to recognize the minor groove of DNA as well [250]. To summarize, there are significant differences between NAC and WRKY, suggesting that they may have evolved from a common ancestor but subsequently diverged into two major TF groups in plants, which underscores the crucial role of NACs in plant evolution. Our tree (Figure 4) supports the view that NAC lineages diverged from WRKY lineages after the divergence of WRKY groups 1N and 1C (sensu Zhang and Wang, 2005 [247]) and prior to the emergence of WRKY genes encompassed in groups 2a; 2b, 2c, 2d, 2e and 3. Although certain WRKY and NAC may exhibit similarities in non-conserved regions apart from the structural domains, some authors reject the view of a direct relationship between WRKY and NAC based solely on these sequences (Bohan Liu, personal communication, 2024). In their view, similarities could be attributed to genetic recombination and sequence exchange processes during the course of evolutionary events.
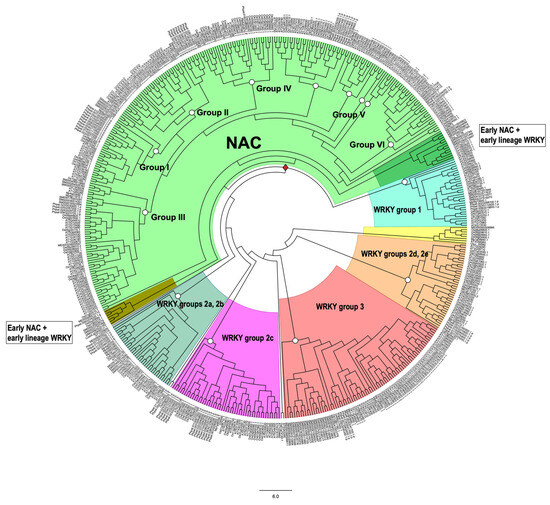
Figure 3.
Maximum-Likelihood phylogenetic analysis of 266 NAC and 268 WRKY protein sequences, aligned with ClustalW, performed in IQ-TREE (2021) under a JTT + R7 model chosen according to BIC. Red diamond indicates the node (BS 85%) showing a sister relationship between WRKY group 1 and NAC genes. White circles designate main groups of NAC genes following Pereira Santana et al. (2015) [251], and WRKY genes according to Zhang and Wang (2005) [247].
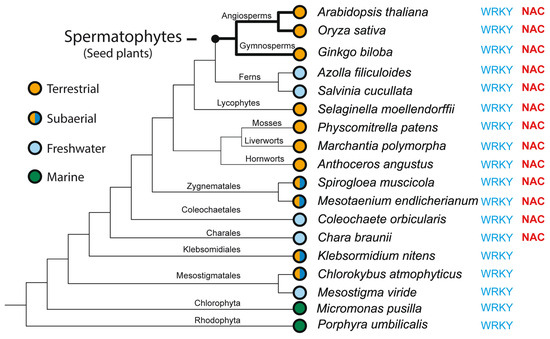
Figure 4.
This tree indicates that Klebsomidiales is the last lineage without NAC TFs, while Charales is the first lineage with WRKY and NACs. For each species, the habitat and documented occurrence of WRKY and NAC gene families are depicted. Adapted from Leebens-Mack et al. 2019 [252] and Wang et al. 2021 [253].
8. New Findings (2020–2024), Concluding Remarks, and Challenges for the Future
A significant number of plant-specific NAC (NAM-ATAF-CUC) genes/proteins have been isolated and studied across various species, yielding several notable findings. Through research outcomes spanning the past two decades, it has become widely recognized that this TF family plays multiple roles in signaling regulatory networks. However, functional studies of NAC genes are still limited and only a few members of this family display well-known functions. Remarkably, particularly notable NAC TFs, including GmNAC30, GmNAC81, ORE1 (ANAC92), OsNAC2, NAP (NAC-like and activated by AP3/PI), ATAF1 (ANAC002), and JUB1 (ANAC042), among others, have undergone thorough analysis at the whole-genome level, including phylogenetic analysis, collinearity analysis, gene structure, motif analysis, cis-element analysis, etc., and examination of gene expression patterns [254,255]. Through these analyses, they have been identified as central hubs regulating both plant development and stress responses. Let us take a look at some very recent significant studies that have not been previously discussed in detail. In soybean, GmNAC30 and GmNAC81 collaborate to regulate the expression of VPE. Likewise, GmNAC81 may negatively affect drought tolerance, not only by activating VPE but also by suppressing ABA signaling [113]. These two findings suggest that GmNAC81 acts as a converging link connecting signals of stress-induced cell death with drought responses [256]. In addition, asparagine(N)-rich protein (NRP) induces GmNAC030 and GmNAC081, which form a nuclear heterodimer to fully activate the expression of VPE, the executor of the cell death program via vacuole collapse. That is, the NRP-NAC-VPE cell death signaling conserved module integrates osmotic and ER stress into a signaling cascade that leads to a cell death fate [257]. To summarize, the integration of stress signals into a cell death pathway seems clear. On the other hand, the in vivo binding of ORE1 (ANAC092/ORESARA 1) to the promoters of two senescence-associated genes like SAG29/SWEET15, involved in nutrient re-mobilization and chlorophyll degradation, and SINA1, a RING E3 ligase, was demonstrated in Arabidopsis. That is, ORE1 directly regulates the expression of SAG29 and SINA1. This feature supports the idea that ORE1 is a master regulator in the process of LS, a biological pathway strictly controlled by gene-regulatory networks [116]. Unfortunately, the mediator subunit required for ORE1 activity, as well as the mechanistic basis for the recognition of the sequence of related genes, remains uncertain. Nevertheless, it was recently demonstrated that MED19a and ORE1 physically interact. This interaction has been observed both in vitro and in vivo during the deficiency of nitrogen, an inducer of LS [258]. Overall, this event highlights an important discovery in plant biology regarding the regulation of LS and the molecular interactions involved in this process. Additionally, it has recently been demonstrated that ORE1 serves as an in vivo phosphorylation substrate for CPK1, a calcium-dependent protein kinase that regulates cell death [259]. In conclusion, elucidating the impact of CPK1-mediated phosphorylation on ORE1 function holds the potential to unveil novel regulatory mechanisms underlying cell fate decisions in response to environmental cues and stress stimuli. Recently, the crystal structure of the ORE1-NAC domain alone and its DNA-binding form were reported. The structure of DNA-bound ORE1-NAC revealed the molecular basis for nucleobase recognition and phosphate backbone interactions [55]. This information is crucial for understanding the precise mechanisms by which ORE1 regulates gene expression and constructs cellular responses to various stimuli.
Rice, as a well-studied species, has its orthologous counterpart to ORE1 in Arabidopsis, known as OsNAC2, and shares similarities in function, particularly in regulating LS. Recent studies have shown that OsNAC2 inhibits SA signaling [260]. It forms a stable nuclear complex with OsEREBP1, disrupting the transmission of SA-mediated signaling. This OsEREBP1-OsNAC2 complex plays a role in co-regulating the expression of target genes. Conversely, OsEREBP1, belonging to the AP2/ERF family, exerts positive regulatory effects on resistance to bacterial blight in rice. It achieves this by interacting with the disease resistance-related protein OsXb22a in the cytoplasm, where it stabilizes OsXb22a. These findings suggest that OsNAC2 plays a role in balancing the distribution of OsEREBP1 between the nucleus and cytoplasm. Additionally, OsNAC2 may indirectly inhibit SA synthesis by regulating OsICS1 (Isochorismate synthase 1), the SA receptor gene OsNPR1 (Nonexpressor of pathogenesis-related genes 1), and the SA synthesis genes OsPAL6 and OsPAL3, thereby modulating plant immunity. OsNAC2 bound to the promotor of OsICS1 and OsNPR1 and suppressed their promotor activities [260]. In conclusion, Zhong et al. (2023) discovered that OsNAC2 is a negative regulator in the resistance to a bacterial blight disease in rice. On the other hand, OsNAC2 plays a crucial role in plant development by directly regulating key genes in the GAs pathway and positively modulating CK signaling. Additionally, OsNAC2 inhibits cell division and serves as an integrator of auxin, GAs, and CK pathways, thereby contributing to the regulation of plant height and root development [126]. This year, the characterization of OsNAC103 revealed its role in repressing cell cycle progression, with OsCYCP2;1, a P-type cyclin protein, identified as a potential target gene [261]. Interestingly, these findings further contribute to our understanding of the regulatory networks governing cell cycle dynamics and plant growth mediated by NAC TFs in rice.
The NAP (NAC-like, activated by AP3/PI) genes found in plants are indeed important members of the NAC family, playing crucial roles in various biological processes, including SL [33,202]. NAPs are highly regulated by ABA. For instance, AtNAP, a senescence-specific TF that plays a key role in promoting LS, participates in GAs-mediated chlorophyll degradation by interacting with GA-insensitive (GAI) and repressor of ga1–3 (RGA). The interaction subsequently impaired the transcriptional activities of NAP to induce the expression of AAO3 and senescence-associated gene SAG113 [262,263]. Recently, the AtNAP-AtCKX3 (cytokinin oxidase 3) regulatory module was characterized [264]. The Y1H experiments demonstrated that AtNAP physically binds to the cis-element of the AtCKX3 promoter to direct its expression. The expression of AtCKX3, regulated by AtNAP, is specific to the LS process, thereby modulating it. This study highlights the importance of this module in regulating CK levels and influencing the LS process [264]. This research further suggests that the AtNAP-AtCKX3 module is implicated in LS by bridging the interaction between two antagonistic plant hormones, ABA and CK. In 2023, a noteworthy breakthrough took place [265]. The authors showed that the regulatory module ABA-AtNAP-SAG113 PP2C controls leaf longevity by dephosphorylating SAG114 SnRK3.25 kinase, a direct target of the regulatory module. The senescence-associated gene SAG114, epistatic to SAG113 PP2C, encodes SnRK3.25 (Calcineurin B-like protein), a serine/threonine protein kinase specifically expressed during LS. Interestingly, SAG114 is localized in the Golgi apparatus. The phosphorylation/dephosphorylation process is yet to be studied [266]. In 2022, the CCCH-type zinc finger gene known as Strong Staygreen (PvSSG) was characterized in Panicum virgatum. PvSSG, through protein–protein interaction, repressed the DNA-binding efficiency of PvNAPs, thereby functioning as an impediment in the progression of LS. Interestingly, the Y2H, pull-down, BiFC, and co-immunoprecipitation (co-IP) analyses provided evidence that PvSSG (SL repressor factor) directly interacted with PvNAPs (SL accelerator factor) in the nucleus, suppressing PvNAPs-induced precocious LS [267]. This study reports a novel regulatory module in LS consisting of PvSSG and a pair of senescence-promoting TFs, PvNAPs. Therefore, the presence of the PvSSG–PvNAPs module demonstrates a finely coordinated process of chlorophyll catabolism and LS. These findings shed light on the regulatory mechanisms involved in LS in switchgrass and offer insight into how protein interactions can influence the progression of this and other similar processes.
As demonstrated throughout this review, the LS process is of singular importance in the life of plants and therefore is extensively studied (i.e., OSNAC5). In the current year, the Borrill group achieved a significant breakthrough in the NACs-LS relationship in wheat (Triticum aestivum L.) [268]. Thus, TaNAC5-1, an ortholog of OSNAC5 expressed in senescing flag leaves, acts as a positive regulator of LS. To characterize TaNAC5-1, the authors employed missense mutations in TaNAC5-A1 and TaNAC5-B1 from a TILLING mutant population and also overexpressed TaNAC5-A1 in wheat. Mutation in TaNAC5-1 resulted in a delayed onset of flag LS, while overexpression of TaNAC5-A1 led to a slightly earlier onset of senescence. Unfortunately, the DAP-seq analysis for TaNAC5-1 yielded limited information. Nonetheless, two serine peptidases and an α/β gliadin gene were identified as potential targets of TaNAC5-1 [268].
The importance of the ATAF-like NAC group in a diverse array of stress-related signaling processes (e.g., pathogenesis) is undeniable. This group consists of four genes in Arabidopsis and two in rice. ATAF1 (ANAC002) and ATAF2 (ANAC081) from Arabidopsis were among the first described NAC members. Analysis of the four Arabidopsis ATAF NAC promoter regions reveals an over-representation of both an ABA-responsive element (YACGTGGC) and G-box (CACGTG), implicating regulation by bZIP TFs and providing some plausible explanation of their ABA and stress responsiveness. Interestingly, ATAF1 has emerged as a potential regulator of Brassinosteroids (BRs) catabolism, whereas ATAF2 promotes LS [88]. Likewise, ATAF1,2, ANAC102, and CCA1 proteins exhibit both self and mutual physical interactions. Additionally, the expression of ATAF2, ANAC102, and ATAF1 is suppressed by BRs [269]. ANAC102 is involved in BRs catabolism and exhibits circadian regulation controlled by phytochromes, and its cellular localization remains uncertain [136,251,252,253,254,269,270,271,272].
The multifunctional JUB1 (ANAC042) TF, whose expression is induced by H2O2, is involved in camalexin biosynthesis, which is the major phytoalexin in Arabidopsis. JUB1 is also involved in the induction and forms the core of a robust regulatory module that triggers DELLA accumulation. Thus, the HB40-JUB1 regulatory network plays an important role in controlling GAs homeostasis during plant growth. HB40 directly activates the transcription of JUNGBRUNNEN1 (JUB1), a key TF that represses growth by suppressing GAs biosynthesis and signaling [271]. Previously, it was shown in tomato that SlDREB1, SlDREB2, and SlDELLA are potential target genes of SlJUB1 under drought stress. To summarize, AtJUB1 directly inhibits the expression of GAs and BRs biosynthetic genes, resulting in the accumulation of DELLA protein. This accumulation inhibits growth and increases plant resistance to stresses. Recently, GhJUB1L1 (homolog gene of AtJUB1) was proved to directly bind to the promoters of GhABI1, GhSOS2, GhCCoAOMT1, GhCesA7, and GhIRX14 in vivo, resulting in the induction of the expression of these five genes in cotton [254]. Zhang et al. (2022) indicate that (i) the expression of JUB1 was activated by CLE14 (CLAVATA3/ENDOSPERM SURROUNDING REGION); (ii) JUB1 functions downstream of the CLE14 signal; (iii) CLE14 expression was also significantly induced by ABA, JA, and SA; (iv) CLE14 is involved in the regulation of ROS accumulation during senescence and under stress conditions, potentially by promoting ROS scavenging activities in a JUB1-dependent manner; and (v) the CLE14-JUB1-ROS module serves as a braking mechanism, in the form of a negative feedback loop, to prevent precocious cell death and ensure that the LS process is undertaken in an orderly manner, such that nutrient remobilization can be accomplished in time [272]. Taken together, the results from Zhang et al. (2022) show that the small peptide CLE14 functions as a novel ‘brake signal’ to regulate age-dependent and stress-induced LS through JUB1-mediated ROS scavenging.
Very recently, and to conclude this section, two adjacent NAC genes named PpNAC1 and PpNAC5 were functionally characterized in peach (Prunus persica L.), demonstrating their roles in regulating fruit maturity date and flavor [136]. The Han group, demonstrated that PpNAC1 and PpNAC5, physically separated by a 19 863-bp DNA region, had several effects as follows: (i) They had pleiotropic effects on plant development, increase in organic acid metabolism and sugar accumulation by activating sugar transport genes. (ii) They participated in organic acid accumulation by activating the expression of the PpGAD gene, which encodes glutamate decarboxylase, contributing to citrate degradation and acidity reduction in peach. Therefore, the GAD gene appears to be a promising target for genetic improvement aimed at enhancing citrate levels in fruits. (iii) They interacted with each other in vivo (Y1H) to form heterodimer complexes that are conserved in eudicots. (iv) Both TFs functioned as ripening enhancers, with the former having a stronger ripening acceleration effect (divergence between their C-terminal regions?), and PpNAC5 could be more crucial for fruit enlargement than for fruit ripening. (v) The orthologs of both TFs were found in all tested angiosperms, but not in gymnosperms [136]. Finally, the results obtained to date demonstrate that NAC1 or NAC5 orthologues are not functionally exchangeable in dry fruit-bearing angiosperms.
The methodologies used for studying the NAC family of TFs have remained relatively consistent over the last twenty years, with a focus on whole-genome analysis and gene expression studies. Techniques such as next-generation sequencing, microarray analysis, and quantitative PCR have been instrumental in elucidating the functions and regulatory roles of NAC genes. Despite the methodological continuity, advances in bioinformatics tools, data analysis algorithms, and high-throughput sequencing technologies have enhanced the efficiency and depth of our understanding of the NAC family. Additionally, the integration of multi-omics approaches, such as combining genomics, transcriptomics, proteomics, and metabolomics data, has provided comprehensive insights into the regulatory networks and functional dynamics of NAC TFs. While the core methodologies may have remained consistent, the refinement and integration of these techniques have undoubtedly contributed to significant advancements in our understanding of the NAC family and its roles in plant development, stress responses, and other biological processes. However, the study of the interaction between TFs and chromatin in vivo is an aspect that is still to be addressed. So far, TFs have been studied in isolation. However, in vivo, TFs often form complexes with other TFs or with other factors (co-repressors, regulators, etc.) (see Section 5). The study of complexes in vivo is lacking; that is, it is necessary to address the study of the complex TF-DNA interaction in vivo. The state of chromatin is crucial for TF function. It is necessary to combine chromatin analysis (e.g., ATAC-seq) with the study of TF binding sites (preferably complexes of TFs-regulatory proteins) (e.g., ChIP-seq). For the study of complexes, techniques such as TAP-tagging or Turbo-ID should be used and combined with studies of Hi-C or similar methods for the 3D chromatin structure. These studies should ideally be dynamic to observe how these complexes vary over time or in response to stress.
Indeed, it would be very interesting to study the mechanisms and endogenous and/or exogenous signals operating in the cell to distribute the NAC TFs in tethering and non-tethering configurations. The phylogenetic investigation into the origin of the NAC family in relation to the WRKY family is also of significance. Figure 4 depicts an accurate and simplified phylogeny of the major lineages in Viridiplantae. In this phylogeny, Charales (specifically Chara braunii) is the first lineage in which the NAC TF appears.
Author Contributions
A.J.M. conceptualized the review outline and wrote the original draft and J.F.-A. prepared the entire phylogenetic study. All authors have read and agreed to the published version of the manuscript.
Funding
Javier Fuertes-Aguilar’s (J.F.-A.) research was funded by NEXTPOL project grant PGC2018-100684-B-I00, Spanish Ministry of Science and Innovation.
Acknowledgments
The authors thank Lluís García-Mir for assistance in bioinformatic tasks. Phylogenetic analysis was possible through the access granted by the Galician Supercomputing Center (CESGA) to its supercomputing infrastructure. The supercomputer FinisTerrae III and its permanent data storage system are funded by the Spanish Ministry of Science and Innovation, the Galician Government, and the European Regional Development Fund (ERDF). The authors would like to thank R. Solano (R.S.) for his invaluable contribution, and S. Mahiwal, P. Agarwal, M-S. Kim, and B. Liu for their assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Martínez-Ainsworth, N.E.; Tenaillon, M.I. Superheroes and masterminds of plant domestication. C. R. Biol. 2016, 339, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.A.; Mishra, S.; Mehrotra, S.; Upadhyay, S.K.; Bagal, D.; Srivastava, V. Plant transcription factors: An overview of their role in plant life. Plant Transcr. Factors 2023, 3–20. [Google Scholar] [CrossRef]
- Ding, Y.; Zhu, J.; Zhao, D.; Liu, Q.; Yang, Q.; Zhang, T. Targeting cis-regulatory elements for rice grain quality improvement. Front. Plant Sci. 2021, 12, 70583. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Guan, Y.; Weng, Y.; Liao, B.; Tong, L.; Hao, Z.; Chen, J.; Shi, J.; Cheng, T. Genome-wide identification of the NAC gene family and its functional analysis in Liriodendron. BMC Plant Biol. 2023, 23, 415. [Google Scholar] [CrossRef]
- Rui, Z.; Pan, W.; Zhao, Q.; Hu, H.; Li, X.; Xing, L.; Jia, H.; She, K.; Nie, X. Genome-wide identification, evolution and expression analysis of NAC gene family under salt stress in wild emmer wheat (Triticum dicoccoides L.). Int. J. Biol. Macromol. 2023, 230, 12337. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Das, S.; Mishra, M.; Choudhury, D.R.; Sharma, K.; Kumari, A.; Sing, R. Emerging roles of NAC transcription factor in medicinal plants: Progress and prospects. 3 Biotech 2021, 11, 425. [Google Scholar] [CrossRef]
- Blanc-Mathieu, R.; Dumas, R.; Turchi, L.; Lucas, J.; Parcy, F. Plant-TFClass: A structural classification for plant transcription factors. Trends Plant Sci. 2024, 29, 40–51. [Google Scholar] [CrossRef]
- Strader, L.; Weijers, D.; Wagner, D. Plant transcription factors—Being in the right place with the right company. Curr. Opin. Plant Biol. 2022, 65, 102136. [Google Scholar] [CrossRef]
- Zemlyanskaya, E.V.; Dolgikh, V.A.; Levitsky, V.G.; Mironova, V. Transcriptional regulation in plants: Using omics data to crack the cis-regulatory code. Curr. Opin. Plant Biol. 2021, 63, 102058. [Google Scholar] [CrossRef]
- De Caprio, J.; Okhol, T. Chromatin Immunoprecipitation. Cold Spring Harb. Protoc. 2020, 2020, 098665. [Google Scholar]
- Zhao, F.; Ma, Q.; Li, Y.; Jiang, M.; Zhou, Z.; Meng, S.; Peng, Y.; Zang, J.; Ye, N. OsNAC2 regulates seed dormancy and germination in rice by inhibiting ABA catabolism. Biochem. Biophys. Res. Commun. 2023, 682, 335–342. [Google Scholar] [CrossRef]
- Huang, C.; Zhao, J.; Huang, Q.; Peng, L.; Huang, Z.; Li, W.; Sun, S.; He, Z.; Wang, Z. Also, OsNAC3 positively regulates seed germination involving ABA pathway (i.e., OsNAC3 can directly bind to the promoter of ABA catabolic gene OsABA8ox1) and cell expansion gene OsEXP4. New Phytol. 2024, 241, 650–664. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Sadreyev, R. Single-cell RNA-seq: Introduction to bioinformatics analysis. Curr. Protoc. Mol. Biol. 2018, 124, e68. [Google Scholar] [CrossRef] [PubMed]
- Munir, A.; Yukun, C.; Xiaohui, C.; Nawaz, M.A.; Iftikhar, J.; Rizwan, M.; Xu, S.; Yuling, L.; Xuan, X.; Zhongxiong, L. Genome-wide identification and comprehensive analyses of NAC transcription factor gene family and expression patterns during somatic embryogenesis in Dimocarpus longan Lour. Plant Physiol. Biochem. 2020, 157, 169–184. [Google Scholar] [CrossRef]
- Zhou, X.; Su, H. DOF transcription factors: Specific regulators of plant biological processes. Front. Plant Sci. 2023, 14, 1044918. [Google Scholar] [CrossRef] [PubMed]
- Khoso, M.A.; Hussain, A.; Nurhasanah, F.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ng, C.K.-Y.; Fan, L.-M. MYB transcription factors, active players in abiotic stress signaling. Environ. Exp. Bot. 2015, 114, 80–91. [Google Scholar] [CrossRef]
- Dröge-Laser, W.; Snoek, B.L.; Snel, B.; Weist, C. The Arabidopsis bZIP transcription factor family—An update. Curr. Opin. Plant Biol. 2018, 45, 36–49. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, Z.; Wang, L.; Jin, B. Plant responses to heat stress: Physiology, transcription, non-coding RNA and epigenetics. Int. J. Mol. Sci. 2021, 22, 117. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, P. The DREB transcription factor, a biomacromolecule, responds to abiotic stress by regulating the expression of stress-related genes. Int. J. Biol. Macrom. 2023, 243, 125231. [Google Scholar] [CrossRef]
- Feng, K.; Hou, X.-L.; Xing, G.-M.; Liu, J.-X.; Duan, A.-Q.; Xu, Z.-S.; Li, M.-Y.; Zhuang, J.; Xiong, A.-S. Advances in AP2/ERF super-family transcription factors in plant. Crit. Rev. Biotechnol. 2020, 40, 750–776. [Google Scholar] [CrossRef] [PubMed]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The no apical meristem gene of Petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T.; Jenkind, J.; Shu, S.; Ishizaki, K.; Yamaoka, S.; Yamaoka, S.; Nishihama, R.; Nakamura, Y.; et al. Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 2017, 171, 287–304.e15. [Google Scholar] [CrossRef]
- Maugarny-Calès, A.; Gonçalves, B.; Jouannic, S.; Melkonian, M.; Wong, G.K.S.; Laufs, P. Apparition of the NAC transcription factors predates the emergence of land plants. Mol. Plant 2016, 9, 1345–1348. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Roy, S. Evaluation of the diversity and phylogenetic implications of NAC transcription factor members of four reference species from the different embryophytic plant groups. Physiol. Mol. Biol. Plants 2019, 25, 347–359. [Google Scholar] [CrossRef]
- Rinerson, C.I.; Rabara, R.C.; Tripathi, P.; Shen, Q.J.; Rushton, P.J. The evolution of WRKY transcription factors. BMC Plant Biol. 2015, 15, 66. [Google Scholar] [CrossRef]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Sablowski, R.W.; Meyerowitz, E.M. A homolog of NO APICAL MERISTEM is an immediate target of the floral homeotic genes APETALA3/PISTILLATA. Cell 1998, 92, 93–103. [Google Scholar] [CrossRef]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Lo Leggio, L.; Skriver, K. NAC transcription factors: Structurally distinct functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Shamimuzzaman, M.; Vodkin, L. Genome-wide identification of binding sites for NAC and YABBY trancription factors and co-regulated genes during soybean seedling development by ChIP-Seq and RNA-Seq. BMC Genom. 2013, 14, 477. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Nevo, E.; Sun, D.; Peng, J. Phylogenetic analyses unravel the evolutionary history of NAC proteins in plants. Evolution 2012, 66, 1833–1848. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Koyama, H.; Bhati, K.K.; Alok, A. The biotechnological importance of the plant-specifc NAC transcription factor family in crop improvement. J. Plant Res. 2021, 134, 475–495. [Google Scholar]
- Liu, G.S.; Li, H.L.; Grierson, D.; Fu, D.Q. NAC transcription factor family regulation of fruit ripening and quality: A review. Cells 2022, 11, 525. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qiao, Y.; Li, C.; Hou, B. The NAC transcription factors play core roles in flowering and ripening fundamental to fruit yield and quality. Front. Plant Sci. 2023, 14, 1095967. [Google Scholar] [CrossRef] [PubMed]
- Welner, D.H.; Deeba, F.; Leggio, L.L.; Skriver, K. NAC transcription factors: From structure to function in stress-associated networks. In Plant Transcription Factors; Academic Press: Cambridge, MA, USA, 2016; Chapter 13; pp. 199–212. [Google Scholar] [CrossRef]
- Jensen, M.K.; Kjaersgaard, T.; Nielsen, M.M.; Galber, P.; Petersen, K.; O’Shea, C.; Skariver, K. The Arabidopsis thaliana NAC transcription factor family: Structure–function relationships and determinants of ANAC019 stress signalling. Biochem. J. 2010, 426, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Li, J.; Hao, Z.; Wei, L.; Ma, J.; Zong, Y.; Li, H. Overexpression of the LcCUC2-like gene in Arabidopsis thaliana alters the cotyledon morphology and increases rosette leaf number. Peer J. 2022, 10, e12615. [Google Scholar] [CrossRef]
- Ernst, H.A.; Olsen, A.N.; Larsen, S.; Lo Leggio, L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef]
- Hussain, R.M.; Ali, M.; Feng, X.; Li, X. The essence of NAC gene family to the cultivation of drought-resistant soybean (Glycine max L. Merr.) cultivars. BMC Plant Biol. 2017, 17, 55. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Y.; Hao, C.; Li, T.; Majeed, U.; Liu, H.; Li, H.; Hou, J.; Zang, X. Wheat NAC-A18 regulates grain starch and storage proteins synthesis and affects grain weight. Theor. Appl. Genet. 2023, 136, 123. [Google Scholar] [CrossRef]
- Liang, C.; Wang, Y.; Zhu, Y.; Tang, J.; Hu, B.; Liu, L.; Ou, S.; Wu, H.; Sun, X.; Chu, J.; et al. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 10013–10018. [Google Scholar] [CrossRef]
- Liu, Y.; Hou, J.; Wang, X.; Li, T.; Majeed, U.; Hao, C.; Zhang, X. The NAC transcription factor NAC019-A1 is a negative regulator of starch synthesis in wheat developing endosperm. J. Exp. Bot. 2020, 71, 5794–5807. [Google Scholar] [CrossRef]
- Zhang, Z.; Dong, J.; Ji, C.; Wu, Y.; Messing, J. NAC-type transcription factors regulate accumulation of starch and protein in maize seeds. Proc. Natl. Acad. Sci. USA 2019, 116, 11223–11228. [Google Scholar] [CrossRef]
- Liang, M.; Li, H.; Zhou, F.; Li, H.; Liu, J.; Hao, Y.; Wang, Y.; Zhao, H.; Han, S. Subcellular distribution of NTL transcription factors in Arabidopsis thaliana. Traffic 2015, 16, 1062–1074. [Google Scholar] [CrossRef]
- Li, P.; Peng, Z.; Xu, P.; Tang, G.; Ma, C.; Zhu, J.; Shan, L.; Wan, S. Genome-wide identification of NAC transcription factors and their functional prediction of abiotic stress response in peanut. Front. Genet. 2021, 12, 30292. [Google Scholar] [CrossRef]
- He, X.-J.; Mu, R.-L.; Cao, W.-H.; Zhang, Z.-G.; Zhang, J.-S.; Chen, A.-Y. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005, 44, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wan, J.X.; Liu, Y.S.; Yang, X.Z.; Shen, R.F.; Zhu, X.F. The NAC transcription factor ANAC017 regulates aluminum tolerance by regulating the cell wall-modifying genes. Plant Phys. 2022, 189, 2517–2534. [Google Scholar] [CrossRef]
- Li, J.; Qiu, J.; Zeng, Q.; Yi, Z.; Zhang, N.; Xu, S.; Jin, J.; Dong, Z.; Chen, L.; Huang, W. OsTOC1 plays dual roles in the regulation of plant circadian clock by functioning as a direct transcription activator or repressor. Cell Rep. 2023, 42, 112765. [Google Scholar] [CrossRef] [PubMed]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef]
- Hussain, Q.; Asim, M.; Zhang, R.; Khan, R.; Farooq, S.; Wu, J. Transcription factors interact with ABA through gene expression and signaling pathways to mitigate drought and salinity stress. Biomolecules 2021, 11, 1159. [Google Scholar] [CrossRef]
- Ren, Y.; Huang, Z.; Jiang, H.; Wang, Z.; Wu, F.; Xiong, Y.; Yao, J. A heat stress responsive NAC transcription factor heterodimer plays key roles in rice grain filling. J. Exp. Bot. 2021, 72, 2947–2964. [Google Scholar] [CrossRef]
- Su, H.; Zhang, S.; Yin, Y.; Zhu, D.; Han, L. Genome-wide analysis of NAM-ATAF1,2-CUC2 transcription factor family in Solanum lycopersicum. J. Plant Biochem. Biot. 2015, 24, 176–183. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, H.; Cai, J.; Li, D.; Song, F. NAC transcription factors in plant immunity. Phytopathol. Res. 2019, 1, 3. [Google Scholar] [CrossRef]
- Chun, I.; Kim, H.J.; Hong, S.; Kim, Y.G.; Kim, M.-S. Structural basis of DNA binding by the NAC transcription factor ORE1, a master regulator of plant senescence. Plant Commun. 2023, 4, 100510. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Q.; Xiong, L.; Lou, Z. A structural view of the conserved domain of rice stress-responsive NAC1. Protein Cell 2011, 2, 55–63. [Google Scholar] [CrossRef]
- Diao, P.; Chen, C.; Zhang, Y.; Meng, Q.; Lv, W.; Ma, N. The role of NAC transcription factor in plant cold response. Plant Signal. Behav. 2020, 15, 1785668. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wang, Q.; Wang, Y.; Cheng, B.; Zhao, Y.S. A maize NAC transcription factor, ZmNAC34, negatively regulates starch synthesis in rice. Plant Cell Rep. 2019, 38, 1473–1484. [Google Scholar] [CrossRef]
- Zhao, S.; Jiang, T.; Zhang, Y.; Zhang, K.; Feng, K.; Wu, P.; Li, L. Identification of the NAC transcription factors and their function in ABA and salinity response in Nelumbo nucifera. Int. J. Mol. Sci 2022, 23, 12394. [Google Scholar] [CrossRef]
- Han, F.; Wang, P.; Chen, X.; Zhao, H.; Zhu, Q.; Song, Y.; Nie, Y.; Guo, M.; Niu, S. An ethylene-induced NAC transcription factor acts as a multiple abiotic stress responsor in conifer. Hort. Res. 2023, 10, uhad130. [Google Scholar] [CrossRef]
- Meng, X.; Liu, S.; Zhang, C.; He, J.; Ma, D.; Wang, X.; Dong, T.; Guo, F.; Cai, J.; Long, T.; et al. The unique sweet potato NAC transcription factor IbNAC3 modulates combined salt and drought stresses. Plant Phys. 2023, 191, 747–771. [Google Scholar] [CrossRef]
- Hofmann, N.R. Endoplasmic reticulum–localized transcription factors and mitochondrial retrograde regulation. Plant Cell 2013, 25, 3155. [Google Scholar] [CrossRef][Green Version]
- Ng, S.; Ivanova, A.; Duncan, O.; Law, S.R.; Aken, O.; Clercq, I.; Wang, Y.; Carrie, C.; Xu, L.; Kmiec, B.; et al. A Membrane-bound NAC transcription factor, ANAC017, mediates mitochondrial retrograde signaling in Arabidopsis. Plant Cell 2013, 25, 3450–3471. [Google Scholar] [CrossRef]
- Seo, P.J. Recent advances in plant membrane-bound transcription factor research: Emphasis on intracellular movement. J. Integr. Plant Biol. 2014, 56, 334–342. [Google Scholar] [CrossRef]
- Wang, D.; Yu, Y.; Liu, Z.; Li, S.; Wang, Z.; Xiang, F. Membrane-bound NAC transcription factors in maize and their contribution to the oxidative stress response. Plant Sci. 2016, 250, 30–39. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, S.Y.; Park, C.M. A membrane-associated NAC transcription factor regulates salt-responsive flowering via FLOWERING LOCUS T in Arabidopsis. Planta 2007, 226, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, P.; Das, R.; Mandal, A.; Kundu, P. Functional characterization of tomato membrane-bound NAC transcription factors. Plant Mol. Biol. 2017, 93, 511–532. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ma, Z.; Sun, W.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide analysis of the NAC transcription factor family in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2019, 20, 113. [Google Scholar] [CrossRef]
- Yang, Q.; Li, B.; Rizwan, H.M.; Sun, K.; Zeng, J.; Shi, M.; Guo, T.; Chen, F. Genome wide identification and comprehensive analyses of NAC transcription factor gene family and expression analysis under Fusarium kyushuense and drought stress conditions in Passiflora edulis. Front. Plant Sci. 2022, 13, 972734. [Google Scholar] [CrossRef]
- Li, H.; Shi, X.; Yu, B.; Zhou, Y.; Chen, S.; Wang, Y.; Peng, Y.; Meyer, R.C.; Smeekens, S.C. The ABI4-induced Arabidopsis ANAC060 transcription factor attenuates ABA signaling and renders seedlings sugar insensitive when present in the nucleus. PLoS Genet. 2014, 10, e1004213. [Google Scholar] [CrossRef]
- Shu, L.; Li, L.; Jiang, Y.; Yan, J. Advances in membrane-tethered NAC transcription factors in plants. Plant Sci. 2024, 342, 112034. [Google Scholar] [CrossRef]
- Yang, Z.T.; Wang, M.J.; Sun, L.; Lu, S.J.; Bi, D.L.; Sun, L.; Song, Z.T.; Zhang, S.S.; Zhou, S.F.; Liu, J.X. The membrane-associated transcription factor NAC089 controls ER-stress-induced programmed cell death in plants. PLoS Genet. 2014, 10, e1004243. [Google Scholar] [CrossRef]
- Chen, Y.N.; Slabaugh, E.; Brandizzi, F. Membrane-tethered transcription factors in Arabidopsis thaliana: Novel regulators in stress response and development. Curr. Opin. Plant Biol. 2008, 11, 695–701. [Google Scholar] [CrossRef]
- Li, P.; Wind, J.J.; Shi, X.; Zhang, H.; Hanson, J.; Smeekens, S.C.; Teng, S. Fructose sensitivity is suppressed in Arabidopsis by the transcription factor ANAC089 lacking the membrane-bound domain. Proc. Natl. Acad. Sci. USA 2011, 108, 3436–3441. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K. Genomics, molecular and evolutionary perspective of NAC transcription factors. PLoS ONE 2020, 15, e0231425. [Google Scholar] [CrossRef]
- Melo, B.P.; Loureco-Tessutti, I.T.; Teixeira, O.; Bezerra, L.; Lins, C.B.; Morgantes, C.V.; Engler, J.A.; Braga, P.A.; Grossi-de-Sá, M.F.; Batista, E.P. Contrasting roles of GmNAC065 and GmNAC085 in natural senescence, plant development, multiple stresses and cell death responses. Sci. Rep. 2021, 11, 11178. [Google Scholar] [CrossRef]
- Yan, J.; Chen, Q.; Cui, X.; Zhao, P.; Gao, S.; Yang, B.; Liu, J.-X.; Tong, T.; Deyholos, K.; Jiang, Y.-Q. Ectopic overexpression of a membrane-tethered transcription factor gene NAC60 from oilseed rape positively modulates programmed cell death and age-triggered leaf senescence. Plant J. 2021, 105, 600–618. [Google Scholar] [CrossRef]
- Fu, C.; Liu, M. Genome-wide identification and molecular evolution of NAC gene family in Dendrobium nobile. Front. Plant Sci. 2023, 14, 1232804. [Google Scholar] [CrossRef]
- Faria, J.A.; Reis, P.; Reis, M.; Rosado, G.L.; Pinheiro, G.L.; Mendes, G.C.; Fontes, E. The NAC domain-containing protein, GmNAC6, is a downstream component of the ER stress-and osmotic stress-induced NRP-mediated cell-death signaling pathway. BMC Plant Biol. 2011, 11, 129. [Google Scholar] [CrossRef]
- Penfold, C.A.; Buchanan-Wollaston, V. Modelling transcriptional networks in leaf senescence. J. Exp. Bot. 2014, 65, 3859–3873. [Google Scholar] [CrossRef]
- Podzimska-Sroka, D.; O’Shea, C.; Gregersen, P.L.; Skriver, K. NAC transcription factors in senescence: From molecular structure to function in crops. Plants 2015, 4, 412–448. [Google Scholar] [CrossRef]
- Kim, H.J.; Nam, H.G.; Lim, P.O. Regulatory network of NAC transcription factors in leaf senescence. Curr. Opin. Plant Biol. 2016, 33, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, J.-H.; Kim, J.; Kim, J.J.; Hong, S.; Kim, J.; Kim, J.H.; Woo, H.R.; Hyeon, C.; Lim, P.O.; et al. Time-evolving genetic networks reveal a NAC troika that negatively regulates leaf senescence in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 4930–4939. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.M.; Zhang, Y.J.; Turečková, V.; Xue, G.P.; Fernie, A.R.; Mueller-Roeber, B.; Balazadeh, S. The NAC transcription factor SlNAP2 regulates leaf senescence and fruit yield in tomato. Plant Physiol. 2018, 177, 1286–1302. [Google Scholar] [CrossRef] [PubMed]
- Bengoa-Luoni, S.; Astigueta, F.H.; Nicosia, S.; Moschen, S.; Fernández, P.; Heinz, R. Transcription factors associated with leaf senescence in crops. Plants 2019, 8, 411. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Upadyay, A.K.; Biradar, H.; Hittalmani, S. OsNAC-like transcription factor involved in regulating seed-storage protein content at different stages of grain filling in rice under aerobic conditions. J. Genet. 2019, 98, 18. [Google Scholar] [CrossRef]
- Yang, T.; Guo, L.; Ji, C.; Wang, H.; Wang, J.; Zheng, X.; Xiao, Q.; Wu, Y. The B3 domain-containing transcription factor ZmABI19 coordinates expression of key factors required for maize seed development and grain filling. Plant Cell. 2021, 33, 104–128. [Google Scholar] [CrossRef] [PubMed]
- Nagahage, I.S.P.; Matsuda, K.; Miyashita, K.; Fujiwara, S.; Mannapperuma, C.; Yamada, T.; Sakamoto, S.; Ishikawa, T.; Nagano, M.; Ohtani, M.; et al. NAC domain transcription factors VNI2 and ATAF2 form protein complexes and regulate leaf senescence. Plant Direct. 2023, 7, e529. [Google Scholar] [CrossRef]
- Wang, L.; Tian, T.; Liang, J.; Li, R.; Xin, X.; Qi, Y.; Zhou, Y.; Fan, Q.; Ning, G.; Becana, M.; et al. A transcription factor of the NAC family regulates nitrate-induced legume nodule senescence. New Phytol. 2023, 238, 2113–2129. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; Ye, Z.H. Global analysis of direct targets of secondary wall NAC master switches in Arabidopsis. Mol. Plant 2010, 3, 1087–1103. [Google Scholar] [CrossRef]
- Li, E.; Bhargava, A.; Qiang, W.; Friedman, M.C.; Forneris, N.; Savidge, R.A.; Johnson, L.A.; Mansfield, S.D.; Ellis, B.E.; Douglas, C.J. The class II KNOX gene KNAT7 negatively regulates secondary wall formation in Arabidopsis and is functionally conserved in Populus. New Phytol. 2012, 194, 102–115. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, J.; Xu, P.; Zhang, R.; Li, L.G. Intron-mediated alternative splicing of wood-associated NAC transcription factor1b regulates cell wall thickening during fiber development in populus species. Plant Physiol. 2014, 164, 765–776. [Google Scholar] [CrossRef]
- Negi, S.; Tak, H.; Ganapathi, T.R. Native vascular related NAC transcription factors are efficient regulator of multiple classes of secondary wall associated genes in banana. Plant Sci. 2017, 265, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, G.Q.; Zou, D.; Yan, J.Q.; Li, Y.; Hu, S.; Li, X.B. The cotton (Gossypium hirsutum) NAC transcription factor (FSN1) as a positive regulator participates in controlling secondary cell wall biosynthesis and modification of fibers. New Phytol. 2018, 217, 625–640. [Google Scholar] [CrossRef]
- Zhang, Q.; Luo, F.; Zhong, Y.; He, J.; Li, L. Modulation of NAC transcription factor NST1 activity by XYLEM NAC DOMAIN1 regulates secondary cell wall formation in Arabidopsis. J. Exp. Bot. 2020, 71, 1449–1458. [Google Scholar] [CrossRef]
- Fang, Y.; Liao, K.; Du, H.; Xu, Y.; Song, H.; Li, X.; Xiong, L. A stress responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2015, 66, 6803–6817. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.-B.; Pan, X.-X.; Jiang, H.-W.; Wu, G.-J. Comparison of the starch synthesis genes between maize and rice: Copies, chromosome location andexpression divergence. Theor. Apple Genet. 2009, 119, 815–825. [Google Scholar] [CrossRef]
- Yan, J.; Tong, T.; Li, X.; Chen, Q.; Dai, M.; Niu, F.; Yang, M.; Deyholos, M.K.; Yang, B.; Jiang, Y.-Q. A Novel NAC-type transcription factor, NAC87, from oilseed rape modulates reactive oxygen species accumulation and cell death. Plant Cell Physiol. 2018, 59, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fengler, K.A.; Hemert, J.L.; Gupta, R.; Gupta, R.; Mongar, N.; Sun, J.; Allen, W.B.; Wang, Y.; Weers, B.; et al. Identification and characterization of a novel stay-green QTL that increases yield in maize. Plant Biotechnol. J. 2019, 17, 2272–2285. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Ohtani, M.; Mitsuda, N.; Kubo, M.; Ohme-Takagi, M.; Fukuda, H.; Demura, T. VND-INTERACTING2, a NAC domain transcription factor, negatively regulates xylem vessel formation in Arabidopsis. Plant Cell 2010, 22, 1249–1263. [Google Scholar] [CrossRef]
- Furuta, K.M.; Furuta, K.M.; Yadav, S.R.; Lehesranta, S.; Belevich, I.; Miyashima, S.; Heo, J.; Vatén, A.; Lindgren, O.; Rybel, B.; et al. Plant development. Arabidopsis NAC45/86 direct sieve element morphogenesis culminating in enucleation. Science 2014, 345, 933–937. [Google Scholar] [CrossRef]
- Matilla, A.J. The Interplay between enucleated sieve elements and companion cells. Plants 2023, 12, 3033. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Maruyama, K.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Tran, L.-S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J. 2004, 39, 863–876. [Google Scholar] [CrossRef]
- Christianson, J.A.; Wilson, L.W.; Llewellyn, D.J.; Dennis, E.S. The Low-oxygen-induced NAC domain transcription factor ANAC 102 affects viability of Arabidopsis seeds following low-oxygen treatment. Plant Physiol. 2009, 149, 1724–1738. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, L.N.; Zhang, Z. The role of major transcription factors in solanaceous food crops under different stress conditions: Current and future perspectives. Plants 2020, 9, 56. [Google Scholar] [CrossRef]
- Zhao, Y.; Antoniou-Kourounioti, R.L.; Calder, G.; Dean, C.; Howard, M. Temperature-dependent growth contributes to long-term cold sensing. Nature 2020, 583, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Alshareef, N.O.; Otterbach, S.L.; Allu, A.D.; Woo, Y.H.; Werk, T.; Kamranfar, I.; Mueller-Roeber, B.; Tester, M.; Balazadeh, S.; Schmöckel, S.M. NAC transcription factors ATAF1 and ANAC055 affect the heat stress response in Arabidopsis. Sci. Rep. 2022, 12, 11264. [Google Scholar] [CrossRef]
- Xu, Y.; Li, P.; Ma, F.; Huang, D.; Xing, W.; Wu, B.; Sun, P.; Xu, B.; Song, S. Characterization of the NAC transcription factor in passion fruit (Passiflora edulis) and functional identification of PeNAC-19 in cold stress. Plants 2023, 12, 1393. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, S.; Zhang, B.; Shang-Guan, K.; Shi, Y.; Zhang, D.; Liu, X.; Wu, K.; Xu, Z.; Fu, X.; et al. A gibberellin-mediated DELLA-NAC signaling cascade regulates cellulose synthesis in rice. Plant Cell 2015, 27, 1681–1696. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yang, Z.T.; Song, Z.T.; Wang, M.J.; Liu, J.X. The plant-specifc transcription factor NAC103 is induced by bZIP60 through a new cis-regulatory element to modulate the unfolded protein response in Arabidopsis. Plant J. 2013, 76, 274–286. [Google Scholar] [CrossRef]
- Ryu, T.H.; Go, Y.S.; Choi, S.H.; Kim, J.I.; Chung, B.Y.; Kim, J.H. SOG1-dependent NAC103 modulates the DNA damage response as a transcriptional regulator in Arabidopsis. Plant J. 2019, 98, 83–96. [Google Scholar] [CrossRef]
- Yang, Z.; Fan, S.; Wang, J.; An, Y.; Guo, Z.; Li, K.; Liu, X. The plasma membrane-associated transcription factor NAC091 regulates unfolded protein response in Arabidopsis thaliana. Plant Sci. 2023, 334, 111777. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.O.; Fraga, O.T.; Pimenta, M.R.; Caetano, H.D.; Machado, J.P.B.; Carpinetti, P.A.; Brustolini, O.J.B.; Quadros, I.P.S.; Reis, P.A.B.; Fontes, E.P. GmNAC81 inversely modulates leaf senescence and drought tolerance. Front. Genet. 2020, 11, 601876. [Google Scholar] [CrossRef] [PubMed]
- Wleklik, K.; Borek, S. Vacuolar processing enzymes in plant programmed cell death and autophagy. Int. J. Mol. Sci. 2023, 24, 1198. [Google Scholar] [CrossRef] [PubMed]
- Mendes, G.C.; Reis, P.A.; Calil, I.P.; Carvalho, H.H.; Aragao, F.J.; Fontes, E.P. GmNAC30 and GmNAC81 integrate the endoplasmic reticulum stress-and osmotic stress-induced cell death responses through a vacuolar processing enzyme. Proc. Natl. Acad. Sci. USA 2013, 110, 19627–19632. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Liu, H.; Tan, S.; Li, Z. Transcription factors-regulated leaf senescence: Current knowledge, challenges and approaches. Int. J. Mol. Sci. 2023, 24, 9245. [Google Scholar] [CrossRef] [PubMed]
- Rauf, M.; Arif, M.; Dortay, H.; Matallana, L.P. ORE1 balances leaf senescence against maintenance by antagonizing G2-like-mediated transcription. EMBO Rep. 2013, 14, 382–388. [Google Scholar] [CrossRef]
- Kim, H.J.; Hong, S.H.; Kim, Y.W.; Lee, H.; Jun, J.H.; Phee, B.-K.; Rupak, T.; Jeon, H.; Lee, Y.; Hong, B.S.; et al. Gene regulatory cascade of senescence-associated NAC transcription factors activated by ETHYLENE-INSENSITIVE2-mediated leaf senescence signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 4023–4036. [Google Scholar] [CrossRef] [PubMed]
- Qiu, K.; Li, Z.; Yang, Z.; Chen, J.; Wu, S.; Zhu, S.; Gao, S.; Gao, J.; Ren, G.; Kuai, B.; et al. EIN3 and ORE1 accelerate de-greening during ethylene-mediated leaf senescence by directly activating chlorophyll catabolic genes in Arabidopsis. PLoS Gene 2015, 11, 1005–1399. [Google Scholar]
- Matallana-Ramírez, L.P.; Rauf, M.; Farage-Barhom, S.; Dortay, H.; Xue, G.-P.; Dröge-Laser, W.; Balazedeh, S.; Mueller-Roeber, B. NAC transcription factor ORE1 and senescence-induced BIFUNCTIONAL NUCLEASE1 (BFN1) constitute a regulatory cascade in Arabidopsis. Mol. Plant 2013, 6, 1438–1452. [Google Scholar] [CrossRef]
- La, V.H.; Nguyen, T.; Ngo, X.B.; Tran, D.; Khuat, H.T.; Bui, T.T.; Tran, T.T.H.; Chung, Y.S.; Nguyen, T.D. At-ore1 gene induces distinct novel H2O2-NACs signaling in regulating the leaf senescence in soybeans (Glycine max L.). Plants 2022, 12, 2110. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Zhang, X.; Lv, B.; Luo, L.; Xi, D.; Shen, J.; Ma, H.; Lu, F.M. OsNAC2 encoding a NAC transcription factor that affects plant height through mediating the gibberellic acid pathway in rice. Plant J. 2015, 82, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Lu, S.; Lv, B.; Zhang, B.; Shen, J.; He, J.; Luo, L.; Xi, D.; Chen, X.; Ming, F. A rice NAC transcription factor promotes leaf senescence via ABA biosynthesis. Plant Physiol. 2017, 174, 1747–1763. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Ding, J.; Zhang, B.; Xi, D.; Ming, F. OsNAC2 positively affects salt-induced cell death and binds to the OsAP37 and OsCOX11 promoters. Plant J. 2018, 94, 454–468. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.B.; Lv, B.; Luo, L.Q.; He, J.M.; Mao, C.J.; Xi, D.D. The MF. NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 7, 40641. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; He, J.; Liu, L.; Deng, Q.; Yao, X.; Liu, C.; Qiao, Y.; Li, P.; Ming, F. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol. J. 2020, 18, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Redillas, M.; Jeong, J.S.; Kim, Y.S.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.-H.; Reuzeau, C.; Kim, J.-K. The overexpression of OsNAC9 alters the root architecture of rice plants enhancing drought resistance and grain yield under field conditions. Plant Biotechnol. J. 2012, 10, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.-H.; Choi, Y.D.; Kim, M.; Reuzeau, C.; Kim, J.-K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Hernández, B.Y.; Núñez-Muñoz, L.; Calderón-Pérz, B.; Xoconostle-Cáceres, B.; Ruiz-Medrano, R. The NAC transcription factor ANAC087 induces aerial rosette development and leaf senescence in Arabidopsis. Front. Plant Sci. 2022, 13, 818107. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhong, Z.; Zhang, H.; Wang, X.; Liu, B.; Yang, L.; Han, X.; Yu, D.; Zheng, X.; Wang, C.; et al. Targeted mutagenesis of NAC transcription factor gene, OsNAC041, leading to salt sensitivity in rice. Rice Sci. 2019, 26, 98–108. [Google Scholar]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef]
- Forlani, S.; Mizzotti, C.; Masiero, S. The NAC side of the fruit: Tuning of fruit development and maturation. BMC Plant Biol. 2021, 21, 238. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.-H.; Zhang, Y.-J.; Yao, J.-L.; Xie, Z.-H.; Zhang, Y.-Y.; Zhang, S.-L.; Gu, C. The NAM/ATAF1/2/CUC2 transcription factor PpNAC.A59 enhances PpERF.A16 expression to promote ethylene biosynthesis during peach fruit ripening. Hortic. Res. 2021, 8, 209. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.H.; Zhou, J.Q.; Wu, C.E.; Yang, S.; Liu, Y.F.; Chai, L.P.; Xue, Z.H. The interplay between ABA/ethylene and NAC TFs in tomato fruit ripening: A review. Plant Mol. Biol. 2021, 106, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Migikovsky, Z.; Yeats, T.H.; Watts, S.; Song, J.; Forney, C.F.; Burgher-MacLellan, K.; Somers, D.J.; Gong, Y.; Zhang, Z.; Vrebalov, J.; et al. Apple ripening is controlled by a NAC transcription factor. Front. Genet. 2021, 12, 671300. [Google Scholar] [CrossRef]
- Zhang, R.-X.; Liu, Y.; Zhang, X.; Chen, X.; Sun, J.; Zhao, Y.; Zhang, J.; Yao, J.-L.; Liao, L. Two adjacent NAC transcription factors regulate fruit maturity date and flavor in peach. New Phytol. 2024, 241, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Cao, H.; Cui, L.; Wang, Y.; Wei, L.; Gent, S.; Yang, L.; Huang, Y.; Bie, Z. CmoNAC1 in pumpkin rootstocks improves salt tolerance of grafted cucumbers by binding to the promoters of CmoRBOHD1, CmoNECD6, CmoAKT1; 2 and CmoHKT1;1 to regulated H2O2, ABA signaling and K+/Na+ homeostasis. Hort. Res. 2023, 10, uhad157. [Google Scholar] [CrossRef]
- Hu, Y.; Li, C.; Zhou, R.; Song, Y.; Lv, Z.; Wang, Q.; Dong, X.; Liu, S.; Feng, C.; Zeng, X.; et al. The transcription factor ZmNAC89 gene is involved in salt tolerance in maize (Zea mays L.). Int. J. Mol. Sci. 2023, 24, 15099. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Ying, S.; Zhang, D.-F.; Shi, Y.-S.; Song, Y.-C.; Wang, T.-Y.; Li, Y. A maize stress-responsive NAC transcription factor, ZmSNAC1, confers enhanced tolerance to dehydration in transgenic Arabidopsis. Plant Cell Rep. 2012, 31, 1701–1711. [Google Scholar] [CrossRef]
- Mao, H.; Yu, L.; Han, R.; Li, Z.; Liu, H. ZmNAC55, a maize stress-responsive NAC transcription factor, confers drought resistance in transgenic Arabidopsis. Plant Physiol. Biochem. 2016, 105, 55–66. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, B.-G.; Chao, Q.; Wang, B.; Zhang, Q.; Zhang, C.; Li, C.; Jin, F.; Yang, F.; Li, X. Function analysis of ZmNAC33, a positive regulator in drought stress response in Arabidopsis. Plant Physiol. Biochem. 2019, 145, 174–183. [Google Scholar] [CrossRef]
- Cao, H.S. Ectopic expression of pumpkin NAC transcription factor CmNAC1 improves multiple abiotic stress tolerance in Arabidopsis. Front. Plant Sci. 2017, 8, 2052. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.L.; Yue, X.F.; Min, Z.; Wang, X.H.; Fang, Y.L.; Zhang, J.X. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC tran-scription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 146, 98–111. [Google Scholar] [CrossRef]
- Mao, H.; Wang, H.; Liu, S.; Yang, X.; Yan, J.; Li, J.; Phan Tran, L.-S.; Qin, F. A transposable element in a NAC gene is associated with drought tolerance in maiz seedlings. Nat. Commun. 2015, 6, 8326. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yan, J.; Liu, W.; Shen, L.; Sheng, Y.; Sun, Y.; Li, Y.; Scheller, H.V.; Jiang, M.; Hou, X.; et al. Phosphorylation of a NAC transcription factor by a calcium/calmodulin-dependent protein kinase regulates abscisic acid-induced antioxidant defense in maize. Plant Physiol. 2016, 171, 1651–1664. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Yan, J.; Xiang, Y.; Zhang, A. Phosphorylation of ZmNAC84 at Ser-113 enhances the rought tolerance by directly modulating ZmSOD2 expression in maize. Biochem. Biophys. Res. Commun. 2021, 567, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought torerance in transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 268. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef]
- Xue, G.P.; Way, H.M.; Richardson, T.; Drenth, J.; Joyce, P.A.; McIntyre, C.L. Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-regulated genes and dehydration tolerance in bread wheat. Mol. Plant 2011, 4, 697–712. [Google Scholar] [CrossRef]
- Zhang, X.; Li, L.; Lang, Z.; Li, D.; He, Y.; Zhao, Y.; Tao, H.; Wei, J.; Li, Q.; Hong, G. Genome-wide characterization of NAC transcription factors in Camellia sinensis and the involvement of CsNAC28 in drought tolerance. Front. Plant Sci. 2022, 13, 1065261. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Borrill, P.; Harrington, S.A.; Uauy, C. Genome-wide sequence and expression analysis of the NAC transcription factor family in polyploid wheat. G3 Genesgenet. 2017, 7, 3019–3029. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchishinozaki, K.; Shinozaki, K.; Tran, L.S.P. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res. 2011, 18, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Shiriga, K.; Sharma, R.; Kumar, K.; Yadav, S.K.; Hossain, F.; Thirunavukkarasu, N. Genome-wide identification and expression pattern of drought-responsive members of the NAC family in maize. Meta Gene 2014, 2, 407–417. [Google Scholar] [CrossRef]
- Singh, A.K.; Sharma, V.; Pal, A.K.; Acharva, V.; Ahuja, P.S. Genome-wide organization and expression profiling of the NAC transcription factor family in potato (Solanum tuberosum L.). DNA Res. 2013, 20, 403–423. [Google Scholar] [CrossRef]
- Hu, W.; Wei, Y.; Xia, Z.; Yan, Y.; Hou, X.; Zou, M.; Lu, C.; Wang, W.; Peng, M. Genome-wide identification and expression analysis of the NAC transcription factor family in cassava. PLoS ONE 2015, 10, e0136993. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, F.; Li, M.Y.; Jiang, Q.; Tan, G.F.; Xiong, A.S. Genome wide analysis of the NAC transcription factor family in Chinese cabbage to elucidate responses to temperature stress. Sci. Hortic. 2014, 165, 82–90. [Google Scholar] [CrossRef]
- Diao, W.; Snyder, J.C.; Wang, S.; Liu, J.; Pan, B.; Guo, G.; Ge, W.; Dawood, M. Genome-wide analyses of the NAC transcription factor gene family in pepper (Capsicum annuum L.): Chromosome location, phylogeny, structure, expression patterns, cis-elements in the promoter, and interaction network. Int. J. Mol. Sci. 2018, 19, 1048–1061. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Gao, L.; Zhang, Y.; Zhang, F.; Yang, X.; Huang, D. Genome-wide investigation of the NAC transcription factor family in melon (Cucumis melo L.) and their expression analysis under salt stress. Plant Cell Rep. 2016, 35, 1827–1839. [Google Scholar] [CrossRef]
- Su, H.; Zhang, S.; Yuan, X.; Chen, C.; Wang, X.F.; Hao, Y.J. Genome-wide analysis and identification of stress-responsive genes of the NAM–ATAF1,2–CUC2 transcription factor family in apple. Plant Physiol. Biochem. 2013, 71, 11–21. [Google Scholar] [CrossRef]
- Bengoa Luoni, S.A.; Cenci, A.; Moschen, S.; Nicosia, S.; Radonic, L.M.; García, J.S.; Langlade, L.B.; Vile, D.; Rovere, C.; Fernández, P. Genome-wide and comparative phylogenetic analysis of senescence-associated NAC transcription factors in sunflower (Helianthus annuus). BMC Genom. 2021, 22, 893. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Quian, Z.; Wang, T.; Zhao, X.; Zhao, X.; Gu, S.; Rao, X.; Lyu, S.; Zhang, R.; He, L.; et al. Genome-wide identification and expression analysis of the NAC transcription factor family in Saccharum spontaneum under different stresses. Plant Signal. Behav. 2022, 17, 2088665. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Ma, L.; Chen, X.; Fei, X.; He, B.; Luo, Y.; Wei, A. Genome-wide identification of the NAC gene family in Zanthoxylum bungeanum and their transcriptional responses to drought stress. Int. J. Mol. Sci. 2022, 23, 4769. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, J.; Zhang, Q.; Dong, A.; Wu, Q.; Zhu, X.; Zhu, X. Genome-wide identification and analysis of the NAC transcription factor gene family in garden asparagus (Asparagus officinalis). Genes 2022, 13, 976. [Google Scholar] [CrossRef]
- He, J.; He, X.; Chang, P.; Jiang, H.Z.; Gong, D.P.; Sun, Q. Genome-wide identification and characterization of TCP family genes in Brassica juncea var. Tumida. Peer J. 2020, 8, e9130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ni, L.; Liu, D.; Fu, Z.; Hua, J.; Lu, Z.; Liu, Z.; Yin, Y.; Li, H.; Gu, C. Genome-wide identification and characterization of NAC family in Hibiscus hamabo Sieb. et Zucc. under various abiotic stresses. Int. J. Mol. Sci. 2022, 23, 3055. [Google Scholar] [CrossRef]
- Sun, M.M.; Liu, X.; Huang, X.-J.; Yang, J.-J.; Qin, P.-T.; Zhou, H.; Jiang, M.-G.; Liao, H.-Z. Genome-wide identification and expression analysis of the NAC gene family in Kandelia obovata, a typical mangrove plant. Curr. Issues Mol. Biol. 2022, 44, 5622–5637. [Google Scholar] [CrossRef] [PubMed]
- Shahnejat-Bushehri, S.; Tarkowska, D.; Sakuraba, Y.; Balazadeh, S. Arabidopsis NAC transcription factor JUB1 regulates GA/BR metabolism and signalling. Nat. Plants 2016, 2, 16013. [Google Scholar] [CrossRef]
- Hu, R.; Qi, G.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145–167. [Google Scholar] [CrossRef]
- Wang, N.; Zheng, Y.; Xin, H.; Fang, L.; Li, S. Comprehensive analysis of NAC domain transcription factor gene family in Vitis vinifera. Plant Cell Rep. 2013, 32, 61–75. [Google Scholar] [CrossRef]
- Rushton, P.J.; Bokowiec, M.T.; Han, S.; Zhang, H.; Brannpck, J.F.; Chen, X.; Laudeman, T.W.; Timbo, M.P. Tobacco transcription factors: Novel insights into transcriptional regulation in the Solanaceae. Plant Physiol. 2008, 147, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Song, L.; Wang, Y.; Guo, C. Genome-wide analysis and expression patterns of the NAC transcription factor family in Medicago truncatula. Physiol. Mol. Biol. Plants 2017, 23, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Song, X.; Duan, W.; Huang, Z.; Liu, G.; Li, Y.; Hou, X. Genome-wide analysis and expression patterns of NAC transcription factor family under different developmental stages and abiotic atresses in chinese cabbage (Brassica rapa). Plant Mol. Biol. Rep. 2014, 32, 1041–1056. [Google Scholar] [CrossRef]
- Yang, T.; Wang, H.; Guo, L.; Wu, X.; Xiao, Q.; Wang, J.; Wang, Q.; Ma, G.; Wang, W.; Wu, Y. ABA-induced phosphorylation of basic leucine zipper 29, ABSCISIC ACID INSENSITIVE-19 and opaque-2 by SnRK2.2 enhances gene transactivation for endosperm filling in maize. Plant Cell. 2022, 34, 1933–1956. [Google Scholar] [CrossRef]
- Bu, Q.; Jiang, H.; Li, C.B.; Zhai, Q.; Zhang, J.; Wu, X.; Sun, J.; Xie, Q.; Li, C. Role of the Arabidopsis thaliana NAC transcription factors ANAC019 and ANAC055 in regulating jasmonic acid-signaled defense responses. Cell Res. 2008, 18, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, L.; Cao, Y.; Qi, C.; Li, S.; Liu, L.; Wang, G.; Mao, A.; Ren, S.; Guo, Y.-D. CsATAF1 positively regulates drought stress tolerance by ABA dependent pathway and promoting ROS scavenging in cucumber. Plant Cell Physiol. 2018, 59, 930–945. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Zhang, J.; Ren, Y.; Li, M.; Tian, S.; Yu, Y.; Zuo, Y.; Gong, G.; Zhang, H.; et al. The NAC transcription factor ClNAC68 positively regulates sugar content and seed development in watermelon by repressing ClINV and ClGH3.6. Hort. Res. 2021, 8, 214. [Google Scholar] [CrossRef]
- Sotta, N.; Sakamoto, T.; Kamiya, T.; Tabata, R.; Yamaguchi, K.; Shigenobu, S.; Yamada, M.; Hasebe, M.; Sawa, S.; Fujiwara, T. NAC103 mutation alleviates DNA damage in an Arabidopsis thaliana mutant sensitive to excess boron. Front. Plant Sci. 2023, 14, 1099816. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, Z.; Zhu, W.; Wang, N.; Bai, M.; Kuang, H.; Cai, C.; Zhong, X.; Kong, F.; Lü, P.; et al. The NAC transcription factors SNAP1/2/3/4 are central regulators mediating high nitrogen responses in mature nodules of soybean. Nat. Commun. 2023, 14, 4711. [Google Scholar] [CrossRef]
- Li, W.; Zeng, Y.; Yin, F.; Wei, R.; Mao, X. Genome-wide identification and comprehensive analysis of the NAC transcription factor family in sunflower during salt and drought stress. Sci. Rep. 2021, 11, 19865. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell Physiol. 2014, 55, 604–619. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.; Kang, K.; Shim, Y.; Sakuraba, Y.; An, G.; Paek, N.-C. Rice ETHYLENE RESPONSE FACTOR 101 promotes leaf senescence through jasmonic acid-mediated regulation of OsNAP and OsMYC2. Front. Plant Sci. 2020, 11, 587620. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Sugahara, S.; Yamada, T.; Kikuchi, K.; Yoshiba, Y.; Hirano, H.Y.; Tsutsumi, N. OsNAC6, a member of the NAC gene family, is induced by various stresses in rice. Genes Genet. Syst. 2005, 80, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, L.; Tian, X.; Liu, S.; Xu, D.; Jin, H.; Song, J.; Dong, Y.; Zhao, D.; Li, G.; et al. TaNAC100 acts as an integrator of seed protein and starch synthesis exerting pleiotropic effects of agronomic traits in wheat. Plant J. 2021, 108, 829–840. [Google Scholar] [CrossRef]
- Gao, Y.; An, K.; Guo, W.; Chen, Y.; Zhang, R.; Zhang, X.; Chang, S.; Rossi, S.; Jin, F.; Cao, X.; et al. The endosperm-specific transcription factor TaNAC019 regulates glutenin and starch accumulation and its elite allele improves wheat grain quality. Plant Cell 2021, 33, 603–622. [Google Scholar] [CrossRef] [PubMed]
- Sen, L.; Luo, G.; Song, Y.; Xu, J.; Ji, J.; Zhang, C.; Gregová, E.; Yang, W.; Li, X.; Sun, J.; et al. A novel NAC family transcription factor SPR suppresses seed storage protein synthesis in wheat. Plant Biotechnol. J. 2012, 19, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Grimault, A.; Gendrot, G.; Chamot, S.; Widiez, T.; Rabillé, H.; Gérentes, M.-F.; Creff, A.; Thévenin, J.; Dubreucq, B.; Ingran, G.C.; et al. ZmZHOUPI, an endosperm-specific basic helix-loop-helix transcription factor involved in maize see development. Plant J. 2015, 84, 574–586. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Yi, Q.; Hu, Y.; Liu, H.; Liu, Y.; Huang, Y. Novel role of ZmaNAC36 in co-expression of starch synthetic genes in maize endosperm. Plant Mol. Biol. 2014, 84, 359–369. [Google Scholar] [CrossRef]
- Dong, Q.; Wang, F.; Kong, J.; Xu, Q.; Li, T.; Chen, L.; Chen, H.; Jiang, H.; Li, C.; Cheng, B. Functional analysis of ZmMADS1a reveals its role in regulating starch biosynthesis in maize endosperm. Sci. Rep. 2019, 9, 3253. [Google Scholar] [CrossRef]
- Chen, E.; Yu, H.; He, J.; Peng, D.; Zhu, P.; Pan, S.; Wu, X.; Wang, J.; Ji, C.; Chao, Z.; et al. The transcription factors ZmNAC128 and ZmNAC130 coordinate with Opaque2 to promote endosperm filling in maize. Plant Cell 2023, 35, 4066–4090. [Google Scholar] [CrossRef]
- Hoth, S.; Morgante, M.; Sanchez, J.P.; Hanafey, M.K.; Tingey, S.V.; Chua, N.H. Genome-wide gene expression profiling in Arabidopsis thaliana reveals new targets of abscisic acid and largely impaired gene regulation in the abi1-1 mutant. J. Cell Sci. 2002, 115, 4891–4900. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Guo, H.-S.; Dallman, G.; Fang, S.; Weissman, A.M.; Chua, N.-H. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature 2002, 419, 167–170. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, Q.; Fang, L.; Yang, W.; Xu, F.; Yan, Y.; Mao, R. Comprehensive analysis of NAC transcription factors in Scutellaria baicalensis and their response to exogenous ABA and GA3. Int. J. Biol. Macr. 2023, 244, 125290. [Google Scholar] [CrossRef] [PubMed]
- Sperotto, R.; Ricachenevsky, F.K.; Duarte, G.L.; Boff, T.; Lopes, K.L.; Sperb, E.R.; Grusal, M.A.; Fett, J.P. Identification of up-regulated genes in flag leaves during rice grain filling and characterization of OsNAC5, a new ABA-dependent transcription factor. Planta 2009, 230, 985–1002. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.K.; Lindemose, S.; Masi, F.; Reimer, J.J.; Nielsen, M.; Perera, V.; Workman, C.T.; Turk, F.; Grant, M.R.; Mundy, J.; et al. ATAF1 transcription factor directly regulates abscisic acid biosynthetic gene NCED3 in Arabidopsis thaliana. FEBS Open Bio 2013, 3, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Kim, D.; Han, S.H.; Kim, S.H.; Piao, W.; Yanagisawa, S.; An, G.; Paek, N.C. Multilayered regulation of membrane-bound ONAC054 is essential for abscisic acid-induced leaf senescence in rice. Plant Cell 2020, 32, 630–649. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Paek, N.C. Overexpression of ONAC054 improves drought stress tolerance and grain yield in rice. Crops 2022, 2, 390–406. [Google Scholar] [CrossRef]
- Xu, B.; Li, H.; Li, Y.; Yu, G.; Zhang, J.; Huang, B. Characterization and transcriptional regulation of chlorophyll-b reductase gene NON-YELLOW COLORING 1 associated with leaf senescence in perennial ryegrass (Lolium perenne L.). Environ. Exp. Bot. 2018, 149, 43–50. [Google Scholar] [CrossRef]
- Breeze, E. Make, modify, move: Multilayered regulation of ONAC054 during ABA-induced leaf senescence. Plant Cell 2020, 32, 532–533. [Google Scholar] [CrossRef]
- Pooam, M.; El-Ballat, E.M.; Jourdan, N.; Ali, H.M.; Hano, C.; Ahmad, M.; El-Esawi, M.A. SNAC3 transcription factor enhances arsenic stress tolerance and grain yield in rice (Oryza sativa L.) through regulating physio-biochemical mechanisms, stress-responsive genes, and cryptochrome-1b. Plants 2023, 12, 2731. [Google Scholar] [CrossRef]
- Chico, J.M.; Lechner, E.; Fernández-Barbero, E.; Canibano, E.; García-Casado, G.; Franco-Zorrilla, J.M.; Hamman, P.; Zamarreño, A.M.; García-Mina, J.M.; Rubio, V.; et al. CUL3BPM E3 ubiquitin ligases regulate MYC2, MYC3, and MYC4 stability and JA responses. Proc. Natl. Acad. Sci. USA 2020, 117, 6205–6215. [Google Scholar] [CrossRef]
- Jiménez-Ibáñez, S.; Boter, M.; Ortigosa, A.; García-Casado, G.; Chini, A.; Lewsey, M.G.; Ecker, J.R.; Ntoukakis, V.; Solano, R. JAZ2 controls stomata dynamics during bacterial invasion. New Phytol. 2017, 213, 1378–1392. [Google Scholar]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress-1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.B.; Li, X.Y.; Liu, X.; Li, X.L.; Li, Y.H.; Li, L. Study on the response to applying ABA and expression changes of ABA induced relative genes in Arabidopsis mutants of three NAC homologous genes. Life Sci. Res. 2015, 19, 114–118. [Google Scholar]
- Hickman, R.; Hill, C.; Penfold, C.A.; Bree, E.; Bowden, L.; Moore, J.D.; Zhang, P.; Jackson, A.; Cooke, E.; Bewicke-Copley, F.; et al. A local regulatory network around three NAC transcription factors in stress responses and senescence in Arabidopsis leaves. Plant J. 2013, 75, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, X.; Li, M.; Yan, Y.; Liu, X.; Li, L. Dual function of NAC072 in ABF3-mediated ABA-responsive gene regulation in Arabidopsis. Front. Plant Sci. 2016, 7, 1075. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Liew, L.C.; Yin, L.; Lewsey, M.G.; Whelan, J.; Berkowitz, O. The retrograde signaling regulator ANAC017 recruits the MKK9–MPK3/6, ethylene, and auxin signaling pathways to balance mitochondrial dysfunction with growth. Plant Cell 2022, 34, 3460–3481. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Yang, H.; Xiang, L.; Wang, Y.; Chan, Z. NAC transcription factor TgNAP promotes tulip petal senescence. Plant Physiol. 2022, 190, 1960–1977. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.Y.; Spivey, N.W.; Zeng, W.; Liu, P.P.; Fu, Z.Q.; Klessig, D.F.; He, S.Y.; Dong, X. Coronatine promotes Pseudomonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulation. Cell Host Microbe 2012, 11, 587–596. [Google Scholar] [CrossRef]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. An apple NAC transcription factor enhances salt stress tolerance by modulating the ethylene response. Physiol. Plant. 2018, 164, 279–289. [Google Scholar] [CrossRef]
- Chung, P.J.; Jung, H.; Choi, Y.D.; Kim, J.K. Genome-wide analyses of direct target genes of four rice NAC-domain transcription factors involved in drought tolerance. BMC Genom. 2018, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.Y.; Wang, S.; Zhang, F.; Chen, F.; Chen, L.; Hao, X.; Pan, Q.; Fu, X.; Li, L.; Su, S.; et al. Overexpression of a novel NAC domain-containing transcription factor gene (AaNAC1) enhances the content of artemisinin and increases tolerance to drought and Botrytis cinerea in Artemisia annua. Plant Cell Physiol. 2016, 57, 1961–1971. [Google Scholar] [CrossRef]
- Lv, Z.; Zhang, L.; Tang, K. New insights into artemisinin regulation. Plant Signal. Behav. 2017, 12, e1366398. [Google Scholar] [CrossRef]
- Kayani, S.; Yanan, M.; Fu, X.; Shen, Q.; Li, Y.; Rahman, S.-U.; Peng, B.; Huang, L.; Tang, K. JA-regulated AaGSW1–AaYABBY5/AaWRKY9 complex regulates artemisinin biosynthesis in Artemisia annua. Plant Cell Physiol. 2023, 64, 771–785. [Google Scholar] [CrossRef]
- Shan, Z.; Jiang, Y.; Li, H.; Guo, J.; Dong, J.; Dong, M.; Zhang, J.; Liu, G. Genome-wide analysis of the NAC transcription factor family in broomcorn millet (Panicum miliaceum L.) and expression analysis under drought stress. BMC Genom. 2020, 21, 96. [Google Scholar] [CrossRef]
- Li, B.; Fan, R.; Yang, Q.; Hu, C.; Sheng, O.; Deng, G.; Dong, T.; Li, C.; Peng, X.; Bi, F.; et al. Genome-wide identification and characterization of the NAC transcription factor family in Musa acuminata and expression analysis during fruit ripening. Int. J. Mol. Sci. 2020, 21, 634. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.-K.; Xu, L.-N.; Leng, Y.-J.; Zhang, M.-Q.; Yang, Q.-Q.; Wang, S.-L.; Jia, S.-W.; Song, T.; Wang, R.-A.; Tao, T.; et al. The OsNAC24-OsNAP protein complex activates OsGBSSI and OsSBEI expression to fine-tune starch biosynthesis in rice endosperm. Plant Biotechnol. J. 2023, 21, 2224–2240. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Liang, X.; Tan, L.; Sun, W.; Dai, X.; Yan, H. Genome-wide identification, evolution and expression profile analysis of NAC transcription factor in Simmondsia chinensis. Curr. Issues Mol. Biol. 2023, 45, 5422–5436. [Google Scholar] [CrossRef]
- Doll, N.M.; Van Hautegem, T.; Schilling, N.; Rycke, R.; Winter, F.; Fendrych, M.; Nowack, M.K. Endosperm cell death promoted by NAC transcription factors facilitates embryo invasion in Arabidopsis. Curr. Biol. 2023, 33, 3785–3795. [Google Scholar] [CrossRef]
- Fourquin, C.; Beauzamy, L.; Chamot, S.; Creff, A.; Goodrich, J.; Boudaoud, A.; Ingram, G. Mechanical stress mediated by both endosperm softening and embryo growth underlies endosperm elimination in Arabidopsis seeds. Development 2016, 143, 3300–3305. [Google Scholar] [CrossRef]
- Feng, F.; Qi, W.; Lv, Y.; Yan, S.; Xu, L.; Yang, W.; Yuan, Y.; Chen, Y.; Zhao, H.; Song, R. OPAQUE11 is a central hub of the regulatory network for maize endosperm development and nutrient metabolism. Plant Cell 2018, 30, 375–396. [Google Scholar] [CrossRef] [PubMed]
- Khöler, C.; Butler, N. Plant development: How to kill the endosperm. Curr. Biol. 2023, 33, R898–R918. [Google Scholar]
- Sun, L.; Liu, L.-P.; Wang, Y.-Z.; Yang, L.; Wang, M.-J.; Liu, J.-X. NAC103, a NAC family transcription factor, regulates ABA response during seed germination and seedling growth in Arabidopsis. Planta 2020, 252, 95. [Google Scholar] [CrossRef]
- Kunieda, T.; Mitsuda, N.; Ohme-Takagi, M.; Takeda, S.; Aida, M.; Tasaka, M.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. NAC family proteins NARS1/NAC2 and NARS2/NAM in the outer integument regulated embryogenesis in Arabidopsis. Plant Cell 2008, 20, 2631–2642. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wu, X.; Wang, W.; Wu, Y. Regulation of seed storage protein synthesis in monocot and dicot plants: A comparative review. Mol. Plant 2023, 16, 145–167. [Google Scholar] [CrossRef]
- Mathew, I.E.; Das, S.; Mahto, A.; Agarwal, P. Three rice NAC transcription factors heteromerize and are associated with seed size. Front. Plant Sci. 2016, 7, 1638. [Google Scholar] [CrossRef]
- Gu, Y.; Han, S.; Chen, L.; Mu, J.; Duan, L.; Li, Y.; Yan, Y.; Li, X. Expression and regulation of genes involved in the reserve starch biosynthesis pathway in hexaploid wheat (Triticum aestivum L.). Crop J. 2021, 9, 440–455. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Z.; Zhang, Q.; Meng, S.; Wei, C. The NAC transcription factors OsNAC20 and OsNAC26 regulate starch and storage protein synthesis. Plant Phys. 2020, 184, 1775–1791. [Google Scholar] [CrossRef]
- Sajeev, N.; Koornneef, M.; Bentsink, L. A commitment for life: Decades of unraveling the molecular mechanisms behind seed dormancy and germination. Plant Cell 2024, 36, 1358–1376. [Google Scholar] [CrossRef]
- Tognacca, R.S.; Botto, S.F. Post-transcriptional regulation of seed dormancy and germination: Current understanding and future directions. Plant Commun. 2021, 2, 100169. [Google Scholar] [CrossRef]
- Sajeev, N.; Bai, B.; Bentsink, L. Seeds: A Unique System to Study Translational Regulation. Trends Plant Sci. 2019, 24, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Matilla, A.J. Exploring breakthroughs in three traits belonging to seed life. Plants 2022, 11, 490. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Mao, C.; Zhong, Q.; Yao, X.; Li, P.; Liu, C.; Ming, F. OsNAC2 is involved in multiple hormonal pathways to mediate germination of rice seeds and establishment of seedling. Front. Plant Sci. 2021, 12, 699303. [Google Scholar] [CrossRef] [PubMed]
- He, H. Environmental Regulation of Seed Performance. Ph.D. Thesis, Wageningen University and Research, Wageningen, The Netherlands, 2014. [Google Scholar]
- Song, S.; Willems, L.; Jiao, A.; Zhao, T.; Schranz, M.E.; Bentsink, L. The membrane associated NAC transcription factors ANAC060 and ANAC040 are functionally redundant in the inhibition of seed dormancy in Arabidopsis thaliana. J. Exp. Bot. 2022, 73, 5514–5528. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Barral, N.; Rodríguez-Gacio, M.C.; Matilla, A.J. Delay of germination-1 (DOG1): A key to understanding seed dormancy. Plants 2020, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Albertos, P.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Fernández-Arbaizar, A.; Nakabayashi, K.; Nambara, K.; Nambara, E.; Godoi, M.; Francos, J.M.; et al. Redox feedback regulation of ANAC089 signaling alters seed germination and stress response. Cell Rep. 2021, 35, 109263. [Google Scholar] [CrossRef]
- Kim, S.G.; Lee, A.K.; Yoon, H.K.; Park, C.M. A membrane-bound NACtranscription factor NTL8 regulates gibberellic acid-mediated salt signaling in Arabidopsis seed germination. Plant J. 2008, 55, 77–88. [Google Scholar] [CrossRef]
- Tian, H.; Wang, X.; Guo, H.; Cheng, Y.; Hou, C.; Chen, J.-G.; Wang, S. NTL regulates trichome formation in Arabidopsis by directly activating R3 MIB genes TRY and TCL1. Plant Physiol. 2017, 174, 510. [Google Scholar] [CrossRef] [PubMed]
- Matilla, A.J. Auxin: Hormonal signal required for seed development and dormancy. Plants 2020, 9, 705. [Google Scholar] [CrossRef]
- Park, J.; Kim, Y.-S.; Kim, S.-G.; Jung, J.-H.; Woo, J.-C.; Park, C.-M. Integration of auxin and salt signals by the NAC transcription factor NTM2 during seed germination in Arabidopsis. Plant Physiol. 2011, 156, 537–549. [Google Scholar] [CrossRef]
- Jung, J.-H.; Park, J. Auxin modulation of salt stress signaling in Arabidopsis seed germination. Plant Signal. Behav. 2011, 6, 1198–1200. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, S.; Nakamura, K. Characterization of a cDNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 5’ upstream regions of genes coding for sporamin and beta-amylase from sweet potato. Mol. Genet. Genet. 1994, 244, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, C.; Wang, H.; Guo, Z. WRKY transcription factors: Evolution, binding, and action. Phytopathol. Res. 2019, 1, 13. [Google Scholar] [CrossRef]
- Mahiwal, S.; Pahuja, S.; Pandey, G.K. Structural-functional relationship of WRKY transcription factors: Unfolding the role of WRKY in plants. Int. J. Biol. Macromol. 2024, 257, 128769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Cao, Y.; Zhao, L.; Zhang, J.; Li, S. WRKY transcription factors: Understanding the functional divergence. Plant Sci. 2023, 334, 111770. [Google Scholar] [CrossRef] [PubMed]
- Dorjee, T.; Cui, Y.; Zhang, Y.; Liu, Q.; Li, X.; Sumbur, B.; Yan, H.; Bing, J.; Geng, Y.; Zhou, Y.; et al. Characterization of NAC gene family in Ammopiptanthus mongolicus and functional analysis of AmNAC24, an osmotic and cold-stress-induced NAC gene. Biomolecules 2024, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Goyal, P.; Devi, R.; Verma, B.; Hussain, S.; Arora, P.; Tabassum, R.; Gupta, S. WRKY transcription factors: Evolution, regulation, and functional diversity in plants. Protoplasma 2023, 260, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Santana, A.; Alcaraz, L.D.; Castaño, E.; Sánchez-Calderón, L.; Sánchez-Teyer, F.; Rodríguez-Zapata, L. Comparative genomics of NAC transcriptional factors in Angiosperms: Implications for the adaptation and diversification of flowering plants. PLoS ONE 2015, 10, e0141866. [Google Scholar]
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef]
- Wang, H.; Chen, W.; Xu, Z.; Chen, M.; Yu, D. Functions of WRKYs in plant growth and development. Trends Plant Sci. 2021, 28, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Bao, C.; Xu, F.; Ma, C.; Huang, L.; Guo, Q.; Luo, M. Silencing GhJUB1L1 (JUB1-like 1) reduces cotton (Gossypium hirsutum) drought tolerance. PLoS ONE 2021, 16, e0259382. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Wu, Y.; Mu, M.; Jiang, J.; Nie, W.; Zhao, S.; Cui, G.; Yin, X. Genome-wide identification and characterization of NAC transcription factor family members in Trifolium pratense and expression analysis under lead stress. BMC Genom. 2024, 25, 128. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, O.; Paes, B.; Silva, I.P.; Braga, P.A.; Pacheco, E. Senescence-associated Glycine max (Gm) NAC genes: Integration of atnural and stress-induced leaf senescence. Int. J. Mol. Sci. 2021, 22, 8287. [Google Scholar]
- Simón, E.B.; Oliveira, C.C.; Fraga, O.T.; Reis, P.A.B.; Fontes, E.P.B. Cell death signaling from endoplasmic reticulum stress: Plant-specific and conserved features. Front Plant Sci. 2022, 13, 835738. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Wu, H.; Xu, H.; Singh, R.M.; Yao, T.; Jang, I.; Chua, N.H. Nutrient status regulates MED19a phase separation for ORESARA1-dependent seescence. New Phytol. 2022, 236, 1779–1795. [Google Scholar] [CrossRef] [PubMed]
- Durian, G.; Sedaghatmehr, M.; Matallana-Ramírez, L.P.; Schilling, S.M.; Schaepe, S.; Guerra, T.; Herde, M.; Witte, C.-P.; Mueller Roeber, B.; Schulze, W.X.; et al. Calcium-dependent protein kinase CPK1 controls cell death by in vivo phosphorylation of senescence master regulator ORE1. Plant Cell 2020, 32, 1610–1625. [Google Scholar] [CrossRef]
- Zhong, Q.; Yu, J.; Yao, X.; Men, X.; Min, F. OsNAC2 maintains the homeostasis of immune responses to bacterial blight through the OsEREBP1 in rice. bioRxiv 2023. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, L.; Guo, C.; Tang, M.; Lian, W.; Chen, S.; Pan, Y.; Xu, X.; Luo, C.; Yi, Y.; et al. OsNAC103, an NAC transcription factor negatively regulates plant height in rice. Planta 2024, 259, 35–53. [Google Scholar] [CrossRef]
- Lei, W.; Li, Y.; Yao, X.; Quiao, K.; Wei, L.; Liu, B.; Zhang, D.; Lin, H. NAP is involved in GA-mediated chlorophyll degradation and leaf senescence by interacting with DELLAs in Arabidopsis. Plant Cell Rep. 2020, 39, 75–87. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Zhou, D.; Zhang, Y.; Song, R.; Li, C.; Li, J.; Gao, J. The roles of gibberellins in regulating leaf development. Plants 2023, 12, 1243. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, B.; Ren, H.; Chen, L.; Watkins, C.B.; Gan, S.-S. The leaf senescence-promoting transcription factor AtNAP activates its direct target gene CYTOKININ OXIDASE 3 to facilitate senescence processes by degrading cytokinins. Mol. Hortic. 2021, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, X.; Gan, S.-S. The ABA-AtNAP-SAG113 PP2C module regulates leaf senescence by dephoshorylating SAG114 SnRK3.25 in Arabidopsis. Mol. Hortic. 2023, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Yu, G.; Lei, S.; Wang, H.; Xu, B. STRONG STAYGREEN inhibits DNA binding of PvNAP transcription factors during leaf senescence in switchgrass. Plant Physiol. 2022, 190, 2045–2058. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Neff, M.M. Two ATAF transcription factors ANAC102 and ATAF1 contribute to the suppression of cytochrome P450-mediated brassinosteroid catabolism in Arabidopsis. Physiol. Plant. 2021, 172, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.; Mogg, S.L.; Soraru, C.; Wallington, E.; Coates, J.; Borrill, P. Wheat NAC transcription factor NAC5-1 is a positive regulator of senescence. bioRxiv 2024. [Google Scholar] [CrossRef]
- Xin, K.; Pan, T.; Gao, S.; Yan, S. A Transcription factor regulates gene expression in chloroplasts. Int. J. Mol. Sci. 2021, 22, 6769. [Google Scholar] [CrossRef] [PubMed]
- Cresta, A.; D’Alessandro, S. Arabidopsis ANAC102, chloroplastic or nucleocytosolic localization? Genes 2023, 14, 438. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Tarkowska, D.; Sedaghatmehr, M.; Welsch, M.; Gupta, S.; Mueller-Roeber, B.; Balazadeh, S. The HB40-JUB1 transcriptional regulatory network controls gibberellin homeostasis in Arabidopsis. Mol. Plant 2022, 15, 322–339. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, C.; Li, K.; Li, X.; Xu, M.; Guo, Y. CLE14 functions as a “brake signal” to suppress age-dependent and stress-induced leaf senescence by promoting JUB1-mediated ROS scavenging in Arabidopsis. Mol. Plant 2022, 15, 179–188. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).