
The Evolution of the WUSCHEL-Related Homeobox Gene Family in Dendrobium Species and Its Role in Sex Organ Development in D. chrysotoxum

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
2.1. Identification and Physicochemical Properties of the WOX Gene Family
2.2. Chromosomal Localization of WOXs
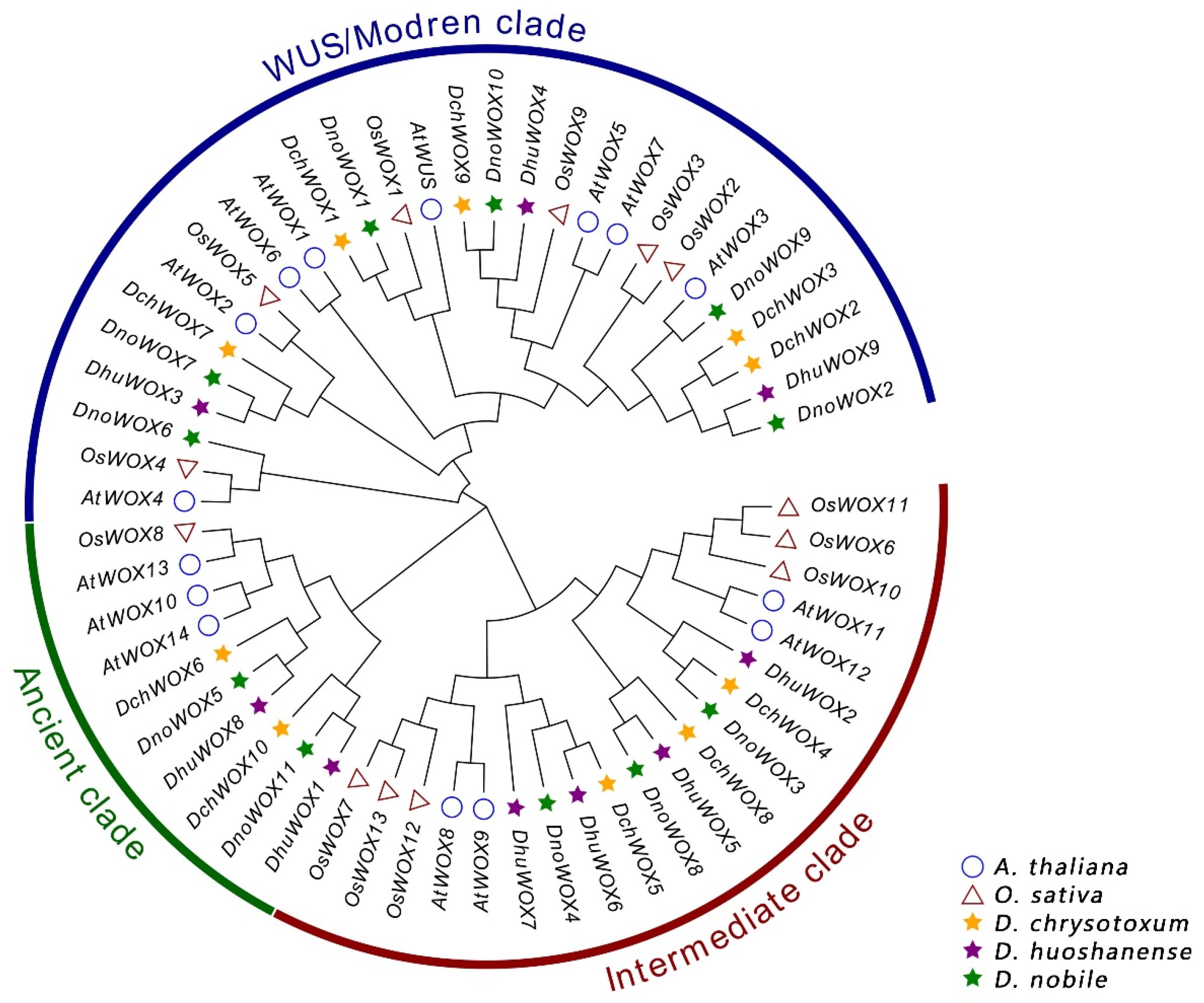
2.3. Phylogenetic Analysis of WOXs
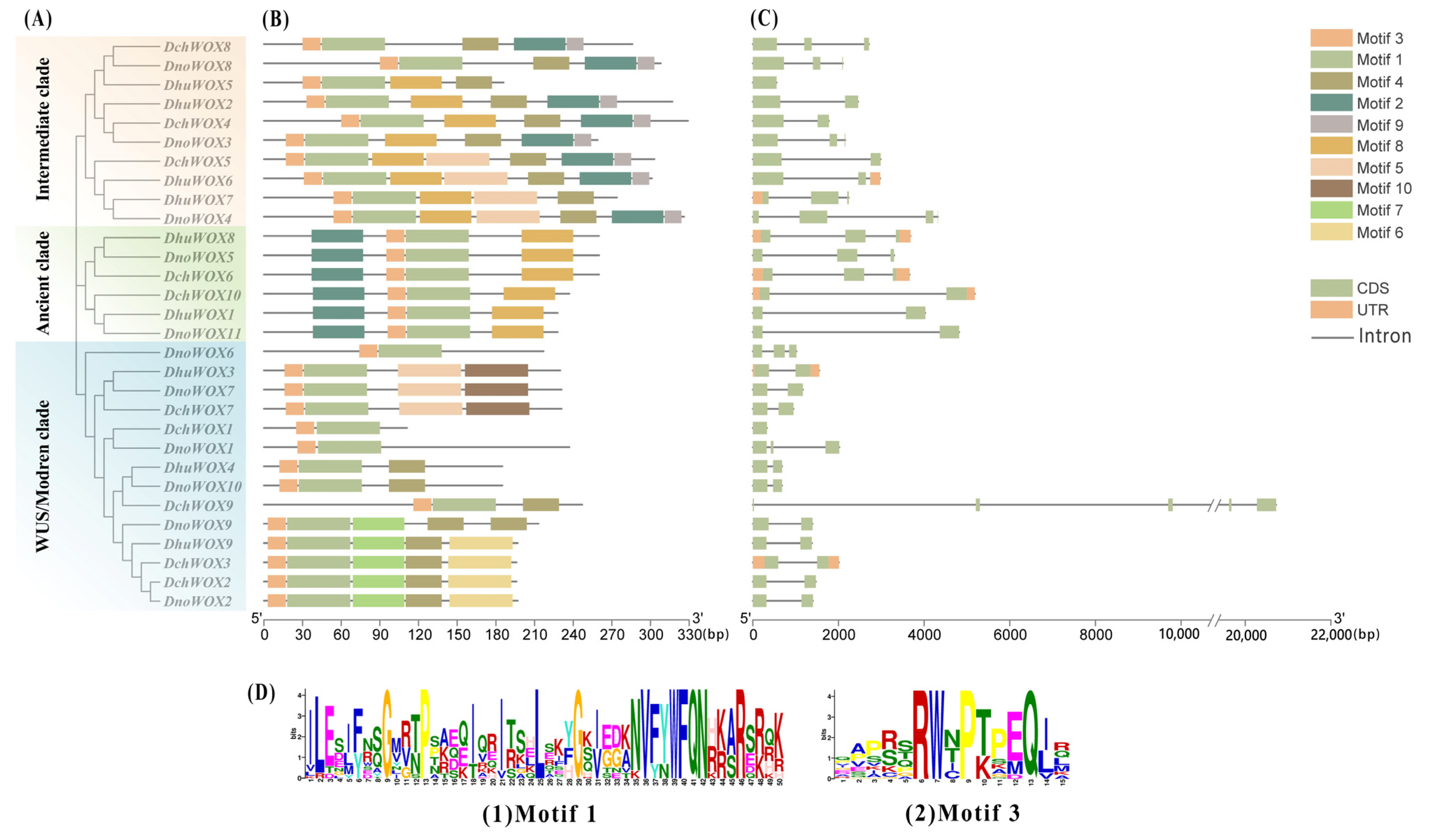
2.4. Gene Structure and Conserved Motifs of WOXs
2.5. Synteny Analysis and Ka/Ks Value of WOX Gene Family
2.6. Cis-Acting Elements Analysis of WOXs
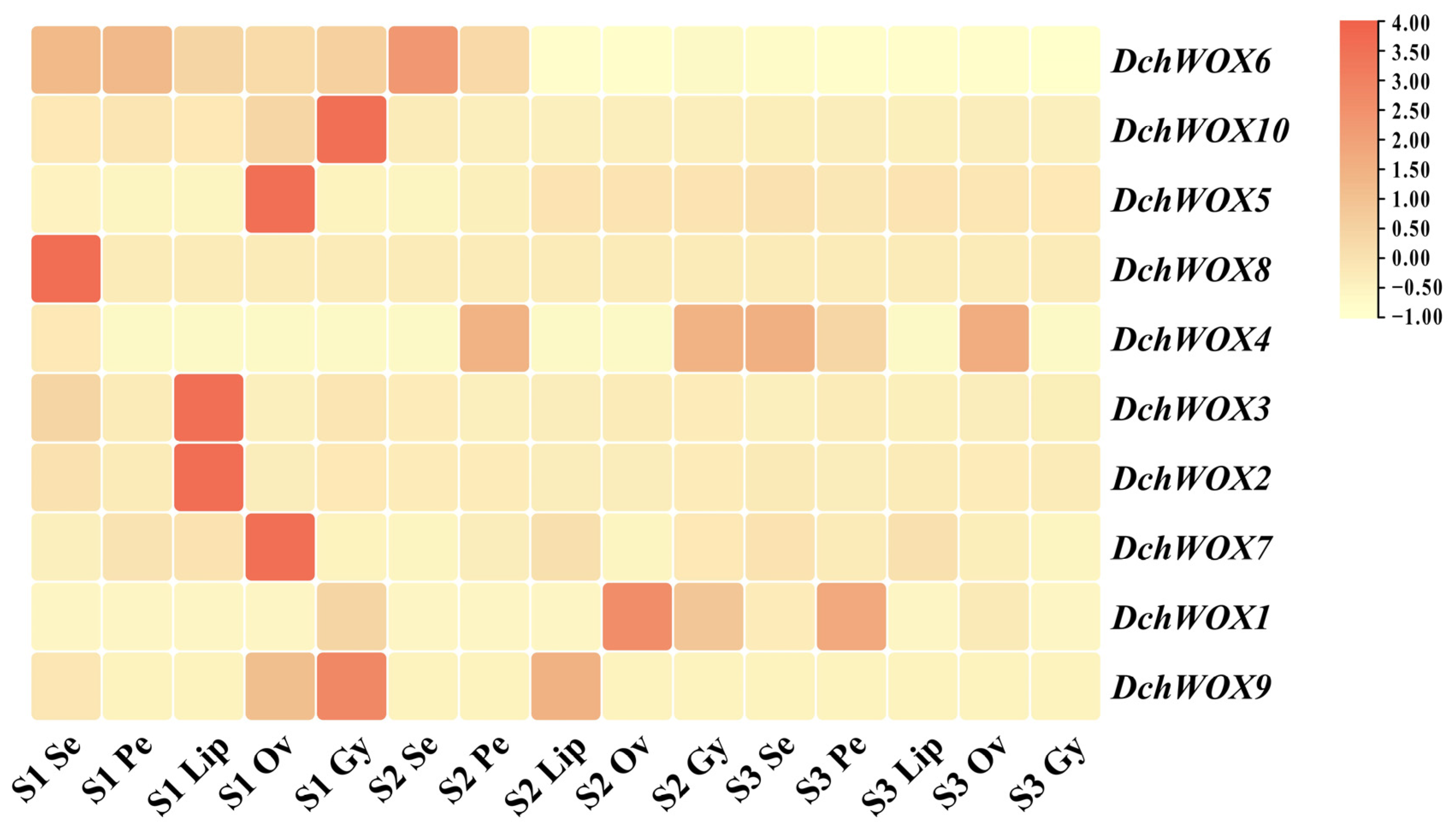
2.7. Expression Pattern Analysis of WOX Gene Family in D. chrysotoxum
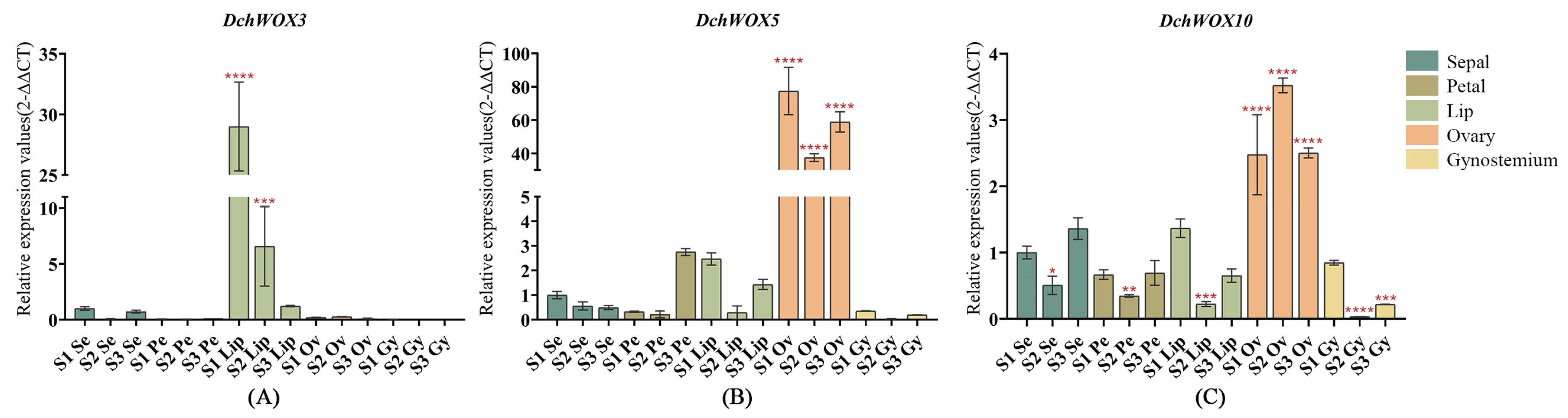
2.8. RT-qPCR Analysis of WOX Genes in D. chrysotoxum
3. Discussion
4. Materials and Methods
4.1. Data Sources
4.2. Identification and Physicochemical Properties of WOXs
4.3. Chromosomal Localization
4.4. Phylogenetic Analysis of WOX Gene Family
4.5. Protein Conservative Domain and Gene Structure Analysis
4.6. Synteny Analysis of WOX Gene Family
4.7. Cis-Acting Regulatory Element Analysis
4.8. Expression Pattern and RT-qPCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gehring, W.J.; Affolter, M.; Bürglin, T. Homeodomain proteins. Annu. Rev. Biochem. 1994, 63, 487. [Google Scholar] [CrossRef] [PubMed]
- Breuninger, H.; Rikirsch, E.; Hermann, M.; Ueda, M.; Laux, T. Differential Expression of WOX Genes Mediates Apical-Basal Axis Formation in the Arabidopsis Embryo. Dev. Cell 2008, 14, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Enrico, C.; Christophe, T.; Michiel, V. The role of WOX genes in flower development. Ann. Bot. 2014, 114, 1545–1553. [Google Scholar]
- Haecker, A.; Groß-Hardt, R.; Geiges, B.; Sarkar, A.; Breuninger, H.; Herrmann, M.; Laux, T. Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development 2004, 131, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Van der Graaff, E.; Laux, T.; Rensing, S.A. The WUS homeobox-containing (WOX) protein family. Genome Biol. 2009, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Lian, G.; Ding, Z.; Wang, Q.; Zhang, D.; Xu, J. Origins and Evolution of WUSCHEL-Related Homeobox Protein Family in Plant Kingdom. Sci. World J. 2014, 2014, 534140. [Google Scholar] [CrossRef] [PubMed]
- Romera-Branchat, M.; Ripoll, J.J.; Yanofsky, M.F.; Pelaz, S. The WOX13 homeobox gene promotes replum formation in the Arabidopsis thaliana fruit. Plant J. 2012, 73, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Deveaux, Y.; Toffano-Nioche, C.; Claisse, G.; Thareau, V.; Morin, H.; Laufs, P.; Moreau, H.; Kreis, M.; Lecharny, A. Genes of the most conserved WOX clade in plants affect root and flower development in Arabidopsis. BMC Evol. Biol. 2008, 8, 291. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Lippman, Z.B.; Cohen, O.; Alvarez, J.P.; Abu-Abied, M.; Pekker, I.; Paran, I.; Eshed, Y.; Zamir, D. The Making of a Compound Inflorescence in Tomato and Related Nightshades. PLoS Biol. 2008, 6, e288. [Google Scholar]
- Wang, W.; Li, G.; Zhao, J.; Chu, H.; Lin, W.; Zhang, D.; Wang, Z.; Liang, W. DWARF TILLER1, a WUSCHEL-Related Homeobox Transcription Factor, Is Required for Tiller Growth in Rice. PLoS Genet. 2014, 10, e1004154. [Google Scholar] [CrossRef]
- Hendelman, A.; Zebell, S.; Rodriguez-Leal, D.; Dukler, N.; Lippman, Z.B. Conserved pleiotropy of an ancient plant homeobox gene uncovered by cis-regulatory dissection. Cell 2021, 184 (Suppl. S1), 1724–1739. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sheng, L.; Xu, Y.; Li, J.; Yang, Z.; Huang, H.; Xu, L. WOX11 and 12 Are Involved in the First-Step Cell Fate Transition during de Novo Root Organogenesis in Arabidopsis. Plant Cell 2014, 26, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.F.X.; Schoof, H.; Haecker, A.; Lenhard, M.; Jürgens, G.; Laux, T. Role of WUSCHEL in Regulating Stem Cell Fate in the Arabidopsis Shoot Meristem. Cell 1998, 95, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Ochatt, S.J.; Kumar, V. WUSCHEL: A master regulator in plant growth signaling. Plant Cell Rep. 2020, 39, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Mitsuda, N.; Ohme-Takagi, M. Arabidopsis WUSCHEL Is a Bifunctional Transcription Factor That Acts as a Repressor in Stem Cell Regulation and as an Activator in Floral Patterning. Plant Cell 2009, 21, 3493–3505. [Google Scholar] [CrossRef] [PubMed]
- Nakata, M.; Matsumoto, N.; Tsugeki, R.; Rikirsch, E.; Okada, K. Roles of the middle domain-specific WUSCHEL-RELATED HOMEOBOX genes in early development of leaves in Arabidopsis. Plant Cell 2012, 24, 519–535. [Google Scholar] [CrossRef]
- Honda, E.; Yew, C.L.; Yoshikawa, T.; Sato, Y.; Hibara, K.I.; Itoh, J.I. LEAF LATERAL SYMMETRY1, a Member of the WUSCHEL-RELATED HOMEOBOX3 Gene Family, Regulates Lateral Organ Development Differentially from Other Paralogs, NARROW LEAF2 and NARROW LEAF3 in Rice. Plant Cell Physiol. 2018, 59, 376–391. [Google Scholar] [CrossRef] [PubMed]
- Batool, H.S.; François, T.J.; Juliane, R.; Kurt, Z.; Andrea, R. Constitutive Overexpression of a Conifer WOX2 Homolog Affects Somatic Embryo Development in Pinus pinaster and Promotes Somatic Embryogenesis and Organogenesis in Arabidopsis Seedlings. Front. Plant Sci. 2022, 13, 838421. [Google Scholar]
- Sarkar, A.K.; Luijten, M.; Miyashima, S.; Lenhard, M.; Hashimoto, T.; Nakajima, K.; Scheres, B.; Heidstra, R.; Laux, T. Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organizers. Nature 2007, 446, 811–814. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Escudero, N.; Zavala-Gonzalez, E.A.; Esteve-Bruna, D.; Blázquez, M.A.; Alabadí, D.; Lopez-Llorca, L.V. Induction of auxin biosynthesis and WOX5 repression mediate changes in root development in Arabidopsis exposed to chitosan. Sci. Rep. 2017, 7, 16813. [Google Scholar] [CrossRef]
- Wang, L.Q.; Wen, S.S.; Wang, R.; Wang, C.; Gao, B.; Lu, M.Z. PagWOX11/12a activates PagCYP736A12 gene that facilitates salt tolerance in poplar. Plant Biotechnol. J. 2021, 19, 2249–2260. [Google Scholar] [CrossRef]
- Sajjad, M.; Wei, X.; Liu, L.; Li, F.; Ge, X. Transcriptome Analysis Revealed GhWOX4 Intercedes Myriad Regulatory Pathways to Modulate Drought Tolerance and Vascular Growth in Cotton. Int. J. Mol. Sci. 2021, 22, 898. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Pan, X.; Jing, Y.; Zhao, Y.; Duan, Y.; Yang, J.; Wang, B.; Liu, Y.; Shen, R.; Cao, Y.; et al. ZmSPL10/14/26 are required for epidermal hair cell fate specification on maize leaf. New Phytol. 2021, 230, 1533–1549. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.J.; Grotewold, E.; Stam, M. Cis-regulatory sequences in plants: Their importance, discovery, and future challenges. Plant Cell 2022, 34, 718–741. [Google Scholar] [CrossRef]
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef]
- Liu, H.; Luo, Y.-B. Protecting Orchids in Nature Reserves: Research and Restoration Needs. Bot. Rev. 2010, 76, 137–139. [Google Scholar] [CrossRef]
- Pridgeon, A.M.; Cribb, P.; Chase, M.W.; Rasmussen, F.N. Genera Orchidacearum Volume 6: Epidendroideae (Part 3); OUP: Oxford, UK, 2014. [Google Scholar]
- Niu, Z.; Zhu, F.; Fan, Y.; Li, C.; Zhang, B.; Zhu, S.; Hou, Z.; Wang, M.; Yang, J.; Xue, Q.; et al. The chromosome-level reference genome assembly for Dendrobium officinale and its utility of functional genomics research and molecular breeding study. Acta Pharm. Sin. B 2021, 11, 2080–2092. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, G.-Q.; Zhang, D.; Liu, X.-D.; Xu, X.-Y.; Sun, W.-H.; Yu, X.; Zhu, X.; Wang, Z.-W.; Zhao, X.; et al. Chromosome-scale assembly of the Dendrobium chrysotoxum genome enhances the understanding of orchid evolution. Hortic. Res. 2021, 8, 183. [Google Scholar] [CrossRef]
- Bangxing, H.; Yi, J.; Jun, D.; Tao, Z.; Fangli, G.; Qun, Z.; Fucheng, Z.; Xiangwen, S.; Hui, D.; Peipei, W.; et al. A chromosome-level genome assembly of Dendrobium huoshanense using long reads and Hi-C data. Genome Biol. Evol. 2020, 12, 2486–2490. [Google Scholar]
- Xu, Q.; Niu, S.-C.; Li, K.-L.; Zheng, P.-J.; Zhang, X.-J.; Jia, Y.; Liu, Y.; Niu, Y.-X.; Yu, L.-H.; Chen, D.-F.; et al. Chromosome-Scale Assembly of the Dendrobium nobile Genome Provides Insights Into the Molecular Mechanism of the Biosynthesis of the Medicinal Active Ingredient of Dendrobium. Front. Genet. 2022, 13, 844622. [Google Scholar] [CrossRef]
- Gasteiger, E. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005. [Google Scholar]
- Lin, H.; Niu, L.; McHale, N.A.; Ohme-Takagi, M.; Mysore, K.S.; Tadege, M. Evolutionarily conserved repressive activity of WOX proteins mediates leaf blade outgrowth and floral organ development in plants. Proc. Natl. Acad. Sci. USA 2013, 110, 366–371. [Google Scholar] [CrossRef]
- Li, H.; Li, C.; Wang, Y.; Qin, X.; Meng, L.; Sun, X. Genome-Wide Analysis of the WOX Transcription Factor Genes in Dendrobium catenatum Lindl. Genes 2022, 13, 1481. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, T.R.; Kanchan, M.; Upadhyay, S.K.; Sembi, J.K. Identification and characterization of WUSCHEL-related homeobox (WOX) gene family in economically important orchid species Phalaenopsis equestris and Dendrobium catenatum. Plant Gene 2018, 14, 37–45. [Google Scholar] [CrossRef]
- Zhang, X.; Zong, J.; Liu, J.; Yin, J.; Zhang, D. Genome-Wide Analysis of WOX Gene Family in Rice, Sorghum, Maize, Arabidopsis and Poplar. J. Integr. Plant Biol. 2010, 52, 1016–1026. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Minuto, M.; Boccacci, P.; Perrone, I.; Vallania, R.; Gribaudo, I. Characterization of expression dynamics of WOX homeodomain transcription factors during somatic embryogenesis in Vitis vinifera. J. Exp. Bot. 2011, 62, 1089. [Google Scholar] [CrossRef] [PubMed]
- Xuemei, Z.; Yingying, G.; Peng, Z.; Meng-Xiang, S. Comparative Analysis of WUSCHEL-Related Homeobox Genes Revealed Their Parent-of-Origin and Cell Type-Specific Expression Pattern During Early Embryogenesis in Tobacco. Front. Plant Sci. 2018, 9, 311. [Google Scholar]
- Shi, L.; Wang, K.; Du, L.; Song, Y.; Li, H.; Ye, X. Genome-Wide Identification and Expression Profiling Analysis of WOX Family Protein-Encoded Genes in Triticeae Species. Multidiscip. Digit. Publ. Inst. 2021, 22, 9325. [Google Scholar] [CrossRef]
- Wang, M.-M.; Liu, M.-M.; Ran, F.; Guo, P.-C.; Ke, Y.-Z.; Wu, Y.-W.; Wen, J.; Li, P.-F.; Li, J.-N.; Du, H. Global Analysis of WOX Transcription Factor Gene Family in Brassica napus Reveals Their Stress- and Hormone-Responsive Patterns. Int. J. Mol. Sci. 2018, 19, 3470. [Google Scholar] [CrossRef]
- Hao, Q.; Zhang, L.; Yang, Y.; Shan, Z.; Zhou, X.A. Genome-Wide Analysis of the WOX Gene Family and Function Exploration of GmWOX18 in Soybean. Plants 2019, 8, 215. [Google Scholar] [CrossRef]
- Ruibin, S.; Xue, Z.; Dan, M.; Chuanliang, L. Identification and Evolutionary Analysis of Cotton (Gossypium hirsutum) WOX Family Genes and Their Potential Function in Somatic Embryogenesis. Int. J. Mol. Sci. 2023, 24, 11077. [Google Scholar] [CrossRef]
- Flagel, L.E.; Wendel, J.F. Gene duplication and evolutionary novelty in plants. New Phytol. 2009, 183, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Gao, X.; Wu, Y.; Wei, C.; Li, M.H.; Liu, D.K.; Liu, Z.J. Identification and Analysis of PEPC Gene Family Reveals Functional Diversification in Orchidaceae and the Regulation of Bacterial-TypePEPC. Int. J. Mol. Sci. 2024, 25, 2055. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guan, C.; Chen, Y.; Shi, Y.; Long, O.; Lin, H.; Zhang, K.; Zhou, M. Evolutionary analysis of MADS-box genes in buckwheat species and functional study of FdMADS28 in flavonoid metabolism. Plant Physiol. Biochem. 2024, 210, 108637. [Google Scholar] [CrossRef]
- Hedman, H.; Zhu, T.; von Arnold, S.; Sohlberg, J.J. Analysis of the WUSCHEL-RELATED HOMEOBOX gene family in the conifer picea abiesreveals extensive conservation as well as dynamic patterns. BMC Plant Biol. 2013, 13, 89. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yang, M.; Liu, X.; Xia, Y.; Hu, R.; Xia, Q.; Jing, D.; Guo, Q. Genome-wide analysis of the WOX gene family and the role of EjWUSa in regulating flowering in loquat (Eriobotrya japonica). Front. Plant Sci. 2022, 13, 1024515. [Google Scholar] [CrossRef]
- Feng, C.; Zou, S.; Gao, P.; Wang, Z. In silico identification, characterization expression profile of WUSCHEL-Related Homeobox (WOX) gene family in two species of kiwifruit. Peer J. 2021, 9, e12348. [Google Scholar] [CrossRef]
- Muhammad Tajo, S.; Pan, Z.; He, S.; Chen, B.; Km, Y.; Mahmood, T.; Bello Sadau, S.; Shahid Iqbal, M.; Gereziher, T.; Suleiman Abubakar, U.; et al. Characterization of WOX genes revealed drought tolerance, callus induction, and tissue regeneration in Gossypium hirsutum. Front. Genet. 2022, 13, 928055. [Google Scholar] [CrossRef]
- Zhang, Z.A.; Liu, M.Y.; Ren, S.N.; Liu, X.; Gao, Y.H.; Zhu, C.Y.; Niu, H.Q.; Chen, B.W.; Liu, C.; Yin, W.; et al. Identification of WUSCHEL-related homeobox gene and truncated small peptides in transformation efficiency improvement in Eucalyptus. BMC Plant Biol. 2023, 23, 604. [Google Scholar] [CrossRef]
- Li, M.; Wang, R.; Liu, Z.; Wu, X.; Wang, J. Genome-wide identification and analysis of the WUSCHEL-related homeobox (WOX) gene family in allotetraploid Brassica napus reveals changes in WOX genes during polyploidization. BMC Genom. 2019, 20, 317. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef]
- Guo, L.; Lu, S.; Liu, T.; Nai, G.; Ren, J.; Gou, H.; Chen, B.; Mao, J. Genome-Wide Identification and Abiotic Stress Response Analysis of PP2C Gene Family in Woodland and Pineapple Strawberries. Int. J. Mol. Sci. 2023, 24, 4049. [Google Scholar] [CrossRef] [PubMed]
- Pham-Thi, M.-T.; Sug, K.J.; Songhwa, C.; Mi, J.K.; Gang-Seob, L.; Dong-Eun, K.; Jong-Joo, C.; Ik, S.S.; Hie, N.B.; Yeon-Ki, K. A WUSCHEL Homeobox Transcription Factor, OsWOX13, Enhances Drought Tolerance and Triggers Early Flowering in Rice. Mol. Cells 2018, 41, 781–798. [Google Scholar]
- Tao, Z.; Ruonan, L.; Jialing, X.; Lang, Y.; Rongchen, W.; Yunde, Z. The YUCCA-Auxin-WOX11 Module Controls Crown Root Development in Rice. Front. Plant Sci. 2018, 9, 523. [Google Scholar]
- Ji, J.; Shimizu, R.; Sinha, N.; Scanlon, M.J. Analyses of WOX4 transgenics provide further evidence for the evolution of theWOXgene family during the regulation of diverse stem cell functions. Plant Signal. Behav. 2014, 5, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Fiume, E.; Fletcher, J.C. Regulation of Arabidopsis Embryo and Endosperm Development by the Polypeptide Signaling Molecule CLE8. Plant Cell 2012, 24, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Z.; Huang, J.; Lin, Z.C.; Wang, F.; Yang, S.M.; Jiang, X.; Ahmad, S.; Zhou, Y.Z.; Lan, S.; Liu, Z.J.; et al. Genome-Wide Analysis of WUSCHEL-Related Homeobox Gene Family in Sacred Lotus (Nelumbo nucifera). Int. J. Mol. Sci. 2023, 24, 14216. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhao, X.; Miao, Y.; Wang, D.; Zhang, Z.; Liu, Y. Genome-Wide Identification and Expression Profile Analysis of the WUSCHEL-Related Homeobox (WOX) Genes in Woodland Strawberry (Fragaria vesca). Horticulturae 2022, 8, 1043. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Xia, R. TBtools-II: A One for All, All for One Bioinformatics Platform for Biological Big-data Mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2015, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Riccucci, E.; Vanni, C.; Vangelisti, A.; Fambrini, M.; Giordani, T.; Cavallini, A.; Mascagni, F.; Pugliesi, C. Genome-Wide Analysis of WOX Multigene Family in Sunflower (Helianthus annuus L.). Int. J. Mol. Sci. 2023, 24, 3352. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Nane | Number of Amino Acids (aa) | Molecular Weight (kDa) | Theoretical PI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| DchWOX6 | 259 | 29.82 | 5.34 | 66.87 | 58.76 | −0.887 | Nucleus |
| DchWOX8 | 285 | 31.33 | 7.68 | 65.78 | 70.07 | −0.395 | Nucleus |
| DchWOX4 | 328 | 35.07 | 8.37 | 70.06 | 79.73 | −0.065 | Nucleus |
| DchWOX3 | 195 | 22.15 | 9.11 | 68.49 | 53.03 | −0.772 | Nucleus |
| DchWOX7 | 230 | 25.85 | 6.32 | 77.88 | 61.09 | −0.693 | Nucleus |
| DchWOX2 | 195 | 22.14 | 9.11 | 69.91 | 53.03 | −0.772 | Nucleus |
| DchWOX10 | 236 | 27.19 | 6.27 | 67.38 | 64.87 | −0.834 | Nucleus |
| DchWOX5 | 302 | 33.16 | 6.24 | 55.35 | 91.06 | −0.098 | Nucleus |
| DchWOX9 | 246 | 28.41 | 9.47 | 74.03 | 77.72 | −0.706 | Nucleus |
| DchWOX1 | 110 | 12.84 | 9.69 | 69.36 | 69.36 | −1.195 | Nucleus |
| DhuWOX4 | 184 | 21.06 | 9.08 | 60.70 | 73.10 | −0.684 | Nucleus |
| DhuWOX7 | 273 | 29.67 | 9.48 | 70.79 | 71.14 | −0.472 | Nucleus |
| DhuWOX1 | 227 | 26.35 | 6.20 | 66.47 | 71.28 | −0.717 | Nucleus |
| DhuWOX3 | 229 | 25.90 | 7.00 | 75.10 | 60.09 | −0.691 | Nucleus |
| DhuWOX6 | 300 | 32.64 | 6.31 | 63.71 | 88.43 | −0.170 | Nucleus |
| DhuWOX8 | 259 | 29.91 | 5.26 | 65.97 | 60.27 | −0.878 | Nucleus |
| DhuWOX5 | 185 | 19.89 | 8.98 | 79.63 | 54.38 | −0.556 | Nucleus |
| DhuWOX9 | 196 | 22.14 | 9.11 | 65.29 | 53.78 | −0.758 | Nucleus |
| DhuWOX2 | 316 | 34.27 | 7.66 | 62.10 | 75.66 | −0.110 | Nucleus |
| DnoWOX11 | 227 | 26.31 | 6.20 | 66.47 | 68.72 | −0.750 | Nucleus |
| DnoWOX7 | 230 | 26.04 | 6.60 | 74.71 | 59.39 | −0.694 | Nucleus |
| DnoWOX4 | 325 | 35.30 | 7.79 | 61.37 | 76.86 | −0.364 | Nucleus |
| DnoWOX6 | 216 | 24.38 | 9.91 | 69.27 | 71.34 | −0.748 | Nucleus |
| DnoWOX2 | 196 | 22.16 | 9.11 | 69.22 | 53.78 | −0.760 | Nucleus |
| DnoWOX8 | 307 | 33.27 | 6.30 | 69.20 | 75.28 | −0.173 | Nucleus |
| DnoWOX10 | 184 | 21.15 | 9.39 | 61.85 | 74.67 | −0.664 | Nucleus |
| DnoWOX5 | 259 | 29.92 | 5.26 | 66.63 | 60.27 | −0.876 | Nucleus |
| DnoWOX1 | 236 | 26.49 | 5.75 | 55.23 | 55.08 | −0.838 | Nucleus |
| DnoWOX3 | 258 | 27.53 | 5.78 | 66.90 | 69.26 | −0.258 | Nucleus |
| DnoWOX9 | 212 | 24.24 | 6.79 | 78.91 | 48.77 | −0.922 | Nucleus |
| Gene 1 | Gene 2 | Ka | Ks | Ka/Ks |
|---|---|---|---|---|
| DchWOX4 | DchWOX8 | 0.332789 | 1.919410 | 0.173381 |
| DhuWOX5 | DhuWOX2 | 0.305003 | 2.326057 | 0.131124 |
| DnoWOX2 | DnoWOX9 | 0.226893 | 1.109762 | 0.204452 |
| DnoWOX3 | DnoWOX8 | 0.267376 | 1.697561 | 0.157506 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; Zheng, Q.; He, X.; Zhao, X.; Zhang, M.; Huang, Y.; Cai, B.; Liu, Z. The Evolution of the WUSCHEL-Related Homeobox Gene Family in Dendrobium Species and Its Role in Sex Organ Development in D. chrysotoxum. Int. J. Mol. Sci. 2024, 25, 5352. https://doi.org/10.3390/ijms25105352
Luo X, Zheng Q, He X, Zhao X, Zhang M, Huang Y, Cai B, Liu Z. The Evolution of the WUSCHEL-Related Homeobox Gene Family in Dendrobium Species and Its Role in Sex Organ Development in D. chrysotoxum. International Journal of Molecular Sciences. 2024; 25(10):5352. https://doi.org/10.3390/ijms25105352
Chicago/Turabian StyleLuo, Xiaoting, Qinyao Zheng, Xin He, Xuewei Zhao, Mengmeng Zhang, Ye Huang, Bangping Cai, and Zhongjian Liu. 2024. "The Evolution of the WUSCHEL-Related Homeobox Gene Family in Dendrobium Species and Its Role in Sex Organ Development in D. chrysotoxum" International Journal of Molecular Sciences 25, no. 10: 5352. https://doi.org/10.3390/ijms25105352
APA StyleLuo, X., Zheng, Q., He, X., Zhao, X., Zhang, M., Huang, Y., Cai, B., & Liu, Z. (2024). The Evolution of the WUSCHEL-Related Homeobox Gene Family in Dendrobium Species and Its Role in Sex Organ Development in D. chrysotoxum. International Journal of Molecular Sciences, 25(10), 5352. https://doi.org/10.3390/ijms25105352