
Beneficial Effects of Bovine Milk Exosomes in Metabolic Interorgan Cross-Talk
by
 , , , and
, , , and
Jorge García-Martínez
1,†, 
Íñigo M. Pérez-Castillo
1,†,
Rafael Salto
2,* ,
,
José M. López-Pedrosa
1,
Ricardo Rueda
1 and
María D. Girón
2 1
Abbott Nutrition R&D, Abbott Laboratories, 18004 Granada, Spain
2
Department of Biochemistry and Molecular Biology II, School of Pharmacy, University of Granada, Campus de Cartuja, 18071 Granada, Spain
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Nutrients 2022, 14(7), 1442; https://doi.org/10.3390/nu14071442
Submission received: 3 March 2022
/
Revised: 25 March 2022
/
Accepted: 28 March 2022
/
Published: 30 March 2022
(This article belongs to the Special Issue Natural Food Ingredients for Human Health: New Insights from a Metabolic Perspective)
Abstract
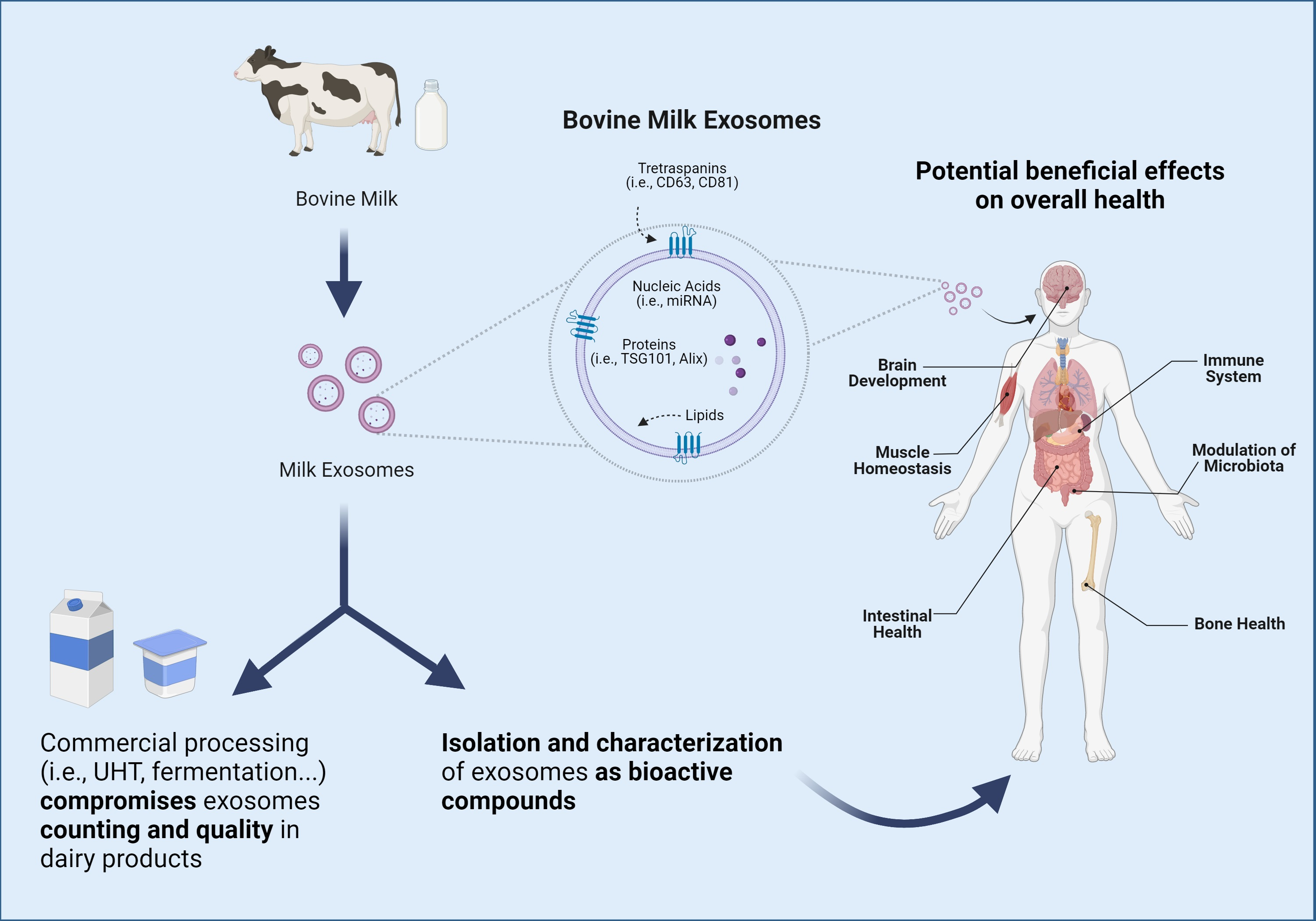
:Extracellular vesicles are membrane-enclosed secreted vesicles involved in cell-to-cell communication processes, identified in virtually all body fluids. Among extracellular vesicles, exosomes have gained increasing attention in recent years as they have unique biological origins and deliver different cargos, such as nucleic acids, proteins, and lipids, which might mediate various health processes. In particular, milk-derived exosomes are proposed as bioactive compounds of breast milk, which have been reported to resist gastric digestion and reach systemic circulation, thus being bioavailable after oral intake. In the present manuscript, we critically discuss the available evidence on the health benefits attributed to milk exosomes, and we provide an outlook for the potential future uses of these compounds. The use of milk exosomes as bioactive ingredients represents a novel avenue to explore in the context of human nutrition, and they might exert important beneficial effects at multiple levels, including but not limited to intestinal health, bone and muscle metabolism, immunity, modulation of the microbiota, growth, and development.
Keywords:
milk; exosomes; extracellular vesicles; miRNA; immunity; intestinal health; bone; muscle; microbiota; neurodevelopment
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Background
Extracellular vesicles (EVs) were first described in the 1960s and 1970s and were considered mere cellular artifacts lacking a biological purpose [1,2]. It was not until the decade of the 1990s that EVs were attributed a role in cell-to-cell communication, thereby gaining increased attention from the scientific community [2,3]. Since then, EVs have been isolated from almost all mammalian cells and are known to be present in virtually all body fluids constituting a burgeoning field of research [4,5].
No consensus on EVs definition has been reached so far. However, EVs are generally considered as membrane-enclosed secreted vesicles that encompass various subsets of different compounds, namely exosomes, ectosomes, microvesicles, microparticles, and apoptotic bodies [6]. These different EVs subsets are characterized based on their size, biological origins, release pathways, functionality, and cargos [7]. In particular, the study of exosomes has witnessed breakthrough progress in recent times. Exosomes are typically considered as molecules sized around 10–200 nm, which originate from repeated invagination of the lipid bilayer membrane of multivesicular bodies [8,9] and can harbor a variety of different cargos and components, such as lipids, proteins, and nucleic acids, thus acting as delivery molecules [10]. Regarding their cargos, the nature of the exosomal content is heterogeneous and depends on its cellular origins and microenvironment [11]. However, the loading of cargos into the exosome is a non-random process that obeys their biological purpose [12]. Hence, given the importance of exosomes in cell-to-cell communication and their presence in diverse body tissues, a database was developed in 2009 with the endeavor of cataloging exosome cargos from all types of tissues [13]. To this date, more than 1960 proteins, 2830 miRNAs, and 1110 lipids have been reported in exosomes [14].
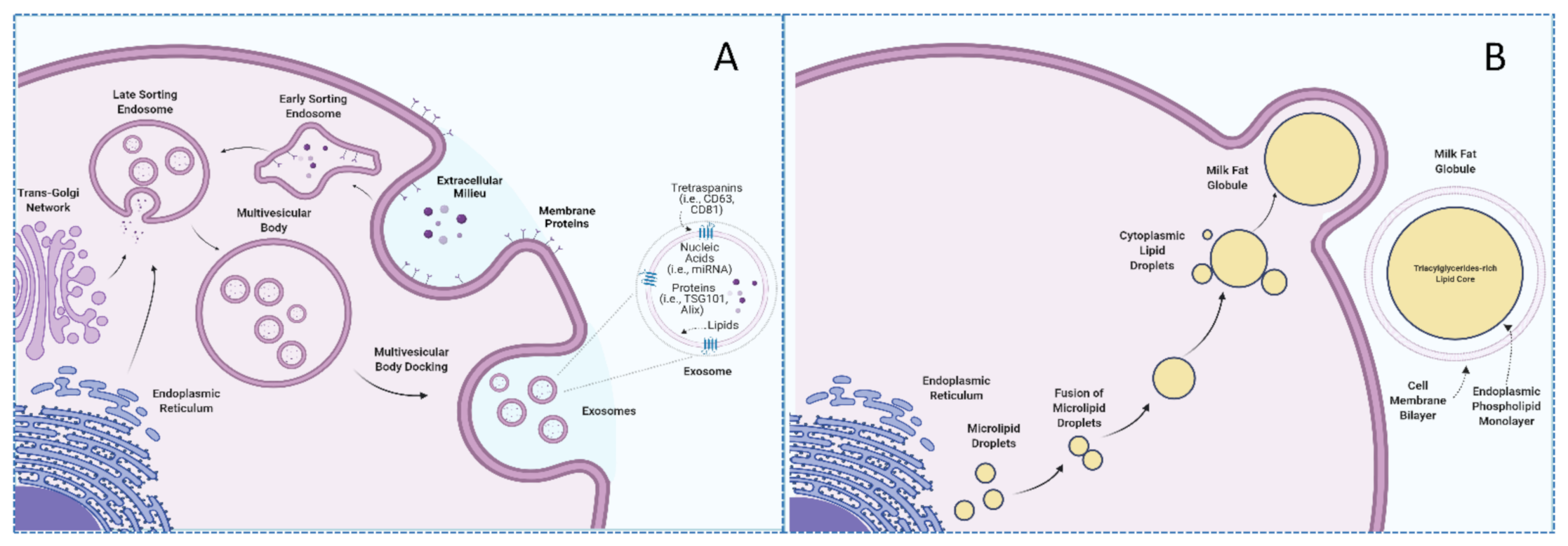
Cellular Origins of Exosomes
From the scientific point of view, one of the most interesting aspects of exosomes resides in their unique biogenesis process. Exosomes biosynthesis begins with the endocytosis of extracellular environment constituents by invagination of the plasma membrane along with the internalization of their associated surface proteins, which leads to the formation of an early-sorting endosome (ESE). The trans-Golgi network and the endoplasmic reticulum can contribute to the content of the ESE, which matures into late-sorting endosomes (LSE) to eventually generate multivesicular-bodies (MVBs). MVBs contain intraluminal vesicles that will constitute future exosomes once the MVB fuses with the plasma membrane [11]. In contrast, larger EVs, such as microvesicles and apoptotic bodies, originate via budding and shedding from the plasma membrane of cells. Several molecules, such as TSG101, ALIX, ESCRT, and ceramides, among others, are involved in the biogenesis of exosomes and, therefore, have been proposed as exosome (specific) markers [5,11,15]. However, these proteins might be involved in the trafficking of other vesicles, thus potentially acting as a confounding factor when conducting research in the field of exosomes [11]. Similarly, exosomes usually display certain proteins in their membranes, namely tetraspanins CD9, CD63, and CD81, which can serve as exosome biomarkers, albeit caution must be warranted as tetraspanins can be indistinctly expressed in the membrane of different vesicles [16].
Due to their biogenesis process, exosomes content resembles that of the donor cell. Thus, the characteristics of exosomes components and cargos will depend mainly on the tissue where they originate. As previously stated, exosomes have been isolated from virtually all human fluids, including plasma, saliva, urine, and breast milk [17]. In this regard, the presence of exosomes in breast milk is of particular interest, given that not only does milk constitute fundamental nutrition for the newborn but also contains bioactive compounds which have been implicated in growth, neurodevelopment, and immunomodulatory processes [18,19,20].
2. Materials and Methods
We comprehensively analyzed the available scientific literature on milk exosomes, including both reviews and original research articles. An extensive search was conducted in the scientific databases and search engines PubMed, ScienceDirect, and Google Scholar using relevant search terms (i.e., “exosomes”, “extracellular vesicles”, “milk”, “dairy”) in combination with Boolean operators (i.e., “AND”, “OR”, “NOT”) with no time restrictions. Additionally, the ProQuest Dialog® search tool was used to screen different electronic databases and repositories such as Embase, BIOSIS Previews, and ProQuest Dissertations and Theses Professional to supplement the literature search.
3. Milk-Derived Exosomes
Milk is a nutrient-rich fluid produced by female mammals whose main purpose is to meet the nutritional requirements of the newborn. Due to its excellent nutritional profile, milk constitutes an important component of the human diet [21]. In addition to nutritional properties, milk contains a wide repertoire of bioactive compounds to which potential health benefits have been attributed [22]. In particular, different EVs, including exosomes and milk fat globules (MFG), are included among described milk bioactive compounds [23]. These vesicles have different sizes, origins, compositions, and functions. Specifically, MFG are far bigger structures (up to 20 μm) constituted by a triacylglyceride-rich core enveloped by a polar lipid trilayer [24], while exosomes are nanosized (10–200 nm), lipid bilayered vesicles with notably lower triacylglyceride content [25]. In the same fashion, lipidomic analyses have suggested that these compounds differ in the presence of phospholipids, with a higher proportion of phosphatidylserine and sphingomyelin being reported in human and bovine milk exosomes in comparison with milk fat globule membranes (MFGM) [25]. Not only lipid but also protein compositions of MFGM and exosomes display notable differences [25,26,27]. According to the BoMiProt database, more than 1300 proteins have been exclusively found in bovine milk exosomes, while 294 have been identified in MFGM [26]. These differences would arise due to their different biogenesis pathways. Whereas exosomes have endosomal origins, MFG are formed in the endoplasmic reticulum and are released into the cytosol as lipid droplets [28,29] (as illustrated in Figure 1).
3.1. Milk Exosomes Isolation and Characterization Methods
The isolation of milk exosomes remains a challenging process. Since extracellular vesicles are an emergent field of research, no consensus has yet been reached on isolation or characterization methods. The International Society for Extracellular Vesicles (ISEV) provided criteria and recommendations for the isolation and analysis of EVs, which can serve as guidance for the accurate reporting of exosomes studies [6,30]. Accordingly, these studies should describe in detail the methods employed to isolate and characterize exosomes and report aspects related to these processes, such as purity, size distribution, and enriched markers assessed [6]. In this regard, the methods used to isolate exosomes from different sources are also applicable to milk exosomes. However, matrix substances, such as milk caseins, which account for >80% of bovine milk proteins, can interfere with the isolation process and the lack of accuracy in separating exosomes from different constituents represents the main limitation of available techniques [31].
The most widely used method to isolate milk exosomes is differential ultracentrifugation, which is considered to be the current “gold standard” technique [31,32]. Sequential ultracentrifugation is needed to separate exosomes from other colloidal structures, and it is usually followed by an additional gradient step (i.e., sucrose gradient) [33,34]. Density gradient ultracentrifugation yields low protein contamination and high purity, but it is a time-consuming process and might be unsuited for large-scale production due to its low sample throughput and potential negative impact on exosome integrity [33,35].
Size-exclusion chromatography (SEC) has also been successfully implemented to isolate exosomes from different origins. SEC relies on the differential elution profiles of different particles running through a stationary phase depending on their size [36]. This technique has been used to isolate bovine milk exosomes improving both yield and time efficiency compared to differential ultracentrifugation [25,37]. On this subject, a method for isolating EVs using a size-exclusion chromatography column in combination with turbidimetry was developed and applied to cow milk samples [38]. SEC presents practical advantages over differential ultracentrifugation, being simpler, less expensive, and reusable [36]. Nonetheless, a recent report suggested that SEC might be incapable of differentiating exosomes from other milk components when assessing industrially processed milk samples [39]. The presence of other contaminants and different EVs subsets might compromise SEC analyses in exosomes studies [31,37]. Overall, most authors favor using a combination of methods to ensure an adequate isolation process [40]. Recently, the combination of ultracentrifugation and SEC, or tangential flow filtration (TFF), along with divalent cation chelation with EDTA, was shown to produce large amounts of pure milk EVs isolates, thus constituting a promising approach [40].
Milk caseins represent an important obstacle to the isolation of milk exosomes. Methods based on acidification have been explored to cause isoelectric precipitation of milk caseins, thus enabling their separation from other milk components [34,41]. Acidification enables faster isolation of milk exosomes and yields significantly higher amounts compared to differential ultracentrifugation [32]. Accordingly, this process has been proposed as the most suitable step to remove caseins [42]. However, some authors have reported deterioration of exosomes’ surfaces after acid treatment [34,41].
Commercial kits were introduced to the market to isolate EVs with similar results to those obtained by ultracentrifugation techniques without requiring specialized equipment [31]. Among these, exosome precipitation solutions (ExoQuickTM, System Biosciences, Palo Alto, CA, USA), and membrane affinity spin columns (ExoEasyTM, QIAGEN, Hilden, Germany) have been applied to milk extracellular vesicles [43,44,45,46,47]. These kits provide rapid EVs isolation but are susceptible to high protein contamination and can promote exosome aggregation [31,48]. Particularly, the incapacity to differentiate different EVs subsets is a common disadvantage of most available methods. Newer techniques, such as asymmetrical flow field-flow fractionation (AF4), have been shown to enable the differentiation between different EVs populations up to 1 nm increments [48,49], albeit low yield and high costs may hinder their implementation [50].
The characterization of milk exosomes is crucial to evaluating the heterogeneity of isolated preparations [6]. Diverse methods are available for this purpose. Transmission electron microscopy (TEM) and cryo-electron microscopy (Cryo-TEM) permit the characterization of exosomes by their size and distribution and are widely used in exosomes research [31,51]. In the same line, adapted high-resolution flow cytometry protocols were demonstrated to enable the characterization and quantification of milk exosomes [52]. These protocols are typically used in conjunction with TEM or cryo-TEM in milk exosome studies [53,54]. Other techniques, such as nanoparticle tracking analysis (NTA), which is usually used in combination with fluorescence detection, can effectively measure particle size, distribution, and concentration in liquid media [31,55] and are applicable to validate the isolation of milk exosomes [56]. Similarly, dynamic light scattering (DLS) has been used to measure the size distribution of milk exosomes [57,58]. However, it is possible that DLS cannot differentiate populations with different sizes, being useful only to evaluate populations of monodisperse samples [29,31]. Large-scale and cost-effective isolation of milk exosomes constitutes a bottleneck for their successful implementation as bioactive ingredients and methods available so far might not be suited for this purpose [59].
As commented, milk exosomes can be characterized based on enriched marker proteins such as tetraspanins (i.e., CD9, CD63), trafficking molecules (i.e., TSG101, ALIX), and chaperones (i.e., HSC70, HSP60) [42,60]. Detection of these markers by Western-Blot is customary in milk exosomes studies [43,61,62]. Finally, the combination of specific antibodies and atomic force microscopy (AFM) provides better resolution than other imaging techniques but requires specialized equipment and has limited throughput [31,42]. Remarkably, given the limitations of existing techniques, the combination of different methods, including microscopy, flow cytometry, and Western blotting, should be considered to ensure an adequate characterization of isolated exosomes [60].
3.2. Milk Exosomes Composition and Cargos
Milk exosomes harbor various cargos with potential roles in intercellular communication, metabolism, and nutrition. Beyond tetraspanins, exosomes can contain many different proteins, including membrane transport proteins, cell adhesion proteins, signaling proteins, enzymes, cytoskeletal proteins, and chaperones [15,63]. Furthermore, exosomes from bovine colostrum were shown to be significantly enriched in proteins implicated in the growth and immune response [64]. Among the most consistently reported proteins of milk exosomes are the lactadherin [65], a glycoprotein that has been observed to play a role in maintaining the intestinal epithelium [66,67]. Besides, bovine milk exosomes were shown to carry bioactive transforming growth factor-β1 (TGF-β1), a cytokine with important immunomodulatory properties [68,69].
Contrary to proteins, the lipid composition of exosomes remains largely unexplored. Lipidomic analyses of milk-derived exosomes have revealed that phosphatidylcholine (PC), phosphatidylserine (PS), phosphatidylethanolamine (PE), and sphingomyelin (SM) are among the most enriched lipids found in these vesicles [25,70,71]. PC has been proposed to ameliorate inflammation via the gut–brain axis in vivo [72]. Likewise, PE might present hypocholesterolemic effects reducing intestinal lipids absorption and SM has been proposed to promote myelin formation and brain neuroplasticity, thus potentially playing a beneficial role in infant neurodevelopment [73,74]. Evidence from human trials attributes a positive effect to MFGM lipids in growth, intestinal health, and infant neurodevelopment [75]. These effects are usually attributed to polar lipids, which are also abundantly expressed in milk exosomes [70]. However, research on the beneficial effects of the lipidic fraction of milk exosomes is lacking and constitutes an interesting avenue to explore [75].
Among exosome components, nucleic acids have attracted the greatest interest due to their biological effects on regulating metabolic processes [5]. Different nucleic acids have been described in milk exosomes, including DNA, mRNA, miRNA, circular RNA, long non-coding RNA, etc. In particular, milk represents one of the richest sources of miRNA [32,76]. MiRNAs are defined as 17–24-nucleotide small noncoding RNA fragments which are responsible for post-transcriptional gene silencing by binding to regions of target mRNAs [77]. In animals, miRNA genes are transcribed to pri-miRNAs that are processed by the nuclear RNase III Drosha, and the resulting intermediates are exported out of the nucleus via exportin-5 to be later cleaved by the RNase III Dicer into a miRNA duplex [78]. The resulting miRNA duplex associates with Argonaute (Ago) proteins, which are part of the RNA-induced silencing complex (miRISC), leading to the removal of the passenger strand and the final maturation of the miRNA [79]. However, miRNAs can also be loaded into exosomes in a completely independent pathway which might implicate multiple mechanisms, namely recognition by hnRNPA2B1 and hnRNPA1 RNA-binding proteins [80,81], an affinity for different cell membrane lipids [82], recognition of specific RNA motifs and configurations, etc. [83]. These processes are illustrated in Figure 2.
Including milk, biological fluids constitute an aggressive environment for miRNA viability. Exosomes protect miRNAs against low pH and RNases, thus enabling their delivery to target cells [35,84,85]. Furthermore, exosomes have been shown to confer protection against the adverse conditions present in the digestive tract, which suggests that the oral intake of these compounds facilitates the availability of their cargos [86,87,88]. Besides protecting their cargos, glycoproteins displayed in milk exosomes facilitate their recognition and uptake by the target cell receptors [70,89].
Interestingly, milk exosomal miRNAs are highly conserved across mammalian species, including humans [90]. A remarkable number of these milk miRNAs are considered to be related to the immune system [91,92]. Particularly, miR-155 has been shown to regulate T cells development and homeostasis [93,94]. Other miRNAs, such as Let-7c, miR-17, miR-30, miR-92, miR-148a, and miR-223, have been linked to the regulation of different populations of immune cells and inflammatory processes [95,96]. Synapse localization is strongly enriched among genes targeted by miRNAs abundantly expressed in milk exosomes, which might positively affect brain development during early life [97]. MiR-181a-5p has been consistently reported to be among the most highly expressed miRNAs in milk exosomes [98,99], and it is considered an antiatherogenic miRNA that downregulates NF-κB activation and vascular inflammation [100,101]. Hence, milk exosomes may promote the transfer of miRNAs from the mother to the offspring, playing a role in the early development of the immune system and mediating different positive health effects [102].
3.3. Bioavailability, Bioaccessibility, and Bioactivity of Milk Exosomes
The capability of milk exosomes to exert biological effects relies on their bioaccessibility and bioavailability after oral intake [103]. Bioaccessibility would consist of the ability of the exosomes and their cargos to survive gastrointestinal digestion. As already mentioned, strong evidence provided by in vitro human digestion models supports that milk exosomes resist gastrointestinal/pancreatic digestion processes [99,104,105,106]. This fact is even more important in infants, given their lower stomach acidity. In this sense, the free synthetic miRNAs that are added to milk are rapidly degraded by an acidic environment and RNases, while disruption of exosomes by ultrasonication releases miRNA content, thus leading to similar effects [107]. Milk exosomes not only protect miRNAs from low pH and RNases but may also exert a protective effect on cargo proteins from proteases [106].
Storage conditions, freezing, and thawing can compromise exosomes stability, negatively influencing their count in milk samples [108,109]. Nonetheless, some studies have observed that exosomes are very stable in harsh conditions. In this sense, it has been shown that exosomes contained in commercial bovine milk harbor miRNAs and immunoregulating proteins such as TFG-β after pasteurization [68]. However, different reports suggest that common industrial processing might compromise commercial milk exosome integrity, thus drastically reducing their count in products such as UHT milk or infant formulas [53]. Similarly, dairy products subjected to fermentation processes and consumer handling (i.e., microwaves) may show a significant decrease in exosome protein and miRNA content [107,110]. Processes such as fermentation may induce exosome lysis and increase RNase levels due to microbial production [107]. In the same line, infant formulas are also subjected to harsh conditions during their production, and thus, the addition of exosomes as an ingredient to resemble the characteristics of human milk must overcome challenges related to industrial processing and storage [111]. These aspects are even more relevant in hydrolyzed formulas, which are subjected to more severe processing conditions [112].
Bioavailability implies gastrointestinal absorption and the subsequent presence in the systemic circulation [103]. Studies evaluating the effects of gastrointestinal digestion on exosomes have also analyzed the uptake of exosomes and their cargos by human intestinal cells in vitro, yielding positive results [99,105]. Furthermore, studies conducted in humans have observed increased concentrations of certain miRNAs in blood mononuclear cells after the consumption of milk [113]. This fact has predisposed authors to evaluate the potential of milk-derived exosomes as carriers to facilitate oral drug administration [114]. Notably, Manca et al. evaluated the bioavailability of fluorescently labeled bovine milk exosomes and their labeled miRNAs in mice, observing that both exosomes and miRNAs were bioavailable after exosomes oral intake and intravenous administration, and were accumulated in different organs, although the distribution of miRNAs showed a unique profile [87]. Moreover, studies have reported that bovine milk exosomes are assimilated by gut bacteria in mice, thus modulating gut microbiota, suggesting that exosomes are involved in a potential cross-talk between animals and bacteria of different species [115,116]. Regarding internalization mechanisms, intestinal cells uptake exosomes through a glycoprotein-mediated endocytosis process. Accordingly, eliminating glycoproteins using proteinase K or trypsin in human and rat intestinal cells leads to a significant decrease in exosome uptake [117]. Similar results are obtained when intestinal cells are treated with endocytosis inhibitors and carbohydrate competitors [117]. Different cells can uptake exosomes by a variety of proposed mechanisms, including clathrin-dependent endocytosis, caveolin-mediated uptake, micropinocytosis, phagocytosis, etc. [118] In this regard, a recent study reported that milk exosomes are phagocyted by mice macrophages through a process mediated by class A scavenger receptors, which is important when assessing their potential as delivery vehicles [119]. Overall, the uptake mechanisms depend on the exosomes source and the type of recipient cell, and several mechanisms may co-exist [8].
As discussed, milk exosomes carry proteins, lipids, and nucleic acids with the potential to regulate immunity, growth, and development [103]. The bioavailability and bioaccessibility of milk exosomes modulate their bioactivity, defined as their capacity to exert functional effects. Remarkably, current scientific evidence suggests that milk exosomes can survive harsh conditions and be bioavailable after oral ingestion. Henceforth, it seems reasonable to evaluate the beneficial effects that these bioactive compounds can mediate in health and metabolism when orally consumed (Figure 3).
4. Beneficial Effects of Bovine Milk Exosomes on Overall Health
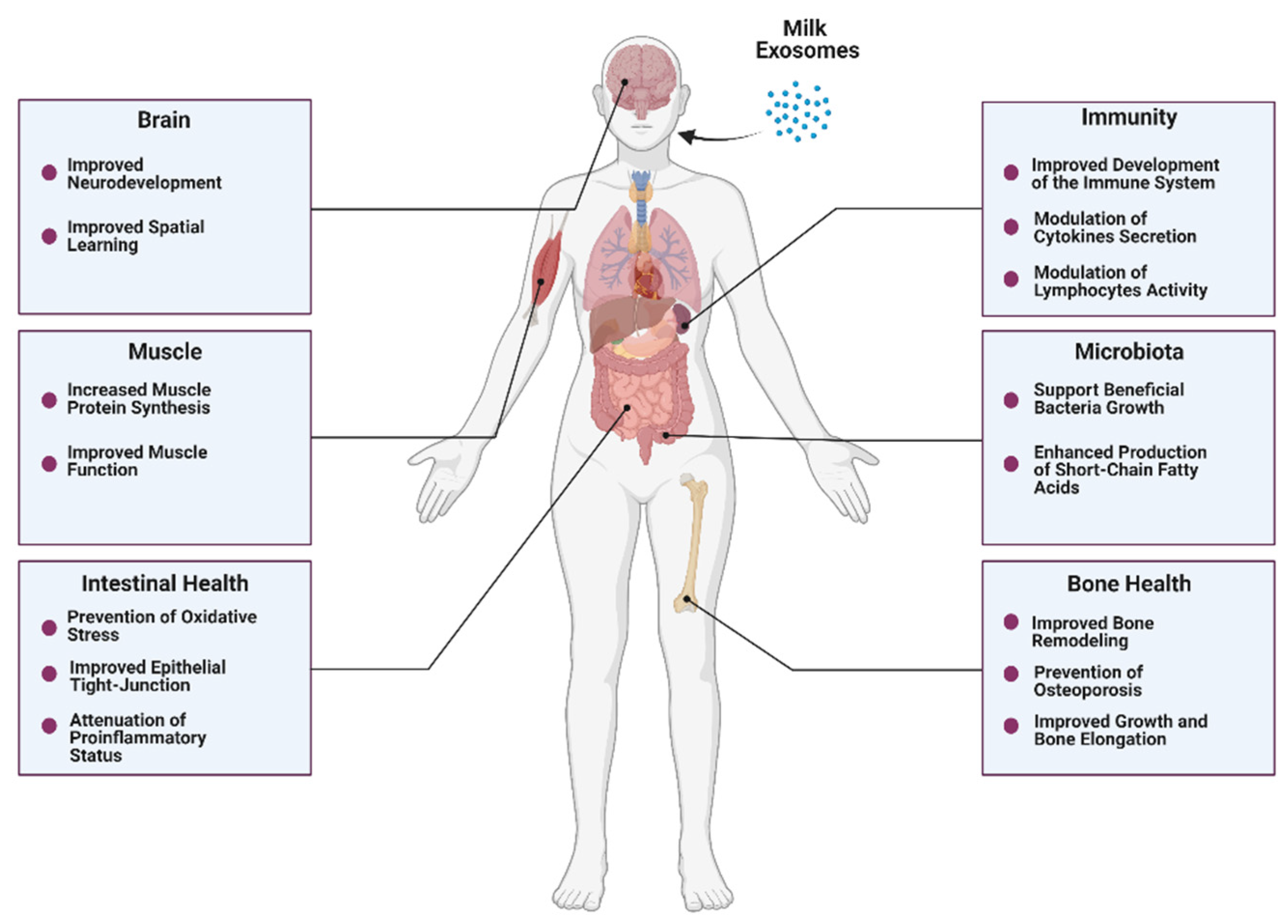
4.1. Intestinal Health
A growing body of scientific evidence indicates that milk exosomes survive digestion processes and are assimilated by intestinal cells via endocytosis, being bioavailable systemically [87,88,99,117,120]. Nonetheless, milk exosomes can also elicit direct effects on intestinal cells when taken up. In this regard, Martin et al. reported that human milk exosomes (HME) exerted a protective effect against H2O2-induced oxidative stress in intestinal epithelial cells [121]. Ensuing studies endeavored to elucidate the underlying mechanism of action. On this subject, Dong et al. exposed intestinal stem cells to H2O2, documenting that the addition of HME led to increased cell viability, which was proposed to be mediated by the upregulation of the highly conserved Wnt/β-catenin axis [122]. Hence, HME might exert beneficial effects on intestinal oxidative stress, which is a key feature of necrotizing enterocolitis (NEC) and intestinal bowel disease (IBD) [123]. Briefly, NEC is a severe intestinal pathology that develops during infancy, whereas IBD is a group of adult chronic disorders, including ulcerative colitis and Crohn’s disease, characterized by inflammatory processes [124]. However, both NEC and IBD share common pathological features, namely compromised intestinal tight junctions, reduced mucosa layer, and increased susceptibility to bacterial components, such as lipopolysaccharides (LPS) [125], and milk exosomes have been reported to convey beneficial effects on various of these aspects.
Both raw and pasteurized human milk exosomes have been shown to reduce the injury caused by hypoxia and LPS in mouse organoids while attenuating the expression of proinflammatory IL-6 in vitro [62]. Similarly, gavage administration of these exosomes to a mouse NEC model led to reduced mucosal injury and inflammation and an increased number of goblet cells and mucosa production regardless of the pasteurization process [62]. Moreover, He et al. evaluated the effects of HME obtained from mothers who delivered term preterm infants on in vivo and in vitro NEC pathological scores. They observed that HME protected against the LPS insult and increased the expression of epithelial tight-junction proteins. Regarding animal models, pretreatment with HME led to reduced intestinal mucosal damage, lower levels of IL-6 and TNF-α, and increased expression of epithelial tight-junction proteins. These effects were independent of the delivery status [126]. Other authors have reported similar effects of HME on the incidence and severity of NEC in rat models [70,127]. In particular, Chen et al. observed that HME isolated from women who gave birth to term or preterm newborns exerted a protective effect on NEC by decreasing the severity of the injury and stimulating the proliferation and migration of intestinal epithelial cells independently of the delivery status [70]. The authors further analyzed the lipidomic profile of HME, finding that term and preterm milk exosomes displayed almost identical lipid compositions. Furthermore, bioinformatic analyses suggested that HME’s most abundant lipids were related to the ERK/MAPK pathway, which is proposed to mediate LPS injury in intestinal cells [70]. Therefore, HME not only reduces oxidative stress but also positively contributes to intestinal epithelial integrity while decreasing intestinal inflammation in NEC models.
In the light of these studies, it can be proposed that exosomes may confer therapeutic properties to human milk in NEC situations. Notably, feeding human breast milk is demonstrated to reduce the incidence of NEC compared to formula feeding [128]. However, mothers delivering preterm and low birth weight infants may be unable to initiate milk expression, thus making donor human milk necessary to ensure adequate lactation [129]. In this context, some authors have evaluated the role that milk exosomes obtained from different animal sources play in NEC as an alternative to human milk. The concomitant administration of bovine milk exosomes to NEC-induced mice was shown to protect against intestinal injury, increase the number of goblet cells, and improve endoplasmic reticulum function in vivo [86]. Similarly, bovine milk exosomes were demonstrated to induce cell proliferation, protect against oxidative stress caused by H2O2 while inhibiting Nrf2 and H01 gene expression, and improve purine nucleotide catabolism and energy status in rat intestinal crypt epithelial cells [110,130,131]. In the same line, milk exosomes originating from different species, including mice, pigs, and yak, have been demonstrated to exert beneficial effects on the features of NEC [45,132,133,134]. The addition of different nutritive compounds to infant formulas to prevent NEC development has been shown to be largely unsuccessful, and further insights into which bioactive human milk compounds are responsible for its effects on NEC prevention are required [135]. In the light of the literature, the effects of bovine milk exosomes on NEC features might resemble those of HME, thus constituting a promising new ingredient for infant formulas to address the development and treatment of NEC.
Regarding IBD, some authors have evaluated the role that milk-derived exosomes play in the characteristics of these intestinal disorders denoted by inflammatory phenotypes. Among them, Wu et al. observed that the reduction of bovine milk exosomes and their miRNA content in diets administered to Mdr1a knockout mice led to exacerbated IBD symptoms compared to exosome/miRNA-sufficient diets [136]. An ensuing pilot study conducted in a different genetic mouse model of ulcerative colitis observed that the administration of bovine milk exosomes improved the macroscopic colitis histopathological scores of treated mice compared to control [137]. Benmoussa et al. evaluated the administration of milk exosomes on colitis outcomes in a dextran sodium sulfate (DSS) -induced colitis mouse model. The authors concluded that milk exosomes partially restored intestinal impermeability, recovered mucin secretion, improved histology scores, reduced colon shortening, and prevented weight loss. In particular, milk exosomes were shown to downregulate the expression of several colitis-associated microRNAs, namely miR-21, miR-29b, and miR-125b [138]. Likewise, Reif et al. reported that the oral administration of exosomes isolated from cow and human milk exerted similar positive effects attenuating the severity of colitis symptoms, reducing IL-6 and TNF-α expression, and downregulating DNMT1 and DNMT3 methyltransferases in DSS-induced colitis mice [69]. On this subject, a recent study conducted by Tong et al. provided further evidence on the beneficial effects of bovine milk exosomes on wide aspects of ulcerative colitis. The authors observed that the treatment of RAW264.7 cells with different concentrations of bovine milk exosomes inhibited inflammatory responses mediated by TLR4-NF-κB and NLRP3 pathways. These results were replicated in vivo in a mouse model of ulcerative colitis. Moreover, the administration of bovine milk exosomes improved wide aspects of cytokine homeostasis, immune response, and modestly improved gut microbiota profile [139]. Thus, bovine milk exosomes constitute a promising therapeutic tool in managing intestinal inflammatory disorders.
Finally, a recent report suggested a potential application of bovine milk exosomes in the management of malnutrition. Exosomes were administered to mice fed a 1% protein diet and were shown to improve defective intestinal epithelial permeability and architecture induced by malnutrition [140]. Further studies are warranted to evaluate the therapeutic value of bovine milk exosomes in malnourished individuals.
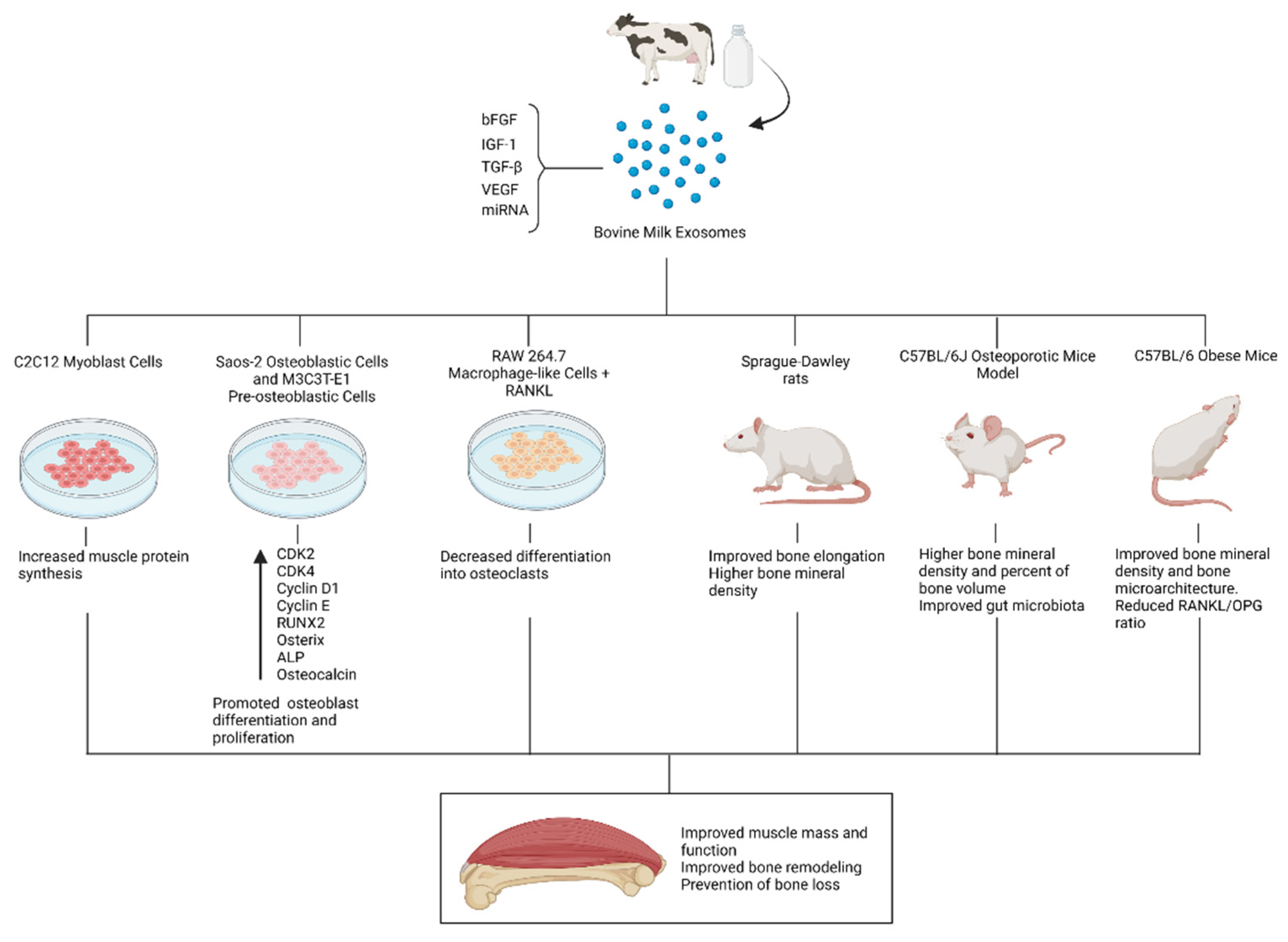
4.2. Bone and Muscle Metabolism
Bone remodeling is a dynamic process characterized by consecutive cycles of bone resorption and formation regulated by specialized cells, namely osteoclasts and osteoblasts [141]. While the attachment of osteoclasts to the bone surface leads to bone resorption mediated by acidification and proteolysis processes, the osteoblasts are responsible for bone formation mediated by tightly regulated processes of matrix production and bone mineralization [141]. Bone remodeling is controlled at the systemic level by hormones responsible for maintaining calcium homeostasis, mainly parathyroid hormone and vitamin D metabolites [141]. Milk is not only a rich source of calcium and vitamin D3 but also contains different minerals and proteins, such as lactoferrin and whey proteins that are involved in bone remodeling processes [142,143,144,145]. Although different mechanisms support a positive role of milk intake in bone health and the prevention of bone fractures, some studies have failed to prove these effects [146]. Nonetheless, milk consists of a complex matrix, and the health effects of individual components might not be completely extrapolated [147]. In this sense, some authors have focused on milk extracellular vesicles to identify milk bioactive compounds with beneficial effects on bone formation. In particular, Oliveira et al. orally administered two different concentrations of bovine milk exosomes (4.7 × 106/mL and 14.3 × 106/mL) to mice for seven weeks observing that treated mice displayed increased osteocytes number and woven bone formation compared to control (PBS) [148]. Furthermore, exposing human mesenchymal stem cells (hMSCs) to bovine milk exosomes led to increased osteoblast differentiation. In an ensuing study, Oliveira et al. showed that milk exosomes treatment on murine bone marrow cells induced an increase in osteoclast differentiation along with an inhibition of the osteoclast activity [149]. Recent research conducted by Go et al. studied the addition of bovine milk exosomes to human osteoblastic Saos-2 cells in vitro, observing an increase in Saos-2 cells proliferation in a time-dependent and dose-dependent manner while promoting the expression of the osteoblast transcription factors RUNX2 and Osterix [150]. Moreover, treatment of Saos-2 and MC3T3-E1 pre-osteoblastic cells with milk exosomes increased alkaline phosphatase (ALP) and osteocalcin (OCN) levels, which are involved in extracellular matrix synthesis processes [150]. Finally, the authors administered 50 mg/kg/day of exosomes to rats for 14 days and reported that the treatment was associated with significantly increased bone mineral density in trabecular and cortical tibiae [150]. In the light of these studies, bovine milk exosomes could constitute food components with the potential to influence different aspects of bone remodeling.
The aforementioned effects of bovine milk exosomes in osteogenesis lay the groundwork for the study of their potential application in settings of bone loss and osteoporosis. Oliveira et al. explored the osteoprotective properties of bovine milk exosomes in obese and ovariectomized mice. The treatment with bovine milk exosomes contributed to a reduced RANKL/OPG ratio in mice fed a high carbohydrate diet, which denotes decreased osteoclast differentiation and activity. In ovariectomized mice, the administration of bovine milk exosomes prevented the loss of mechanical resistance and improved several features of femur microarchitecture [151]. In addition, the administration of exosomes was also associated with decreased number and activity of osteoclast cells in ovariectomized mice by reducing the local and systemic RANKL/OPG ratio [151]. Other authors have explored the use of bovine milk exosomes to improve bone health in the context of osteoporosis. Yun et al. treated MC3T3-E1 and RAW 264.7 cells with bovine colostrum exosomes. No cytotoxicity was evident in MC3T3-E1 cells exposed up to 500 ng/mL of exosomes, while 100 ng/mL was enough to enhance cell proliferation. Regarding the RAW 264.7 cells assay, exosomes were shown to inhibit osteoclast differentiation after exposure to RANKL and macrophage-colony-stimulating factor [98]. Furthermore, 8-week treatment with colostrum exosomes before the onset of steroid-induced osteoporosis exerted a positive effect on the femoral bone volume and bone mass density [98]. Thus, bovine milk exosomes appear to be a promising element to explore the prevention of bone loss and osteoporotic processes.
Milk intake has been associated with improved linear growth in various observational and interventional studies [152,153,154]. In the same line, milk allergy has been linked to a lower final height in adulthood [155]. Not only has milk intake been associated with linear growth, but it also may be beneficial for malnourished children undergoing catch-growth [145]. Importantly, relationships between growth and milk intake can be subjected to reverse causality and residual confounding. Notwithstanding this fact, the potential effects of milk on stature growth have been attributed to different milk components, such as proteins, minerals, and prebiotic carbohydrates [156]. These effects are usually proposed to be mediated through the stimulation of IGF-1 synthesis, which constitutes a major regulator of growth and bone elongation [157,158,159]. Interestingly, some authors have proposed that breastfeeding is linked to higher IGF-1 levels later in life, which implies a programming effect of breastmilk on linear growth patterns [160]. MiRNAs abundantly expressed in milk exosomes, namely miRNA-148a and miRNA-29, have been hypothesized to induce IGF-1-mediated growth [118,161].
Bone and muscle constitute a functional unit [162]. Not only adequate bone remodeling but also an appropriate muscle function are desirable to support a healthy musculoskeletal status. Nutritional interventions targeting both bone mass and muscle function may have profound implications for mobility and quality of life. In this regard, milk contains valuable proteins with important anabolic effects in terms of muscle protein synthesis [163]. Nonetheless, different milk components might contribute to these anabolic effects. On this subject, Mobley et al. explored the effects that exosomes isolated from the whey fraction of bovine milk had on features of muscle anabolism in vitro. Seeding C2C12 myoblast cells with milk exosomes led to a significant increase in muscle protein synthesis 12 and 24 h after treatment. Although mTORC1 was not affected 6–24 h after the treatment, a short-term activation could not be discarded [164]. The following study evaluated the administration of bovine milk exosomes to C57BL/6 mice in vivo, observing only modest effects on gene expression and amino acid content in treated animals with no relevant impact on muscle strength [165]. These results were in line with those obtained by Parry et al., who documented that a bovine milk exosome-depleted diet elicited anabolic and transcriptomic effects in rat muscle, whereas an exosome-sufficient diet was not associated with any anabolic outcome [166]. Overall, in vitro results suggested a potential role for bovine whey exosomes in muscle anabolism, but evidence from in vivo studies is conflictive. Differences in tissue distribution patterns of milk exosomes might be responsible for these heterogeneous results [165,166], yet cross-communication between different organs should not be discarded [33]. Promising in vitro results should not be eclipsed by these discrepancies, and further research on this subject is warranted to elucidate the potential role of milk exosomes in muscle anabolism. A summary of the preclinical evidence on the potential beneficial effects of bovine milk exosomes on musculoskeletal health is illustrated in Figure 4.
4.3. Immunity
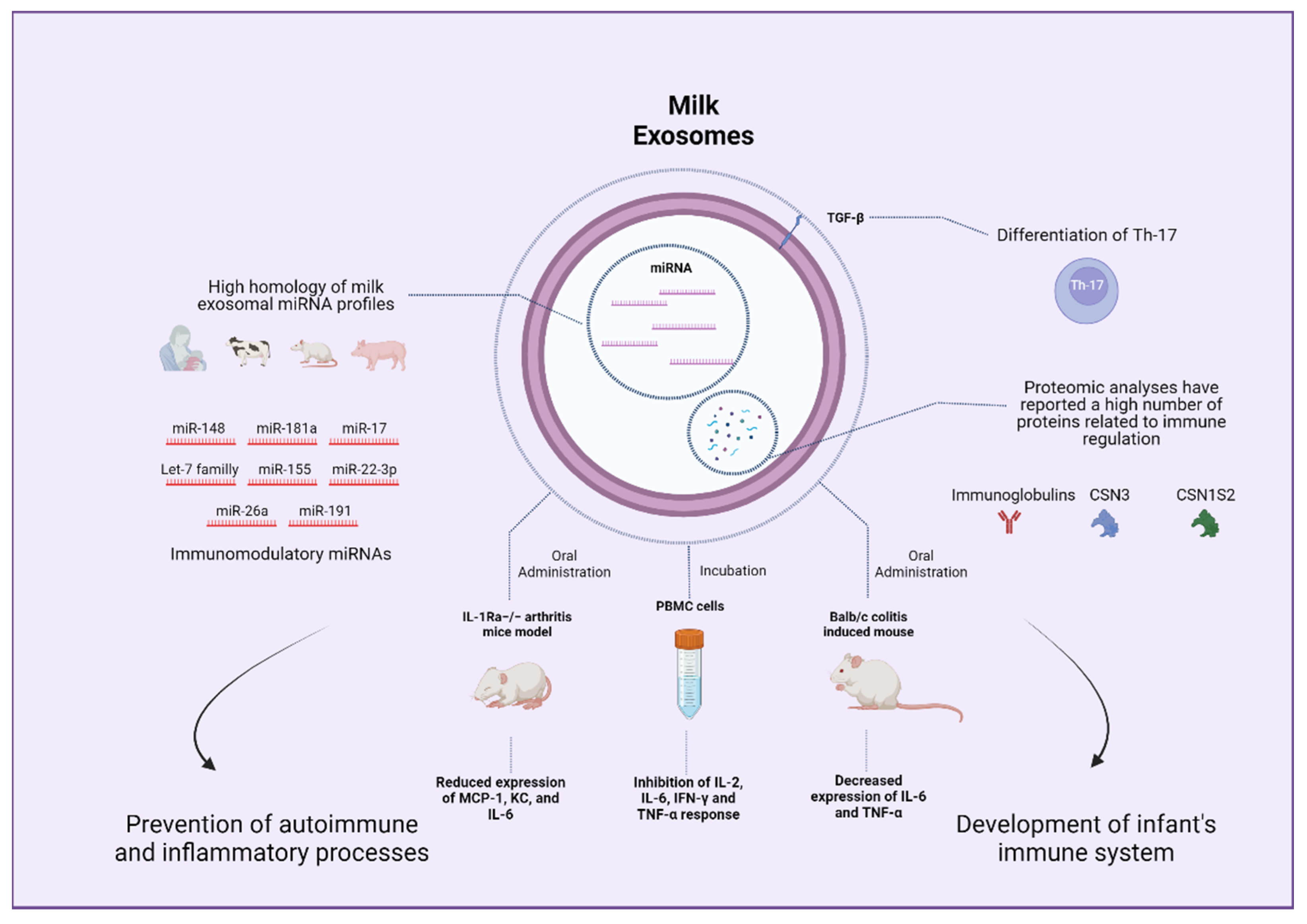
Breast milk is a dynamic composition of nutrients and bioactive factors that changes throughout lactation according to the needs of the developing infant [167]. A solid body of literature supports that breastfeeding confers protection against infections, allergic diseases, and inflammatory processes in infants [168,169,170,171]. In this sense, a wide number of breast milk components present immunoregulatory properties and might contribute to these effects, including immunoglobulins, oligosaccharides, glycoproteins, living cells from the mother, and probiotic microorganisms, among others [167]. Exosomes are among the pool of bioactive compounds with immunoregulatory features identified in milk. Admyre et al. reported the presence of exosome-like vesicles in human breast milk, which, when incubated with peripheral blood mononuclear cells (PBMC), led to inhibited production of IL-2, IFN-γ, and TNF-α along with increased production of IL-5 [61]. These effects were proposed to be mediated by the regulation of T-lymphocytes response [61]. Authors postulated that these effects might contribute to the benefits of breast milk in the development of the infant’s immune system. Kosaka et al. examined milk exosomes cargos observing that these vesicles contained miRNAs with the potential to induce B-lymphocytes differentiation, namely miR-181 and miR-155 [172]. Following an in-depth analysis of human milk exosomal miRNA reported that immune-related miRNAs are present in high quantities in this medium with low variation among healthy individuals [92]. Nowadays, it is well-accepted that milk exosomes carry a notable number of miRNAs with potential immunomodulatory effects [173]. The potential immunomodulatory properties of milk exosomes might be at least partially responsible for the already discussed benefits on intestinal health and other health outcomes [174].
Interestingly, not only human breast milk but also milk from different mammals display a high miRNA content, and exosome immune-related miRNA profiles are similar across species [175]. In particular, miRNAs members of the let-7 family and miR-148a were shown to be highly conserved in milk exosome samples obtained from a wide variety of different mammals and have been implicated in several aspects of the immune function [90,120,176,177]. More precisely, members of the miR-148/152 family were demonstrated to inhibit the TLR-mediated expression of MHC II and cytokine production by targeting CaMKIIα in dendritic cells [178]. MiRNA-148a has also been linked to the regulation and function of lymphocytes B and T and might play a role in the prevention of inflammatory and autoimmune disorders [179]. Similarly, members of the let-7 miRNA family play an important role in regulating TLR4 signaling, contributing to host defense responses in settings of infection [180]. Furthermore, let-7 has been linked to processes of macrophage activation and modulation of the adaptative immune response [180]. Recently, immune-related miRNAs, such as miR-181a, miR-26a, and miR-191, were found to be among the most abundantly expressed in colostrum and mature bovine milk exosomes [98]. Hence, it can be inferred that miRNAs present in milk exosomes from different species might serve the purpose of contributing to the development of the infant’s immune system.
Different cargos might also contribute to the immunomodulatory properties of milk exosomes. Regarding proteins, Benmoussa et al. analyzed possible pathways impacted by proteins contained in milk exosomes observing that identified proteins were associated with different immunity pathways related to neutrophil degranulation and the innate immune system, among other aspects [181]. Specifically, Pieters et al. observed that transforming growth factor-β (TGF-β) was consistently expressed in exosomes isolated from commercial semi-skimmed cow milk [68]. TGF-β cytokine is crucial for the differentiation of Th17 cells [182]. To assess exosomes activity, authors co-incubated murine spleen T-cells with a Th17 differentiation cocktail replacing TGF-β with milk exosomes, confirming the differentiation of Th17 cells, which was repressed by the blockade of TGF-β using monoclonal antibodies [68]. These results aligned with a study evaluating similar settings [183]. Likewise, another study reported TGF-β1 as a cargo of both cow and human milk exosomes [69]. Curiously, one report on proteomic and functional enrichment analysis of bovine milk exosomes suggested that immune-related proteins are more abundant in colostrum compared to mature milk [64]. Nonetheless, a recent proteomic analysis of late-stage lactation bovine milk exosomes revealed a large number of proteins that were mostly related to metabolism and immune system pathways [184]. These findings underscore that not only nucleic acids but also proteins might contribute to the immunomodulatory properties of exosomes isolated from different types of milk. Finally, as commented, data regarding the lipid content of exosomes and its potential effects on immune outcomes are limited and might constitute an interesting field to explore.
Research on the regulatory effects of milk exosomes in different immune processes has been reported. In this regard, different macrophage cell lines have been consistently shown to take up milk exosomes. A recent study observed that the treatment of RAW264.7 macrophages with bovine exosomes before LPS stimulation contributed to a decreased inflammatory response and a downregulated cytokine secretion mediated by inhibited NF-κB pathway [185]. Other studies conducted in RAW264.7 cells have demonstrated that bovine milk exosomes stimulate macrophages proliferation without stimulating nitric oxide or proinflammatory cytokines production while promoting the expression of proteins involved in cell cycle and proliferation [186]. Furthermore, bovine milk exosomes were shown to exert a protective effect against the cytotoxic action of cisplatin, a chemotherapeutic drug, in RAW264.7 cells [186]. Regarding immune effects on different cells, Arntz et al. pre-incubated mice splenocytes with different concentrations of exosomes (20 and 200 μg/mL) to later stimulate the cells with LPS. Exosomes treatment led to a pronounced reduction in proinflammatory markers TNF-α and monocyte chemoattractant protein-1 (MCP-1) in response to the LPS insult compared to control [183]. Furthermore, IL-1Ra knockout mice, which display symptoms of polyarticular arthritis, were administered different doses of bovine milk exosomes. Mice treated with 1200 μg/mL of exosomes showed a considerable delay in arthritis onset and reduced arthritis symptoms, which implies a potential role of bovine milk exosomes in autoimmune and inflammatory diseases [183]. Interestingly, milk exosomes have been suggested to confer protection against HIV infection. Milk exosomes were shown to express soluble mucin 1 (MUC1) and incubation of monocyte-derived dendritic cells (MDDCs) with exosomes was observed to protect against HIV-1 infection in vitro [187]. Moreover, milk exosomes were taken up by MDDCs within 4 h and prevented the transfer of HIV-1 from MDDCs to CD4+ T cells, whereas plasma-derived exosomes did not exhibit any of these effects [187]. Overall, the literature supports an immunomodulatory role of milk exosomes as diet components that may impact an infant’s development and different immune processes (Figure 5).
4.4. Microbiota
EVs have been proposed to facilitate a cross-talk process between the host and the microbiome [188]. Milk exosome microRNAs display unique distribution profiles and are accumulated in the intestine, among other organs [87]. Importantly, a fraction of dietary exosomes and their cargos are not absorbed and can reach further sections of the intestine [87]. Small compounds such as plant-derived nanoparticles, which contain proteins and miRNAs [189], have been observed to be taken up by gut bacteria and elicit changes in the gut microbiota composition [190]. Hence, it seems reasonable to conceive a potential interaction between dietary milk exosomes and the gut microbiota.
Yu et al. characterized three EVs from different sources, namely pasteurized bovine milk, coconut water, and adipose-derived stem cells, and compared their uptake by common representative gut bacteria and analyzed their effects on bacterial growth [191]. The authors observed that exosomes and exosome-like nanoparticles were capable of supporting bacterial growth and modulating gene expression in vitro. Specifically, milk exosomes promoted the growth of E. coli and L. plantarum commensal strains [191]. Zhou et al. evaluated the long-term administration of either bovine milk exosome-sufficient or bovine milk exosome-depleted diets to C57BL/6 mice and analyzed bacterial communities ex vivo. Three phyla, seven families, and 52 operational taxonomic units were differentially abundant between mice allocated to the exosomes-sufficient diet compared to the exosomes-depleted diet group [116]. An ensuing study conducted by Tong et al. evaluated the impact that an eight-week duration treatment with different bovine milk exosomes concentrations had on features of gut microbiota and intestinal immunity in C57BL/6 mice [115]. Authors reported that milk exosomes led to a higher relative abundance of Clostridiaceae, Ruminococcaceae, and Lachnospiraceae compared to control (PBS) [115]. Furthermore, SCFAs production was enhanced in treated animals displaying increased levels of acetate, propionate, and butyrate, and the administration of moderate quantities of milk exosomes correlated to increased levels of IgA and sIgA in mice’s intestine [115]. Notably, a recent study conducted by the same authors documented that the administration of milk exosomes to a DSS-induced colitis mice model restored gut bacteria relative abundance near the levels displayed by control mice [139]. Finally, another recent report confirmed a protective effect of milk exosomes and a miRNA-sufficient diet in the severity of C. difficile infection in C57BL/6 mice when compared to the administration of a milk exosomes and miRNA depleted diet [192]. As commented, milk exosomes elicit immunomodulatory effects and might convey beneficial effects on intestinal health. Hence, the effects of milk exosomes on gut microbiota should be considered within this context. In the light of these results, milk exosomes constitute valuable milk bioactive compounds that might influence wide aspects of intestinal health and microbiota.
4.5. Neurodevelopment
Breastfeeding has been consistently associated with long-term positive cognitive outcomes [193]. Accordingly, epidemiological studies have linked breastfeeding to higher intelligence quotient scores; and improved cognitive, language, and motor skills later in life, compared to formula feeding [193]. Different breast milk components might contribute to brain development and cognitive function [194,195]. Preclinical evidence suggests that milk exosomes and their cargos cross the brain-blood barrier and accumulate in the brain following oral intake [87,196]. On this subject, the depletion of dietary bovine milk exosomes was reported to impair cognitive performance in C57BL/6 mice compared to an exosome-sufficient diet [197]. Mice displayed improved spatial learning and memory when fed the exosome-sufficient diet compared to the exosome-depleted diet [197]. The depletion of bovine milk exosomes and their RNA cargos from the diet was linked to increased hepatic purine metabolites in mice and higher plasma and urine excretion of purine metabolites in humans, which led the authors to speculate that the regulation of purine metabolism by milk exosomes might mediate their positive effects at cognitive level [198]. In this context, it could be proposed that exosomes take part in the neurodevelopmental properties of breast milk. Accordingly, numerous miRNAs contained in milk exosomes, such as miR-148a, miR-141-3p, miR-375, and miR-107, might be involved in nervous system pathways and brain development [199,200]. However, sphingomyelin and other phospholipids present in milk exosomes have been demonstrated to promote neurodevelopment in preclinical research and clinical studies [196]. Furthermore, miRNAs targeting glycosphingolipid biosynthesis pathways were shown to be highly expressed in preterm breast milk, which has a major role in brain development [201]. As commented, different mechanisms support a potential role of milk exosomes in infant neurodevelopment and further research is warranted.
5. Limitations
Milk exosomes are promising ingredients, which can act as drug delivery vehicles, and contain bioactive cargos with the potential to influence different health outcomes. However, some studies have cast doubts on their capacity to elicit significant effects following oral intake. Several preclinical and clinical studies have failed to show a positive association between milk intake and serum miRNA levels [202,203]. Nonetheless, these studies did not specifically focus on exosomal miRNA and aspects related to sample conservation and the use of miRNA reference sequences might compromise their results [103]. In the same line, studies in favor of milk miRNA availability in humans (i.e., [204]) have been subjected to criticism due to methodological concerns [205,206]. On the contrary, preclinical research on the assimilation of exosomal miRNA has yielded more promising results [87,160]. The effects of milk exosomes are at least partially attributed to their nucleic acid content. Yet, miRNAs not only must survive industrial processing and storage conditions but also persist after oral intake, being absorbed, and reaching the targeting cells in sufficient amounts to exert a significant effect [103]. There is a need to evaluate if the promising results observed in animal models can be extrapolated to clinical research. Clinical studies evaluating the effect that the depletion of milk exosomes compared to exosome-sufficient milk has on serum miRNA profile are lacking and might shed light on milk exosome bioavailability. In the same fashion, studies conducted on human organoids would provide insights into exosomes bioactivity [174]. Finally, future studies should consider the integrity of isolated exosomes when assessing their bioavailability after oral intake.
Isolation protocols and characterization methods should be harmonized to ensure the replicability of milk exosomes studies [207,208]. Furthermore, isolation methods should be optimized to enable large-scale production without compromising exosome bioactivity in compliance with good manufacturing practices [88]. The purification of exosomes is important to avoid other milk constituents or contaminants which might jeopardize their quality in preclinical research [174,208], and rigorous compliance with the MISEV guidelines is necessary to attribute milk exosomes effects reported in exosomes studies [209].
Some reviews have raised concerns about the possible deleterious effects of milk exosome intake. Accordingly, milk exosomes would contribute to the development of diabetes, atherosclerosis, osteoporosis, Parkinson’s disease, diverse types of cancer, and even all-cause mortality mediated by different mechanisms such as over-activation of the mechanistic target of rapamycin complex 1 (mTORC1), among others [210,211,212,213,214]. These effects would not be attributed to fermented milk because fermented milk displays reduced exosome miRNA, branched-chain amino acids, and protein content due to bacterial activity [212]. On the other hand, a recent paper by one of these authors extensively disclosed the beneficial effects of milk exosomes on infant intestinal health, β-cells maintenance, bone homeostasis, adipogenesis, and neurodevelopment [118]. These contradictory data suggest that the consumption of non-fermented milk leads to strikingly opposed effects depending on the consumer’s age. However, this argument is not supported by epidemiological studies evaluating milk or dairy products intakes, and some of the proposed pathways are independent of the consumer’s age. According to the FAO, there is no solid evidence regarding the deleterious effects of milk and normal intakes of dairy products on any health outcome [215].
6. Future Perspectives
Due to their high biocompatibility and stability, milk exosomes have been proposed as promising vehicles for the delivery of hydrophilic and lipophilic bioactive compounds [216]. In contraposition to artificial delivery systems, these formulations might present lower immunogenicity, superior bioavailability, increased capacity to cross biological barriers, and the ability to confer protection against harsh conditions, thus improving the biological effects of delivered molecules while reducing their potential toxicity [32,59,217,218]. Specifically, the use of milk exosomes to deliver epicatechin gallate (ECG), a naturally occurring polyphenolic compound with neuroprotective properties, was shown to improve its antioxidative and antiapoptotic effects while inhibiting autophagy in a rotenone-induced model of Parkinson’s disease in vitro, compared to the incubation with free ECG [217]. Similarly, milk exosomes have been explored to improve the bioavailability of different natural compounds with positive results, including curcumin and anthocyanidins [219,220]. Curcumin is a bioactive polyphenol with anti-inflammatory and antioxidative properties, which has been proposed to exert beneficial effects on obesity and protect against neural damage, among other functions [221,222,223]. In the same line, anthocyanidins derived from bilberries have been observed to inhibit T-cell cytokine signaling and IFN-γ transduction in settings of ulcerative colitis [224]. However, these compounds present poor bioavailability after oral intake. Modulating the bioavailability of these compounds is crucial for improving their use as nutraceutical ingredients [225,226]. Milk exosomes represent promising natural delivery vehicles that might enhance the bioavailability of bioactive ingredients contained in these formulations.
To establish milk exosomes as ingredients and delivery vehicles, it is mandatory to overcome some limitations of current isolation and characterization methods, mainly those related to the inability to produce inexpensive and large-scale quantities of milk exosomes [40]. For instance, novel microfluidics-based techniques yield high purity exosomes and are automatable but lack scalability and high sample capacity [59,218]. Optimization of current ultracentrifugation and filtration protocols (i.e., TFF), as well as the use of mixed approaches, including combinations of different characterization techniques, might be best suited for the large-scale production of high-quality milk exosomes [40,227].
Besides the already-mentioned cargos, milk exosomes might contain or be associated with different compounds with potential beneficial effects, such as antibodies, functional lipids, or oligosaccharides. As recently reported by He et al., several milk oligosaccharides, including 2-fucosyllactose with potential prebiotic and immunomodulatory properties [228], were isolated from human colostrum and mature milk exosomes [229]. The in vivo administration of these encapsulated oligosaccharides was shown to modulate macrophages activation in vitro, attenuate adherent-invasive Escherichia coli infection in dextran sodium sulfate mice models in vivo, and prevent LPS-induced inflammation and intestinal damage in these animals [229].
Beyond miRNAs, different nucleic acids contained in milk exosomes have been suggested to play roles in different health outcomes. Long noncoding RNAs (lncRNAs) are transcripts of more than 200 nucleotides that do not translate into proteins [230]. LncRNAs identified in human breast milk exosomes were proposed to be implicated in infant metabolism, neonatal immunity, and development [231]. Not only lncRNAs but also different nucleic acids, such as circular RNAs (circRNAs), covalently closed RNA molecules capable of modulating miRNAs activity, have been reported in milk exosomes [232]. CircRNAs identified in bovine milk-derived exosomes were linked to genes involved in the cytoplasm, endoplasmic reticulum, transport, and transcription factors [232]. In an ensuing study, Zeng et al. performed functional analysis on circRNAs and lncRNAs identified in pig milk-derived exosomes predicting that these would be involved in features of intestinal barrier status, including adherence junction, tight junction, and inflammation [230]. Although further research is needed, some authors have postulated potential roles for milk exosome-derived circRNAs in developing intestine and gut microbiota during early life [233].
Proteomic analyses have concluded that various immunoglobulin components are contained in bovine milk exosomes [26,64]. Interestingly, Betker et al. observed that the “neonatal” Fc receptor (FcRn), a receptor expressed throughout life and involved in IgG binding and transport, participates in milk exosomes uptake [114]. Furthermore, the authors demonstrated the presence of substantial amounts of both bound and unbound bovine IgG in milk exosome isolates [114]. The presence of antibodies among exosomal cargos further supports their immunomodulatory properties.
As we review throughout the present manuscript, milk exosomes have been linked to the prevention of disorders linked to perinatal health such as NEC [134] and might modulate wide aspects of immunity and development, but studies analyzing infant formulae have reported very low quantities of exosomal miRNAs in these formulations [109,234]. Several authors have considered the addition of miRNA-containing milk exosomes to infant formulae as a strategy to reduce the prevalence of intestinal disorders [134,235]. Similarly, the addition of miRNA cargos related to the development of the immune system, such as miR-155, might contribute to the prevention of atopic disease in infants [29]. Other miRNA cargos linked to neurodevelopment, lipid, and glucose metabolism might support adequate infant growth and development [95]. Overall, the benefits of breastfeeding are well-established in the literature, and the natural presence of milk exosomes in breast milk provides a new dimension to perinatal nutrition. Infant formulae might benefit from the addition of these natural compounds as bioactive ingredients. Furthermore, infants and adults might take advantage of the potential positive effects of milk exosomes in areas such as muscle strength, osteoporosis processes, immunity, and intestinal health.
7. Conclusions
The presence of exosomes in breast milk might represent a shift in the paradigm in human nutrition and constitutes a burgeoning field of research. Milk exosomes contain a variety of bioactive cargos, which have been reported to exert beneficial effects at multiple levels, including but not limited to immunity, intestinal and musculoskeletal health, gut microbiota, growth, and development. Further research is necessary to extrapolate to clinical studies the promising results observed in preclinical studies.
Author Contributions
Conceptualization, R.R., J.M.L.-P., J.G.-M., M.D.G. and R.S.; investigation, J.G.-M. and J.M.L.-P.; writing—original draft preparation, J.G.-M. and Í.M.P.-C.; writing—review and editing, R.R., J.M.L.-P., M.D.G. and R.S.; supervision, R.R. and J.M.L.-P.; funding acquisition, R.R. and J.M.L.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. Article processing charges are covered by Granada University.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
J.G.-M., Í.M.P.-C., J.M.L.-P. and R.R. have no relevant interests to declare other than their affiliation with Abbott Laboratories. Abbott produces infant formulas and other pediatric and adult nutritional products.
References
- Witwer, K.W.; Thery, C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J. Extracell. Vesicles 2019, 8, 1648167. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkach, M.; Kowal, J.; Thery, C. Why the need and how to approach the functional diversity of extracellular vesicles. Philos Trans. R Soc. Lond. B Biol. Sci. 2018, 373, 20160479. [Google Scholar] [CrossRef] [PubMed]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug. Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Zempleni, J.; Sukreet, S.; Zhou, F.; Wu, D.; Mutai, E. Milk-Derived Exosomes and Metabolic Regulation. Annu. Rev. Anim. Biosci. 2019, 7, 245–262. [Google Scholar] [CrossRef]
- Lotvall, J.; Hill, A.F.; Hochberg, F.; Buzas, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cell Commun. Signal. 2021, 19, 47. [Google Scholar] [CrossRef]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [Green Version]
- Samanta, S.; Rajasingh, S.; Drosos, N.; Zhou, Z.; Dawn, B.; Rajasingh, J. Exosomes: New molecular targets of diseases. Acta Pharm. Sin. 2018, 39, 501–513. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Zempleni, J.; Aguilar-Lozano, A.; Sadri, M.; Sukreet, S.; Manca, S.; Wu, D.; Zhou, F.; Mutai, E. Biological Activities of Extracellular Vesicles and Their Cargos from Bovine and Human Milk in Humans and Implications for Infants. J. Nutr. 2017, 147, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ExoCarta. Available online: http://exocarta.org/ (accessed on 10 November 2021).
- Dai, J.; Su, Y.; Zhong, S.; Cong, L.; Liu, B.; Yang, J.; Tao, Y.; He, Z.; Chen, C.; Jiang, Y. Exosomes: Key players in cancer and potential therapeutic strategy. Signal. Transduct. Target 2020, 5, 145. [Google Scholar] [CrossRef]
- Andreu, Z.; Yanez-Mo, M. Tetraspanins in extracellular vesicle formation and function. Front Immunol. 2014, 5, 442. [Google Scholar] [CrossRef] [Green Version]
- Lässer, C.; Alikhani, V.S.; Ekström, K.; Eldh, M.; Paredes, P.T.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human saliva, plasma and breast milk exosomes contain RNA: Uptake by macrophages. J. Transl. Med. 2011, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.W.; Nam, M.S. Bioactive Peptides in Milk and Dairy Products: A Review. Korean J. Food. Sci. Anim. Resour. 2015, 35, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Gartner, L.M.; Morton, J.; Lawrence, R.A.; Naylor, A.J.; O’Hare, D.; Schanler, R.J.; Eidelman, A.I. Breastfeeding and the use of human milk. Pediatrics 2005, 115, 496–506. [Google Scholar]
- Timby, N.; Domellof, E.; Hernell, O.; Lonnerdal, B.; Domellof, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 860–868. [Google Scholar] [CrossRef] [Green Version]
- Fox, P.F. Milk: An overview. In Milk Proteins: From Expression to Food; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Hill, D.R.; Newburg, D.S. Clinical applications of bioactive milk components. Nutr. Rev. 2015, 73, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Thaler, J.; Nieuwland, R. Extracellular Vesicles in Human Milk. Pharmaceuticals 2021, 14, 1050. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C. Milk fat globules enveloped by their biological membrane: Unique colloidal assemblies with a specific composition and structure. Curr. Opin. Colloid. Interface Sci. 2011, 16, 391–404. [Google Scholar] [CrossRef]
- Blans, K.; Hansen, M.S.; Sorensen, L.V.; Hvam, M.L.; Howard, K.A.; Moller, A.; Wiking, L.; Larsen, L.B.; Rasmussen, J.T. Pellet-free isolation of human and bovine milk extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2017, 6, 1294340. [Google Scholar] [CrossRef] [PubMed]
- Maity, S.; Bhat, A.H.; Giri, K.; Ambatipudi, K. BoMiProt: A database of bovine milk proteins. J. Proteom. 2020, 215, 103648. [Google Scholar] [CrossRef]
- Reinhardt, T.A.; Lippolis, J.D.; Nonnecke, B.J.; Sacco, R.E. Bovine milk exosome proteome. J. Proteom. 2012, 75, 1486–1492. [Google Scholar] [CrossRef]
- Heid, H.W.; Keenan, T.W. Intracellular origin and secretion of milk fat globules. Eur. J. Cell Biol. 2005, 84, 245–258. [Google Scholar] [CrossRef]
- De la Torre Gomez, C.; Goreham, R.V.; Bech Serra, J.J.; Nann, T.; Kussmann, M. “Exosomics”-A Review of Biophysics, Biology and Biochemistry of Exosomes With a Focus on Human Breast Milk. Front. Genet. 2018, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Buzas, E.I.; Bemis, L.T.; Bora, A.; Lasser, C.; Lotvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J. Extracell. Vesicles 2013, 2, 20360. [Google Scholar] [CrossRef]
- Gandham, S.; Su, X.; Wood, J.; Nocera, A.L.; Alli, S.C.; Milane, L.; Zimmerman, A.; Amiji, M.; Ivanov, A.R. Technologies and Standardization in Research on Extracellular Vesicles. Trends Biotechnol. 2020, 38, 1066–1098. [Google Scholar] [CrossRef]
- Adriano, B.; Cotto, N.M.; Chauhan, N.; Jaggi, M.; Chauhan, S.C.; Yallapu, M.M. Milk exosomes: Nature’s abundant nanoplatform for theranostic applications. Bioact. Mater. 2021, 6, 2479–2490. [Google Scholar] [CrossRef]
- Munir, J.; Lee, M.; Ryu, S. Exosomes in Food: Health Benefits and Clinical Relevance in Diseases. Adv. Nutr. 2020, 11, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Shimizu, K.; Rahman, M.; Ishikawa, H.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Efficient method for isolation of exosomes from raw bovine milk. Drug Dev. Ind. Pharm. 2019, 45, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo-Acebo, L.; Hazas, M.L.L.; Tome-Carneiro, J.; Gil-Cabrerizo, P.; San-Cristobal, R.; Busto, R.; Garcia-Ruiz, A.; Davalos, A. Bovine Milk-Derived Exosomes as a Drug Delivery Vehicle for miRNA-Based Therapy. Int. J. Mol. Sci. 2021, 22, 1105. [Google Scholar] [CrossRef]
- Monguio-Tortajada, M.; Galvez-Monton, C.; Bayes-Genis, A.; Roura, S.; Borras, F.E. Extracellular vesicle isolation methods: Rising impact of size-exclusion chromatography. Cell Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef] [PubMed]
- Vaswani, K.; Koh, Y.Q.; Almughlliq, F.B.; Peiris, H.N.; Mitchell, M.D. A method for the isolation and enrichment of purified bovine milk exosomes. Reprod. Biol. 2017, 17, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Kaddour, H.; Lyu, Y.; Shouman, N.; Mohan, M.; Okeoma, C.M. Development of Novel High-Resolution Size-Guided Turbidimetry-Enabled Particle Purification Liquid Chromatography (PPLC): Extracellular Vesicles and Membraneless Condensates in Focus. Int. J. Mol. Sci. 2020, 21, 5361. [Google Scholar] [CrossRef]
- Hansen, M.S.; Gregersen, S.B.; Rasmussen, J.T. Bovine milk processing impacts characteristics of extracellular vesicle isolates obtained by size-exclusion chromatography. Int. Dairy J. 2021, 127, 105212. [Google Scholar] [CrossRef]
- Marsh, S.R.; Pridham, K.J.; Jourdan, J.; Gourdie, R.G. Novel Protocols for Scalable Production of High Quality Purified Small Extracellular Vesicles from Bovine Milk. Nanotheranostics 2021, 5, 488–498. [Google Scholar] [CrossRef]
- Rahman, M.M.; Shimizu, K.; Yamauchi, M.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Acidification effects on isolation of extracellular vesicles from bovine milk. PLoS ONE 2019, 14, e0222613. [Google Scholar] [CrossRef] [Green Version]
- Cintio, M.; Polacchini, G.; Scarsella, E.; Montanari, T.; Stefanon, B.; Colitti, M. MicroRNA Milk Exosomes: From Cellular Regulator to Genomic Marker. Animals 2020, 10, 1126. [Google Scholar] [CrossRef]
- Yamada, T.; Inoshima, Y.; Matsuda, T.; Ishiguro, N. Comparison of Methods for Isolating Exosomes from Bovine Milk. J. Veter.-Med. Sci. 2012, 74, 1523–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X. Isolation of Extracellular Vesicles from Breast Milk. In Extracellular Vesicles: Methods and. Protocols; Kuo, W.P., Jia, S., Eds.; Springer: New York, NY, USA, 2017; pp. 351–353. [Google Scholar] [CrossRef]
- Hock, A.; Miyake, H.; Li, B.; Lee, C.; Ermini, L.; Koike, Y.; Chen, Y.; Maattanen, P.; Zani, A.; Pierro, A. Breast milk-derived exosomes promote intestinal epithelial cell growth. J. Pediatr. Surg. 2017, 52, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Colitti, M.; Sgorlon, S.; Licastro, D.; Stefanon, B. Differential expression of miRNAs in milk exosomes of cows subjected to group relocation. Res. Vet. Sci. 2019, 122, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Tsukasaki, Y.; Dasgupta, S.; Mukhopadhyay, N.; Ikebe, M.; Sauter, E.R. Exosomes in Human Breast Milk Promote EMT. Clin. Cancer. Res. 2016, 22, 4517–4524. [Google Scholar] [CrossRef] [Green Version]
- Vahkal, B.; Kraft, J.; Ferretti, E.; Chung, M.; Beaulieu, J.F.; Altosaar, I. Review of Methodological Approaches to Human Milk Small Extracellular Vesicle Proteomics. Biomolecules 2021, 11, 833. [Google Scholar] [CrossRef]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef]
- Phillips, W.; Willms, E.; Hill, A.F. Understanding extracellular vesicle and nanoparticle heterogeneity: Novel methods and considerations. Proteomics 2021, 21, e2000118. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef]
- Nolan, J.P.; Duggan, E. Analysis of Individual Extracellular Vesicles by Flow Cytometry. Methods Mol. Biol. 2018, 1678, 79–92. [Google Scholar] [CrossRef]
- Kleinjan, M.; van Herwijnen, M.J.; Libregts, S.F.; van Neerven, R.J.; Feitsma, A.L.; Wauben, M.H. Regular Industrial Processing of Bovine Milk Impacts the Integrity and Molecular Composition of Extracellular Vesicles. J. Nutr. 2021, 151, 1416–1425. [Google Scholar] [CrossRef]
- Gao, H.N.; Guo, H.Y.; Zhang, H.; Xie, X.L.; Wen, P.C.; Ren, F.Z. Yak-milk-derived exosomes promote proliferation of intestinal epithelial cells in an hypoxic environment. J. Dairy Sci. 2019, 102, 985–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, S.; Deighan, C.; Bandeira, E.; Kwak, K.J.; Rahman, M.; Nana-Sinkam, P.; Lee, L.J.; Paulaitis, M.E. Analyzing the miRNA content of extracellular vesicles by fluorescence nanoparticle tracking. Nanomedicine 2017, 13, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Vaswani, K.; Mitchell, M.D.; Holland, O.J.; Qin Koh, Y.; Hill, R.J.; Harb, T.; Davies, P.S.W.; Peiris, H. A Method for the Isolation of Exosomes from Human and Bovine Milk. J. Nutr. Metab. 2019, 2019, 5764740. [Google Scholar] [CrossRef] [PubMed]
- Reif, S.; Elbaum Shiff, Y.; Golan-Gerstl, R. Milk-derived exosomes (MDEs) have a different biological effect on normal fetal colon epithelial cells compared to colon tumor cells in a miRNA-dependent manner. J. Transl. Med. 2019, 17, 325. [Google Scholar] [CrossRef]
- Baddela, V.S.; Nayan, V.; Rani, P.; Onteru, S.K.; Singh, D. Physicochemical Biomolecular Insights into Buffalo Milk-Derived Nanovesicles. Appl. Biochem. Biotechnol. 2016, 178, 544–557. [Google Scholar] [CrossRef]
- Zhong, J.; Xia, B.; Shan, S.; Zheng, A.; Zhang, S.; Chen, J.; Liang, X.-J. High-quality milk exosomes as oral drug delivery system. Biomaterials 2021, 277, 121126. [Google Scholar] [CrossRef]
- Lässer, C.; Eldh, M.; Lötvall, J. Isolation and Characterization of RNA-Containing Exosomes. JoVE J. Vis. Exp. 2012, 59, e3037. [Google Scholar] [CrossRef]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filen, J.J.; Lahesmaa, R.; Norman, M.; Neve, E.P.; Scheynius, A.; Gabrielsson, S. Exosomes with immune modulatory features are present in human breast milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef]
- Miyake, H.; Lee, C.; Chusilp, S.; Bhalla, M.; Li, B.; Pitino, M.; Seo, S.; O’Connor, D.L.; Pierro, A. Human breast milk exosomes attenuate intestinal damage. Pediatr. Surg. Int. 2020, 36, 155–163. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, N.; Yan, T.; Shi, Y.N.; Chen, J.; Zhang, C.J.; Xie, X.J.; Liao, D.F.; Qin, L. The crosstalk: Exosomes and lipid metabolism. Cell Commun. Signal. 2020, 18, 119. [Google Scholar] [CrossRef]
- Samuel, M.; Chisanga, D.; Liem, M.; Keerthikumar, S.; Anand, S.; Ang, C.S.; Adda, C.G.; Versteegen, E.; Jois, M.; Mathivanan, S. Bovine milk-derived exosomes from colostrum are enriched with proteins implicated in immune response and growth. Sci. Rep. 2017, 7, 5933. [Google Scholar] [CrossRef] [PubMed]
- Vaswani, K.M.; Peiris, H.; Qin Koh, Y.; Hill, R.J.; Harb, T.; Arachchige, B.J.; Logan, J.; Reed, S.; Davies, P.S.W.; Mitchell, M.D. A complete proteomic profile of human and bovine milk exosomes by liquid chromatography mass spectrometry. Expert Rev. Proteom. 2021, 18, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.F.; Zuo, X.L.; Wang, X.; Ensslin, M.A.; Koti, V.; Hsueh, W.; Raymond, A.S.; Shur, B.D.; Tan, X.D. Milk fat globule-EGF factor 8/lactadherin plays a crucial role in maintenance and repair of murine intestinal epithelium. J. Clin. Investig. 2007, 117, 3673–3683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Lei, Y.; He, X.; Liu, D.; He, Z. Role of lactadherin in intestinal barrier integrity in experimental neonatal necrotizing enterocolitis. J. Cell Biochem. 2019, 120, 19509–19517. [Google Scholar] [CrossRef]
- Pieters, B.C.; Arntz, O.J.; Bennink, M.B.; Broeren, M.G.; van Caam, A.P.; Koenders, M.I.; van Lent, P.L.; van den Berg, W.B.; de Vries, M.; van der Kraan, P.M.; et al. Commercial cow milk contains physically stable extracellular vesicles expressing immunoregulatory TGF-beta. PLoS ONE 2015, 10, e0121123. [Google Scholar] [CrossRef]
- Reif, S.; Elbaum-Shiff, Y.; Koroukhov, N.; Shilo, I.; Musseri, M.; Golan-Gerstl, R. Cow and Human Milk-Derived Exosomes Ameliorate Colitis in DSS Murine Model. Nutrients 2020, 12, 2589. [Google Scholar] [CrossRef]
- Chen, W.; Chen, X.; Qian, Y.; Wang, X.; Zhou, Y.; Yan, X.; Yu, B.; Yao, S.; Yu, Z.; Zhu, J.; et al. Lipidomic Profiling of Human Milk Derived Exosomes and Their Emerging Roles in the Prevention of Necrotizing Enterocolitis. Mol. Nutr. Food. Res. 2021, 65, e2000845. [Google Scholar] [CrossRef]
- Yassin, A.M.; Abdel Hamid, M.I.; Farid, O.A.; Amer, H.; Warda, M. Dromedary milk exosomes as mammary transcriptome nano-vehicle: Their isolation, vesicular and phospholipidomic characterizations. J. Adv. Res. 2016, 7, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Zhang, Q.; Dong, Z.; Yan, Y.; Fu, Y.; Liu, X.; Zhao, B.; Duan, X. Phosphatidylcholine Ameliorates LPS-Induced Systemic Inflammation and Cognitive Impairments via Mediating the Gut–Brain Axis Balance. J. Agric. Food. Chem. 2020, 68, 14884–14895. [Google Scholar] [CrossRef]
- Anto, L.; Warykas, S.W.; Torres-Gonzalez, M.; Blesso, C.N. Milk Polar Lipids: Underappreciated Lipids with Emerging Health Benefits. Nutrients 2020, 12, 1001. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Cheong, L.Z.; Zhang, X.; Ali, A.H.; Jin, Q.; Wei, W.; Wang, X. Dietary Sphingomyelin Metabolism and Roles in Gut Health and Cognitive Development. Adv. Nutr. 2021, nmab117. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Anaya, J.; Jimenez-Flores, R. Symposium review: The relevance of bovine milk phospholipids in human nutrition-Evidence of the effect on infant gut and brain development. J. Dairy Sci. 2019, 102, 2738–2748. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and Exosomal MicroRNA: Trafficking, Sorting, and Function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [Green Version]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Yu, X.; Odenthal, M.; Fries, J.W. Exosomes as miRNA Carriers: Formation-Function-Future. Int. J. Mol. Sci. 2016, 17, 2028. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Perez-Hernandez, D.; Vazquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sanchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [Green Version]
- Janas, T.; Janas, M.M.; Sapon, K.; Janas, T. Mechanisms of RNA loading into exosomes. FEBS Lett. 2015, 589, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Koga, Y.; Yasunaga, M.; Moriya, Y.; Akasu, T.; Fujita, S.; Yamamoto, S.; Matsumura, Y. Exosome can prevent RNase from degrading microRNA in feces. J. Gastrointest. Oncol. 2011, 2, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Sharples, R.A.; Scicluna, B.J.; Hill, A.F. Exosomes provide a protective and enriched source of miRNA for biomarker profiling compared to intracellular and cell-free blood. J. Extracell. Vesicles 2014, 3, 23743. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hock, A.; Wu, R.Y.; Minich, A.; Botts, S.R.; Lee, C.; Antounians, L.; Miyake, H.; Koike, Y.; Chen, Y.; et al. Bovine milk-derived exosomes enhance goblet cell activity and prevent the development of experimental necrotizing enterocolitis. PLoS ONE 2019, 14, e0211431. [Google Scholar] [CrossRef] [PubMed]
- Manca, S.; Upadhyaya, B.; Mutai, E.; Desaulniers, A.T.; Cederberg, R.A.; White, B.R.; Zempleni, J. Milk exosomes are bioavailable and distinct microRNA cargos have unique tissue distribution patterns. Sci. Rep. 2018, 8, 11321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutai, E.; Ramer-Tait, A.E.; Zempleni, J. MicroRNAs in bovine milk exosomes are bioavailable in humans but do not elicit a robust pro-inflammatory cytokine response. ExRNA 2020, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Sukreet, S.; Silva, B.V.R.E.; Adamec, J.; Cui, J.; Zempleni, J. Galactose and Sialo-galactose Modifications in Glycoproteins on the Surface of Bovine Milk Exosome Are Essential for Exosome Uptake in Non-bovine Species (OR34-07-19). Curr. Dev. Nutr. 2019, 3 (Suppl. 1), nzz031-OR34. [Google Scholar] [CrossRef]
- Van Herwijnen, M.J.C.; Driedonks, T.A.P.; Snoek, B.L.; Kroon, A.M.T.; Kleinjan, M.; Jorritsma, R.; Pieterse, C.M.J.; Hoen, E.; Wauben, M.H.M. Abundantly Present miRNAs in Milk-Derived Extracellular Vesicles Are Conserved Between Mammals. Front. Nutr. 2018, 5, 81. [Google Scholar] [CrossRef]
- Simpson, M.R.; Brede, G.; Johansen, J.; Johnsen, R.; Storro, O.; Saetrom, P.; Oien, T. Human Breast Milk miRNA, Maternal Probiotic Supplementation and Atopic Dermatitis in Offspring. PLoS ONE 2015, 10, e0143496. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-related microRNAs are abundant in breast milk exosomes. Int. J. Biol. Sci. 2012, 8, 118–123. [Google Scholar] [CrossRef]
- Kohlhaas, S.; Garden, O.A.; Scudamore, C.; Turner, M.; Okkenhaug, K.; Vigorito, E. Cutting edge: The Foxp3 target miR-155 contributes to the development of regulatory T cells. J. Immunol. 2009, 182, 2578–2582. [Google Scholar] [CrossRef]
- Stephen, B.J.; Pareek, N.; Saeed, M.; Kausar, M.A.; Rahman, S.; Datta, M. Xeno-miRNA in Maternal-Infant Immune Crosstalk: An Aid to Disease Alleviation. Front Immunol. 2020, 11, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrillo-Lozano, E.; Sebastian-Valles, F.; Knott-Torcal, C. Circulating microRNAs in Breast Milk and Their Potential Impact on the Infant. Nutrients 2020, 12, 3066. [Google Scholar] [CrossRef] [PubMed]
- Carr, L.E.; Virmani, M.D.; Rosa, F.; Munblit, D.; Matazel, K.S.; Elolimy, A.A.; Yeruva, L. Role of Human Milk Bioactives on Infants’ Gut and Immune Health. Front. Immunol. 2021, 12, 604080. [Google Scholar] [CrossRef] [PubMed]
- Lonnerdal, B. Human Milk MicroRNAs/Exosomes: Composition and Biological Effects. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Maburutse, B.E.; Kang, M.; Park, M.R.; Park, D.J.; Kim, Y.; Oh, S. Short communication: Dietary bovine milk-derived exosomes improve bone health in an osteoporosis-induced mouse model. J. Dairy Sci. 2020, 103, 7752–7760. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Du, X.; Li, J.; Lonnerdal, B. Human milk exosomes and their microRNAs survive digestion in vitro and are taken up by human intestinal cells. Mol. Nutr. Food. Res. 2017, 61, 1700082. [Google Scholar] [CrossRef]
- Su, Y.; Yuan, J.; Zhang, F.; Lei, Q.; Zhang, T.; Li, K.; Guo, J.; Hong, Y.; Bu, G.; Lv, X.; et al. MicroRNA-181a-5p and microRNA-181a-3p cooperatively restrict vascular inflammation and atherosclerosis. Cell Death Dis. 2019, 10, 365. [Google Scholar] [CrossRef] [Green Version]
- Du, X.-J.; Lu, J.-M.; Sha, Y. MiR-181a inhibits vascular inflammation induced by ox-LDL via targeting TLR4 in human macrophages. J. Cell Physiol. 2018, 233, 6996–7003. [Google Scholar] [CrossRef]
- Le Doare, K.; Holder, B.; Bassett, A.; Pannaraj, P.S. Mother’s Milk: A Purposeful Contribution to the Development of the Infant Microbiota and Immunity. Front. Immunol. 2018, 9, 361. [Google Scholar] [CrossRef] [Green Version]
- Benmoussa, A.; Provost, P. Milk MicroRNAs in Health and Disease. Compr. Rev. Food. Sci. Food. Saf. 2019, 18, 703–722. [Google Scholar] [CrossRef] [Green Version]
- Lonnerdal, B.; Du, X.; Liao, Y.; Li, J. Human milk exosomes resist digestion in vitro and are internalized by human intestinal cells. FASEB J. 2015, 29, 121–123. [Google Scholar] [CrossRef]
- Kahn, S.; Liao, Y.; Du, X.; Xu, W.; Li, J.; Lonnerdal, B. Exosomal MicroRNAs in Milk from Mothers Delivering Preterm Infants Survive in Vitro Digestion and Are Taken Up by Human Intestinal Cells. Mol. Nutr. Food. Res. 2018, 62, e1701050. [Google Scholar] [CrossRef] [PubMed]
- Rani, P.; Vashisht, M.; Golla, N.; Shandilya, S.; Onteru, S.K.; Singh, D. Milk miRNAs encapsulated in exosomes are stable to human digestion and permeable to intestinal barrier in vitro. J. Funct. Foods 2017, 34, 431–439. [Google Scholar] [CrossRef]
- Howard, K.M.; Jati Kusuma, R.; Baier, S.R.; Friemel, T.; Markham, L.; Vanamala, J.; Zempleni, J. Loss of miRNAs during processing and storage of cow’s (Bos taurus) milk. J. Agric. Food Chem. 2015, 63, 588–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeyaram, A.; Jay, S.M. Preservation and Storage Stability of Extracellular Vesicles for Therapeutic Applications. AAPS J. 2017, 20, 1. [Google Scholar] [CrossRef]
- Leiferman, A.; Shu, J.; Upadhyaya, B.; Cui, J.; Zempleni, J. Storage of Extracellular Vesicles in Human Milk, and MicroRNA Profiles in Human Milk Exosomes and Infant Formulas. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 235–238. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, Z.; Sun, L.; Li, P. Fermentation Results in Quantitative Changes in Milk-Derived Exosomes and Different Effects on Cell Growth and Survival. J. Agric. Food. Chem. 2017, 65, 1220–1228. [Google Scholar] [CrossRef]
- Mukhopadhya, A.; Santoro, J.; Moran, B.; Useckaite, Z.; O’Driscoll, L. Optimisation and comparison of orthogonal methods for separation and characterisation of extracellular vesicles to investigate how representative infant milk formula is of milk. Food. Chem. 2021, 353, 129309. [Google Scholar] [CrossRef]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Bovine milk contains microRNA and messenger RNA that are stable under degradative conditions. J. Dairy Sci. 2012, 95, 4831–4841. [Google Scholar] [CrossRef] [Green Version]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. J. Nutr. 2014, 144, 1495–1500. [Google Scholar] [CrossRef]
- Betker, J.L.; Angle, B.M.; Graner, M.W.; Anchordoquy, T.J. The Potential of Exosomes From Cow Milk for Oral Delivery. J. Pharm. Sci. 2019, 108, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Hao, H.; Zhang, X.; Zhang, Z.; Lv, Y.; Zhang, L.; Yi, H. Oral Administration of Bovine Milk-Derived Extracellular Vesicles Alters the Gut Microbiota and Enhances Intestinal Immunity in Mice. Mol. Nutr. Food Res. 2020, 64, 1901251. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Paz, H.A.; Sadri, M.; Cui, J.; Kachman, S.D.; Fernando, S.C.; Zempleni, J. Dietary bovine milk exosomes elicit changes in bacterial communities in C57BL/6 mice. Am. J. Physiol. Gastrointest. Liver. Physiol. 2019, 317, G618–G624. [Google Scholar] [CrossRef]
- Wolf, T.; Baier, S.R.; Zempleni, J. The Intestinal Transport of Bovine Milk Exosomes Is Mediated by Endocytosis in Human Colon Carcinoma Caco-2 Cells and Rat Small Intestinal IEC-6 Cells. J. Nutr. 2015, 145, 2201–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnik, B.C.; Stremmel, W.; Weiskirchen, R.; John, S.M.; Schmitz, G. Exosome-Derived MicroRNAs of Human Milk and Their Effects on Infant Health and Development. Biomolecules 2021, 11, 851. [Google Scholar] [CrossRef]
- Khanam, A.; Yu, J.; Zempleni, J. Class A scavenger receptor-1/2 facilitates the uptake of bovine milk exosomes in murine bone marrow-derived macrophages and C57BL/6J mice. Am. J. Physiol. Cell Physiol. 2021, 321, C607–C614. [Google Scholar] [CrossRef]
- Izumi, H.; Tsuda, M.; Sato, Y.; Kosaka, N.; Ochiya, T.; Iwamoto, H.; Namba, K.; Takeda, Y. Bovine milk exosomes contain microRNA and mRNA and are taken up by human macrophages. J. Dairy Sci. 2015, 98, 2920–2933. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Patel, M.; Williams, S.; Arora, H.; Brawner, K.; Sims, B. Human breast milk-derived exosomes attenuate cell death in intestinal epithelial cells. Innate Immun. 2018, 24, 278–284. [Google Scholar] [CrossRef]
- Dong, P.; Zhang, Y.; Yan, D.Y.; Wang, Y.; Xu, X.; Zhao, Y.C.; Xiao, T.T. Protective Effects of Human Milk-Derived Exosomes on Intestinal Stem Cells Damaged by Oxidative Stress. Cell Transpl. 2020, 29, 963689720912690. [Google Scholar] [CrossRef] [Green Version]
- Aydemir, C.; Dilli, D.; Uras, N.; Ulu, H.O.; Oguz, S.S.; Erdeve, O.; Dilmen, U. Total oxidant status and oxidative stress are increased in infants with necrotizing enterocolitis. J. Pediatr. Surg. 2011, 46, 2096–2100. [Google Scholar] [CrossRef]
- Podolsky, D.K. Inflammatory Bowel Disease. N. Engl. J. Med. 1991, 325, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.; Camacho Salazar, R.; Chen, Y.H. Animal models for studying epithelial barriers in neonatal necrotizing enterocolitis, inflammatory bowel disease and colorectal cancer. Tissue Barriers 2017, 5, e1356901. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Liu, G.; Zhu, X. Human breast milk-derived exosomes may help maintain intestinal epithelial barrier integrity. Pediatr. Res. 2021, 90, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Pisano, C.; Galley, J.; Elbahrawy, M.; Wang, Y.; Farrell, A.; Brigstock, D.; Besner, G.E. Human Breast Milk-Derived Extracellular Vesicles in the Protection Against Experimental Necrotizing Enterocolitis. J. Pediatr. Surg. 2020, 55, 54–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, L.S.; Parks, O.B.; Good, M. A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis. Nutrients 2019, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Kantorowska, A.; Wei, J.C.; Cohen, R.S.; Lawrence, R.A.; Gould, J.B.; Lee, H.C. Impact of Donor Milk Availability on Breast Milk Use and Necrotizing Enterocolitis Rates. Pediatrics 2016, 137, e20153123. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, X.; Shi, Z.; Shen, L.; Zhang, J.; Zhang, J. Bovine milk exosomes attenuate the alteration of purine metabolism and energy status in IEC-6 cells induced by hydrogen peroxide. Food. Chem. 2021, 350, 129142. [Google Scholar] [CrossRef]
- Wang, L.; Shi, Z.; Wang, X.; Mu, S.; Xu, X.; Shen, L.; Li, P. Protective effects of bovine milk exosomes against oxidative stress in IEC-6 cells. Eur. J. Nutr. 2021, 60, 317–327. [Google Scholar] [CrossRef]
- Gao, H.N.; Hu, H.; Wen, P.C.; Lian, S.; Xie, X.L.; Song, H.L.; Yang, Z.N.; Ren, F.Z. Yak milk–derived exosomes alleviate lipopolysaccharide-induced intestinal inflammation by inhibiting PI3K/AKT/C3 pathway activation. J. Dairy Sci. 2021, 104, 8411–8424. [Google Scholar] [CrossRef]
- Xie, M.Y.; Chen, T.; Xi, Q.Y.; Hou, L.J.; Luo, J.Y.; Zeng, B.; Li, M.; Sun, J.J.; Zhang, Y.L. Porcine milk exosome miRNAs protect intestinal epithelial cells against deoxynivalenol-induced damage. Biochem. Pharm. 2020, 175, 113898. [Google Scholar] [CrossRef]
- Xie, M.Y.; Hou, L.J.; Sun, J.J.; Zeng, B.; Xi, Q.Y.; Luo, J.Y.; Chen, T.; Zhang, Y.L. Porcine Milk Exosome MiRNAs Attenuate LPS-Induced Apoptosis through Inhibiting TLR4/NF-kappaB and p53 Pathways in Intestinal Epithelial Cells. J. Agric. Food. Chem. 2019, 67, 9477–9491. [Google Scholar] [CrossRef] [PubMed]
- Good, M.; Sodhi, C.P.; Hackam, D.J. Evidence-based feeding strategies before and after the development of necrotizing enterocolitis. Expert Rev. Clin. Immunol. 2014, 10, 875–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Kittana, H.; Shu, J.A.-O.X.; Kachman, S.A.-O.X.; Cui, J.; Ramer-Tait, A.A.-O.; Zempleni, J.A.-O. Dietary Depletion of Milk Exosomes and Their MicroRNA Cargos Elicits a Depletion of miR-200a-3p and Elevated Intestinal Inflammation and Chemokine (C-X-C Motif) Ligand 9 Expression in Mdr1a(-/-) Mice. Curr. Dev. Nutr. 2019, 3, nzz122. [Google Scholar] [CrossRef]
- Stremmel, W.; Weiskirchen, R.; Melnik, B.C. Milk Exosomes Prevent Intestinal Inflammation in a Genetic Mouse Model of Ulcerative Colitis: A Pilot Experiment. Inflamm. Intestig. Dis. 2020, 5, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Benmoussa, A.; Diallo, I.; Salem, M.; Michel, S.; Gilbert, C.; Sevigny, J.; Provost, P. Concentrates of two subsets of extracellular vesicles from cow’s milk modulate symptoms and inflammation in experimental colitis. Sci. Rep. 2019, 9, 14661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, L.; Hao, H.; Zhang, Z.; Lv, Y.; Liang, X.; Liu, Q.; Liu, T.; Gong, P.; Zhang, L.; Cao, F.; et al. Milk-derived extracellular vesicles alleviate ulcerative colitis by regulating the gut immunity and reshaping the gut microbiota. Theranostics 2021, 11, 8570–8586. [Google Scholar] [CrossRef]
- Maghraby, M.K.; Li, B.; Chi, L.; Ling, C.; Benmoussa, A.; Provost, P.; Postmus, A.C.; Abdi, A.; Pierro, A.; Bourdon, C.; et al. Extracellular vesicles isolated from milk can improve gut barrier dysfunction induced by malnutrition. Sci. Rep. 2021, 11, 7635. [Google Scholar] [CrossRef]
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef]
- Superti, F. Lactoferrin from Bovine Milk: A Protective Companion for Life. Nutrients 2020, 12, 2562. [Google Scholar] [CrossRef]
- Takada, Y.; Kobayashi, N.; Matsuyama, H.; Kato, K.; Yamamura, J.; Yahiro, M.; Kumegawa, M.; Aoe, S. Whey protein suppresses the osteoclast-mediated bone resorption and osteoclast cell formation. Int. Dairy J. 1997, 7, 821–825. [Google Scholar] [CrossRef]
- Toba, Y.; Takada, Y.; Matsuoka, Y.; Morita, Y.; Motouri, M.; Hirai, T.; Suguri, T.; Aoe, S.; Kawakami, H.; Kumegawa, M.; et al. Milk Basic Protein Promotes Bone Formation and Suppresses Bone Resorption in Healthy Adult Men. Biosci. Biotechnol. Biochem. 2001, 65, 1353–1357. [Google Scholar] [CrossRef] [PubMed]
- Yackobovitch-Gavan, M.; Phillip, M.; Gat-Yablonski, G. How Milk and Its Proteins Affect Growth, Bone Health, and Weight. Horm. Res. Paediatr. 2017, 88, 63–69. [Google Scholar] [CrossRef]
- Weaver, C.M. Milk consumption and bone health. JAMA Pediatr. 2014, 168, 12–13. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D. Dairy Foods, Obesity, and Metabolic Health: The Role of the Food Matrix Compared with Single Nutrients. Adv. Nutr. 2019, 10, 917S–923S. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.C.; Arntz, O.J.; Blaney Davidson, E.N.; van Lent, P.L.; Koenders, M.I.; van der Kraan, P.M.; van den Berg, W.B.; Ferreira, A.V.; van de Loo, F.A. Milk extracellular vesicles accelerate osteoblastogenesis but impair bone matrix formation. J. Nutr. Biochem. 2016, 30, 74–84. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Di Ceglie, I.; Arntz, O.J.; van den Berg, W.B.; van den Hoogen, F.H.; Ferreira, A.V.; van Lent, P.L.; van de Loo, F.A. Milk-Derived Nanoparticle Fraction Promotes the Formation of Small Osteoclasts But Reduces Bone Resorption. J. Cell Physiol. 2017, 232, 225–233. [Google Scholar] [CrossRef]
- Go, G.; Jeon, J.; Lee, G.; Lee, J.H.; Lee, S.H. Bovine milk extracellular vesicles induce the proliferation and differentiation of osteoblasts and promote osteogenesis in rats. J. Food. Biochem. 2021, 45, e13705. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Pieters, B.C.H.; Guimaraes, P.B.; Duffles, L.F.; Heredia, J.E.; Silveira, A.L.M.; Oliveira, A.C.C.; Teixeira, M.M.; Ferreira, A.V.M.; Silva, T.A.; et al. Bovine Milk Extracellular Vesicles Are Osteoprotective by Increasing Osteocyte Numbers and Targeting RANKL/OPG System in Experimental Models of Bone Loss. Front. Bioeng. Biotechnol. 2020, 8, 891. [Google Scholar] [CrossRef]
- Grasgruber, P.; Sebera, M.; Hrazdira, E.; Cacek, J.; Kalina, T. Major correlates of male height: A study of 105 countries. Econ. Hum. Biol. 2016, 21, 172–195. [Google Scholar] [CrossRef] [Green Version]
- DeBoer, M.D.; Agard, H.E.; Scharf, R.J. Milk intake, height and body mass index in preschool children. Arch. Dis. Child. 2015, 100, 460–465. [Google Scholar] [CrossRef] [Green Version]
- Marshall, T.A.; Curtis, A.M.; Cavanaugh, J.E.; Warren, J.J.; Levy, S.M. Higher Longitudinal Milk Intakes Are Associated with Increased Height in a Birth Cohort Followed for 17 Years. J. Nutr. 2018, 148, 1144–1149. [Google Scholar] [CrossRef] [PubMed]
- Sinai, T.; Goldberg, M.R.; Nachshon, L.; Amitzur-Levy, R.; Yichie, T.; Katz, Y.; Monsonego-Ornan, E.; Elizur, A. Reduced Final Height and Inadequate Nutritional Intake in Cow’s Milk-Allergic Young Adults. J. Allergy Clin. Immunol. Pract. 2019, 7, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Grenov, B.; Larnkjær, A.; Mølgaard, C.; Michaelsen, K.F. Role of Milk and Dairy Products in Growth of the Child. Nestle Nutr. Inst. Workshop Ser. 2020, 93, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Mølgaard, C.; Larnkjær, A.; Arnberg, K.; Michaelsen, K.F. Milk and growth in children: Effects of whey and casein. Nestle Nutr. Workshop Ser. Paediatr. Programme 2011, 67, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Racine, H.L.; Serrat, M.A. The Actions of IGF-1 in the Growth Plate and Its Role in Postnatal Bone Elongation. Curr. Osteoporos. Rep. 2020, 18, 210–227. [Google Scholar] [CrossRef]
- Martin, R.M.; Holly, J.M.; Gunnell, D. Milk and linear growth: Programming of the igf-I axis and implication for health in adulthood. Nestle Nutr. Workshop Ser. Paediatr. Programme 2011, 67, 79–97. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Chen, T.; Xie, M.; Li, M.; Zeng, B.; Sun, R.; Zhu, Y.; Ye, D.; Wu, J.; Sun, J.; et al. Oral Administration of Bovine and Porcine Milk Exosome Alter miRNAs Profiles in Piglet Serum. Sci. Rep. 2020, 10, 6983. [Google Scholar] [CrossRef]
- Melnik, B.C.; Schmitz, G. MicroRNAs: Milk’s epigenetic regulators. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 427–442. [Google Scholar] [CrossRef]
- Fricke, O.; Schoenau, E. The ‘Functional Muscle-Bone Unit’: Probing the relevance of mechanical signals for bone development in children and adolescents. Growth Horm. IGF Res. 2007, 17, 1–9. [Google Scholar] [CrossRef]
- Wilkinson, S.B.; Tarnopolsky, M.A.; MacDonald, M.J.; MacDonald, J.R.; Armstrong, D.; Phillips, S.M. Consumption of fluid skim milk promotes greater muscle protein accretion after resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein beverage. Am. J. Clin. Nutr. 2007, 85, 1031–1040. [Google Scholar] [CrossRef]
- Mobley, C.B.; Mumford, P.W.; McCarthy, J.J.; Miller, M.E.; Young, K.C.; Martin, J.S.; Beck, D.T.; Lockwood, C.M.; Roberts, M.D. Whey protein-derived exosomes increase protein synthesis and hypertrophy in C2-C12 myotubes. J. Dairy Sci. 2017, 100, 48–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiferman, A.; Shu, J.; Grove, R.; Cui, J.; Adamec, J.; Zempleni, J. A diet defined by its content of bovine milk exosomes and their RNA cargos has moderate effects on gene expression, amino acid profiles and grip strength in skeletal muscle in C57BL/6 mice. J. Nutr. Biochem. 2018, 59, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Parry, H.A.; Mobley, C.B.; Mumford, P.W.; Romero, M.A.; Haun, C.T.; Zhang, Y.; Roberson, P.A.; Zempleni, J.; Ferrando, A.A.; Vechetti, I.J., Jr.; et al. Bovine Milk Extracellular Vesicles (EVs) Modification Elicits Skeletal Muscle Growth in Rats. Front. Physiol. 2019, 10, 436. [Google Scholar] [CrossRef]
- Cacho, N.T.; Lawrence, R.M. Innate Immunity and Breast Milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Dee, D.; Li, C.M.; Hoffman, H.J.; Grummer-Strawn, L.M. Breastfeeding and risk of infections at 6 years. Pediatrics 2014, 134 (Suppl. 1), S13–S20. [Google Scholar] [CrossRef] [Green Version]
- Ladomenou, F.; Moschandreas, J.; Kafatos, A.; Tselentis, Y.; Galanakis, E. Protective effect of exclusive breastfeeding against infections during infancy: A prospective study. Arch. Dis. Child. 2010, 95, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Henrick, B.M.; Yao, X.D.; Nasser, L.; Roozrogousheh, A.; Rosenthal, K.L. Breastfeeding Behaviors and the Innate Immune System of Human Milk: Working Together to Protect Infants against Inflammation, HIV-1, and Other Infections. Front. Immunol. 2017, 8, 1631. [Google Scholar] [CrossRef] [Green Version]
- Kramer, M.S. Breastfeeding and allergy: The evidence. Ann. Nutr. Metab. 2011, 59 (Suppl. 1), 20–26. [Google Scholar] [CrossRef]
- Kosaka, N.; Izumi, H.; Sekine, K.; Ochiya, T. microRNA as a new immune-regulatory agent in breast milk. Silence 2010, 1, 7. [Google Scholar] [CrossRef] [Green Version]
- Zeng, B.; Chen, T.; Luo, J.Y.; Zhang, L.; Xi, Q.Y.; Jiang, Q.Y.; Sun, J.J.; Zhang, Y.L. Biological Characteristics and Roles of Noncoding RNAs in Milk-Derived Extracellular Vesicles. Adv. Nutr. 2021, 12, 1006–1019. [Google Scholar] [CrossRef]
- Aarts, J.; Boleij, A.; Pieters, B.C.H.; Feitsma, A.L.; van Neerven, R.J.J.; Ten Klooster, J.P.; M’Rabet, L.; Arntz, O.J.; Koenders, M.I.; van de Loo, F.A.J. Flood Control: How Milk-Derived Extracellular Vesicles Can Help to Improve the Intestinal Barrier Function and Break the Gut-Joint Axis in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 703277. [Google Scholar] [CrossRef] [PubMed]
- Alsaweed, M.; Hartmann, P.E.; Geddes, D.T.; Kakulas, F. MicroRNAs in Breastmilk and the Lactating Breast: Potential Immunoprotectors and Developmental Regulators for the Infant and the Mother. Int. J. Env. Res. Public Health 2015, 12, 13981–14020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, B.; Kim, Y.; Park, D.J.; Oh, S. Comparative analysis of dietary exosome-derived microRNAs from human, bovine and caprine colostrum and mature milk. J. Anim. Sci. Technol. 2021, 63, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xie, Y.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Milk exosome-derived miRNAs from water buffalo are implicated in immune response and metabolism process. BMC Vet. Res. 2020, 16, 123. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhan, Z.; Xu, L.; Ma, F.; Li, D.; Guo, Z.; Li, N.; Cao, X. MicroRNA-148/152 impair innate response and antigen presentation of TLR-triggered dendritic cells by targeting CaMKIIalpha. J. Immunol. 2010, 185, 7244–7251. [Google Scholar] [CrossRef]
- Friedrich, M.; Pracht, K.; Mashreghi, M.F.; Jack, H.M.; Radbruch, A.; Seliger, B. The role of the miR-148/-152 family in physiology and disease. Eur. J. Immunol. 2017, 47, 2026–2038. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S. Recent findings regarding let-7 in immunity. Cancer Lett. 2018, 434, 130–131. [Google Scholar] [CrossRef]
- Benmoussa, A.; Gotti, C.; Bourassa, S.; Gilbert, C.; Provost, P. Identification of protein markers for extracellular vesicle (EV) subsets in cow’s milk. J. Proteom. 2019, 192, 78–88. [Google Scholar] [CrossRef]
- Mangan, P.R.; Harrington, L.E.; O’Quinn, D.B.; Helms, W.S.; Bullard, D.C.; Elson, C.O.; Hatton, R.D.; Wahl, S.M.; Schoeb, T.R.; Weaver, C.T. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature 2006, 441, 231–234. [Google Scholar] [CrossRef]
- Arntz, O.J.; Pieters, B.C.; Oliveira, M.C.; Broeren, M.G.; Bennink, M.B.; de Vries, M.; van Lent, P.L.; Koenders, M.I.; van den Berg, W.B.; van der Kraan, P.M.; et al. Oral administration of bovine milk derived extracellular vesicles attenuates arthritis in two mouse models. Mol. Nutr. Food. Res. 2015, 59, 1701–1712. [Google Scholar] [CrossRef]
- Rahman, M.M.; Takashima, S.; Kamatari, Y.O.; Shimizu, K.; Okada, A.; Inoshima, Y. Comprehensive Proteomic Analysis Revealed a Large Number of Newly Identified Proteins in the Small Extracellular Vesicles of Milk from Late-Stage Lactating Cows. Animals 2021, 11, 2506. [Google Scholar] [CrossRef] [PubMed]
- Ascanius, S.R.; Hansen, M.S.; Ostenfeld, M.S.; Rasmussen, J.T. Milk-Derived Extracellular Vesicles Suppress Inflammatory Cytokine Expression and Nuclear Factor-κB Activation in Lipopolysaccharide-Stimulated Macrophages. Dairy 2021, 2, 165–178. [Google Scholar] [CrossRef]
- Matic, S.; D’Souza, D.H.; Wu, T.; Pangloli, P.; Dia, V.P. Bovine Milk Exosomes Affect Proliferation and Protect Macrophages against Cisplatin-Induced Cytotoxicity. Immunol. Investig. 2020, 49, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Naslund, T.I.; Paquin-Proulx, D.; Paredes, P.T.; Vallhov, H.; Sandberg, J.K.; Gabrielsson, S. Exosomes from breast milk inhibit HIV-1 infection of dendritic cells and subsequent viral transfer to CD4+ T cells. AIDS 2014, 28, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smythies, L.E.; Smythies, J.R. Exosomes in the gut. Front. Immunol. 2014, 5, 104. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Zhuang, X.; Wang, Q.; Jiang, H.; Deng, Z.B.; Wang, B.; Zhang, L.; Kakar, S.; Jun, Y.; Miller, D.; et al. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles. Mol. Nutr. Food Res. 2014, 58, 1561–1573. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e8. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Zhao, Z.; Xu, X.; Li, M.; Li, P. Characterization of three different types of extracellular vesicles and their impact on bacterial growth. Food Chem. 2019, 272, 372–378. [Google Scholar] [CrossRef]
- Wang, S.; Auchtung, J.; Zempleni, J. Milk Exosomes Protect Human Microbiota Associated-Mice Against Clostridioides difficile Infection. Curr. Dev. Nutr. 2021, 5, 1189. [Google Scholar] [CrossRef]
- Horta, B.L.; de Sousa, B.A.; de Mola, C.L. Breastfeeding and neurodevelopmental outcomes. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 174–178. [Google Scholar] [CrossRef]
- Koo, W.; Tank, S.; Martin, S.; Shi, R. Human milk and neurodevelopment in children with very low birth weight: A systematic review. Nutr. J. 2014, 13, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottolini, K.M.; Andescavage, N.; Keller, S.; Limperopoulos, C. Nutrition and the developing brain: The road to optimizing early neurodevelopment: A systematic review. Pediatr. Res. 2020, 87, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, N.L.; Aasmul-Olsen, K.; Venkatasubramanian, R.; Nygaard, M.K.E.; Sprenger, R.R.; Heckmann, A.B.; Ostenfeld, M.S.; Ejsing, C.S.; Eskildsen, S.F.; Mullertz, A.; et al. Dairy-Derived Emulsifiers in Infant Formula Show Marginal Effects on the Plasma Lipid Profile and Brain Structure in Preterm Piglets Relative to Soy Lecithin. Nutrients 2021, 13, 718. [Google Scholar] [CrossRef] [PubMed]
- Mutai, E.; Zhou, F.; Zempleni, J. Depletion of Dietary Bovine Milk Exosomes Impairs Sensorimotor Gating and Spatial Learning in C57BL/6 Mice. FASEB J. 2017, 31, 150–154. [Google Scholar] [CrossRef]
- Aguilar-Lozano, A.; Baier, S.; Grove, R.; Shu, J.; Giraud, D.; Leiferman, A.; Mercer, K.E.; Cui, J.; Badger, T.M.; Adamec, J.; et al. Concentrations of Purine Metabolites Are Elevated in Fluids from Adults and Infants and in Livers from Mice Fed Diets Depleted of Bovine Milk Exosomes and their RNA Cargos. J. Nutr. 2018, 148, 1886–1894. [Google Scholar] [CrossRef] [Green Version]
- Tome-Carneiro, J.; Fernandez-Alonso, N.; Tomas-Zapico, C.; Visioli, F.; Iglesias-Gutierrez, E.; Davalos, A. Breast milk microRNAs harsh journey towards potential effects in infant development and maturation. Lipid encapsulation can help. Pharm. Res. 2018, 132, 21–32. [Google Scholar] [CrossRef]
- Chiurazzi, M.; Cozzolino, M.; Reinelt, T.; Nguyen, T.D.; Elke Chie, S.; Natalucci, G.; Miletta, M.C. Human Milk and Brain Development in Infants. Reprod. Med. 2021, 2, 107–117. [Google Scholar] [CrossRef]
- Carney, M.C.; Tarasiuk, A.; DiAngelo, S.L.; Silveyra, P.; Podany, A.; Birch, L.L.; Paul, I.M.; Kelleher, S.; Hicks, S.D. Metabolism-related microRNAs in maternal breast milk are influenced by premature delivery. Pediatr. Res. 2017, 82, 226–236. [Google Scholar] [CrossRef]
- Auerbach, A.; Vyas, G.; Li, A.; Halushka, M.; Witwer, K. Uptake of dietary milk miRNAs by adult humans: A validation study. F1000Research 2016, 5, 721. [Google Scholar] [CrossRef]
- Laubier, J.; Castille, J.; Le Guillou, S.; Le Provost, F. No effect of an elevated miR-30b level in mouse milk on its level in pup tissues. RNA Biol. 2015, 12, 26–29. [Google Scholar] [CrossRef]
- Wang, L.; Sadri, M.; Giraud, D.; Zempleni, J. RNase H2-Dependent Polymerase Chain Reaction and Elimination of Confounders in Sample Collection, Storage, and Analysis Strengthen Evidence That microRNAs in Bovine Milk Are Bioavailable in Humans. J. Nutr. 2018, 148, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromm, B.; Tosar, J.P.; Lu, Y.; Halushka, M.K.; Witwer, K.W. Human and Cow Have Identical miR-21-5p and miR-30a-5p Sequences, Which Are Likely Unsuited to Study Dietary Uptake from Cow Milk. J. Nutr. 2018, 148, 1506–1507. [Google Scholar] [CrossRef] [PubMed]
- Mar-Aguilar, F.; Arreola-Triana, A.; Mata-Cardona, D.; Gonzalez-Villasana, V.; Rodriguez-Padilla, C.; Resendez-Perez, D. Evidence of transfer of miRNAs from the diet to the blood still inconclusive. PeerJ 2020, 8, e9567. [Google Scholar] [CrossRef]
- O’Reilly, D.; Dorodnykh, D.; Avdeenko, N.V.; Nekliudov, N.A.; Garssen, J.; Elolimy, A.A.; Petrou, L.; Simpson, M.R.; Yeruva, L.; Munblit, D. Perspective: The Role of Human Breast-Milk Extracellular Vesicles in Child Health and Disease. Adv. Nutr. 2021, 12, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Gadeval, A.; Kathar, U.; Sengupta, P.; Kalia, K.; Tekade, R.K. Emerging roles and biopharmaceutical applications of milk derived exosomes. J. Drug. Deliv. Sci. Technol. 2021, 64, 102577. [Google Scholar] [CrossRef]
- Ong, S.L.; Blenkiron, C.; Haines, S.; Acevedo-Fani, A.; Leite, J.A.S.; Zempleni, J.; Anderson, R.C.; McCann, M.J. Ruminant Milk-Derived Extracellular Vesicles: A Nutritional and Therapeutic Opportunity? Nutrients 2021, 13, 2505. [Google Scholar] [CrossRef]
- Melnik, B.C. Synergistic Effects of Milk-Derived Exosomes and Galactose on alpha-Synuclein Pathology in Parkinson’s Disease and Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 1059. [Google Scholar] [CrossRef]
- Melnik, B.C.; Schmitz, G. Exosomes of pasteurized milk: Potential pathogens of Western diseases. J. Transl. Med. 2019, 17, 3. [Google Scholar] [CrossRef] [Green Version]
- Melnik, B.C.; Schmitz, G. Pasteurized non-fermented cow’s milk but not fermented milk is a promoter of mTORC1-driven aging and increased mortality. Ageing Res. Rev. 2021, 67, 101270. [Google Scholar] [CrossRef]
- Melnik, B.C. Lifetime Impact of Cow’s Milk on Overactivation of mTORC1: From Fetal to Childhood Overgrowth, Acne, Diabetes, Cancers, and Neurodegeneration. Biomolecules 2021, 11, 404. [Google Scholar] [CrossRef]
- Wehbe, Z.; Kreydiyyeh, S. Cow’s milk may be delivering potentially harmful undetected cargoes to humans. Is it time to reconsider dairy recommendations? Nutr. Rev. 2021, 80, 874–888. [Google Scholar] [CrossRef] [PubMed]
- FAO. Milk and Dairy Products in Human Nutrition; FAO: Rome, Italy, 2013. [Google Scholar]
- Meng, W.; He, C.; Hao, Y.; Wang, L.; Li, L.; Zhu, G. Prospects and challenges of extracellular vesicle-based drug delivery system: Considering cell source. Drug Deliv. 2020, 27, 585–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Sun, X.; Huang, M.; Ma, Q.; Du, L.; Cui, Y. Enhanced Neuroprotective Effects of Epicatechin Gallate Encapsulated by Bovine Milk-Derived Exosomes against Parkinson’s Disease through Antiapoptosis and Antimitophagy. J. Agric. Food Chem. 2021, 69, 5134–5143. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; Aqil, F.; Tyagi, N.; Gupta, R. Milk exosomes: A biogenic nanocarrier for small molecules and macromolecules to combat cancer. Am. J. Reprod. Immunol. 2021, 85, e13349. [Google Scholar] [CrossRef]
- Vashisht, M.; Rani, P.; Onteru, S.K.; Singh, D. Curcumin Encapsulated in Milk Exosomes Resists Human Digestion and Possesses Enhanced Intestinal Permeability in vitro. Appl. Biochem. Biotechnol. 2017, 183, 993–1007. [Google Scholar] [CrossRef]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, B.; Shen, J.; Wan, L.; Zhu, Y.; Yi, T.; Xiao, Z. The Beneficial Effects of Quercetin, Curcumin, and Resveratrol in Obesity. Oxid. Med. Cell Longev. 2017, 2017, 1459497. [Google Scholar] [CrossRef]
- Alappat, L.; Awad, A.B. Curcumin and obesity: Evidence and mechanisms. Nutr. Rev. 2010, 68, 729–738. [Google Scholar] [CrossRef]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M. Curcumin and Health. Molecules 2016, 21, 264. [Google Scholar] [CrossRef]
- Roth, S.; Spalinger, M.R.; Gottier, C.; Biedermann, L.; Zeitz, J.; Lang, S.; Weber, A.; Rogler, G.; Scharl, M. Bilberry-Derived Anthocyanins Modulate Cytokine Expression in the Intestine of Patients with Ulcerative Colitis. PLoS ONE 2016, 11, e0154817. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of Curcumin: Problems and Promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Ichiyanagi, T.; Shida, Y.; Rahman, M.M.; Hatano, Y.; Konishi, T. Bioavailability and Tissue Distribution of Anthocyanins in Bilberry (Vaccinium myrtillus L.) Extract in Rats. J. Agric. Food. Chem. 2006, 54, 6578–6587. [Google Scholar] [CrossRef] [PubMed]
- Busatto, S.; Vilanilam, G.; Ticer, T.; Lin, W.L.; Dickson, D.W.; Shapiro, S.; Bergese, P.; Wolfram, J. Tangential Flow Filtration for Highly Efficient Concentration of Extracellular Vesicles from Large Volumes of Fluid. Cells 2018, 7, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, R.; Ma, J.; Yan, Q.; Yang, S.; Fan, Z.; Jiang, Z. Biochemical characterization of a novel α-L-fucosidase from Pedobacter sp. and its application in synthesis of 3′-fucosyllactose and 2′-fucosyllactose. Appl. Microbiol. Biotechnol. 2020, 104, 5813–5826. [Google Scholar] [CrossRef]
- He, Y.; He, Z.; Leone, S.; Liu, S. Milk Exosomes Transfer Oligosaccharides into Macrophages to Modulate Immunity and Attenuate Adherent-Invasive E. coli (AIEC) Infection. Nutrients 2021, 13, 3198. [Google Scholar] [CrossRef]
- Zeng, B.; Chen, T.; Luo, J.; Xie, M.; Wei, L.; Xi, Q.; Sun, J.; Zhang, Y. Exploration of Long Non-coding RNAs and Circular RNAs in Porcine Milk Exosomes. Front. Genet. 2020, 11, 652. [Google Scholar] [CrossRef]
- Karlsson, O.; Rodosthenous, R.S.; Jara, C.; Brennan, K.J.; Wright, R.O.; Baccarelli, A.A.; Wright, R.J. Detection of long non-coding RNAs in human breastmilk extracellular vesicles: Implications for early child development. Epigenetics 2016, 11, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, D.; Wang, Y.; Li, M.; Fang, X.; Chen, H.; Zhang, C. The landscape of circular RNAs and mRNAs in bovine milk exosomes. J. Food. Compos. Anal. 2019, 76, 33–38. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Guan, L.L. Noncoding RNAs: Regulatory Molecules of Host-Microbiome Crosstalk. Trends Microbiol. 2021, 29, 713–724. [Google Scholar] [CrossRef]
- Alsaweed, M.; Lai, C.T.; Hartmann, P.E.; Geddes, D.T.; Kakulas, F. Human milk miRNAs primarily originate from the mammary gland resulting in unique miRNA profiles of fractionated milk. Sci. Rep. 2016, 6, 20680. [Google Scholar] [CrossRef]
- Madden, J.W. Human breast milk exosomes may protect against necrotizing enterocolitis in preterm infants. Pediatric Res. 2021, 90, 244–245. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Biogenesis of milk exosomes and milk fat globules. (A), milk exosomes; (B), milk fat globules.
Figure 1.
Biogenesis of milk exosomes and milk fat globules. (A), milk exosomes; (B), milk fat globules.
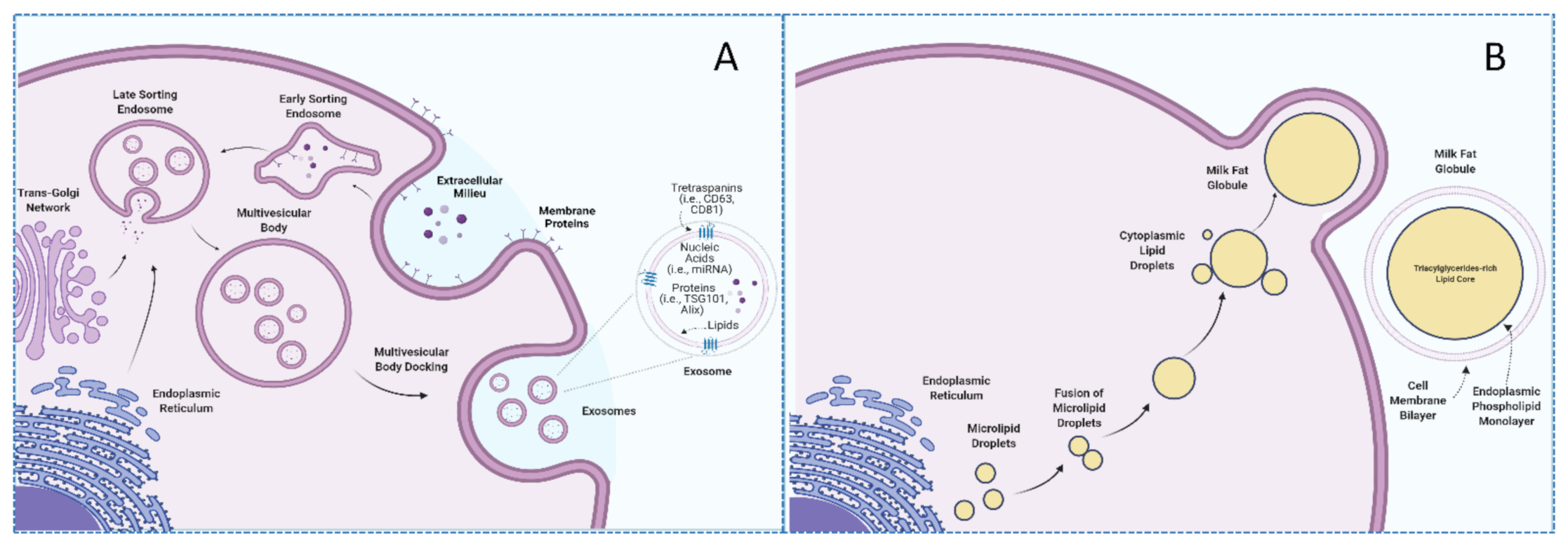
Figure 2.
MiRNA synthesis and loading into milk exosomes.
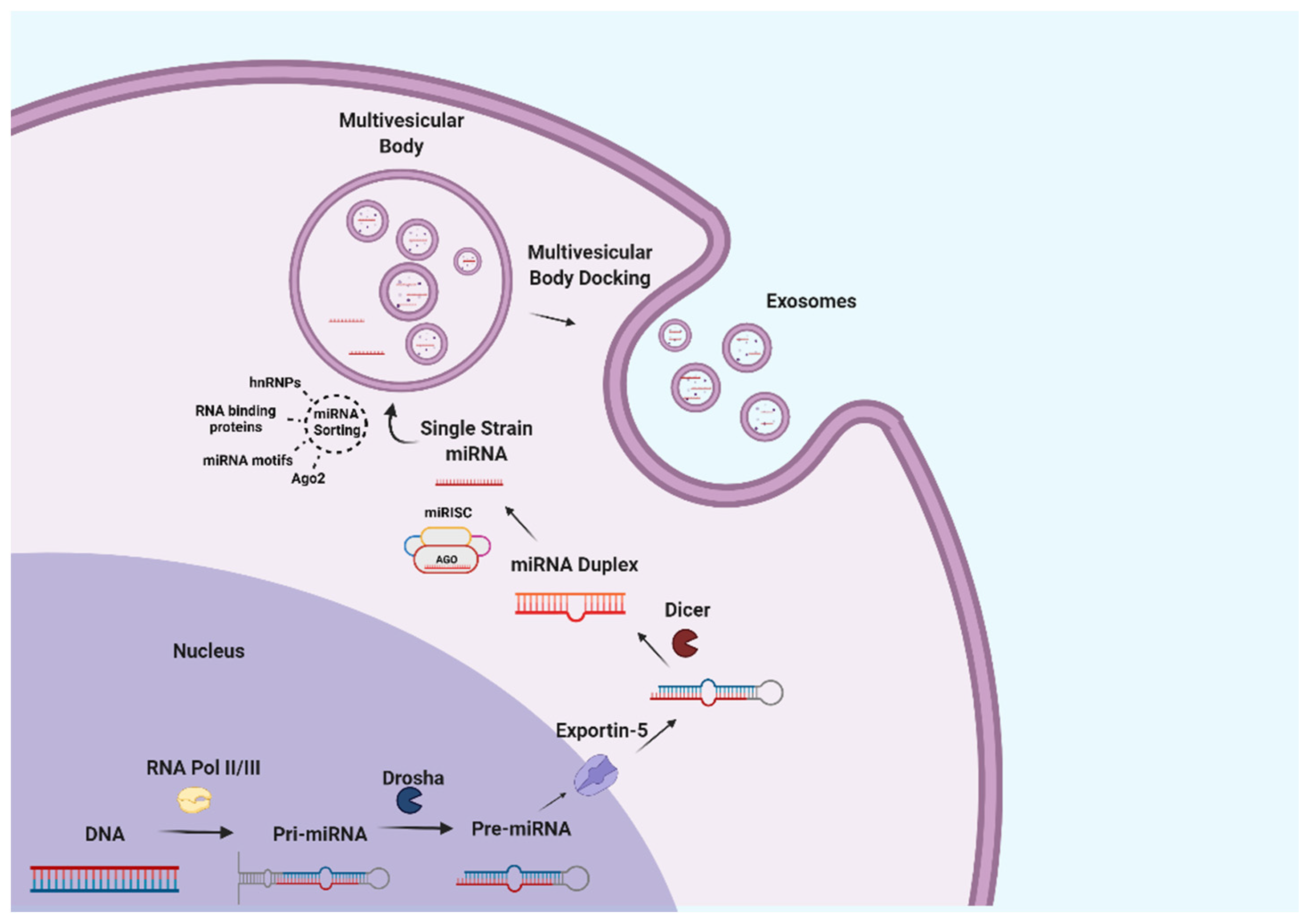
Figure 3.
Potential beneficial effects of milk exosomes on overall health.
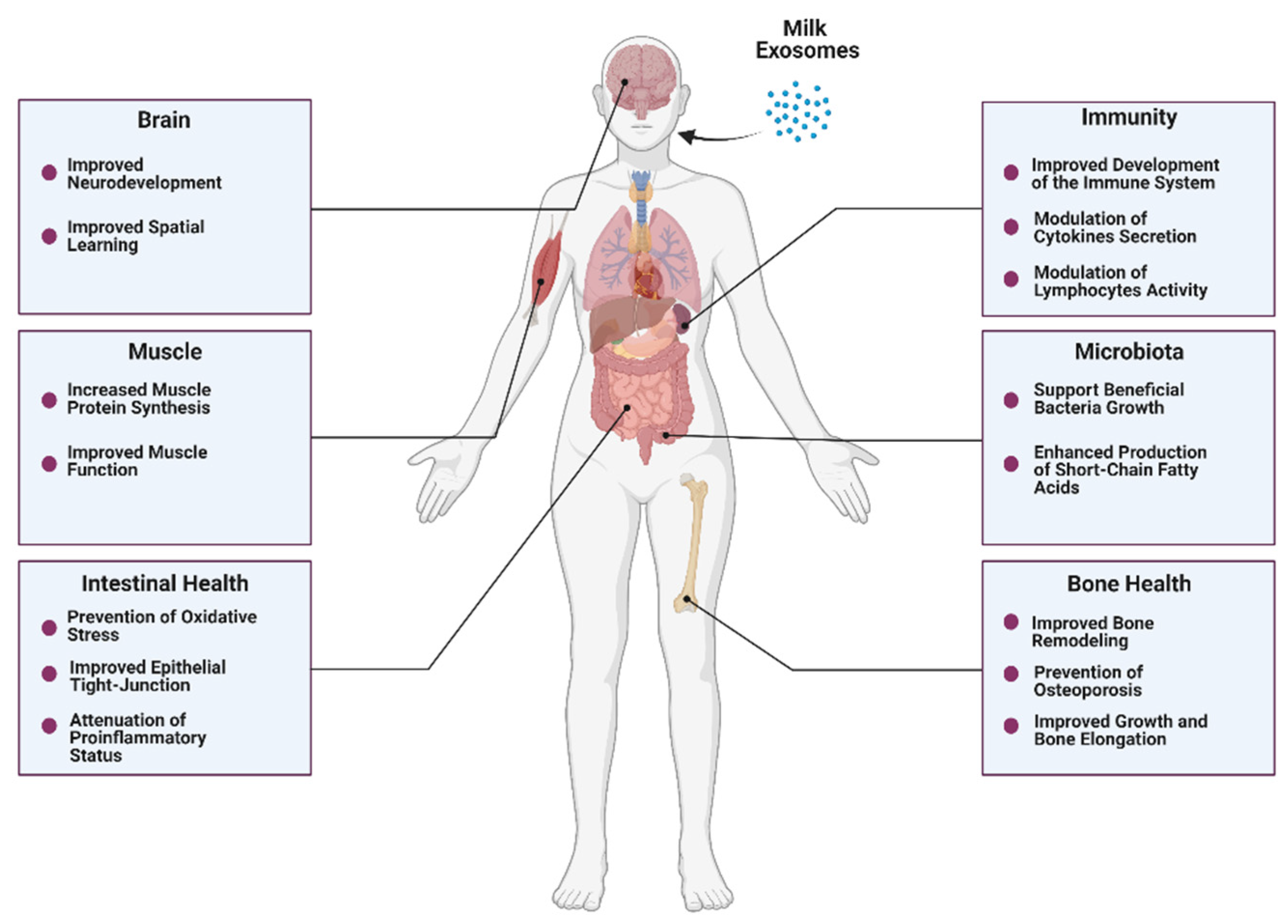
Figure 4.
Potential beneficial effects of milk exosomes on the musculoskeletal system.

Figure 5.
Potential beneficial effects of milk exosomes on the immune system.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
García-Martínez, J.; Pérez-Castillo, Í.M.; Salto, R.; López-Pedrosa, J.M.; Rueda, R.; Girón, M.D. Beneficial Effects of Bovine Milk Exosomes in Metabolic Interorgan Cross-Talk. Nutrients 2022, 14, 1442. https://doi.org/10.3390/nu14071442
AMA Style
García-Martínez J, Pérez-Castillo ÍM, Salto R, López-Pedrosa JM, Rueda R, Girón MD. Beneficial Effects of Bovine Milk Exosomes in Metabolic Interorgan Cross-Talk. Nutrients. 2022; 14(7):1442. https://doi.org/10.3390/nu14071442
Chicago/Turabian StyleGarcía-Martínez, Jorge, Íñigo M. Pérez-Castillo, Rafael Salto, José M. López-Pedrosa, Ricardo Rueda, and María D. Girón. 2022. "Beneficial Effects of Bovine Milk Exosomes in Metabolic Interorgan Cross-Talk" Nutrients 14, no. 7: 1442. https://doi.org/10.3390/nu14071442
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.