Electroantennographic Responses of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae) to a Range of Volatile Compounds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Odor Stimuli
2.3. Electroantennography (EAG)
2.4. Data Analysis
3. Results
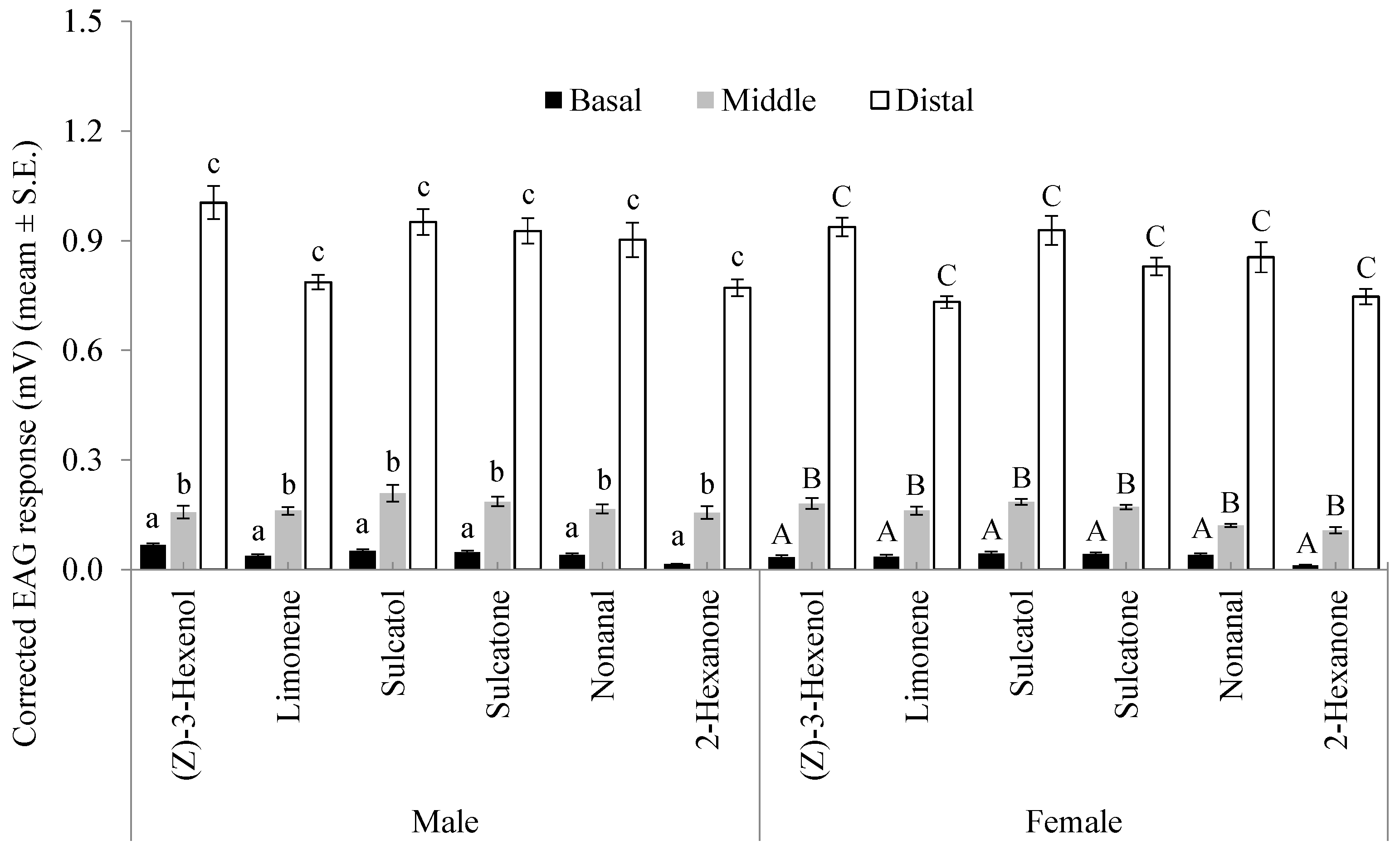
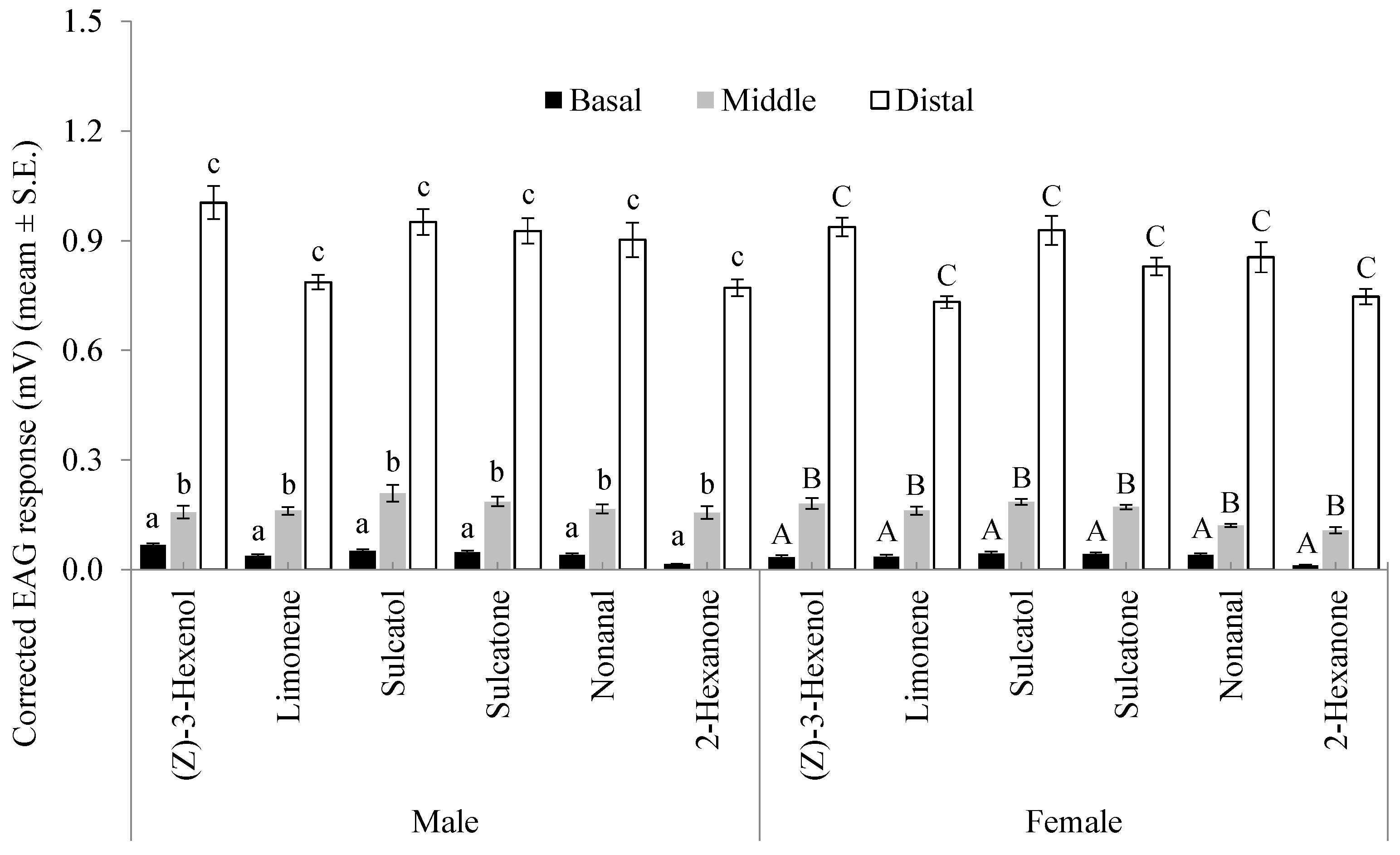
3.1. Chemoreceptivity of Flagellomeres
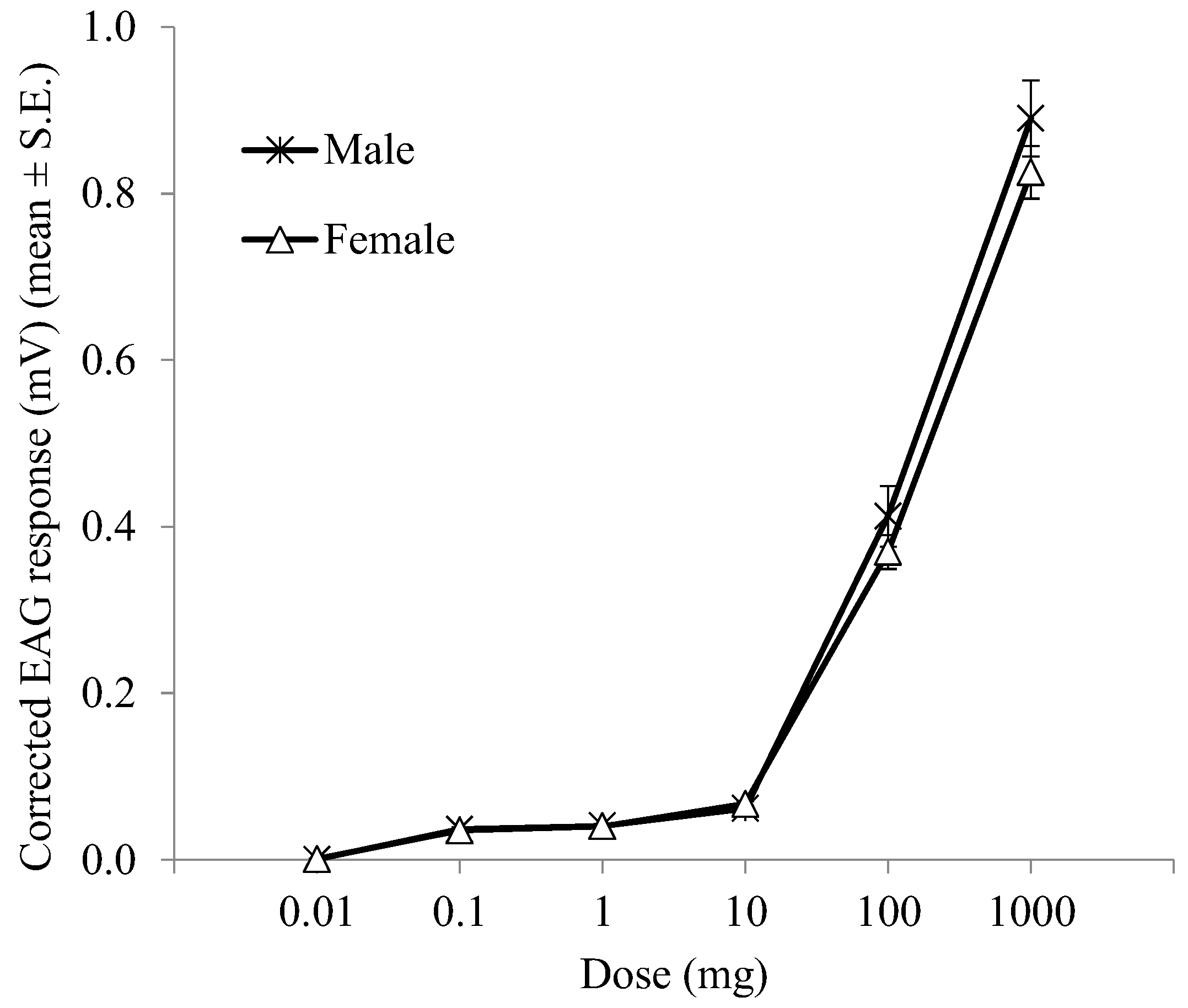
3.2. Sensitivity of Distal Flagellomeres
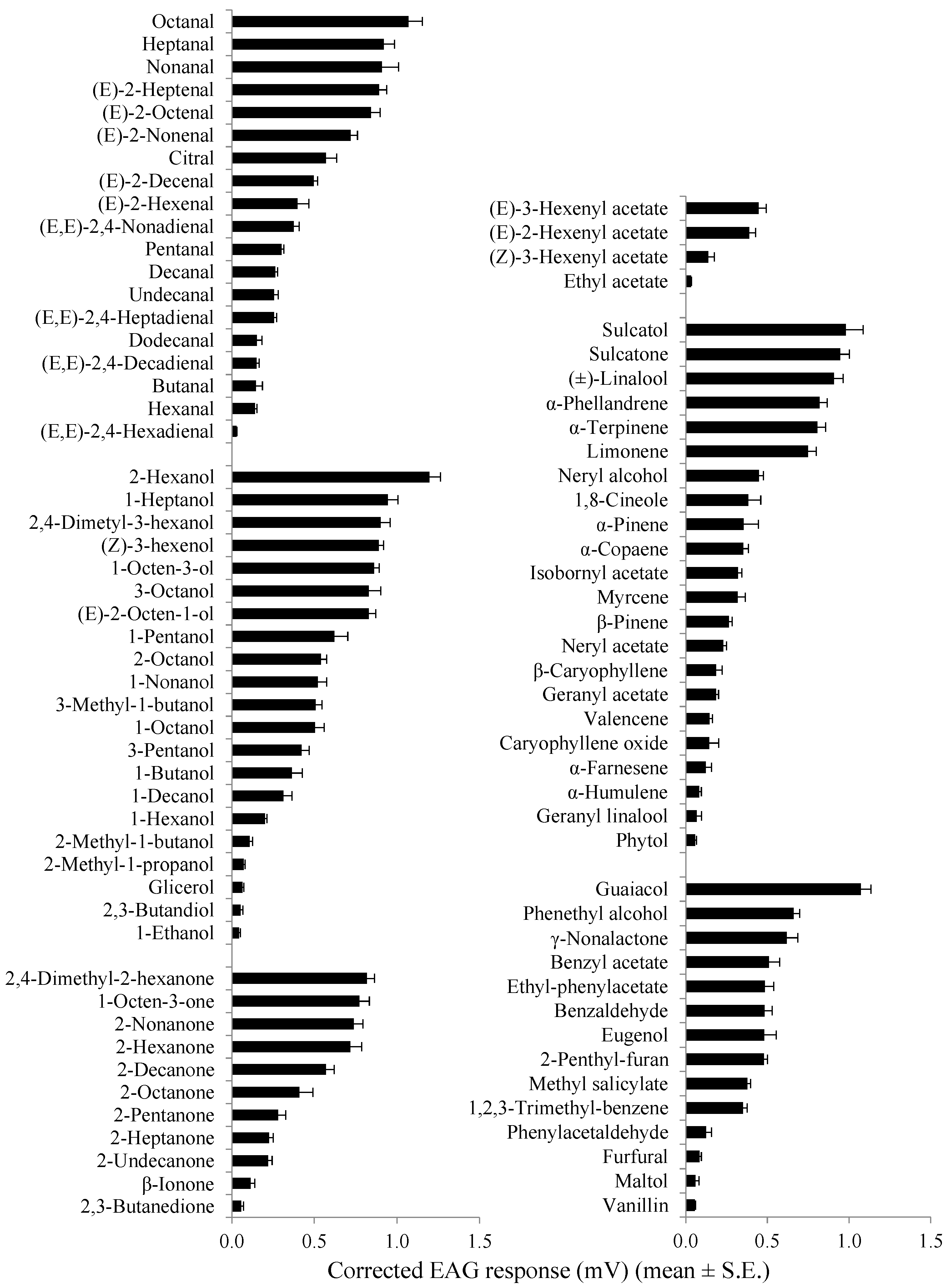
3.3. Selectivity of Distal Flagellomeres
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- CABI. Aromia bungii. In Invasive Species Compendium; CAB International: Wallingford, UK, 2019; Available online: www.cabi.org/isc (accessed on 2 July 2019).
- Hua, L.Z. List of Chinese Insects; Zhongshan (Sun Yat-sen) University Press: Guangzhou, China, 2002; Volume II, p. 612. [Google Scholar]
- Smith, J.W. NPAG report. In Aromia bungii (Faldermann): Redneck Longhorned Beetle Coleoptera/Cerambycidae; New Pest Advisory Group (NPAG), Plant Epidemiology and Risk Analysis Laboratory, Center for Plant Health Science & Technology, APHIS, USDA: Washington, DC, USA, 2009. [Google Scholar]
- EPPO. First Report of Aromia bungii in Germany: Addition to the EPPO Alert List. EPPO Reporting Service. September 2012. Available online: https://gd.eppo.int/reporting/article-1917 (accessed on 2 July 2019).
- EPPO. First Report of Aromia bungii in Italy. EPPO Reporting Service. October 2012. Available online: https://gd.eppo.int/reporting/article-2410 (accessed on 2 July 2019).
- Garonna, A.P.; Nugnes, F.; Epinosa, B.; Griffo, R.; Benchi, D. Aromia bungii, nuovo tarlo asiatico ritrovato in Campania (Aromia bungii, a new Asian worm found in Campania). Inf. Agrar. 2013, 69, 60–62. (In Italian) [Google Scholar]
- Xu, T.; Yasui, H.; Teale, S.A.; Fujiwara-Tsujii, N.; Wickham, J.D.; Fukaya, M.; Hansen, L.; Kiriyama, S.; Hao, D.; Nakano, A.; et al. Identification of a male-produced sex-aggregation pheromone for a highly invasive cerambycid beetle, Aromia bungii. Sci. Rep. 2017, 7, 7330. [Google Scholar] [CrossRef] [PubMed]
- EPPO. Pest Risk Analysis for Aromia bungii; EPPO: Paris, France, 2014; Available online: http://www.eppo.int/QUARANTINE/Pest_Risk_Analysis/PRA_intro.htm (accessed on 2 July 2019).
- Cocquempot, C. Aromia bungii. EPPO datasheet on pests recommended for regulation. EPPO Bull. 2015, 45, 4–8. [Google Scholar] [CrossRef]
- Nugnes, F.; Russo, E.; Garonna, A.P.; Griffo, R. L’attività di monitoraggio ed eradicazione di Aromia bungii (Faldermann) in Campania. In Proceedings of the XXIV Congresso Nazionale Italiano di Entomologia, Orosei, Italy, 9–14 June 2014; p. 113. [Google Scholar]
- European Union. Commission Implementing Decision (EU) 2018/1503 of 8 October 2018 as regards measures to prevent the introduction into and the spread within the Union of Aromia bungii (Faldermann). Off. J. EU 2018, L 254, 9–18. [Google Scholar]
- Gressitt, J.L. Destructive Long-Horned Beetle borers at Canton, China. In Special Publication 1. Lingnan Natural History Survey and Museum; Lingnan University: Canton, China, 1942; pp. 1–60. [Google Scholar]
- Carella, D. Piano d’azione regionale per la lotta al Cerambicide Aromia bungii-VI aggiornamento; Decreto Dirigenziale n. 31. Bollettino Ufficiale della Regione Campania, Italy. 2019; 1–16. [Google Scholar]
- Zou, Y.; Hansen, L.; Xu, T.; Teale, S.A.; Hao, D.; Millar, J.G. Optimizing pheromone-based lures for the invasive red-necked longhorn beetle, Aromia bungii. J. Pest Sci. 2019, 92, 1217–1225. [Google Scholar] [CrossRef]
- Wei, J.R.; Liu, X.B.; Niu, Y.L.; Wang, J.J. Identification of volatiles released from the living adult Aromia bungii Faldermann. For. Pest Dis. 2013, 32, 8–10, (In Chinese with English abstract). [Google Scholar]
- Wang, W.C.; Cao, D.D.; Jin, M.; Wei, J.R. (R)-(+)-citronellal identified as a female-produced sex pheromone of Aromia bungii Faldermann (Coleoptera: Cerambycidae). Egypt. J. Biol. Pest Control 2018, 28, 77. [Google Scholar] [CrossRef]
- Dickens, J.C. Olfaction in the boll weevil Anthonomus grandis Boh (Coleoptera: Curculionidae): Electroantennogram studies. J. Chem. Ecol. 1984, 10, 1759–1785. [Google Scholar] [CrossRef]
- Visser, J.H.; Piron, P.G.M.; Hardie, J. The aphids’ peripheral perception of plant volatiles. Entomol. Exp. Appl. 1996, 80, 35–38. [Google Scholar] [CrossRef]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef]
- Reagel, P.F.; Ginzel, M.D.; Hanks, L.M. Aggregation and mate location in the red milkweed beetle (Coleoptera: Cerambycidae). J. Insect Behav. 2002, 15, 811–830. [Google Scholar] [CrossRef]
- Allison, J.D.; Borden, J.H.; Seybold, S.J. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, L.Y. Mating behavior of a flower visiting longhorn beetle Zorion guttigerum (Westwood) (Coleoptera: Cerambycidae: Cerambycinae). Naturwissenschaften 2005, 92, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Li, J.G.; Jin, Y.J.; Luo, Y.Q.; Xu, Z.C.; Chen, H.J. Leaf volatiles for host tree Acer negundo: Diurnal rhythm and behavior responses of Anoplophora glabripennis to volatiles in field. Acta Bot. Sin. 2003, 45, 177–182. [Google Scholar]
- Nehme, M.E.; Keena, M.A.; Zhang, A.; Baker, T.C.; Hoover, K. Attraction of Anoplophora glabripennis to male-produced pheromone and plant volatiles. Environ. Entomol. 2009, 38, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Collignon, R.M.; Swift, I.P.; Zou, Y.; McElfresh, J.S.; Hanks, L.M.; Millar, J.G. The influence of host plant volatiles on the attraction of longhorn beetles to pheromones. J. Chem. Ecol. 2016, 42, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Millar, J.G.; Hanks, L.M. Chemical ecology of Cerambycids. In Cerambycidae of the World: Biology and Pest Management; Wang, Q., Ed.; CRC Press/Tayler & Francis Group: Boca Raton, FL, USA, 2017; pp. 161–208. [Google Scholar]
- Nakamuta, K.; Leal, W.S.; Nakashima, T.; Tokoro, M. Increase of trap catches by a combination of male sex pheromones and floral attractant in longhorn beetle, Anaglyptus subfasciatus. J. Chem. Ecol. 1997, 23, 1635–1640. [Google Scholar] [CrossRef]
- Silk, P.J.; Lemay, M.A.; LeClair, G.; Sweeney, J.; Magee, D. Behavioral and electrophysiological responses of Tetropium fuscum (Coleoptera: Cerambycidae) to pheromone and spruce volatiles. Environ. Entomol. 2010, 39, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, J.; De Groot, P.; Macdonald, L.; Smith, S.; Cocquempot, C.; Kenis, M.; Gutowski, J.M. Host volatile attractants and traps for detection of Tetropium fuscum (F.), Tetropium castaneum (L.) and other longhorned beetles (Coleoptera: Cerambycidae). Environ. Entomol. 2004, 33, 844–854. [Google Scholar] [CrossRef]
- Fujiwara-Tsujii, N.; Yasui, H.; Wakamura, S.; Hashimoto, I.; Minamishima, M. The white-spotted longicorn beetle, Anoplophora malasiaca (Coleoptera: Cerambycidae), with blueberry as host plant, utilizes host chemicals for male orientation. Appl. Entomol. Zool. 2012, 47, 103–110. [Google Scholar] [CrossRef]
- Yasui, H. Chemical communication in mate location and recognition in the white-spotted longicorn beetle, Anoplophora malasiaca (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 2009, 44, 183–194. [Google Scholar] [CrossRef]
- Yasui, H.; Fujiwara-Tsujii, N.; Wakamura, S. Volatile attractant phytochemicals for a population of white-spotted longicorn beetles Anoplophora malasiaca (Thomson) (Coleoptera: Cerambycidae) fed on willow differ from attractants for a population fed on citrus. Chemoecology 2011, 21, 51–58. [Google Scholar] [CrossRef]
- Ibeas, F.; Gallego, D.; Diez, J.J.; Pajares, J.A. An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerambycidae). J. Appl. Ent. 2007, 131, 13–20. [Google Scholar] [CrossRef]
- Miller, D.R.; Asaro, C.; Crowe, C.M.; Duerr, D.A. Bark beetle pheromones and pine volatiles: Attractant kairomone lure blend for longhorn beetles (Cerambycidae) in pine stands of the southeastern United States. J. Econ. Entomol. 2011, 104, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D. Electrophysiological investigations on the olfactory specificity of sexual attracting substances in different species of moth. J. Insect Physiol. 1962, 8, 15–30. [Google Scholar] [CrossRef]
- Van Der Pers, J.N.C. Comparison of electroantennogram response spectra to plant volatiles in seven species of Yponomeuta and in the torticid Adoxophyes orana. Entomol. Exp. Appl. 1981, 30, 181–192. [Google Scholar] [CrossRef]
- Di Palma, A.; Pistillo, M.; Griffo, R.; Garonna, A.P.; Germinara, G.S. Scanning electron microscopy of the antennal sensilla and their secretion analysis in adults of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae). Insects 2019, 10, 88. [Google Scholar] [CrossRef] [PubMed]
- Kaissling, K.E.; Thorson, J. Insect olfactory sensilla: Structural, chemical and electrical aspects of the functional organization. In Receptors for Neurotransmitters, Hormones and Pheromones in Insects; Sattelle, D.B., Hall, L.M., Hildebrand, J.G., Eds.; Elsevier: Amsterdam, The Netherlands, 1980; pp. 261–282. [Google Scholar]
- Light, M.D.; Kamm, J.A.; Buttery, R.G. Electroantennogram response of alfalfa seed chalcid, Bruchophagus roddi (Hymenoptera: Eurytomidae) to host- and non host-plant volatiles. J. Chem. Ecol. 1992, 18, 333–352. [Google Scholar] [CrossRef]
- Raguso, R.A.; Light, D.M. Electroantennogram responses of male Sphinx perelegans hawkmoths to floral and ‘green leaf volatiles’. Entomol. Exp. Appl. 1998, 86, 287–293. [Google Scholar] [CrossRef]
- Den Otter, C.J.; Tchicaya, T.; Schutte, A.M. Effects of age, sex and hunger on the antennal olfactory sensitivity of tsetse flies. Physiol. Entomol. 1991, 16, 173–182. [Google Scholar] [CrossRef]
- Germinara, G.S.; Ganassi, S.; Pistillo, M.O.; Di Domenico, C.; De Cristofaro, A.; Di Palma, A.M. Antennal olfactory responses of adult meadow spittlebug, Philaenus spumarius, to volatile organic compounds (VOCs). PLoS ONE 2017, 12, e0190454. [Google Scholar] [CrossRef]
- Germinara, G.S.; De Cristofaro, A.; Rotundo, G. Antennal olfactory responses to individual cereal volatiles in Theocolax elegans (Westwood) (Hymenoptera: Pteromalidae). J. Stored Prod. Res. 2009, 45, 195–200. [Google Scholar] [CrossRef]
- Yang, H.; Yang, W.; Liang, X.Y.; Yang, M.F.; Yang, C.P.; Zhu, T.H.; Wu, X.L. The EAG and behavioral responses of Batocera horsfieldi (Coleoptera: Cerambycidae) to the composition of volatiles. J. Kans. Entomol. Soc. 2011, 84, 217–231. [Google Scholar] [CrossRef]
- Elgar, A.; Zhang, D.; Wang, Q.; Wittwer, B.; Pham, H.T.; Johnson, T.L.; Freelance, C.B.; Coquilleau, M. Insect antennal morphology: The evolution of diverse solutions to odorant perception. Yale J. Biol. Med. 2018, 91, 457–469. [Google Scholar] [PubMed]
- Barata, E.N.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M.; Mustaparta, H. Identification of host and nonhost semiochemicals of eucalyptus woodborer Phoracantha semipunctata by gas chromatography-electroantennography. J. Chem. Ecol. 2000, 26, 1877–1895. [Google Scholar] [CrossRef]
- Lopes, O.; Barata, N.E.; Mustaparta, H.; Araújo, J. Fine structures of antennal sensilla basiconica and their detection of plant volatiles in the eucalyptus woodborer, Phoracantha semipunctata Fabricius (Coleoptera, Cerambycidae). Arthropod Struct. Dev. 2002, 31, 1–13. [Google Scholar] [CrossRef]
- Silk, P.J.; Sweeney, J.; Wu, J.; Price, J.; Gutowski, J.M.; Kettela, E.G. Evidence for a male-produced pheromone in Tetropium fuscum (F.) and Tetropium cinnamopterum (Kirby) (Coleoptera: Cerambycidae). Naturwissenschaften 2007, 94, 697–701. [Google Scholar] [CrossRef]
- Yang, H.; Yang, W.; Yang, M.F.; Yang, C.P.; Zhu, T.H.; Huang, Q. Effects of plant volatiles on the EAG and behavioral responses of Batocera horsfieldi Hope (Coleoptera: Cerambycidae). J. Agric. Urban Entomol. 2010, 27, 20–32. [Google Scholar] [CrossRef]
- Raguso, R.A.; Light, D.M.; Pickersky, E. Electroantennogram responses of Hyles lineata (Onagraceae) and other moth-pollinated flowers. J. Chem. Ecol. 1996, 22, 1735–1766. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M. The Pherobase: Database of Insect Pheromones and Semiochemicals. 2019. Available online: http://www.pherobase.com (accessed on 19 June 2019).
- Prokopy, R.J.; Phelan, P.L.; Wright, S.E.; Minalga, A.J.; Barger, R.; Leskey, T.C. Compounds from host fruit odor attractive to adult plum curculios (Coleoptera: Curculionidae). J. Entomol. Sci. 2001, 36, 122–134. [Google Scholar] [CrossRef]
- Schorkopf, D.L.P. Male meliponine bees (Scaptotrigona aff. depilis) produce alarm pheromones to which workers respond with fight and males with flight. J. Comp. Physiol. A 2016, 202, 667–678. [Google Scholar] [CrossRef]
- Torto, B.; Suazo, A.; Alborn, H.; Tumlinson, J.H.; Teal, P.E.A. Response of the small hive beetle (Aethina tumida) to a blend of chemicals identified from honeybee (Apis mellifera) volatiles. Apidologie 2005, 36, 523–532. [Google Scholar] [CrossRef]
- Mikolajczak, K.L.; Zilkowski, B.W.; Smith, C.R., Jr.; Burkholder, W.E. Volatile food attractants for Oryzaephilus surinamensis (L.) from oats. J. Chem. Ecol. 1984, 10, 301–309. [Google Scholar] [CrossRef]
- McGraw, B.A.; Rodriguez-Saona, C.; Holdcraft, R.; Szendrei, Z.; Koppenhfer, A.M. Behavioral and electrophysiological responses of Listronotus maculicollis (Coleoptera: Curculionidae) to volatiles from intact and mechanically damaged annual bluegrass. Environ. Entomol. 2011, 40, 412–419. [Google Scholar] [CrossRef]
- Dettner, K.; Reissenweber, F. The defensive secretion of Omaliinae and Proteininae (Coleoptera: Staphylinidae): Its chemistry, biological and taxonomic significance. Biochem. Syst. Ecol. 1991, 19, 291–303. [Google Scholar] [CrossRef]
- Huber, D.P.W.; Gries, R.; Borden, J.H.; Pierce, H.D., Jr. A survey of antennal responses by five species of coniferophagous bark beetles (Coleoptera: Scolytidae) to bark volatiles of six species of angiosperm trees. Chemoecology 2000, 10, 103–113. [Google Scholar] [CrossRef]
- Obeng-Ofori, D.; Torto, B.; Njagi, P.G.N.; Hassanali, A.; Amiani, H. Fecal volatiles as part of the aggregation pheromone complex of the desert locust, Schistocerca gregaria (Forskal) (Orthoptera: Acrididae). J. Chem. Ecol. 1994, 20, 2077–2087. [Google Scholar] [CrossRef]
- Molnár, B.P.; Tóth, Z.; Kárpáti, Z. Synthetic blend of larval frass volatiles repel oviposition in the invasive box tree moth, Cydalima perspectalis. J. Pest Sci. 2017, 90, 873–885. [Google Scholar] [CrossRef]
- Byrne, K.J.; Swigar, A.A.; Silverstein, R.M.; Borden, J.H.; Stokkink, E. Sulcatol: Population aggregation pheromone in the scolytid beetle, Gnathotrichus sulcatus. J. Insect Physiol. 1974, 20, 1895–2000. [Google Scholar] [CrossRef]
- Borden, J.H.; McLean, J.A. Secondary attraction in Gnathotricus retusus and cross attraction of G. sulcatus (Coleoptera: Scolytidae). J. Chem. Ecol. 1979, 5, 79–88. [Google Scholar] [CrossRef]
- Fletchmann, C.A.H.; Berisford, C.W. Identification of sulcatol, a potential pheromone of the ambrosia beetle Gnathotricus materiarus (Col., Scolytidae). J. Appl. Entomol. 2003, 127, 189–194. [Google Scholar] [CrossRef]
- Quiroz, A.; Petterson, J.; Pickett, J.A.; Wadhams, L.J.; Niemeyer, H.M. Semiochemicals mediating spacing behavior of bird cherry-oat aphid, Rhopalosiphum padi feeding on cereals. J. Chem. Ecol. 1997, 23, 2599–2607. [Google Scholar] [CrossRef]
- Gatti Liguori, P.; Zerba, E.; Gonzalez Audino, P. Anatomical site of pheromone accumulation and temporal pattern of pheromone emission in the ambrosia beetle, Megaplatypus mutatus. Physiol. Entomol. 2011, 36, 201–207. [Google Scholar] [CrossRef]
- Algarvio, R.; Teixeira, C.; Barata, E.; Pickett, J.; Novas, P.C.; Figueiredo, D. Identification of a putative aggregation pheromone from males Platypus cylindrus (Coleoptera: Platypodidae). In Proceedings of the 19th Annual Meeting International Society of Chemical Ecology, Hamburg, Germany, 3–7 August 2002; p. 151. [Google Scholar]
- Borden, J.H.; Wilson, I.M.; Gries, R.; Chong, L.J.; Pierce, H.D., Jr.; Gries, G. Volatiles from the bark of trembling aspen, Populus tremuloides Michx. (Salicaceae) disrupt secondary attraction by the mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera: Scolytidae). Chemoecology 1998, 8, 69–75. [Google Scholar] [CrossRef]
- Bernays, E.A.; Oppenheim, S.; Chapman, R.F.; Kwon, H.; Gould, F. Taste sensitivity of insect herbivores to deterrents is greater in specialists than in generalists: A behavioural test of the hypothesis with two closely related caterpillars. J. Chem. Ecol. 2000, 26, 547–563. [Google Scholar] [CrossRef]
- Linn, C., Jr.; Nojima, S.; Roelofs, W. Antagonistic effects of nonhost fruit volatiles on chemically-mediated discrimination of host fruit by Rhagoletis pomonella flies infesting apple, hawthorn (Crataegus spp.) and flowering dogwood (Cornus florida). Èntomol. Exp. Appl. 2005, 114, 97–105. [Google Scholar] [CrossRef]
- Germinara, G.S.; De Cristofaro, A.; Rotundo, G. Chemical cues for host location by the chestnut gall wasp, Dryocosmus kuriphilus. J. Chem. Ecol. 2011, 37, 49–56. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
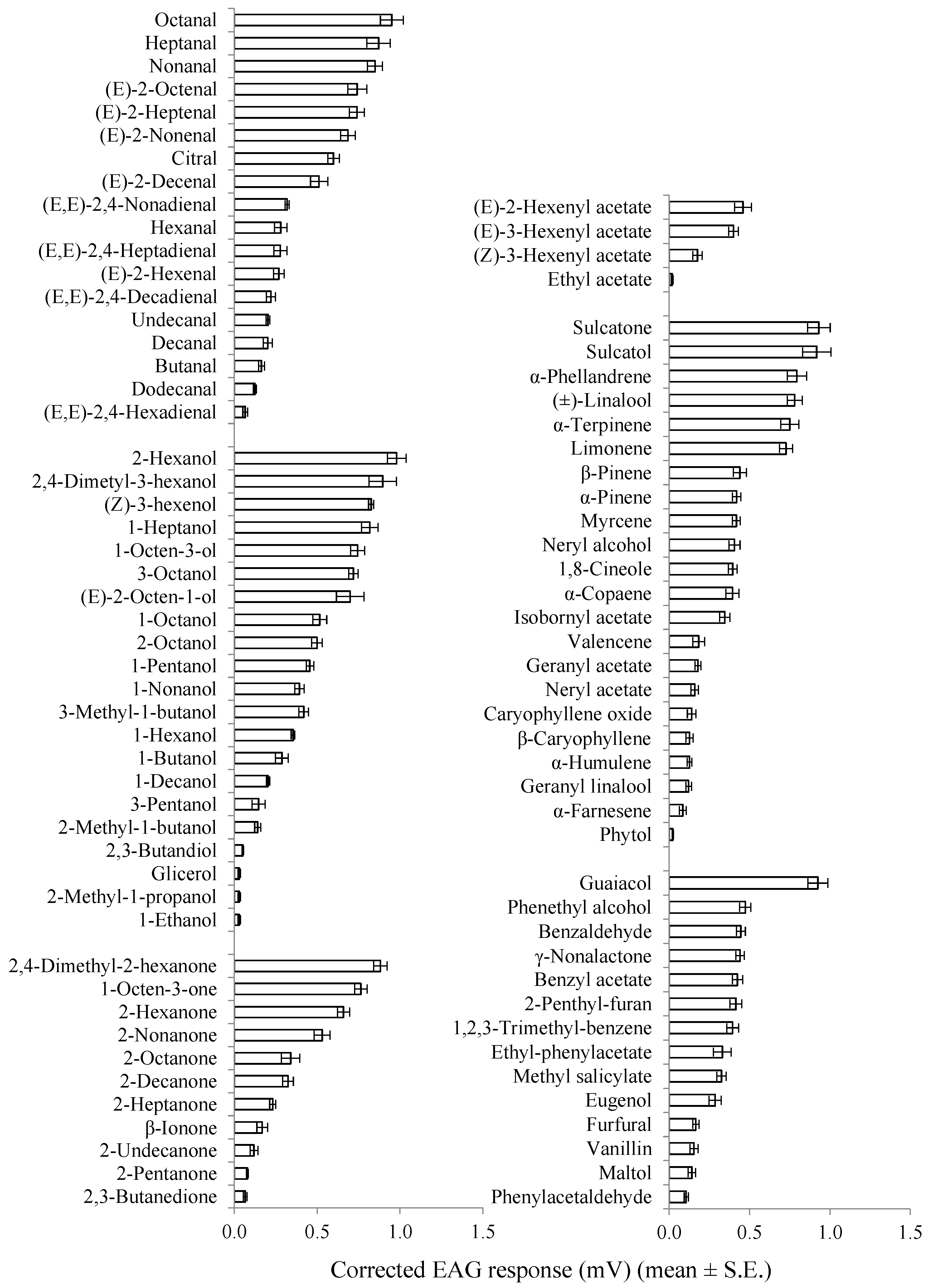
| Class Compound a | Chemical Purity (%) | Corrected EAG Response in mV (Mean ± SE) | |
|---|---|---|---|
| Males | Females | ||
| Aliphatic aldehydes | |||
| Butanal | 99.0 | 0.15 ± 0.04 | 0.16 ± 0.02 |
| Pentanal | 95.0 | 0.30 ± 0.01 | 0.27 ± 0.02 |
| Hexanal ** | 98.0 | 0.14 ± 0.01 | 0.28 ± 0.04 |
| Heptanal | 95.0 | 0.92 ± 0.07 | 0.87 ± 0.07 |
| Octanal | 99.0 | 1.07 ± 0.08 | 0.95 ± 0.07 |
| Nonanal | 95.0 | 0.91 ± 0.10 | 0.85 ± 0.04 |
| Decanal | 95.0 | 0.26 ± 0.01 | 0.20 ± 0.03 |
| Undecanal | 97.0 | 0.26 ± 0.03 | 0.20 ± 0.01 |
| Dodecanal | 92.0 | 0.15 ± 0.03 | 0.12 ± 0.01 |
| (E)-2-Hexenal | 99.0 | 0.40 ± 0.07 | 0.27 ± 0.03 |
| (E)-2-Heptenal * | 97.0 | 0.89 ± 0.05 | 0.74 ± 0.04 |
| (E)-2-Octenal | 94.0 | 0.84 ± 0.05 | 0.74 ± 0.06 |
| (E)-2-Nonenal | 97.0 | 0.72 ± 0.04 | 0.69 ± 0.04 |
| (E)-2-Decenal | 95.0 | 0.50 ± 0.02 | 0.51 ± 0.05 |
| (E,E)-2,4-Hexadienal * | 95.0 | 0.03 ± 0.01 | 0.07 ± 0.01 |
| (E,E)-2,4-Heptadienal | 88.0 | 0.26 ± 0.02 | 0.28 ± 0.04 |
| (E,E)-2,4-Nonadienal | 85.0 | 0.38 ± 0.03 | 0.32 ± 0.01 |
| (E,E)-2,4-Decadienal | 85.0 | 0.15 ± 0.02 | 0.22 ± 0.03 |
| Citral | 95.0 | 0.57 ± 0.07 | 0.60 ± 0.03 |
| Aliphatic alcohols | |||
| 1-Ethanol | 99.8 | 0.04 ± 0.01 | 0.03 ± 0.01 |
| 1-Butanol | 99.0 | 0.36 ± 0.06 | 0.29 ± 0.04 |
| 1-Pentanol | 99.0 | 0.62 ± 0.08 | 0.46 ± 0.02 |
| 3-Pentanol ** | 98.0 | 0.42 ± 0.05 | 0.15 ± 0.04 |
| 1-Hexanol ** | 98.0 | 0.20 ± 0.01 | 0.35 ± 0.01 |
| 2-Hexanol | 99.0 | 1.20 ± 0.07 | 0.98 ± 0.06 |
| 1-Heptanol | 98.0 | 0.94 ± 0.06 | 0.82 ± 0.05 |
| 1-Octanol | 98.0 | 0.50 ± 0.06 | 0.52 ± 0.04 |
| 2-Octanol | 96.0 | 0.54 ± 0.04 | 0.50 ± 0.03 |
| 3-Octanol | 99.0 | 0.83 ± 0.07 | 0.72 ± 0.03 |
| 1-Octen-3-ol | 98.0 | 0.86 ± 0.03 | 0.74 ± 0.04 |
| 1-Nonanol | 98.0 | 0.52 ± 0.05 | 0.39 ± 0.03 |
| 1-Decanol | 98.0 | 0.31 ± 0.05 | 0.20 ± 0.01 |
| (Z)-3-Hexenol | 98.0 | 0.89 ± 0.03 | 0.83 ± 0.05 |
| (E)-2-Octen-1-ol | 97.0 | 0.83 ± 0.04 | 0.70 ± 0.08 |
| 2-Methyl-1-butanol | 99.0 | 0.07 ± 0.01 | 0.03 ± 0.01 |
| 2-Methyl-1-propanol | 99.5 | 0.11 ± 0.02 | 0.14 ± 0.02 |
| 3-Methyl-1-butanol | 99.0 | 0.51 ± 0.04 | 0.42 ± 0.03 |
| 2,4-Dimetyl-3-hexanol | 99.0 | 0.90 ± 0.06 | 0.90 ± 0.90 |
| 2,3-Butandiol | 98.0 | 0.05 ± 0.01 | 0.05 ± 0.01 |
| Glicerol | 99.0 | 0.06 ± 0.01 | 0.03 ± 0.01 |
| Aliphatic ketones | |||
| 2-Pentanone ** | 97.0 | 0.28 ± 0.05 | 0.08 ± 0.01 |
| 2-Hexanone | 99.0 | 0.72 ± 0.07 | 0.66 ± 0.04 |
| 2-Heptanone | 98.0 | 0.22 ± 0.03 | 0.23 ± 0.02 |
| 2-Octanone | 98.0 | 0.41 ± 0.08 | 0.34 ± 0.06 |
| 2-Nonanone * | 99.0 | 0.74 ± 0.05 | 0.53 ± 0.05 |
| 2-Decanone ** | 98.0 | 0.57 ± 0.05 | 0.32 ± 0.03 |
| 2-Undecanone * | 99.0 | 0.22 ± 0.03 | 0.12 ± 0.02 |
| 1-Octen-3-one | 96.0 | 0.77 ± 0.06 | 0.76 ± 0.04 |
| β-Ionone | 95.0 | 0.11 ± 0.02 | 0.17 ± 0.03 |
| 2,4-dimetyl-2-hexanone | 98.0 | 0.82 ± 0.04 | 0.88 ± 0.04 |
| 2,3-Butanedione | 97.0 | 0.06 ± 0.01 | 0.06 ± 0.01 |
| Aliphatic esters | |||
| Ethyl acetate * | 99.0 | 0.03 ± 0.01 | 0.02 ± 0.01 |
| (E)-2-Hexenyl acetate | 98.0 | 0.39 ± 0.04 | 0.46 ± 0.05 |
| (E)-3-Hexenyl acetate | 98.0 | 0.45 ± 0.05 | 0.40 ± 0.03 |
| (Z)-3-Hexenyl acetate | 98.0 | 0.14 ± 0.04 | 0.18 ± 0.03 |
| Terpenes | |||
| α-Pinene | 98.0 | 0.35 ± 0.09 | 0.42 ± 0.03 |
| β-Pinene ** | 98.0 | 0.26 ± 0.02 | 0.44 ± 0.04 |
| Limonene | 97.0 | 0.75 ± 0.05 | 0.73 ± 0.04 |
| α-Farnesene | 95.0 | 0.12 ± 0.04 | 0.08 ± 0.02 |
| β-Caryophyllene | 80.0 | 0.19 ± 0.04 | 0.13 ± 0.02 |
| Myrcene | 92.0 | 0.32 ± 0.05 | 0.42 ± 0.02 |
| α-Terpinene | 95.0 | 0.81 ± 0.05 | 0.75 ± 0.06 |
| 1.8-Cineole | 99.0 | 0.38 ± 0.08 | 0.40 ± 0.03 |
| (±)-Linalool | 99.0 | 0.91 ± 0.06 | 0.78 ± 0.05 |
| Sulcatone (6-Methyl-5-hepten-2-one) | 99.0 | 0.95 ± 0.06 | 0.91 ± 0.07 |
| Sulcatol (6-Methyl-5-hepten-2-ol) | 99.0 | 0.98 ± 0.11 | 0.92 ± 0.09 |
| Neryl alcohol | 98.0 | 0.45 ± 0.03 | 0.41 ± 0.03 |
| Neryl acetate * | 98.0 | 0.23 ± 0.02 | 0.16 ± 0.02 |
| Geranyl acetate | 97.0 | 0.18 ± 0.02 | 0.18 ± 0.02 |
| Isobornyl acetate | 95.0 | 0.32 ± 0.02 | 0.35 ± 0.03 |
| α-Phellandrene | 95.0 | 0.82 ± 0.05 | 0.79 ± 0.06 |
| Geranyl linalool | 95.0 | 0.07 ± 0.03 | 0.12 ± 0.02 |
| Phytol * | 97.0 | 0.06 ± 0.01 | 0.02 ± 0.01 |
| α-Copaene | 90.0 | 0.35 ± 0.03 | 0.39 ± 0.04 |
| Valencene | 70.0 | 0.15 ± 0.02 | 0.18 ± 0.04 |
| α-Farnesene | 95.0 | 0.12 ± 0.04 | 0.08 ± 0.02 |
| β-Caryophyllene | 80.0 | 0.19 ± 0.04 | 0.13 ± 0.02 |
| Caryophyllene oxide | 95.0 | 0.14 ± 0.06 | 0.14 ± 0.03 |
| α-Humulene | 96.0 | 0.08 ± 0.01 | 0.13 ± 0.01 |
| Aromatics | |||
| Guaiacol | 98.0 | 1.07 ± 0.06 | 0.93 ± 0.06 |
| Eugenol * | 99.0 | 0.48 ± 0.07 | 0.29 ± 0.04 |
| Maltol * | 99.0 | 0.06 ± 0.02 | 0.14 ± 0.02 |
| Phenethyl alcohol ** | 99.0 | 0.66 ± 0.04 | 0.47 ± 0.04 |
| Furfural | 99.0 | 0.08 ± 0.01 | 0.17 ± 0.02 |
| Phenylacetaldehyde | 90.0 | 0.12 ± 0.03 | 0.11 ± 0.01 |
| Vanillin | 99.0 | 0.06 ± 0.01 | 0.15 ± 0.03 |
| Benzaldehyde | 99.0 | 0.48 ± 0.05 | 0.45 ± 0.03 |
| Methyl salicylate | 99.0 | 0.38 ± 0.02 | 0.33 ± 0.03 |
| 1,2,3-Trimethyl-benzene | 90.0 | 0.35 ± 0.03 | 0.39 ± 0.04 |
| γ-Nonalactone * | 98.0 | 0.62 ± 0.07 | 0.44 ± 0.03 |
| 2-Penthyl-furan | 98.0 | 0.48 ± 0.02 | 0.41 ± 0.04 |
| Benzyl acetate | 99.0 | 0.51 ± 0.07 | 0.42 ± 0.03 |
| Ethyl-phenylacetate | 98.0 | 0.48 ± 0.05 | 0.33 ± 0.06 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germinara, G.S.; Pistillo, M.; Griffo, R.; Garonna, A.P.; Di Palma, A. Electroantennographic Responses of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae) to a Range of Volatile Compounds. Insects 2019, 10, 274. https://doi.org/10.3390/insects10090274
Germinara GS, Pistillo M, Griffo R, Garonna AP, Di Palma A. Electroantennographic Responses of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae) to a Range of Volatile Compounds. Insects. 2019; 10(9):274. https://doi.org/10.3390/insects10090274
Chicago/Turabian StyleGerminara, Giacinto S., Marco Pistillo, Raffaele Griffo, Antonio P. Garonna, and Antonella Di Palma. 2019. "Electroantennographic Responses of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae) to a Range of Volatile Compounds" Insects 10, no. 9: 274. https://doi.org/10.3390/insects10090274