

Attenuation of Pseudomonas aeruginosa Virulence by Pomegranate Peel Extract

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Microbial Cells and Growth Conditions
2.2. Pomegranate Peel Extract
2.3. Total Microbial Growth, Biofilm Formation and Regrowth Assays
2.4. Mass Spectrometry Analysis of PomeGr Extract
2.5. Mass Spectrometry Analysis of P. aeruginosa AI
2.6. Statistical Analysis
3. Results
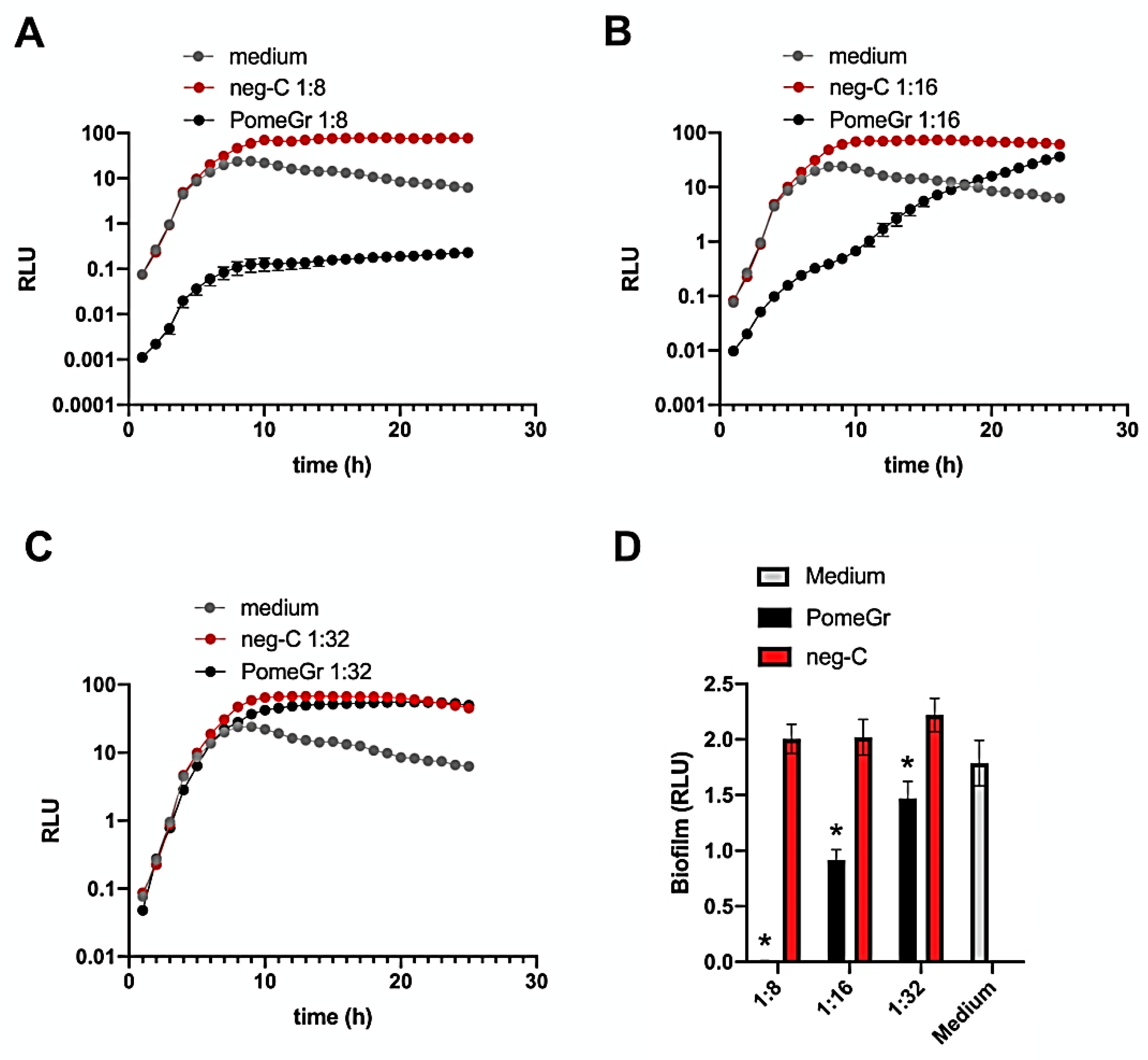
3.1. PomeGr Effects on Pseudomonas Growth and Biofilm Formation
3.2. Phenolic Compounds Profile of PomeGr Exposed or Not to P. aeruginosa
3.3. PomeGr Effects on AI Release by P. aeruginosa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gale, M.J.; Maritato, M.S.; Chen, Y.L.; Abdulateef, S.S.; Ruiz, J.E. Pseudomonas aeruginosa causing inflammatory mass of the nasopharynx in an immunocompromised HIV infected patient: A mimic of malignancy. IDCases 2015, 2, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Otto, M. Molecular basis of in vivo biofilm formation by bacterial pathogens. Chem. Biol. 2012, 19, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Zur, K.B.; Mandell, D.L.; Gordon, R.E.; Holzman, I.; Rothschild, M.A. Electron microscopic analysis of biofilm on endotracheal tubes removed from intubated neonates. Otolaryngol.–Head Neck Surg. 2004, 130, 407–414. [Google Scholar] [CrossRef]
- Ryder, C.; Byrd, M.; Wozniak, D.J. Role of polysaccharides in Pseudomonas aeruginosa biofilm development. Curr. Opin. Microbiol. 2007, 10, 644–648. [Google Scholar] [CrossRef]
- Cécile, R.; Laurent, G.; Jean, G. Biofilm-detached cells, a transition from a sessile to a planktonic phenotype: A comparative study of adhesion and physiological characteristics in Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2008, 290, 135–142. [Google Scholar]
- Mah, T.F.; O’Toole, G.A. Mechanisms of Biofilm Resistance to Antimicrobial Agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Venturi, V. Regulation of quorum sensing in Pseudomonas. FEMS Microbiol. Rev. 2006, 30, 274–291. [Google Scholar] [CrossRef]
- Pesci, E.C.; Pearson, J.P.; Seed, P.C.; Iglewski, B.H. Regulation of las and rhl quorum sensing in Pseudomonas aeruginosa. J. Bacteriol. 1997, 179, 3127–3132. [Google Scholar] [CrossRef]
- Dubern, J.F.; Diggle, S.P. Quorum sensing by 2-alkyl-4-quinolones in Pseudomonas aeruginosa and other bacterial species. Mol. Biosyst. 2008, 4, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Wu, J.; Deng, Y.; Wang, J.; Wang, C.; Wang, J.; Chang, C.; Dong, Y.; Williams, P.; Zhang, L.H. A cell-cell communication signal integrates quorum sensing and stress response. Nat. Chem. Biol. 2013, 9, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 2016, 6, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, T.H.; Bjarnsholt, T.; Jensen, P.Ø.; Givskov, M.; Høiby, N. Targeting quorum sensing in Pseudomonas aeruginosa biofilms: Current and emerging inhibitors. Future Microbiol. 2013, 8, 901–921. [Google Scholar] [CrossRef]
- Ueda, A.; Wood, T.K. Connecting quorum sensing, c-di-GMP, pel polysaccharide, and biofilm formation in Pseudomonas aeruginosa through tyrosine phosphatase TpbA (PA3885). PLoS Pathog. 2009, 5, e1000483. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.Ø.; Bjarnsholt, T.; Phipps, R.; Rasmussen, T.B.; Calum, H.; Christoffersen, L.; Moser, C.; Williams, P.; Pressler, T.; Givskov, M.; et al. Rapid necrotic killing of polymorphonuclear leukocytes is caused by quorum-sensing-controlled production of rhamnolipid by Pseudomonas aeruginosa. Microbiology 2007, 153 (Pt 5), 1329–1338. [Google Scholar] [CrossRef]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Shrout, J.D.; Chopp, D.L.; Just, C.L.; Hentzer, M.; Givskov, M.; Parsek, M.R. The impact of quorum sensing and swarming motility on Pseudomonas aeruginosa biofilm formation is nutritionally conditional. Mol. Microbiol. 2006, 62, 1264–1277. [Google Scholar] [CrossRef]
- Banin, E.; Vasil, M.L.; Greenberg, E.P. Iron and Pseudomonas aeruginosa biofilm formation. Proc. Natl. Acad. Sci. USA 2005, 102, 11076–11081. [Google Scholar] [CrossRef]
- Ciofu, O.; Tolker-Nielsen, T.; Jensen, P.Ø.; Wang, H.; Høiby, N. Antimicrobial resistance, respiratory tract infections and role of biofilms in lung infections in cystic fibrosis patients. Adv. Drug Deliv. Rev. 2015, 85, 7–23. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. β-Lactams and β-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.-G.; Liu, Y.-C.; Chang, C.-Y. Inhibiting N-acylhomoserine lactone synthesis and quenching Pseudomonas quinolone quorum sensing to attenuate virulence. Front. Microbiol. 2016, 6, 1173–1179. [Google Scholar]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic compounds as beneficial phytochemicals in pomegranate (Punica granatum L.) peel: A review. Food Chem. 2018, 261, 75–86. [Google Scholar] [CrossRef]
- Odorici, A.; Colombari, B.; Bellini, P.; Meto, A.; Venturelli, I.; Blasi, E. Novel Options to Counteract Oral Biofilm Formation: In Vitro Evidence. Int. J. Environ. Res. Public Health 2022, 19, 8056. [Google Scholar] [CrossRef]
- Colombari, B.; Tagliazucchi, D.; Odorici, A.; Pericolini, E.; Foltran, I.; Pinetti, D.; Meto, A.; Peppoloni, S.; Blasi, E. Pomegranate Extract Affects Fungal Biofilm Production: Consumption of Phenolic Compounds and Alteration of Fungal Autoinducers Release. Int. J. Environ. Res. Public Health 2022, 19, 14146. [Google Scholar] [CrossRef]
- Abu El-Wafa, W.M.; Ahmed, R.H.; Ramadan, M.A. Synergistic effects of pomegranate and rosemary extracts in combination with antibiotics against antibiotic resistance and biofilm formation of Pseudomonas aeruginosa. Braz. J. Microbiol. 2020, 51, 1079–1092. [Google Scholar] [CrossRef]
- Choi, K.H.; Schweizer, H.P. Mini-Tn7 insertion in bacteria with single attTn7 sites: Example Pseudomonas aeruginosa. Nat. Protoc. 2006, 1, 153–161. [Google Scholar] [CrossRef]
- Pericolini, E.; Colombari, B.; Ferretti, G.; Iseppi, R.; Ardizzoni, A.; Girardis, M.; Sala, A.; Peppoloni, S.; Blasi, E. Real-time monitoring of Pseudomonas aeruginosa biofilm formation on endotracheal tubes in vitro. BMC Microbiol. 2018, 18, 84–93. [Google Scholar] [CrossRef]
- Colombari, B.; Alfano, G.; Gamberini, C.; Cappelli, G.; Blasi, E. EDTA and Taurolidine Affect Pseudomonas aeruginosa Virulence In Vitro-Impairment of Secretory Profile and Biofilm Production onto Peritoneal Dialysis Catheters. Microbiol. Spectr. 2021, 9, e0104721. [Google Scholar] [CrossRef]
- Peppoloni, S.; Pericolini, E.; Colombari, B.; Pinetti, D.; Cermelli, C.; Fini, F.; Prati, F.; Caselli, E.; Blasi, E. The b-lactamase inhibitor boronic acid derivative SM23 as a new anti-Pseudomonas aeruginosa biofilm. Front. Microbiol. 2020, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- Kupnik, K.; Primožič, M.; Vasić, K.; Knez, Ž.; Leitgeb, M. A Comprehensive Study of the Antibacterial Activity of Bioactive Juice and Extracts from Pomegranate (Punica granatum L.) Peels and Seeds. Plants 2021, 10, 1554. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.; Gullón, B.; Gullón, P.; Gomes, A.; Tavaria, F. Bioactive packaging using antioxidant extracts for the prevention of microbial food-spoilage. Food Funct. 2016, 7, 3273–3282. [Google Scholar] [CrossRef]
- Gosset-Erard, C.; Zhao, M.; Lordel-Madeleine, S.; Ennahar, S. Identification of punicalagin as the bioactive compound behind the antimicrobial activity of pomegranate (Punica granatum L.) peels. Food Chem. 2021, 352, 129396. [Google Scholar] [CrossRef] [PubMed]
- Pirzadeh, M.; Caporaso, N.; Rauf, A.; Shariati, M.A.; Yessimbekov, Z.; Khan, M.U.; Imran, M.; Mubarak, M.S. Pomegranate as a source of bioactive constituents: A review on their characterization, properties and applications. Crit. Rev. Food Sci. Nutr. 2020, 61, 982–999. [Google Scholar] [CrossRef]
- Nagoba, B.S.; Deshmukh, S.R.; Wadher, B.J.; Mahabaleshwar, L.; Gandhi, R.C.; Kulkarni, P.B.; Mane, V.A.; Deshmukh, J.S. Treatment of superficial pseudomonal infections with citric acid: An effective and economical approach. J. Hosp. Infect. 1998, 40, 155–157. [Google Scholar] [CrossRef]
- Stanojevic, D.; Comic, I.; Stefanovic, O.; Solujic-Sukdolak, S.L. Antimicrobial effects of sodium benzoate, sodium nitrite and potassium sorbate and their synergistic action in vitro. Bulg. J. Agric. Sci. 2009, 15, 307–311. [Google Scholar]
- Cao, B.; Christophersen, L.; Thomsen, K.; Sonderholm, M.; Bjarnsholt, T.; Jensen, P.O. Antibiotic penetration and bacterial killing in a Pseudomonas aeruginosa biofilm model. J. Antimicrob. Chemother. 2015, 70, 2057–2063. [Google Scholar] [CrossRef]
- Kamble, E.; Pardesi, K. Antibiotic Tolerance in Biofilm and Stationary-Phase Planktonic Cells of Staphylococcus aureus. Microb. Drug Resist. 2021, 27, 3–12. [Google Scholar] [CrossRef]
- Bar-Ya’akov, I.; Tian, L.; Amir, R.; Holland, D. Primary metabolites, anthocyanins, and hydrolyzable tannins in the pomegranate fruit. Front. Plant Sci. 2019, 10, 1–19. [Google Scholar] [CrossRef]
- Paczkowski, J.E.; Mukherjee, S.; McCready, A.R.; Cong, J.P.; Aquino, C.J.; Kim, H.; Henke, B.R.; Smith, C.D.; Bassler, B.L. Flavonoids Suppress Pseudomonas aeruginosa Virulence through Allosteric Inhibition of Quorum-sensing Receptors. J. Biol. Chem. 2017, 292, 4064–4076. [Google Scholar] [CrossRef] [PubMed]
- Sarabhai, S.; Sharma, P.; Capalash, N. Ellagic acid derivatives from Terminalia chebula Retz. downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS ONE 2013, 8, e53441. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Treatment | 3-oxo-C12-HSL | C4-HSL | PQS | IQS |
|---|---|---|---|---|
| Medium | 1.65 × 108 | 3.15 × 108 | 2.51 × 1010 | 2.09 × 106 |
| PomeGr 1:8 | 1.89 × 106 | 3.99 × 107 | 4.42 × 106 | n.d. |
| PomeGr 1:16 | 6.58 × 107 | 3.92 × 107 | 1.93 × 108 | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peppoloni, S.; Colombari, B.; Tagliazucchi, D.; Odorici, A.; Ventrucci, C.; Meto, A.; Blasi, E. Attenuation of Pseudomonas aeruginosa Virulence by Pomegranate Peel Extract. Microorganisms 2022, 10, 2500. https://doi.org/10.3390/microorganisms10122500
Peppoloni S, Colombari B, Tagliazucchi D, Odorici A, Ventrucci C, Meto A, Blasi E. Attenuation of Pseudomonas aeruginosa Virulence by Pomegranate Peel Extract. Microorganisms. 2022; 10(12):2500. https://doi.org/10.3390/microorganisms10122500
Chicago/Turabian StylePeppoloni, Samuele, Bruna Colombari, Davide Tagliazucchi, Alessandra Odorici, Cristiano Ventrucci, Aida Meto, and Elisabetta Blasi. 2022. "Attenuation of Pseudomonas aeruginosa Virulence by Pomegranate Peel Extract" Microorganisms 10, no. 12: 2500. https://doi.org/10.3390/microorganisms10122500
APA StylePeppoloni, S., Colombari, B., Tagliazucchi, D., Odorici, A., Ventrucci, C., Meto, A., & Blasi, E. (2022). Attenuation of Pseudomonas aeruginosa Virulence by Pomegranate Peel Extract. Microorganisms, 10(12), 2500. https://doi.org/10.3390/microorganisms10122500