
Stress Resistance of Saccharomyces cerevisiae Strains Overexpressing Yeast Polyphosphatases

Federal Research Center, Pushchino Scientific Center for Biological Research of the Russian Academy of Sciences, Skryabin Institute of Biochemistry and Physiology of Microorganisms, 142290 Pushchino, Russia
*
Author to whom correspondence should be addressed.
Stresses 2022, 2(1), 17-25; https://doi.org/10.3390/stresses2010002
Submission received: 24 December 2021
/
Revised: 12 January 2022
/
Accepted: 21 January 2022
/
Published: 24 January 2022
Abstract
:Inorganic polyphosphate (polyP) is an important factor in the stress resistance of microorganisms. The polyphosphate-overexpressing strains of yeast S. cerevisiae were used as a model for studying the inter-relationship between stress resistance and polyP level. We compared the polyP level and resistance to the oxidative, manganese, cadmium, and alkaline stresses in parent stain CRN and in strains overexpressing the four yeast polyphosphatases: Ppx1, Ppn1, Ppn2, and Ddp1. Strains overexpressing Ppx1, Ppn1, and Ppn2 have lower polyP content and the strain overexpressing Ddp1 has the same polyP content as the parent strain. The strains overexpressing Ppn1 and Ddp1 show higher resistance to peroxide and manganese. The strain overexpressing Ppx1 showed a decrease in peroxide resistance. The strain overexpressing Ppn2 was more resistant to alkaline and peroxide stresses. A similar increase in resistance to the manganese and peroxide stresses of strains overexpressing Ppn1 and Ddp1, which differ in polyP content, led to the conclusion that there is no direct relationship between polyP content and variations in this resistance. Thus, we speculate about the potential role of inositol pyrophosphates as signaling molecules in stress response.
1. Introduction
Inorganic polyphosphate (polyP), a linear polymer containing a few to several hundred orthophosphate residues, participates in various processes regulating vital activities [1,2,3,4], including stress response in microbial cells [1,5,6,7,8,9]. PolyP and polyphosphate kinases are involved in the induction of the synthesis of RpoS, an RNA polymerase subunit in bacteria that provides the expression of the genes responsible for adaptation to the stationary stage and are necessary for bacterial cell motility, biofilm formation, and virulence [1]. PolyP prevents iron stress and oxidative DNA damage by cisplatin, an anticancer drug and antimicrobial agent; polyP is a crucial iron reservoir that blocks ROS formation during iron stress [10]. PolyP synthesis is vital for controlling cellular adenosine 5’-triphosphate homeostasis, and maintaining both respiratory and photosynthetic electron transport upon sulfur deprivation in Chlamydomonas reinhardtii [11].
PolyP is an important factor in overcoming heavy metal stresses [12]. Cu2+ resistance depends on polyP in many microbial species [13]. The increase in the number of polyP inclusions in the cells of Ochrobactrum anthropi directly correlate with the concentration of Cr(III) in culture medium [14]. The Mn–phosphate complex (Mn–Pi) plays a key role in cellular resistance to oxidative stress [15]. In the cells of radioresistant bacterium Deinococcus radiodurans, the PPKDr gene encoding polyphosphate kinase and the PPXDr gene encoding Mn-dependent polyphosphatase exhibited different expression profiles under oxidative stress [15]. First, PPKDr was upregulated, resulting in the accumulation of polyP, which chelated Mn2+ ions. Second, ppkDr expression level decreased, while ppxDr was upregulated and polyphosphatase hydrolyzed polyP-Mn, followed by phosphate (Pi) release and Mn–Pi formation [16].
Yeast can accumulate high polyP levels [17,18]. In yeast, this polyanion participate is cell cycle regulation [19] in overcoming alkali [20] and manganese stresses [21]. The possible role of polyP in the virulence of Candida albicans, Cryptococcus neoformans, and other fungal pathogens is under discussion [22,23]. Under manganese stress, an increase in the content and chain length of acid-soluble polyP was observed in the cells of Saccharomyces cerevisiae [24] and Cryptococcus humicola [25]. In the cytoplasm of Candida guilliermondii, the size of electron-dense volutin granules containing polyP increased under the influence of X-irradiation [26].
The significant role of polyP in stress overcoming by microorganisms is due to their ability to accumulate large polyP pools in depending on environmental conditions. Chemical properties that determine the participation of polyP in stress overcoming are as follows: being polyanions, polyP chelate heavy metal ions [1]; hydrolysis of polyP provides formation of phosphate, which is necessary for restoring of intracellular pH at alkaline stress [20] and formation of Mn–P complexes at oxidative stress [15].
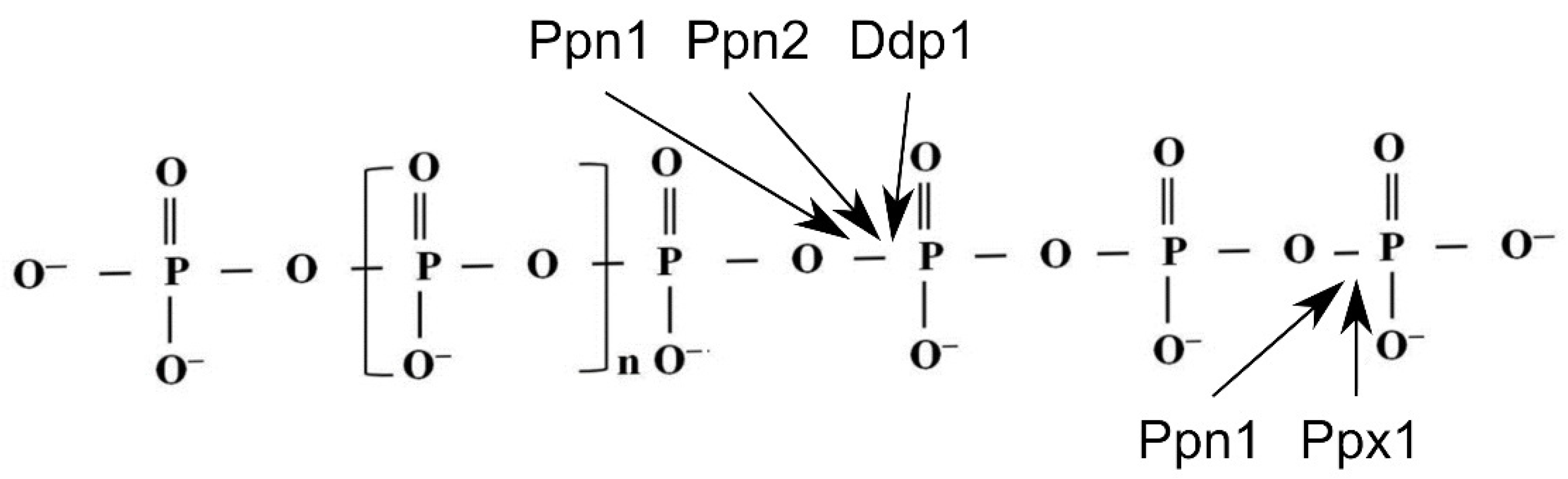
The multicomponent system of polyP metabolism in yeast includes polyP-degrading enzymes with exopolyphosphatase and endopolyphosphatase activities; exopolyphosphatases (polyphosphate phosphohydrolases, EC 3.6.1.11) cleave Pi from the end of the polyP chain, whereas endopolyphosphatases (polyphosphate depolymerases, EC 3.6.1.10.) split long polyP molecules to shorter ones. The four enzymes of S. cerevisiae involved in polyP degradation are exopolyphosphatase Ppx1 [27], dual exo/endopolyphoshatase Ppn1 [28], and endopolyphosphatases Ddp1 [29] and Ppn2 [30,31]. Polyphosphatases vary in the mechanisms of polyP hydrolysis (Figure 1), ability to cleave polyPs of different chain lengths, cellular localization and probably the role in polyP metabolism [27,28,29,30,31]; they do not show obvious similarities in amino acid sequences and belong to different protein families.
We constructed polyphosphatase-overexpressing yeast strains using a vector with a strong constitutive promoter from the S. cerevisiae aldehyde-triphosphate dehydrogenase (TDH3) gene and the ∆ppn1 null mutant as a recipient strain [31]. Earlier, we showed that the Ppn1-overexpressing strain shows higher resistance to manganese and oxidative stresses [21], while the Ppn2-overexpressing strain is more resistant to alkaline and oxidative stresses [32]. Different culture media with various Pi levels were used in our previous studies. PolyP level in yeast varies is highly dependent on culture conditions. Hence, this study compares resistance to oxidative, alkaline, and heavy metal stresses of the parent strain and four strains overexpressing polyphosphatases Ppx1, Ppn1, Ppn2, and Ddp1, cultivated under the same culture conditions, including growth stage and Pi concentration.
2. Materials and Methods
2.1. Strains and Growth Conditions
S. cerevisiae strains overexpressing polyphosphatases Ppn1 (CRN/PPN1), Ppx1 (CRN/PPX1), Ddp1 (CRN/DDP1), and Ppn2 (CRN/PPN2) were constructed in our previous study [31,33] on the basis of the CRN parent strain [34]. The strains’ genotypes are indicated in Supplementary Table S1. We used strain CRN (MATa ade2 his3 ura3 ppn1∆::CgTRP1) as recipient because the wild-type strain transformed with the vector containing PPN1 gene had significantly reduced viability.
Strain CRN was maintained on an agarized synthetic YNB minimal medium with uracil; transformant strains were maintained on an agarized synthetic YNB minimal medium without uracil [33]. The medium contained (per 1 L): bacto-yeast nitrogen bases (Difco, Detroit, MI, USA), 6.7 g; glucose, 20 g; L-tryptophan, L-histidine, L-methionine and adenine, 20 mg of each; and L-leucine, 60 mg. For the cultivation of CRN, uracil was added at a concentration of 20 mg/L.
For the experiments, all yeast strains were cultured in a YPD medium (2% glucose, 2% peptone, 1% yeast extract, w/v) for 24 h at 29 °C with shaking at 145 rpm. The medium contained 3 mM phosphate (Pi).
2.2. Pi and polyP Extraction and Assay
Biomass samples were separated from the culture medium by centrifugation at 5000× g for 15 min, and washed twice with distilled water using centrifugation at 5000× g for 10 min. Acid-soluble polyP1 was extracted from biomass with 0.5 M HClO4 at 0 °C as described earlier [33]. PolyP content in the extracts was estimated by phosphate releasing after 20 min treatment with 0.5 M HClO4 at 90 °C. The amount of acid-insoluble polyP2 remaining in the biomass was estimated by the phosphate released after 20 min treatment of the biomass samples with 0.5 M HClO4 at 100 °C. Phosphate was measured by a colorimetric method [35]. The amount of polyP was normalized to the weight of biomass samples standardized by using centrifugation conditions applied in our previous studies, 5000× g for 15 min [33].
2.3. Polyphosphatase Activity Assay
The exopolyphosphatase and endopolyphosphatase activities were measured in cell-free extract and crude membrane fraction. Yeast cells were harvested, washed twice with distilled water at 5000× g for 10 min, and spheroplasts were obtained as described earlier [33]. Spheroplasts were broken in a glass Potter-Elvehjem homogenizer in 0.1 M sorbitol, 25 mM Tris-HCl, pH 7.2, supplemented with 0.5 mM phenylmethylsulfonyl fluoride (Sigma-Aldrich, St. Louis, MO, USA), and centrifuged at 5000× g for 10 min. The supernatant was collected and centrifuged at 15,000× g for 60 min. The obtained supernatants were cell-free extracts. Pellets (crude membrane fraction) were resuspended in 25 mM Tris-HCl, pH 7.2.
Exopolyphosphatase activity was determined at 30 °C by phosphate formation rate [33]. The incubation medium contained 50 mM Tris-HCl, pH 7.2, 2.5 mM MgSO4 and 200 mM NH4Cl and 2.5 mM polyP (as phosphate) with the average chain lengths of 208 phosphate residues (polyP208) (Monsanto, Creve Coeur, MO, USA). In case of exopolyphosphatase activity, the amount of the enzyme forming 1 µmole P in 1 min was taken as a unit of activity (U).
The endopolyphosphatase activity was analyzed by a decrease in polyP208 chain length. The incubation medium contained 50 mM Tris-HCl, pH 7.2, 200 mM NH4Cl, 1 mM MgSO4, 9 mM polyP208 and cell-free extract or crude membrane fraction samples. Incubation time was 30 min at 30°. The reaction was stopped by adding HClO4 to a concentration of 0.5 M. Further sample preparation and electrophoresis in 30% PAAG with 7 M urea were performed as described [28].
BSA was used as a standard for protein assay with Bradford reagent (Thermo Fisher, Waltham, MA, USA) according to the instructions of the company.
2.4. Determination of Yeast Sensitivity to Peroxide, Alkali, and Heavy Metal Ions
Yeast samples normalized by cell concentration (0.5 × 107 cell/mL) were added to the wells of sterile plates containing the YPD medium and different concentrations of Cd(CH3COO)2×2H2O, MnSO4, hydrogen peroxide, or KOH, cultured for 24 h, and optical density was measured at 594 nm with an EFOS photometer (Sapphire, Russia).
2.5. Statistics
Experiments were repeated four times, and results are presented as the mean with standard deviation (Excel). The experiment with electrophoresis was repeated twice.
3. Results
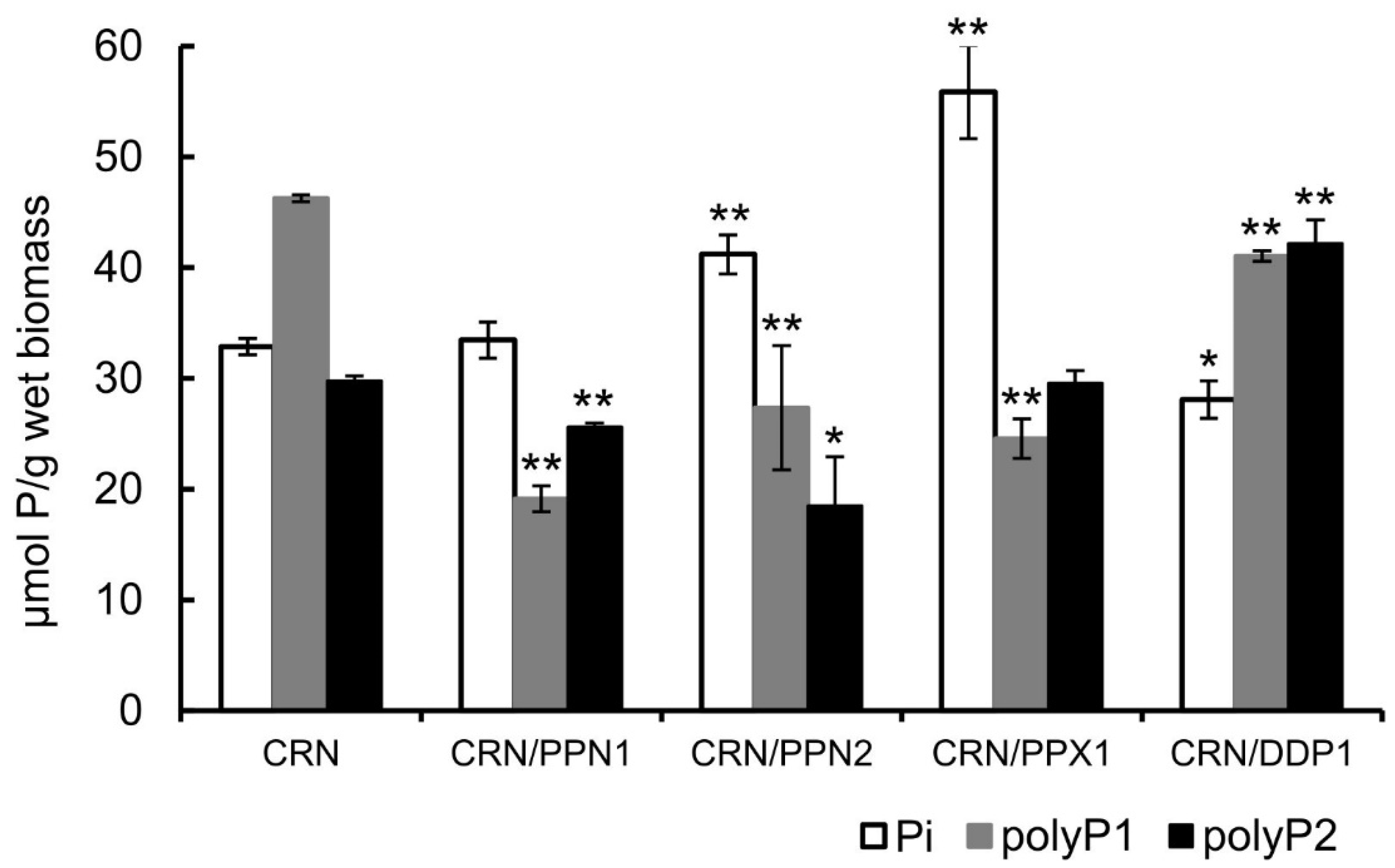
In this study, we cultivated the parent strain and polyphosphatase-overexpressing strains in the same YPD medium containing 3 mM phosphate (Pi). There were no differences in the growth of the strains in this medium (data not shown). Hence, it was possible to correctly compare the polyP content in the cells of all strains under study. Pi level was higher in the cells of the CRN/PPX1 and CRN/PPN2 strains (Figure 2). The content of acid-soluble polyP was lower in cells overexpressing Ppx1, Ppn1, and Ppn2 polyphosphatases, while the content of acid-insoluble polyP was lower only in cells overexpressing Ppn2 polyphosphatase (Figure 2).
Yeast polyphosphatases differ in subcellular localization. Ppx1 [27] and Ddp1 [29] are mainly localized in the cytoplasm [27], while Ppn1 and Ppn2 are mainly localized in vacuoles [28,30]. Ppn2 and Ppn1 are able to form a complex in the vacuolar membrane [30]. In the case of overexpressing strains, all four polyphosphatases were observed in cell-free extracts [31]. So, we proved the exopolyphosphatase and endopolyphosphatase activities in cell-free extracts and crude membrane fractions of all strains under study (Table 1 and Figure 3). As expected, exopolyphosphatase activities showed the greatest increase in strains overexpressing Ppx1 and Ppn1. In the case of the cell-free extract obtained from the cells of the CRN/PPX1 strain, there was complete hydrolysis of polyP (Figure 3).
Endopolyphosphatase activity was estimated by the decrease in the chain length of polyP208 used as substrate. The content of cellular polyPs in samples used in this experiment was undetectable on electrophoregram (not shown). There was a strong increase in endopolyphosphatase activity, both in the cell-free extract and the crude membrane fraction obtained from the CRN/PPN1 and CRN/PPN2 strains. As regards the CRN/DDP1 strain, the increase in exo- and endopolyphosphatase activities was slight and occurred in the cell-free extract (Figure 3). The decrease in polyP content is in agreement with the increase in polyphosphatase activities in the cells of polyphosphatase-overexpressing strains.
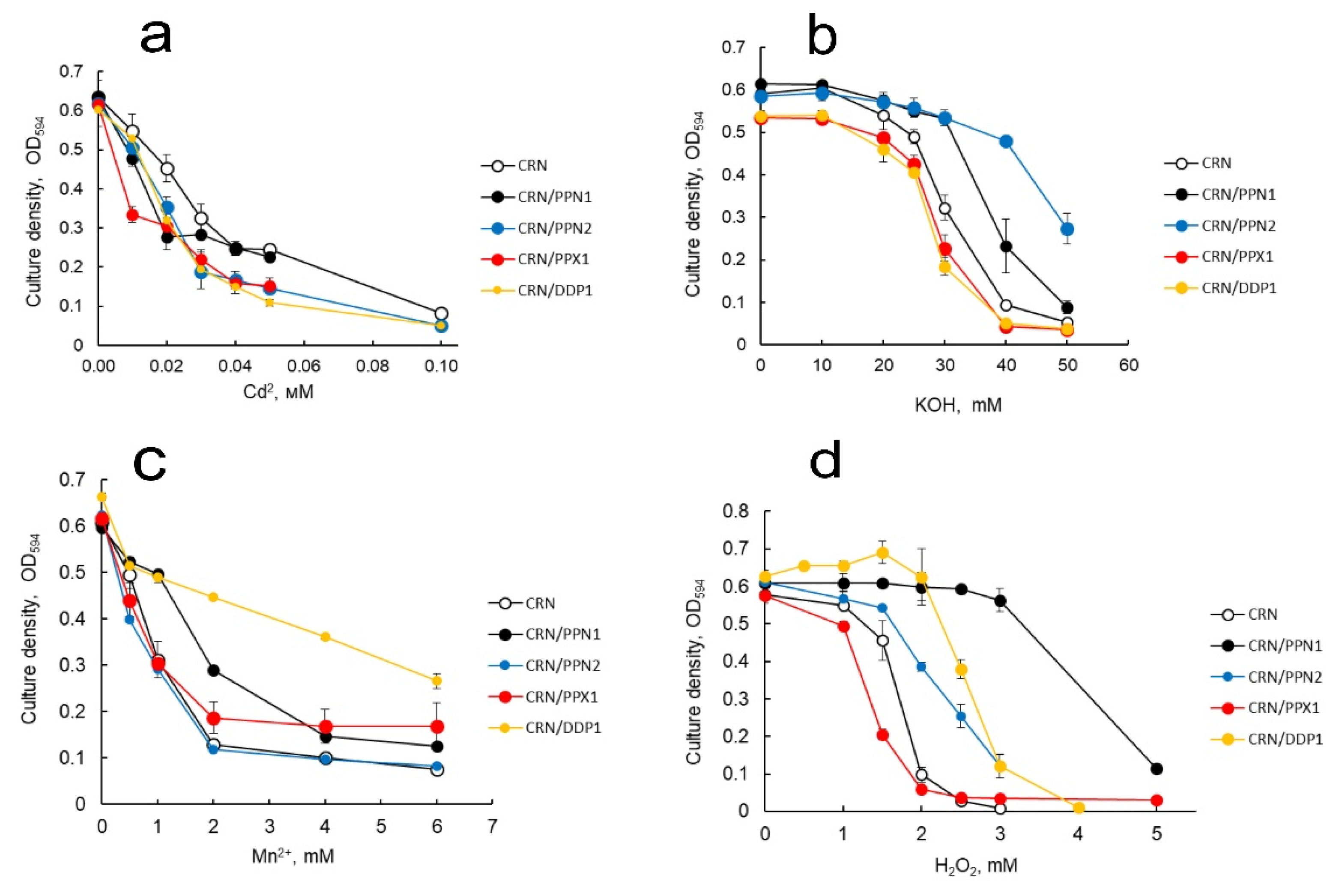
Thus, there were three strains with lower polyP content (CRN/PPX1, CRN/PPN1, and CRN/PPN2) and one strain with unchanged polyP content (CRN/DDP1). Strains were compared with respect to their resistance to alkali, heavy metals, and peroxide. Resistance to Cd2+ was almost the same for all tested overexpressing strains, and lower than this value for parent strain CRN (Figure 4a).
The I50 for Cd2+ was near 0.04 mM and 0.02 mM for the parent strain CRN and overexpressing strains, respectively. The Ppn2-overexpressing strain was more resistant to alkali; the Ppn1 strain also showed higher resistance to alkali (Figure 4b). The resistance of Ddp1 and Ppx1 to alkali was similar to that of the parent strain. The strains overexpressing Ppn1 and Ddp1 were more resistant to peroxide and Mn2+ among the strains tested (Figure 4c,d). The Ppn2-overexpressing strain shows an increase in resistance to peroxide but not to manganese (Figure 4c,d).
4. Discussion
The relationship between polyP content and changes in stress resistance in the cells of S. cerevisiae was nondirect. For example, CRN/PPN1 and CRN/DDP1 strains displayed enhanced resistance to peroxide and manganese, though the content of polyP in the CRN/PPN1 strain was reduced; in CRN/DDP1, it was similar to parent strain CRN. The peroxide and manganese resistance of the CRN/PPN1 strain was associated with the activation of the expression of large groups of genes involved in general stress response: the genes associated with the response to external stimulus, plasma membrane organization, and oxidation/reduction [21]. The question of compounds triggering these expression changes is still under discussion. The comparison of polyP content and the oxidative and manganese stress resistance in the CRN/PPN1 and CRN/DDP1 strains allows for us to speculate that inositol pyrophosphates may serve as a signal to preadaptive state of the cells of above strains. The role of inositol pyrophosphates in the stress response of yeast cells is highly significant and implemented through transcription factors [36]. Ddp1 is a diphosphoinositol-polyphosphate phosphohydrolase [29], and the overexpression of this enzyme must lead to a decrease in the level of diphosphoinositol polyphosphate. As regards Ppn1, there are no published data on their ability to hydrolyze diphosphoinositol polyphosphate. It is difficult to obtain data on the ability of enzymes to hydrolyze inositol pyrophosphates because these compounds are very labile and not available as commercial products. Methods for the analysis of these compounds in cells are relatively recent and require special equipment [37]. The findings of the present work draw attention to the participation of diphosphoinositol polyphosphate in stress response, and the necessity for the further analysis of inter-relationships between polyphosphates and these signaling molecules. These inter-relationships are quite complex and variable among microorganism species [38,39].
There are no available data on the abilities Ppn2 to hydrolyze signal compounds. So, we explain the increased resistance of Ppn2 overexpressing strain to alkali and peroxide by the fact that the cells of this strain are able to faster hydrolyze polyP, both for buffering the cytoplasm and for the formation of an antioxidant complex of phosphate and manganese.
The CRN/PPX1 strain showed no increase in resistance and even a decrease in peroxide resistance, though the content of polyP in this strain under the cultivation conditions used in this work was lower, as in the CRN/PPN1 strain. Ppx1 did not hydrolyze inositol pyrophosphates [29], and there were consequently no grounds to expect a decrease in diphosphoinositol polyphosphate level in the Ppx1-overexpressing strain. Instead of this, Ppx1 could hydrolyze cAMP [31]. Cyclic AMP signaling pathways are involved in the protection against oxidative stress [40]. A more than 20 times increase in Ppx1 activity in the cells of CRN/PPX1 strain (Table 1) could probably influence the process of overcoming oxidative stress.
Stress-tolerant yeast strains were considered to be promising models for the development of new bioremediation technologies [41,42,43]. Strains overexpressing Ppn1 and Ddp1 polyphosphatases with increased resistance to oxidative stress and heavy metal stress are of interest for future research in this area.
5. Conclusions
In this study, we demonstrated that the overexpression of yeast polyphosphatases in the cells of S. cerevisiae alters their stress resistance, independently of changes in polyP content. Strains overexpressing Ppx1, Ppn1, and Ppn2 had lower polyP content, and the strain overexpressing Ddp1 had the same polyP content as that of the parent strain CRN. Strains overexpressing Ppn1 and Ddp1 showed higher resistance to peroxide and manganese. The strain overexpressing Ppx1 showed a decrease in peroxide resistance. The strain overexpressing Ppn2 was more resistant to alkaline and peroxide stresses. The difference in the stress resistance of strains overexpressing polyphosphatases may be due to the activities of some polyphosphatases to hydrolyze some signal compounds. For example, a similar increase in resistance to manganese and peroxide stresses of strains overexpressing polyphosphatase Ppn1 and inositol pyrophosphate hydrolase Ddp1 suggests the possible role of inositol pyrophosphates in observed effects.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/article/10.3390/stresses2010002/s1. Figure S1: A pixel densitometry graph of electroforegram shown in Figure 3 obtained by Image Engineering—ImageJ tool (https://imagej.nih.gov/nih-image/moredocs/Engineering/ImgEngr.html, accessed on 22 December 2021); Table S1: The construction of expression vectors and the genetic map of plasmids pMB1 were published earlier [33]; Table S2: Statistical significance estimates for data shown in Figure 4. Two-tailed Student’s t-test was computed in MS Excel software comparing the data from each polyphosphataseoverexpressing strain against CRN.
Author Contributions
Conceptualization, T.K.; methodology, T.K. and N.A.; investigation, N.A., L.R., L.L. and L.T.; writing, T.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rao, N.N.; Gómez-García, M.R.; Kornberg, A. Inorganic Polyphosphate: Essential for Growth and Survival. Annu. Rev. Biochem. 2009, 78, 605–647. [Google Scholar] [CrossRef] [PubMed]
- Albi, T.; Serrano, A. Inorganic polyphosphate in the microbial world. Emerging roles for a multifaceted biopolymer. World J. Microbiol. Biotechnol. 2016, 32, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Jakob, U. Inorganic Polyphosphate, a Multifunctional Polyanionic Protein Scaffold. J. Biol. Chem. 2019, 294, 2180–2190. [Google Scholar] [CrossRef] [Green Version]
- Desfougères, Y.; Saiardi, A.; Azevedo, C. Inorganic Polyphosphate in Mammals: Where’s Wally? Biochem. Soc. Trans. 2020, 48, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Rosigkeit, H.; Kneißle, L.; Obruča, S.; Jendrossek, D. The Multiple Roles of Polyphosphate in Ralstonia eutropha and Other Bacteria. Microb. Physiol. 2021, 31, 63–177. [Google Scholar] [CrossRef] [PubMed]
- Denoncourt, A.; Downey, M. Model Systems for Studying Polyphosphate Biology: A Focus on Microorganisms. Curr. Genet. 2021, 67, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Ropelewska, M.; Gross, M.H.; Konieczny, I. DNA and Polyphosphate in Directed Proteolysis for DNA Replication Control. Front. Microbiol. 2020, 11, 585717. [Google Scholar] [CrossRef]
- Sultana, S.; Foti, A.; Dahl, J.U. Bacterial Defense Systems against the Neutrophilic Oxidant Hypochlorous Acid. Infect. Immun. 2020, 88, e00964-19. [Google Scholar] [CrossRef] [PubMed]
- Gross, M.H.; Konieczny, I. Polyphosphate Induces the Proteolysis of ADP-bound Fraction of Initiator to Inhibit DNA Replication Initiation upon Stress in Escherichia coli. Nucleic Acids Res. 2020, 48, 5457–5466. [Google Scholar] [CrossRef]
- Beaufay, F.; Quarles, E.; Franz, A.; Katamanin, O.; Wholey, W.Y.; Jakob, U. Polyphosphate Functions In Vivo as an Iron Chelator and Fenton Reaction Inhibitor. mBio 2020, 11, e01017-20. [Google Scholar] [CrossRef]
- Sanz-Luque, E.; Bhaya, D.; Grossman, A.R. Polyphosphate: A Multifunctional Metabolite in Cyanobacteria and Algae. Front. Plant Sci. 2020, 11, 938. [Google Scholar] [CrossRef] [PubMed]
- Kulakovskaya, T. Inorganic Polyphosphates and Heavy Metal Resistance in Microorganisms. World J. Microbiol. Biotechnol. 2018, 34, 139. [Google Scholar] [CrossRef] [PubMed]
- Rivero, M.; Torres-Paris, C.; Muñoz, R.; Cabrera, R.; Navarro, C.A.; Jerez, C.A. Inorganic Polyphosphate, Exopolyphosphatase, and Pho84-like Transporters May Be Involved in Copper Resistance in Metallosphaera sedula DSM 5348T. Archaea 2018, 2018, 5251061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villagrasa, E.; Egea, R.; Ferrer-Miralles, N.; Solé, A. Genomic and Biotechnological Insights on Stress-linked Polyphosphate Production Induced by Chromium(III) in Ochrobactrum anthropi DE2010. World J. Microbiol. Biotechnol. 2020, 36, 97. [Google Scholar] [CrossRef] [PubMed]
- Reddi, A.R.; Jensen, L.T.; Culotta, V.C. Manganese homeostasis in Saccharomyces cerevisiae. Chem. Rev. 2009, 109, 4722–4732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Xie, Z.; Wang, B.; Yu, N.; Zhao, J.; Zhou, Y.; Hua, Y.; Tian, B. Dynamic Polyphosphate Metabolism Coordinating with Manganese Ions Defends against Oxidative Stress in the Extreme Bacterium Deinococcus radiodurans. Appl. Environ. Microbiol. 2021, 87, e02785-20. [Google Scholar] [CrossRef]
- Vagabov, V.M.; Trilisenko, L.V.; Kulakovskaya, T.V.; Kulaev, I.S. Effect of a Carbon Source on Polyphosphate Accumulation in Saccharomyces Cerevisiae. FEMS Yeast Res. 2008, 8, 877–882. [Google Scholar] [CrossRef] [Green Version]
- Christ, J.J.; Blank, L.M. Saccharomyces cerevisiae Containing 28% Polyphosphate and Production of a Polyphosphate-Rich Yeast Extract Thereof. FEMS Yeast Res. 2019, 19, foz011. [Google Scholar] [CrossRef]
- Bru, S.; Martínez-Laínez, J.M.; Hernández-Ortega, S.; Quandt, E.; Torres-Torronteras, J.; Martí, R.; Canadell, D.; Ariño, J.; Sharma, S.; Jiménez, J.; et al. Polyphosphate Is Involved in Cell Cycle Progression and Genomic Stability in Saccharomyces cerevisiae. Mol. Microbiol. 2016, 101, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Serra-Cardona, A.; Canadell, D.; Ariño, J. Coordinate Responses to Alkaline pH Stress in Budding Yeast. Microb. Cell 2015, 2, 182–196. [Google Scholar] [CrossRef] [Green Version]
- Trilisenko, L.; Zvonarev, A.; Valiakhmetov, A.; Penin, A.A.; Eliseeva, I.A.; Ostroumov, V.; Kulakovskiy, I.V.; Kulakovskaya, T. The Reduced Level of Inorganic Polyphosphate Mobilizes Antioxidant and Manganese-Resistance Systems in Saccharomyces cerevisiae. Cells 2019, 8, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeh, M.; Ahmed, Y.; Quinn, J. Phosphate Acquisition and Virulence in Human Fungal Pathogens. Microorganisms 2017, 5, 48. [Google Scholar] [CrossRef]
- Köhler, J.R.; Acosta-Zaldívar, M.; Qi, W. Phosphate in Virulence of Candida albicans and Candida glabrata. J. Fungi 2020, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreeva, N.; Ryazanova, L.; Dmitriev, V.; Kulakovskaya, T.; Kulaev, I. Adaptation of Saccharomyces cerevisiae to Toxic Manganese Concentration Triggers Changes in Inorganic Polyphosphates. FEMS Yeast Res. 2013, 13, 463–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreeva, N.; Ryazanova, L.; Dmitriev, V.; Kulakovskaya, T.; Kulaev, I. Cytoplasmic inorganic polyphosphate participates in the heavy metal tolerance of Cryptococcus humicola. Folia Microbiol. 2014, 59, 381–3899. [Google Scholar] [CrossRef] [PubMed]
- Hovnanyan, K.; Marutyan, S.; Marutyan, S.; Hovnanyan, M.; Navasardyan, L.; Trchounian, A. Ultrastructural Investigation of Acidocalcisomes and ATPase Activity in Yeast Candida Guilliermondii NP-4 as ‘Complementary’ Stress-Targets. Lett. Appl. Microbiol. 2020, 71, 413–419. [Google Scholar] [CrossRef]
- Wurst, H.; Shiba, T.; Kornberg, A. The Gene for a Major Exopolyphosphatase of Saccharomyces cerevisiae. J. Bacteriol. 1995, 177, 898–906. [Google Scholar] [CrossRef] [Green Version]
- Andreeva, N.; Trilisenko, L.; Eldarov, M.; Kulakovskaya, T. Polyphosphatase PPN1 of Saccharomyces cerevisiae: Switching of Exopolyphosphatase and Endopolyphosphatase Activities. PLoS ONE 2015, 10, e0119594. [Google Scholar] [CrossRef]
- Lonetti, A.; Szijgyarto, Z.; Bosch, D.; Loss, O.; Azevedo, C.; Saiardi, A. Identification of an Evolutionarily Conserved Family of Inorganic Polyphosphate Endopolyphosphatases. J. Biol. Chem. 2011, 286, 31966–31974. [Google Scholar] [CrossRef] [Green Version]
- Gerasimait, R.; Mayer, A. Ppn2, a Novel Zn2+-Dependent Polyphosphatase in the Acidocalcisome-like Yeast Vacuole. J. Cell Sci. 2017, 130, 1625–1636. [Google Scholar] [CrossRef] [Green Version]
- Andreeva, N.; Ledova, L.; Ryazanova, L.; Tomashevsky, A.; Kulakovskaya, T.; Eldarov, M. Ppn2 Endopolyphosphatase Overexpressed in Saccharomyces cerevisiae: Comparison with Ppn1, Ppx1, and Ddp1 Polyphosphatases. Biochimie 2019, 163, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Ryazanova, L.P.; Ledova, L.A.; Andreeva, N.A.; Zvonarev, A.N.; Eldarov, M.A.; Kulakovskaya, T.V. Inorganic Polyphosphate and Physiological Properties of Saccharomyces cerevisiae Yeast Overexpressing Ppn2. Biochemistry 2020, 85, 516–522. [Google Scholar] [CrossRef]
- Eldarov, M.A.; Baranov, M.V.; Dumina, M.V.; Shgun, A.A.; Andreeva, N.A.; Trilisenko, L.V.; Kulakovskaya, T.V.; Ryasanova, L.P.; Kulaev, I.S. Polyphosphates and Exopolyphosphatase Activities in the Yeast Saccharomyces cerevisiae under Overexpression of Homologous and Heterologous PPN1 Genes. Biochemistry 2013, 78, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Sethuraman, A.; Rao, N.N.; Kornberg, A. The Endopolyphosphatase Gene: Essential in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2001, 98, 8542–8547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinonen, J.K.; Lahti, R.J. A New and Convenient Colorimetric Determination of Inorganic Orthophosphate and its Application to the Assay of Inorganic Pyrophosphatase. Anal. Biochem. 1981, 113, 313–317. [Google Scholar] [CrossRef]
- Morrissette, V.A.; Rolfes, R.J. The Intersection Between Stress Responses and Inositol Pyrophosphates in Saccharomyces cerevisiae. Curr. Genet. 2020, 66, 901–910. [Google Scholar] [CrossRef]
- Qiu, D.; Eisenbeis, V.B.; Saiardi, A.; Jessenm, H.J. Absolute Quantitation of Inositol Pyrophosphates by Capillary Electrophoresis Electrospray Ionization Mass Spectrometry. J. Vis. Exp. 2021, 174, e62847. [Google Scholar] [CrossRef]
- Pascual-Ortiz, M.; Walla, E.; Fleig, U.; Saiardi, A. The PPIP5K Family Member Asp1 Controls Inorganic Polyphosphate Metabolism in S. pombe. J. Fungi 2021, 37, 626. [Google Scholar] [CrossRef]
- Desfougères, Y.; Portela-Torres, P.; Qiu, D.; Livermore, T.M.; Harmel, R.K.; Borghi, F.; Jessen, H.J.; Fiedler, D.; Saiardi, A. The Inositol Pyrophosphate Metabolism of Dictyostelium discoideum does not Regulate Inorganic Polyphosphate (polyP) Synthesis. Adv. Biol. Regul. 2021, 10, 100835. [Google Scholar] [CrossRef]
- De la Torre-Ruiz, M.A.; Mozo-Villarías, A.; Pujol, N.; Petkova, M.I. How Budding Yeast Sense and Transduce the Oxidative Stress Signal and the Impact in Cell Growth and Morphogenesis. Curr. Protein Pept. Sci. 2010, 11, 669–679. [Google Scholar] [CrossRef]
- Li, C.; Yu, J.; Wang, D.; Li, L.; Yang, X.; Ma, H.; Xu, Y. Efficient Removal of Zinc by Multi-stress-tolerant Yeast Pichia kudriavzevii A16. Bioresour. Technol. 2016, 206, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Feng, J.; Dai, Y.; Chang, S. Biosorption of Strontium Ions from Simulated High-level Liquid Waste by Living Saccharomyces cerevisiae. Environ. Sci. Pollut. Res. 2018, 25, 17194–17206. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.L.; Reynolds, E.E.; Belcher, A.M. Designing Yeast as Plant-like Hyperaccumulators for Heavy Metals. Nat. Commun. 2019, 10, 5080. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the action of polyphosphatases on polyP. Exopolyphosphatase Ppx1 cleave Pi from the end of the polyP chain. Endopolyphosphatases Ppn2 and Ddp1 split long-chained polyP into shorter ones. Polyphosphatase Ppn1 displays both exo- and endopolyphosphatase activities.
Figure 1.
Schematic representation of the action of polyphosphatases on polyP. Exopolyphosphatase Ppx1 cleave Pi from the end of the polyP chain. Endopolyphosphatases Ppn2 and Ddp1 split long-chained polyP into shorter ones. Polyphosphatase Ppn1 displays both exo- and endopolyphosphatase activities.

Figure 2.
Pi (orthophosphate) and polyP content in cells of S. cerevisiae strains overexpressing polyphosphatases Ppx1, Ppn1, Ppn2, or Ddp1 and parent strain CRN. Cells were cultivated in YPD medium (2% glucose, 2% peptone, 1% yeast extract, w/v) containing 3 mM Pi for 24 h at 29 °C with shaking at 145 rpm. Acid-soluble polyP1 extracted from biomass with 0.5 M HClO4 at 0 °C. Amount of acid-insoluble polyP2 remaining in the biomass was estimated by the phosphate released after 20 min treatment of biomass samples with 0.5 M HClO4 at 100 °C. Amount of polyP normalized to weight of biomass samples standardized by centrifugation at 5000× g for 15 min (wet biomass). Statistical significance assessed against CRN with Student’s t-test: * p < 0.05, ** p < 0.01.
Figure 2.
Pi (orthophosphate) and polyP content in cells of S. cerevisiae strains overexpressing polyphosphatases Ppx1, Ppn1, Ppn2, or Ddp1 and parent strain CRN. Cells were cultivated in YPD medium (2% glucose, 2% peptone, 1% yeast extract, w/v) containing 3 mM Pi for 24 h at 29 °C with shaking at 145 rpm. Acid-soluble polyP1 extracted from biomass with 0.5 M HClO4 at 0 °C. Amount of acid-insoluble polyP2 remaining in the biomass was estimated by the phosphate released after 20 min treatment of biomass samples with 0.5 M HClO4 at 100 °C. Amount of polyP normalized to weight of biomass samples standardized by centrifugation at 5000× g for 15 min (wet biomass). Statistical significance assessed against CRN with Student’s t-test: * p < 0.05, ** p < 0.01.

Figure 3.
Decrease in chain length of polyP208 (commercial preparation of polyP with average chain length of 208 phosphate residues) after incubation with cell-free extract or crude membrane fraction samples obtained from cells of S. cerevisiae strains overexpressing polyphosphatases Ppx1, Ppn1, Ppn2, or Ddp1, and parent strain CRN. Incubation medium contained 50 mM Tris-HCl, pH 7.2, 200 mM NH4Cl, 1 mM MgSO4, 9 mM polyP208 and cell-free extract or crude membrane fraction samples. Amounts of total protein added to reaction mixture were 157, 90, 60, 59, and 63 μg for cell-free extracts preparations from cells of CRN, CRN/PPX1, CRN/PPN1, CRN/PPN1, and CRN/DDP1 strains, respectively. Amounts of total protein added to reaction mixture were 61, 55, 14, 16, and 60 μg for crude membrane preparations from cells of CRN, CRN/PPX1, CRN/PPN1, CRN/PPN1, and CRN/DDP1 strains, respectively. C-control, polyP208 was incubated without polyphosphatase-containing samples. PolyP markers were from Sigma-Aldrich (polyP75, polyP25, and polyP15 were polyP with average chain length of 75, 25 and 15 phosphate residues, respectively. Electrophoresis was performed in 30% PAAG with 7 M urea, staining with toluidine blue. A pixel densitometry graph of electrophoregrams is shown in Supplementary Figure S1.
Figure 3.
Decrease in chain length of polyP208 (commercial preparation of polyP with average chain length of 208 phosphate residues) after incubation with cell-free extract or crude membrane fraction samples obtained from cells of S. cerevisiae strains overexpressing polyphosphatases Ppx1, Ppn1, Ppn2, or Ddp1, and parent strain CRN. Incubation medium contained 50 mM Tris-HCl, pH 7.2, 200 mM NH4Cl, 1 mM MgSO4, 9 mM polyP208 and cell-free extract or crude membrane fraction samples. Amounts of total protein added to reaction mixture were 157, 90, 60, 59, and 63 μg for cell-free extracts preparations from cells of CRN, CRN/PPX1, CRN/PPN1, CRN/PPN1, and CRN/DDP1 strains, respectively. Amounts of total protein added to reaction mixture were 61, 55, 14, 16, and 60 μg for crude membrane preparations from cells of CRN, CRN/PPX1, CRN/PPN1, CRN/PPN1, and CRN/DDP1 strains, respectively. C-control, polyP208 was incubated without polyphosphatase-containing samples. PolyP markers were from Sigma-Aldrich (polyP75, polyP25, and polyP15 were polyP with average chain length of 75, 25 and 15 phosphate residues, respectively. Electrophoresis was performed in 30% PAAG with 7 M urea, staining with toluidine blue. A pixel densitometry graph of electrophoregrams is shown in Supplementary Figure S1.

Figure 4.
Effects of (a) Cd2+, (b) alkali, (c) Mn2+, and (d) hydrogen peroxide on growth of S. cerevisiae strains overexpressing polyphosphatases Ppx1, Ppn1, Ppn2, or Ddp1 and parent strain CRN. Statistical significance estimates shown in Supplementary Table S2. Two-tailed Student’s t-test was computed in MS Excel software comparing data from each polyphosphatase-overexpressing strain against CRN strain.
Figure 4.
Effects of (a) Cd2+, (b) alkali, (c) Mn2+, and (d) hydrogen peroxide on growth of S. cerevisiae strains overexpressing polyphosphatases Ppx1, Ppn1, Ppn2, or Ddp1 and parent strain CRN. Statistical significance estimates shown in Supplementary Table S2. Two-tailed Student’s t-test was computed in MS Excel software comparing data from each polyphosphatase-overexpressing strain against CRN strain.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Exopolyphosphatase activities (U/mg protein) in cell-free extracts and crude membrane fractions of S. cerevisiae CRN parent strain and strains overexpressing polyphosphatases.
Table 1.
Exopolyphosphatase activities (U/mg protein) in cell-free extracts and crude membrane fractions of S. cerevisiae CRN parent strain and strains overexpressing polyphosphatases.
| Strain | Cell-Free Extract | Crude Membrane Fraction |
|---|---|---|
| CRN | 0.342 ± 0.033 | 0.094 ± 0.005 |
| CRN/PPX1 | 8.04 ± 0.44 | 1.72 ± 0.04 |
| CRN/PPN1 | 1.64 ± 0.123 | 4.6 ± 0.18 |
| CRN/PPN2 | 0.874 ± 0.44 | 0.39 ± 0.023 |
| CRN/DDP1 | 0.22 ± 0.0136 | 0.099 ± 0.0072 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Andreeva, N.; Ryazanova, L.; Ledova, L.; Trilisenko, L.; Kulakovskaya, T. Stress Resistance of Saccharomyces cerevisiae Strains Overexpressing Yeast Polyphosphatases. Stresses 2022, 2, 17-25. https://doi.org/10.3390/stresses2010002
AMA Style
Andreeva N, Ryazanova L, Ledova L, Trilisenko L, Kulakovskaya T. Stress Resistance of Saccharomyces cerevisiae Strains Overexpressing Yeast Polyphosphatases. Stresses. 2022; 2(1):17-25. https://doi.org/10.3390/stresses2010002
Chicago/Turabian StyleAndreeva, Nadeshda, Lubov Ryazanova, Larisa Ledova, Ludmila Trilisenko, and Tatiana Kulakovskaya. 2022. "Stress Resistance of Saccharomyces cerevisiae Strains Overexpressing Yeast Polyphosphatases" Stresses 2, no. 1: 17-25. https://doi.org/10.3390/stresses2010002