
Teamwork Makes the String Work: A Pilot Test of the Loose String Task with African Crested Porcupines (Hystrix cristata)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
2.3. Procedure
2.3.1. Phase 1: Individual Pulling
2.3.2. Phase 2: Simultaneous Pulling
2.3.3. Video Coding
3. Results
3.1. Reliability
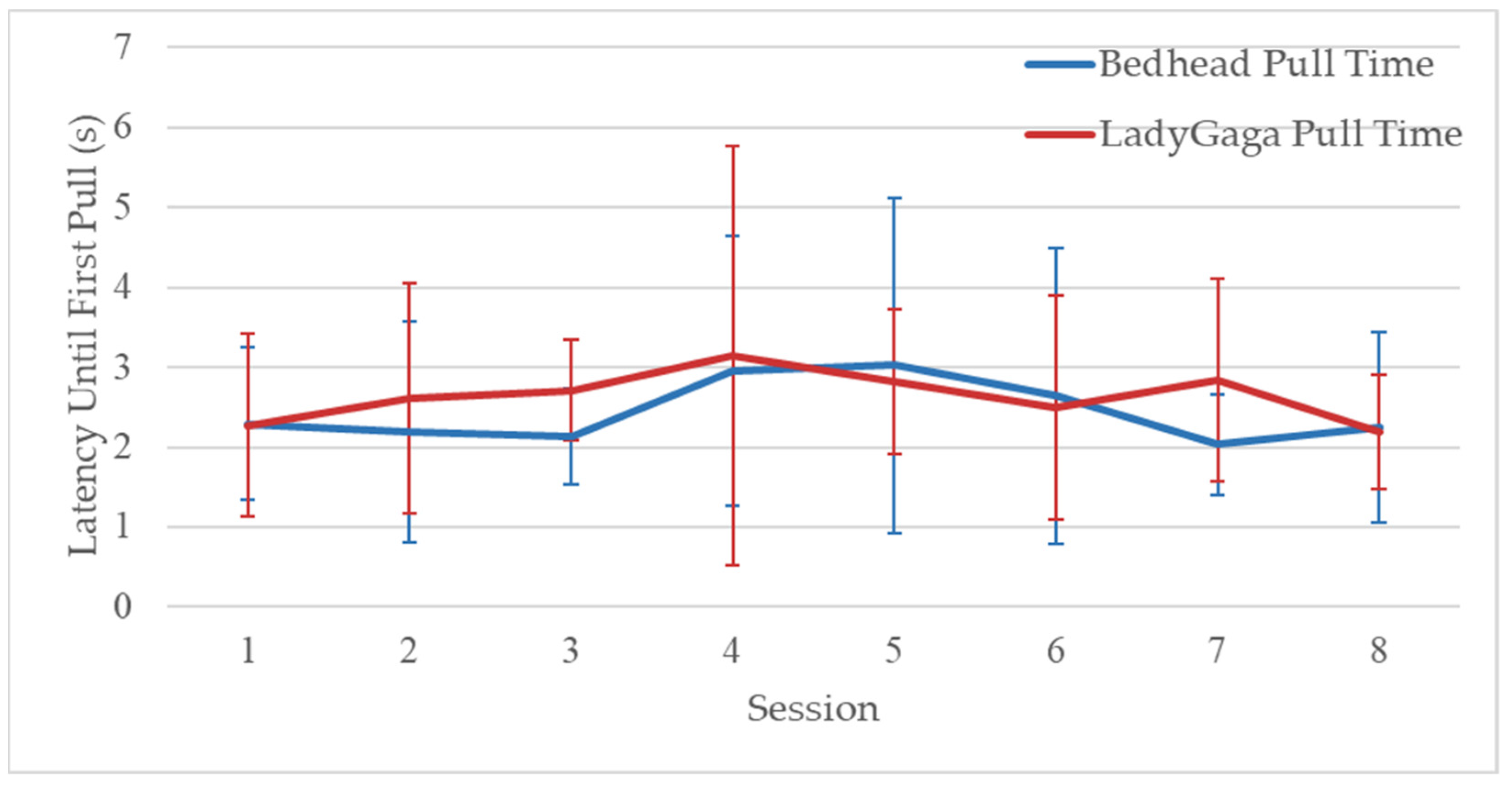
3.2. Phase 1: Individual Pulling
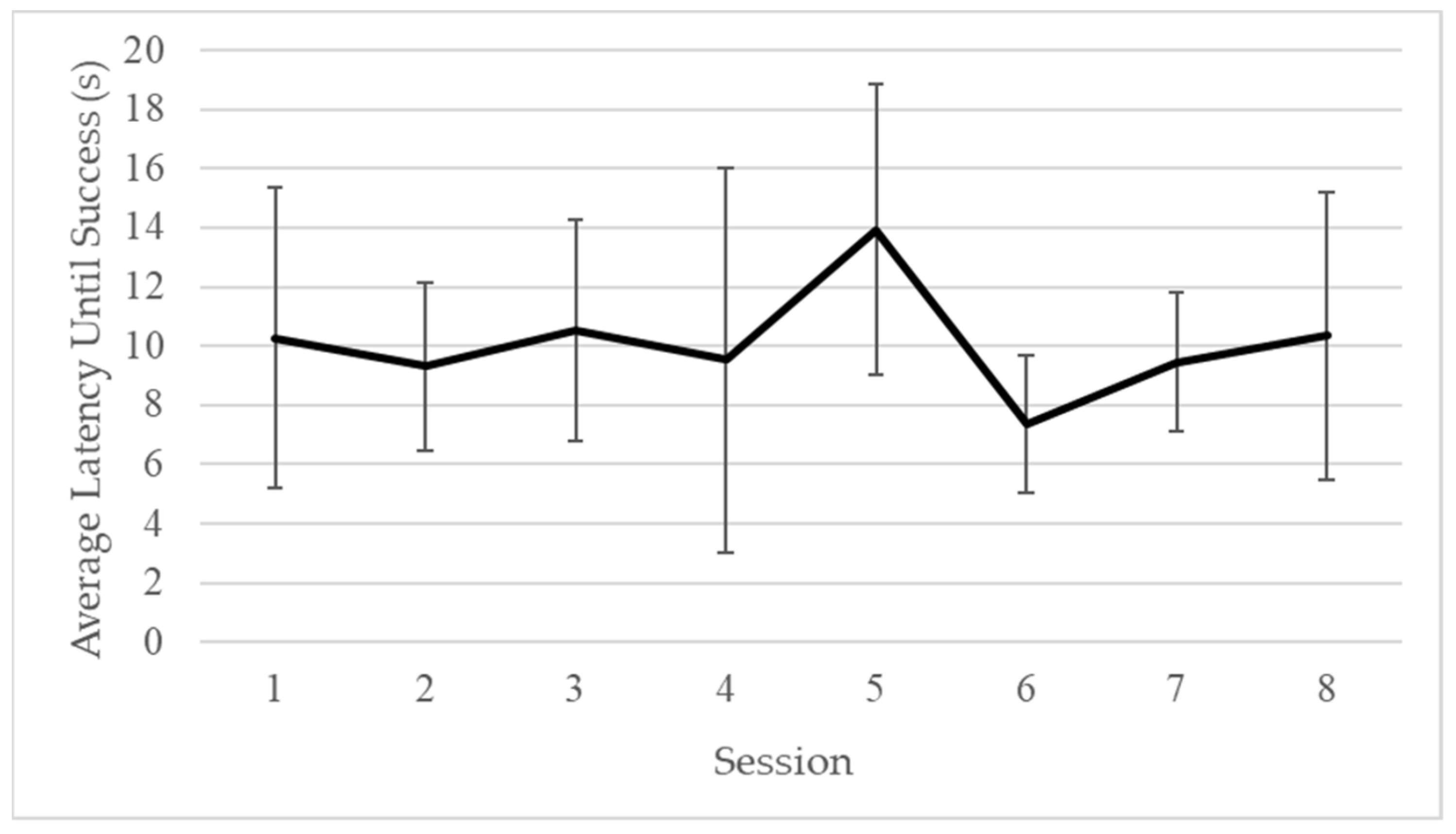
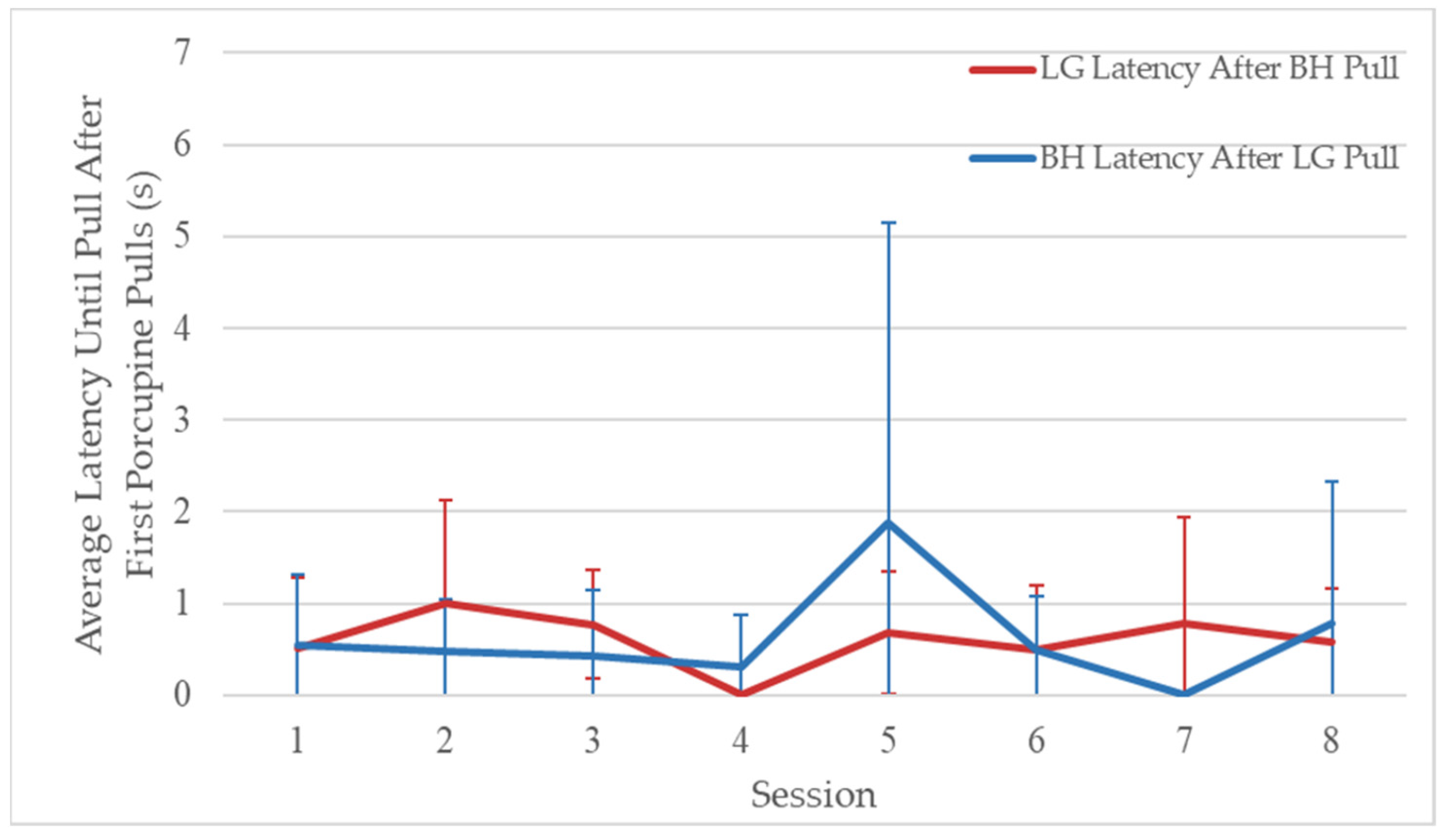
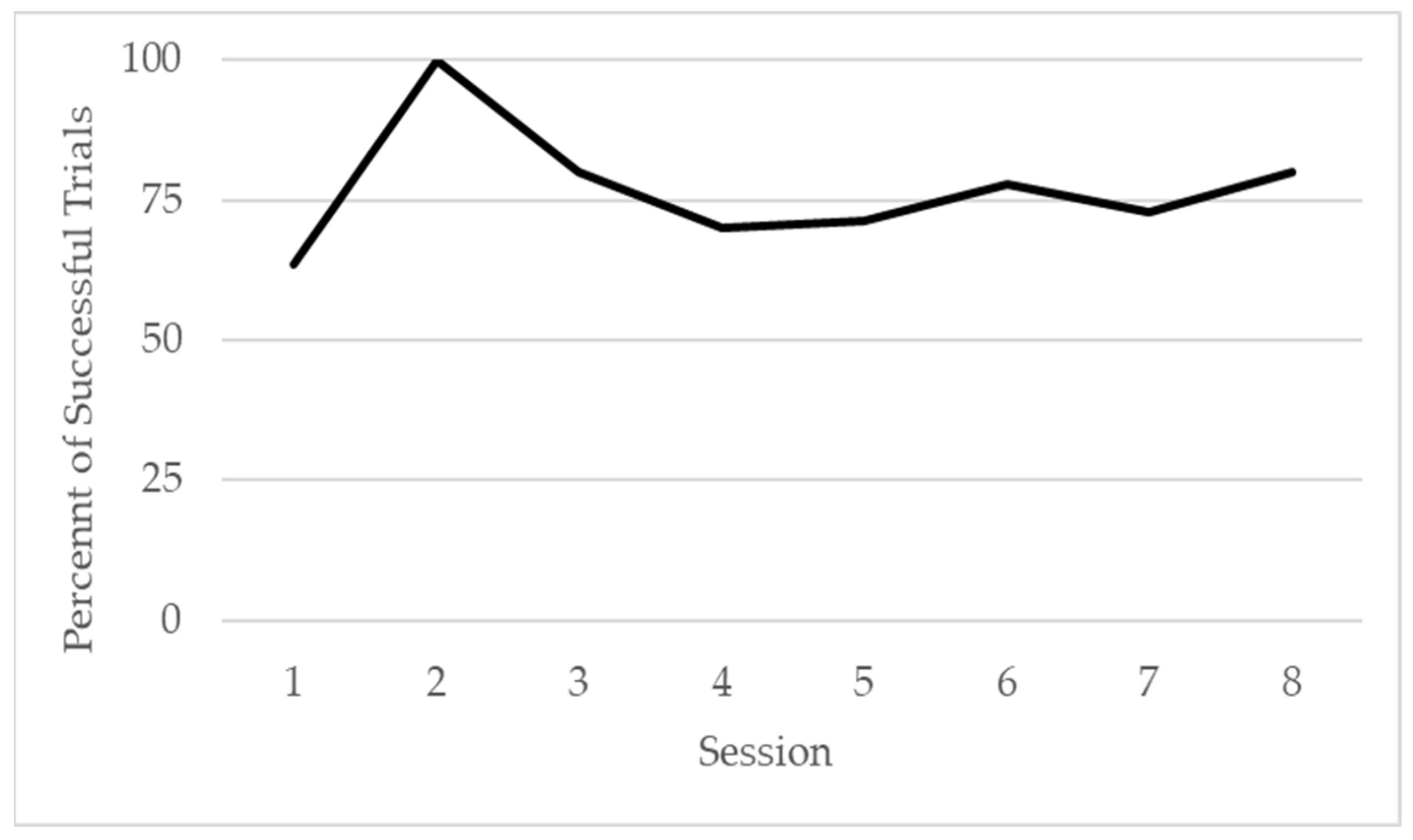
3.3. Phase 2: Simultaneous Pulling
4. Discussion
Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Creel, S.; Creel, N.M. Communal hunting and pack size in African wild dogs, Lycaon pictus. Anim. Behav. 1995, 50, 1325–1339. [Google Scholar] [CrossRef]
- Bossema, I.; Benus, R.F. Territorial defence and intra-pair cooperation in the carrion crow (Corvus corone). Behav. Ecol. Sociobiol. 1985, 16, 99–104. [Google Scholar] [CrossRef]
- Boesch, C.; Boesch, H. Hunting behavior of wild chimpanzees in the Tai National Park. Am. J. Phys. Anthropol. 1989, 78, 547–573. [Google Scholar] [CrossRef] [PubMed]
- Schoech, S.J. Physiology of helping in Florida scrub-Jays: When these birds are young, they delay reproduction and help others raise their offspring. The hormone prolactin may influence that cooperation. Am. Sci. 1998, 86, 70–77. [Google Scholar] [CrossRef]
- Stander, P.E. Cooperative hunting in lions: The role of the individual. Behav. Ecol. Sociobiol. 1992, 29, 445–454. [Google Scholar] [CrossRef]
- Bronstein, J.L. The scope for exploitation within mutualistic interactions. In Genetic and Cultural Evolution of Cooperation; Hammerstein, P., Ed.; MIT Press: London, UK, 2003; pp. 185–202. [Google Scholar]
- Noë, R. Cooperation experiments: Coordination through communication versus acting apart together. Anim. Behav. 2006, 71, 1–18. [Google Scholar] [CrossRef]
- Hamilton, W.D. The genetical evolution of social behaviour II. J. Theor. Biol. 1964, 7, 17–52. [Google Scholar] [CrossRef]
- Trivers, R.L. The evolution of reciprocal altruism. Q. Rev. Biol. 1971, 46, 35–57. [Google Scholar] [CrossRef]
- Brosnan, S.F.; Silk, J.B.; Henrich, J.; Mareno, M.C.; Lambeth, S.P.; Schapiro, S.J. Chimpanzees (Pan troglodytes) do not develop contingent reciprocity in an experimental task. Anim. Cogn. 2009, 12, 587–597. [Google Scholar] [CrossRef]
- Hamann, K.; Warneken, F.; Greenberg, J.R.; Tomasello, M. Collaboration encourages equal sharing in children but not in chimpanzees. Nature 2011, 476, 328. [Google Scholar] [CrossRef]
- Hirata, S.; Fuwa, K. Chimpanzees (Pan troglodytes) learn to act with other individuals in a cooperative task. Primates 2007, 48, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Melis, A.P.; Hare, B.; Tomasello, M. Chimpanzees recruit the best collaborators. Science 2006, 311, 1297–1300. [Google Scholar] [CrossRef] [PubMed]
- Rekers, Y.; Haun, D.B.; Tomasello, M. Children, but not chimpanzees, prefer to collaborate. Curr. Biol. 2011, 21, 1756–1758. [Google Scholar] [CrossRef]
- Suchak, M.; Eppley, T.M.; Campbell, M.W.; de Waal, F.B. Ape duos and trios: Spontaneous cooperation with free partner choice in chimpanzees. PeerJ 2014, 2, e417. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Tanaka, M. Do chimpanzees (Pan troglodytes) spontaneously take turns in a reciprocal cooperation task? J. Comp. Psychol. 2009, 123, 242–249. [Google Scholar] [CrossRef]
- Seed, A.M.; Clayton, N.S.; Emery, N.J. Cooperative problem solving in rooks (Corvus frugilegus). Proc. R. Soc. Lond. B Biol. Sci. 2008, 275, 1421–1429. [Google Scholar] [CrossRef]
- Vonk, J.; Vincent, J.; O’Connor, V. It’s hard to be social alone: Cognitive complexity as transfer within and across domains. Comp. Cogn. Behav. Rev. 2021, 16, 33–67. [Google Scholar] [CrossRef]
- Gräfenhain, M.; Behne, T.; Carpenter, M.; Tomasello, M. Young children’s understanding of joint commitments. Dev. Psychol. 2009, 45, 1430. [Google Scholar] [CrossRef]
- Baciadonna, L.; Cornero, F.M.; Emery, N.J.; Clayton, N.S. Convergent evolution of complex cognition: Insights from the field of avian cognition into the study of self-awareness. Learn. Behav. 2021, 49, 9–22. [Google Scholar] [CrossRef]
- Clayton, N.S. Ways of thinking: From crows to children and back again. Q. J. Exp. Psychol. 2015, 68, 209–241. [Google Scholar] [CrossRef]
- Jaakkola, K.; Guarino, E.; Donegan, K.; King, S.L. Bottlenose dolphins can understand their partner’s role in a cooperative task. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180948. [Google Scholar] [CrossRef] [PubMed]
- Lea, S.E.; Osthaus, B. In what sense are dogs special? Canine cognition in comparative context. Learn. Behav. 2018, 46, 335–363. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, M.K.; Lockwood, P.L.; Rushworth, M.F. Neural mechanisms of social cognition in primates. Annu. Rev. Neurosci. 2018, 41, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Burkart, J.M.; Hrdy, S.B.; Van Schaik, C.P. Cooperative breeding and human cognitive evolution. Evol. Anthropol. Issues News Rev. 2009, 18, 175–186. [Google Scholar] [CrossRef]
- Burkart, J.M.; Van Schaik, C.P. Cognitive consequences of cooperative breeding in primates? Anim. Cogn. 2010, 13, 1–19. [Google Scholar] [CrossRef]
- Duque, J.F.; Stevens, J.R. Voluntary food sharing in pinyon jays: The role of reciprocity and dominance. Anim. Behav. 2016, 122, 135–144. [Google Scholar] [CrossRef]
- Horn, L.; Scheer, C.; Bugnyar, T.; Massen, J.J. Proactive prosociality in a cooperatively breeding corvid, the azure-winged magpie (Cyanopica cyana). Biol. Lett. 2016, 12, 20160649. [Google Scholar] [CrossRef]
- Leete, J.A.; Vonk, J. In mixed company: Two macaws are self-regarding in a symbolic prosocial choice task. Behav. Ecol. Sociobiol. 2022, 76, 1–14. [Google Scholar] [CrossRef]
- Clarke, F.M.; Faulkes, C.G. Dominance and queen succession in captive colonies of the eusocial naked mole–rat, Heterocephalus glaber. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1997, 264, 993–1000. [Google Scholar] [CrossRef]
- Lacey, E.A.; Ebensperger, L.A.; Wolff, J.O.; Sherman, P.W. Social structure in octodontid and ctenomyid rodents. In Rodent Societies: An Ecological and Evolutionary Perspective; Wolff, J.O., Sherman, P.W., Eds.; University of Chicago Press: Chicago, IL, USA, 2007; pp. 403–415. [Google Scholar]
- Corsini, M.T.; Lovari, S.; Sonnino, S. Temporal activity patterns of crested porcupines Hystrix cristata. J. Zool. 1995, 236, 43–54. [Google Scholar] [CrossRef]
- Monetti, L.; Massolo, A.; Sforzi, A.; Lovari, S. Site selection and fidelity by crested porcupines for denning. Ethol. Ecol. Evol. 2005, 17, 149–159. [Google Scholar] [CrossRef]
- Mori, E.; Menchetti, M.; Lucherini, M.; Sforzi, A.; Lovari, S. Timing of reproduction and paternal cares in the crested porcupine. Mamm. Biol. 2016, 81, 345–349. [Google Scholar] [CrossRef]
- Viviano, A.; Amori, G.; Luiselli, L.; Oebel, H.; Bahleman, F.; Mori, E. Blessing the rains down in Africa: Spatiotemporal behaviour of the crested porcupine Hystrix cristata (Mammalia: Rodentia) in the rainy and dry seasons, in the African savannah. Trop. Zool. 2020, 33. [Google Scholar] [CrossRef]
- Coppola, F.; Guerrieri, D.; Simoncini, A.; Varuzza, P.; Vecchio, G.; Felicioli, A. Evidence of scavenging behaviour in crested porcupine. Sci. Rep. 2020, 10, 12297. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.J.; Dorey, N.R. An examination of shaping with an African Crested Porcupine (Hystrix cristata). J. Appl. Anim. Welf. Sci. 2021, 24, 372–378. [Google Scholar] [CrossRef]
- Woods, C.A. Erethizon dorsatum. Mammal Species 1973, 29, 1–6. [Google Scholar] [CrossRef]
- Crawford, M.P. The cooperative solving of problems by young chimpanzees. Comp. Psychol. Monogr. 1937, 14, 1–88. [Google Scholar]
- Chalmeau, R. Do chimpanzees cooperate in a learning task? Primates 1994, 35, 385–392. [Google Scholar] [CrossRef]
- Mendres, K.A.; de Waal, F.B. Capuchins do cooperate: The advantage of an intuitive task. Anim. Behav. 2000, 60, 523–529. [Google Scholar] [CrossRef]
- Plotnik, J.M.; Lair, R.; Suphachoksahakun, W.; de Waal, F.B. Elephants know when they need a helping trunk in a cooperative task. Proc. Natl. Acad. Sci. USA 2011, 108, 5116–5121. [Google Scholar] [CrossRef]
- Ostojić, L.; Clayton, N.S. Behavioural coordination of dogs in a cooperative problem-solving task with a conspecific and a human partner. Anim. Cogn. 2014, 17, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall-Pescini, S.; Schwarz, J.F.; Kostelnik, I.; Virányi, Z.; Range, F. Importance of a species’ socioecology: Wolves outperform dogs in a conspecific cooperation task. Proc. Natl. Acad. Sci. USA 2017, 114, 11793–11798. [Google Scholar] [CrossRef] [PubMed]
- Möslinger, H.; Kotrschal, K.; Huber, L.; Range, F.; Virányi, Z. Cooperative string-pulling in wolves. J. Vet. Behav. Clin. Appl. Res. 2009, 4, 99. [Google Scholar] [CrossRef]
- Péron, F.; Rat-Fischer, L.; Lalot, M.; Nagle, L.; Bovet, D. Cooperative problem solving in African grey parrots (Psittacus erithacus). Anim. Cogn. 2011, 14, 545–553. [Google Scholar] [CrossRef]
- Schwing, R.; Reuillon, L.; Conrad, M.; Noë, R.; Huber, L. Paying attention pays off: Kea improve in loose-string cooperation by attending to partner. Ethology 2020, 126, 246–256. [Google Scholar] [CrossRef]
- Scheid, C.; Noë, R. The performance of rooks in a cooperative task depends on their temperament. Anim. Cogn. 2010, 13, 545–553. [Google Scholar] [CrossRef]
- Hare, B.; Melis, A.P.; Woods, V.; Hastings, S.; Wrangham, R. Tolerance allows bonobos to outperform chimpanzees on a cooperative task. Curr. Biol. 2007, 17, 619–623. [Google Scholar] [CrossRef]
- Melis, A.P.; Hare, B.; Tomasello, M. Engineering cooperation in chimpanzees. Anim. Behav. 2006, 72, 275–286. [Google Scholar] [CrossRef]
- Heaney, M.; Gray, R.D.; Taylor, A.H. Keas perform similarly to chimpanzees and elephants when solving collaborative tasks. PLoS ONE 2017, 12, e0169799. [Google Scholar] [CrossRef]
- Schwing, R.; Jocteur, E.; Wein, A.; Noë, R.; Massen, J.J. Kea cooperate better with sharing affiliates. Anim. Cogn. 2016, 19, 1093–1102. [Google Scholar] [CrossRef]
- Massen, J.J.; Ritter, C.; Bugnyar, T. Tolerance and reward equity predict cooperation in ravens (Corvus corax). Sci. Rep. 2015, 5, 15021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, I.F.; Osvath, M. The string-pulling paradigm in comparative psychology. J. Comp. Psychol. 2015, 129, 89. [Google Scholar] [CrossRef] [PubMed]
- Seed, A.M.; Jensen, K. Large-scale cooperation. Nature. 2011, 472, 424–425. [Google Scholar] [CrossRef] [PubMed]
- Stringham, S.F. Salmon fishing by bears and the dawn of cooperative predation. J. Comp. Psychol. 2012, 126, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Drea, C.M.; Carter, A.N. Cooperative problem solving in a social carnivore. Anim. Behav. 2009, 78, 967–977. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truax, J.; Vonk, J.; Vincent, J.L.; Bell, Z.K. Teamwork Makes the String Work: A Pilot Test of the Loose String Task with African Crested Porcupines (Hystrix cristata). J. Zool. Bot. Gard. 2022, 3, 448-462. https://doi.org/10.3390/jzbg3030034
Truax J, Vonk J, Vincent JL, Bell ZK. Teamwork Makes the String Work: A Pilot Test of the Loose String Task with African Crested Porcupines (Hystrix cristata). Journal of Zoological and Botanical Gardens. 2022; 3(3):448-462. https://doi.org/10.3390/jzbg3030034
Chicago/Turabian StyleTruax, Jordyn, Jennifer Vonk, Joy L. Vincent, and Zebulon Kade Bell. 2022. "Teamwork Makes the String Work: A Pilot Test of the Loose String Task with African Crested Porcupines (Hystrix cristata)" Journal of Zoological and Botanical Gardens 3, no. 3: 448-462. https://doi.org/10.3390/jzbg3030034