
Bone-Regulating MicroRNAs and Resistance Exercise: A Mini-Review


1
Department of Health and Exercise Science, University of Oklahoma, Norman, OK 73019, USA
2
Department of Kinesiology, California State University, San Bernardino, 5500 University Parkway, San Bernardino, CA 92407, USA
3
Department of Health and Human Performance, University of Texas Rio Grande Valley, Edinburg, TX 78539, USA
*
Author to whom correspondence should be addressed.
Osteology 2022, 2(1), 11-20; https://doi.org/10.3390/osteology2010002
Submission received: 22 October 2021
/
Revised: 20 December 2021
/
Accepted: 29 December 2021
/
Published: 3 January 2022
(This article belongs to the Special Issue Bone Biomechanics: Diseases, Treatment and Rehabilitation)
Abstract
:MicroRNAs (miRNA) are a class of short noncoding RNA that play important roles in controlling gene expression. Many miRNAs have been identified as being important regulators of bone cell function, thus affecting the bone remodeling processes. In addition to being expressed in specific tissues and exerting intracellular effects, miRNAs can enter the blood where they can be taken up by other tissues. These circulating miRNAs (c-miRNA) also have clinical significance as biomarkers of musculoskeletal diseases as they are tissue-specific, are stable and easily detectable, and require minimally invasive procedures. This mini-review discusses miRNAs with regulatory roles in bone metabolism and c-miRNA responses to acute bouts of resistance exercise. MiRNA responses (e.g., upregulation/downregulation of expression) vary depending on the resistance exercise protocol characteristics and the age of the participants. There are gaps in the literature that need to be addressed as most of the resistance exercise studies focused on miRNAs that regulate skeletal muscle in male participants.
1. Introduction
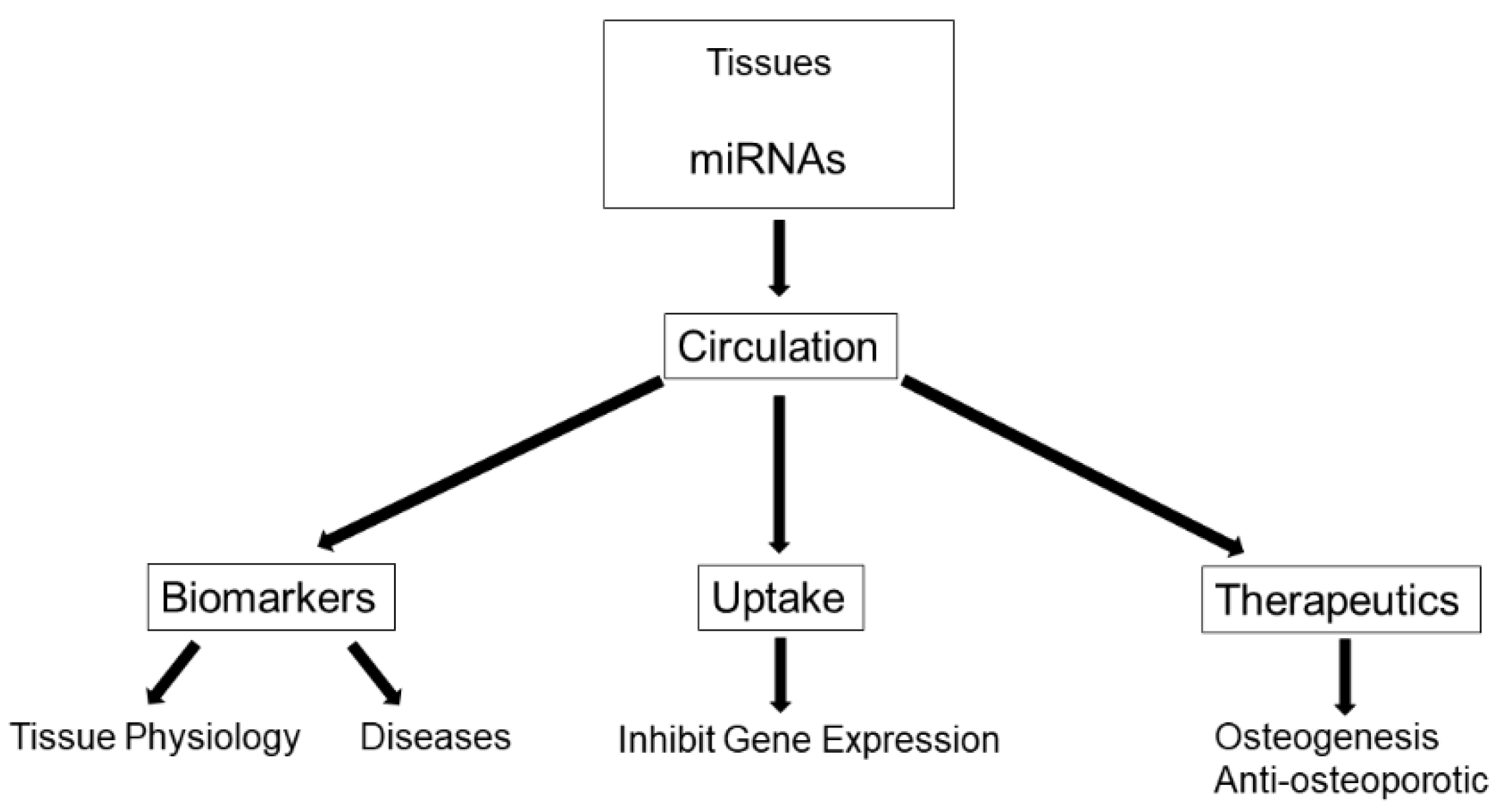
A rapidly emerging topic in osteoporosis research is the use of microRNAs (miRNA, miR) as biomarkers of bone status since they are key regulatory players in bone remodeling [1,2,3]. MiRNAs are a class of short (21–22 nucleotides (nt) in length) noncoding RNA that play important roles in controlling gene expression; specifically as negative regulators of gene expression [4]. A large number of miRNAs are important regulators of bone cell function, with some miRNAs promoting or suppressing bone formation and others promoting or suppressing bone resorption [1,3]. In addition to being expressed in specific tissues and having intracellular effects, miRNAs can enter into the blood in response to various stimuli including tissue injury and mechanical stress. These circulating miRNAs (c-miRNA) have clinical significance as biomarkers of musculoskeletal diseases as they are tissue-specific, are stable and easily detectable, and require minimally invasive procedures (e.g., venous blood sample) [2]. Recent findings documenting dysregulated miRNAs in the serum of osteoporosis patients [5,6,7], have generated much excitement as they support the use of circulating miRNAs as biomarkers for bone loss and osteoporosis [2] as well as highlight their therapeutic potential for bone disease [8] (Figure 1).
Besides their basic tissue physiology effects, c-miRNAs may have important regulatory functions for physiological adaptations to exercise training, and may be useful as biomarkers of exercise responses. These topics are of growing interest to exercise scientists (refer to [9,10] for in depth reviews). High intensity resistance exercise has been shown to have positive effects on bone mineral density [11] and bone turnover markers [12]. Muscle contraction can stimulate osteogenesis by producing a strain stimulus and through the release of miRNAs. Much of the literature on miRNA responses to resistance exercise has focused on miRNAs that regulate skeletal muscle; however, miRNAs also may play a regulatory role in bone adaptations to resistance exercise. This mini-review will discuss miRNA physiology, the regulatory role of miRNAs in bone metabolism and osteoporosis, and miRNA responses to mechanical loading and resistance exercise.
2. MiRNA Physiology
MiRNA genes are initially transcribed by RNA polymerase II from the introns of messenger RNA (mRNA) in the nucleus to form primary miRNA, which is folded and contains a hairpin structure [13]. The enzyme complex containing Drosha and DiGeorge syndrome critical region 8 (DGCR8) edit the primary miRNA to pre-miRNA. Pre-miRNA is transported to the cytoplasm via Exportin-5. Once in the cytoplasm, the pre-miRNA is bound to an Argonaut protein. The enzyme, Dicer, cleaves the hairpin structure of the pre-miRNA to form a miRNA duplex. One strand of the duplex disassociates and is degraded. The remaining strand, now a mature miRNA, bound to the Argonaute protein forms the RNA-Induced Silencing Complex (RISC). The RISC complex negatively regulates protein translation post-transcription by selectively binding nt 2–9 to the 3′ untranslated region of mRNA. Binding to mRNA either interferes with protein translation or leads to the degradation of the mRNA. Multiple RNAs are targetable by a single MiRNA, and greater than 60% of human genes are affected by them [14].
3. Regulatory Role of MiRNAs in Bone Metabolism
Previous research has indicated that miRNAs are involved in all stages of osteogenesis by regulating differentiation, proliferation, apoptosis, and activity of different bone cells: osteocytes, osteoblasts, osteoclasts, and bone marrow stem cells (Table 1) [3]. Bone-regulating miRNAs are those expressed in osteoblast lineage cells for regulation of bone formation by either direct repression of inhibitors of osteoblast differentiation or by their response to osteogenic signals, such as BMP (Bone Morphogenetic Protein), to promote osteogenesis. A growing number of MiRNAs have been identified to regulate osteoblastogenesis and bone formation by targeting inhibitors of osteogenesis or osteogenic factors [3]. Some MiRNAs stimulate osteoblastogenesis, such as miR-15b, -20a, whereas some MiRNAs inhibit osteoblastogenesis, such as miR-23a-3p, -100-5p, and -133a-3p. This occurs primarily through regulating BMP-RUNX2 (Runt-Related Transcription Factor 2) and Wnt (Wingless-Type MMTV Integration Site Family) signaling pathways. For example, miR-100-5p directly targets BMPR2 (BMP receptor type II) and inhibits osteoblastogenesis. Research on the role of miRNAs in osteocytes is in its early stages, however, miRNAs are expressed in osteocytes and have been shown to regulate osteocyte differentiation [15]. MiRNAs also regulate the differentiation and proliferation of chondrocytes; miR-125b-5p is reported to stimulate the differentiation of chondrocytes in mouse by repressing an inhibitor of chondrogenesis [16].
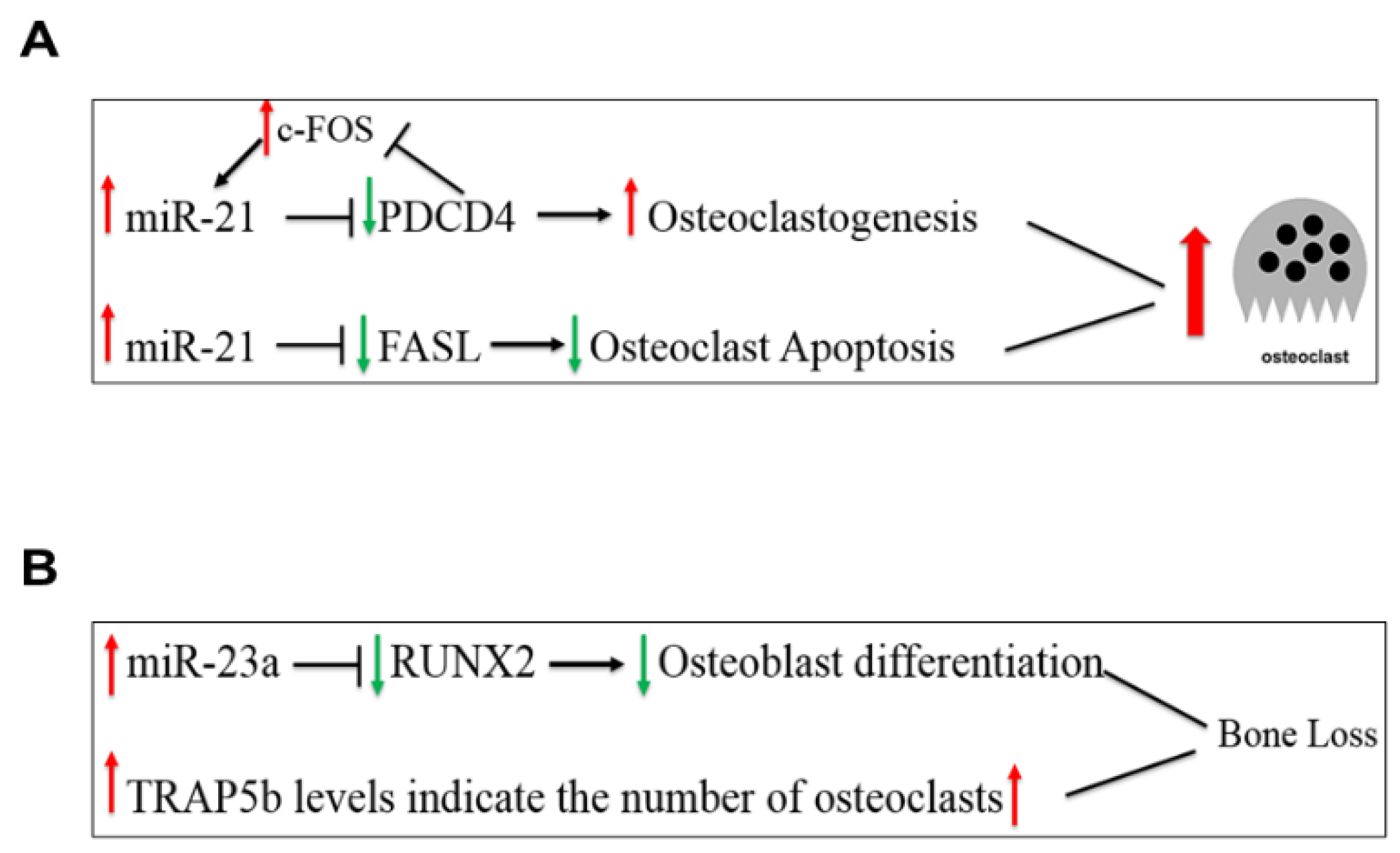
MiR-21-5p, a widely studied miRNA, is ubiquitously expressed and has regulatory roles in both osteoblast and osteoclast activities [2,3,17,18]. MiR-21-5p promotes bone formation by inhibiting SMAD7 (Small Mothers Against Decapentaplegic 7), a protein that inhibits osteoblast differentiation via the BMP and TGF-β (Transforming Growth Factor-Beta) pathways. MiR-21-5p also promotes osteoclastogenesis via a positive feedback loop involving c-Fos, a factor that promotes osteoclast formation, and PDCD4 (Programmed Cell Death Protein 4), a protein that acts to inhibit osteoclast formation through c-Fos inhibition [17,19]. In the positive feedback loop, c-Fos upregulates miR-21-5p expression, which then suppresses PDCD4 expression, thereby reducing its inhibitory effect on c-Fos (Figure 2A) [20]. Estrogen deficiency in postmenopausal women upregulates miR-21-5p, which acts to suppress the transcription of FASL (Fas Ligand) [21]. As a result, osteoclast apoptosis is inhibited, promoting bone resorption activity. Another miRNA, miR-23a-3p targets the RUNX2 and SMAD3 (Small Mothers Against Decapentaplegic 3) pathways, inhibiting osteoblast differentiation [22], thus high levels of miR-23a-3p inhibit bone formation, and the net imbalance favors bone resorption (Figure 2B). We recently reported a positive relationship between miR-23a-3p and the bone resorption marker, TRAP5b (Transforming Growth Factor-Beta), in postmenopausal women that may be attributed to their regulatory roles on bone resorption since high serum TRAP5b concentrations indicate a greater number of osteoclasts, which favors bone resorption [23].
4. MiRNAs and Osteoporosis
MiRNAs regulate osteogenesis and are associated with bone diseases such as osteoporosis (refer to review [3]) and may serve as biomarkers for osteoporosis. Wang et al. [24] measured miRNAs in circulating monocytes in 20 postmenopausal women. The cohort was dichotomized into high and low BMD groups and compared for miRNA expression levels. Of the miRNAs assessed, miR-133a-3p was expressed significantly higher in the low BMD group. The target for miR-133a-3p in bone is RUNX2, and higher levels in tissue would have a negative impact on bone formation.
Seeliger et al. [5] analyzed circulating and bone tissue miRNAs in osteoporotic (n = 10) and non-osteoporotic (n = 10) patients with hip fractures. In the two sample types, 5 miRNAs (miR-21-5p, -23a-3p, -24, -100-5p, -125b-5p) were expressed higher in osteoporotic patients. MiR-21-5p may affect bone through inhibition of PDCD4, which allows for osteoclast survival. MiR-23a-3p and -24 are implicated in inhibition of RUNX2 for osteocyte formation. MiR-100-5p may negatively affect bone formation by inhibiting BMPR2, thus halting the growth of pre-osteoblasts into mature osteoblasts. MiR-125b-5p may also affect formation by inhibiting mesenchymal stem cell differentiation towards an osteoblastic lineage. Weilner at al. [25] also assessed miRNAs in osteoporotic fracture patients. Serum samples from 37 postmenopausal women (>65 years) were collected, 19 of which had recent fractures due to osteoporosis. Samples were taken within two weeks of surgery. Differentially expressed miRNAs included three upregulated (miR-10a-5p, 10b-5p, -22-3p) and three downregulated (miR-133b -328-3p, let-7g-5p). A validation cohort was also used to validate the differentially expressed miRNAs. It was found that only miR-22-3p, -328-3p and let-7g-5p reached significance. In the case of miR-22-3p, the first study saw significant upregulation while the validation cohort had significant downregulation. Of note, there were no differences between fracture patients and controls for miR-21-5p. Panach et al. [6] assessed c-miRNA in eight osteoporotic fracture patients (mean 80 years) compared to five controls with osteoarthritis. They found that miR-122-5p, -125-5p, -21-5p were significantly upregulated in the fracture patients. MiR-21-5p was highly correlated with the bone resorption marker, CTX-I (C-telopeptide of Type I collagen cross-links) and several miRNAs (miR-21-5p, -125b-5p, -122-5p, -210) were positively correlated with bone formation markers (osteocalcin, bone-specific alkaline phosphatase). Although miR-21-5p can have an osteogenic effect, it appears that in fracture patients, miR-21-5p serves to improve osteoclastic function to a greater degree. Kelch et al. [7] found that miR-21-5p expression was higher in both serum and in bone tissue of male and female patients with osteoporotic fractures versus normal controls, suggesting a role for miR-21-5p in bone loss. Both cultured osteoclast and osteoblast cells from bone tissue of osteoporotic fracture patients showed upregulation of miR-21-5p expression.
In a recent study, we assessed circulating miRNAs in 75 postmenopausal women aged 60–85 years based on osteoporosis and sarcopenia status [23]. We measured miR-1-3p, -21-5p, -23a-3p, -24-3p, -100-5p, -125b-5p, -133a-3p, -206, miRNAs associated with targets in muscle and bone, and the bone resorption markers, CTX-I and TRAP5b. There were no significant differences in c-miRNA expression between osteoporotic and non-osteoporotic and sarcopenic and non-sarcopenic participants; however, there were potentially biologically relevant differences in fold changes of miR-21-5p and miR-23a-3p, which were upregulated in osteoporotic women, and in miR-125b-5p which was downregulated in the osteoporotic group.
5. MiRNAs and Mechanical Loading
In vitro studies have shown that mechanical loading changes miRNA expression, which in turn, may be important regulatory mechanisms for the exercise-induced adaptations in bone metabolism [26]. Cell line studies provide evidence that miR-21-5p is sensitive to mechanical loading as it was upregulated in human periodontal stem cells by a stretch load [27] but downregulated in MC3T3-E1 cells by fluid shear stress promoting osteogenic differentiation [28]. In mice, miR-21-5p deficiency inhibited osteoclast function and bone resorption leading to increased trabecular bone accrual [17].
5.1. Skeletal Muscle MiRNAs
MiRNAs are involved in the regulation of hypertrophy and other muscular adaptations. Table 2 shows a summary of skeletal muscle and serum/plasma miRNA responses to acute bouts of resistance exercise studies in humans. Drummond et al. [29] compared miRNA expression in skeletal muscle following acute resistance exercise and essential amino acids ingestion in young (mean 29 years) and older (mean 70 years) men. MiR-1 expression was downregulated in young but not in older men following the exercise stimulus, indicating that aging results in a dysregulated miRNA response after an anabolic stimulus. Rivas et al. [30] also compared the skeletal muscle miRNA responses to acute resistance exercise in younger and older men. Similar to Drummond et al. [29], they found that alterations in miRNA expression were age-dependent, as a large number of miRNA were downregulated and 1 miRNA (miR-486-5p) upregulated in young men, whereas there were no miRNA changes in skeletal muscle of older men after the resistance exercise bout.
Telles et al. [31] compared skeletal muscle miRNA responses in young men to three types of acute exercise protocols; resistance exercise, high-intensity interval exercise, and both resistance and high-intensity interval exercise performed concurrently. They used a targeted approach for the miRNA analysis, selecting 8 miRNAs with known regulatory roles in skeletal muscle development. Their main findings were that 7/8 miRNAs were upregulated up to 8 h after acute exercise, with 6 miRNA (miR-1-3p, -133a-3p, -133b-3p, -181a-3p, -378a-5p, -486) showing similar responses for all 3 exercise protocols, and 2 miRNAs (miR-23a-3p, -206) having greater changes in expression for the resistance exercise compared to the high-intensity interval exercise protocol.
5.2. Circulating MiRNAs
Sawada et al. [32] assessed c-miRNA responses in 12 young men (~29 years) performing bench press and bilateral leg press. Participants performed five sets of ten repetitions at 70% 1 RM and had serum samples taken before, immediately after, 60 min, 1 day, and 3 days after exercise. There was no significant change for miR-133 nor miR-21-5p post-exercise, only downregulation of miR-146a and miR-221 three days post-exercise and upregulation of miR-149a 24 h post-exercise. MiR-21-5p had a weak positive correlation with the catecholamines, epinephrine and norepinephrine, and testosterone and IGF-1 were weakly positively correlated with miR-222.
C-miRNA expression changes to acute exercise may be attenuated by aging. Margolis et al. [33] reported different patterns of c-miRNA expression in response to acute high intensity resistance exercise with aging as c-miRNA profiles were downregulated in older men but upregulated in younger men. Three miRNAs, miR-19b-3p, -206, -486, significantly predicted skeletal muscle aging. Of the 10 c-miRNA that were differentially expressed in young versus older men, 6 (miR-19a-3p, -19b-3p, -20a-5p, -26b-5p, -143-3p, -195-5p) were associated with inhibition of the PI3K-Akt pathway, which would increase activation of the Akt-mTOR pathway and increase cellular protein synthesis.
D’Souza et al. [34] observed the effects of an acute resistance exercise bout on the expression of miRNAs in skeletal muscle tissue and plasma in 9 healthy young males. Subjects performed 6 sets of 8–10 repetitions for leg press and 8 sets of 8–10 repetitions for knee extension. Samples were collected pre, 2 h, and 4 h post-exercise. From skeletal muscle biopsy, miR-133a-3p, -206a increased 2 h post- and miR-146a-5p increased 4 h post-exercise. MiR-23a-3p decreased 2 h post- and further decreased 4 h post-exercise. Of the c-miRNA, miR-133a-3p and -149-5p were elevated from baseline, while miR-1, -208a, and -499 were undetected. Only miR-133a-3p had increased expression in both plasma and skeletal muscle tissue and may serve as a marker for muscle damage.
Cui et al. [35] assessed the time-course of c-miRNA expression from three different resistance exercise protocols in young men. The three protocols consisted of five exercises: bench press, squat, pulldown, overhead press, and standing dumbbell curl and were performed at different volumes and intensities. The strength-endurance (SE) group performed three sets of 16–20 repetitions at 40% 1RM with one-min rest intervals. The muscular hypertrophy (MH) group performed three sets of 12 repetitions at 70% of 1RM with two-min rest intervals. The maximum strength (MS) group performed four sets of six repetitions at 90% of 1RM with three-min rest intervals. For the SE group, miR-532 fold changes increased 1 h post-exercise and remained elevated 24 h later while miR-208b decreased immediately post-exercise and remained depressed. The MH group showed decreased expression for miR-21, and -133a, immediately post-exercise, and miR-221 1 h post-exercise. MiR133b was upregulated 24 h post-exercise, while miR-181a and miR-206 were upregulated 1 h post-exercise, then returned to baseline 24 h later. For the MS group, miR-133a significantly decreased expression immediately post- and returned to baseline 1 h post-exercise. MiR-133b increased expression from immediately post- to 1 h post-exercise. The authors concluded that miR-133a may be a potential biomarker of physiological muscle strain for resistance exercise at very high intensities (e.g., 90% 1RM). The changes seen in miR-21 may be indicative of an initial pro-inflammatory response, followed by an anti-inflammatory response.
Evaluating c-miRNA responses to exercise may provide insight into the potential benefits of exercise in ameliorating the negative effects of aging on musculoskeletal health. There is a paucity of data in the literature on c-miRNA responses to acute exercise in postmenopausal women. We recently studied the effects of acute resistance exercise and whole-body vibration on expression of selected c-miRNAs in physically active postmenopausal women aged 65–76 years [36]. This randomized crossover design study compared c-miRNA responses to a bout of resistance exercise (RE) (3 sets 10 reps 70% 1 repetition maximum (1RM), 5 exercises) and a bout of whole-body vibration (WBV) (5 sets 1 min bouts 20 Hz 3.38 mm peak to peak displacement. We targeted 4 c-miRNAs (miR-21-5p, -23a-3p, -133a-3p, -148a-3p) that have been shown in the literature to regulate bone metabolism. The main finding was that c-miR-21-5p expression was downregulated from 60 min post to 24 h post-exercise for only the whole-body vibration protocol. Absolute changes in the bone resorption marker (TRAP5b) were negatively correlated with c-miR-21-5p fold changes for both whole-body vibration and resistance exercise modalities and there were significant moderate to strong negative correlations between baseline c-miRNAs and bone status variables. Based on our findings, whole-body vibration induces a sufficient mechanical stimulus for altering c-miR-21-5p expression. The lack of c-miRNA responses to the high intensity resistance exercise protocol may be related to the older age of the postmenopausal women, since previous studies [29,30,33] have reported age-associated differences in miRNA responses to a resistance exercise stimulus.

{kind=link}
{kind=link}
{kind=link}
Table 2.
MiRNA Responses to Acute Resistance Exercise (RE) Protocols.
| Study | RE Protocol | Target Population | Sample | Upregulation | MiRNA Responses Downregulation | No Change |
|---|---|---|---|---|---|---|
| Drummond et al. [29] | Leg extension 8 sets 10 reps 70% 1RM | Young men (n = 6) Older men (n = 6) | Muscle | Young only miR-1 | Young miR-133a miR-206 All in older men | |
| Sawada et al. [32] | Bench press Leg Press 5 sets 10 reps 70% 1RM | Young men (n = 12) | Serum | miR-149 | miR-146a miR-221 | |
| Rivas et al. [30] | Bilateral Knee Extension 3 sets 10 reps 80% 1RM | Young men (n = 8) Older men (n = 8) | Muscle | Young only miR-486-5p | Young only miR-23b-3p -24-3p, -26a-3p -27a-3p, -27b-3p -29c-3p, -30a-5p -30d-5p, -95-3p -126-3p, -133a -133b, -140-3p -181a-3p, -378a-5p | All in older men |
| Margolis et al. [33] | Bilateral Knee Extension, Leg Press 3 sets 10 reps 80% 1RM | Young men (n = 9) Older men (n = 9) | Serum | Young miR-221-3p -222-3p -206 | Older miR-31-5p -124-3p, -211-5p -375 | |
| D’Souza et al. [34] | Leg press 6 sets 8–10 reps 80% 1RM Knee extension 8 sets 8–10 reps 80% 1RM | Young men (n = 9) | Muscle Plasma | Muscle miR-23a-3p, -133a-3p, -146a-5p, -206a, -378b -486-5p Plasma miR-133a-3p -149-5p | ||
| Cui et al. [35] | Bench press, Squat, Pull-down, Overhead Press, Dumbbell Curl Hypertrophy 3 sets 12 reps 70% 1RM Strength 4 sets 6 reps 90% 1RM Strength Endurance | Young men (n = 45) | Plasma | Hypertrophy miR-133b -181a, -206 Strength miR-133b Strength-Endurance miR-532 | Hypertrophy miR-21 -133a, -221 Strength miR-133a Strength-Endurance miR-208b | |
| 3 sets 16–20 reps 40% 1RM | ||||||
| Telles et al. [31] | Leg press Knee extension 4 sets 8–12 RM High-intensity interval exercise (HIIE) Concurrent resistance and high-intensity interval exercise (CON) | Young men (n = 9) | Muscle | all 3 protocols miR-1-3p -133a-3p -133b-3p -181a-3p -486, RE > HIIE, CON miR-23a-3p,-206 | all 3 protocols miR-378a-5p | |
| Buchanan [36] | Leg press Shoulder press Lat pulldown Knee extension Hip adduction 3 sets 10 reps 70–75% 1RM Whole-body vibration (WBV) 5–1 min sets 3.38 mm peak-to-peak displacement 2.7 g | Postmenopausal women (n = 10) | Serum | WBV only miR-21-5p | all for RE |
miR—MicroRNA; RM—Repetition Maximum.
6. Conclusions
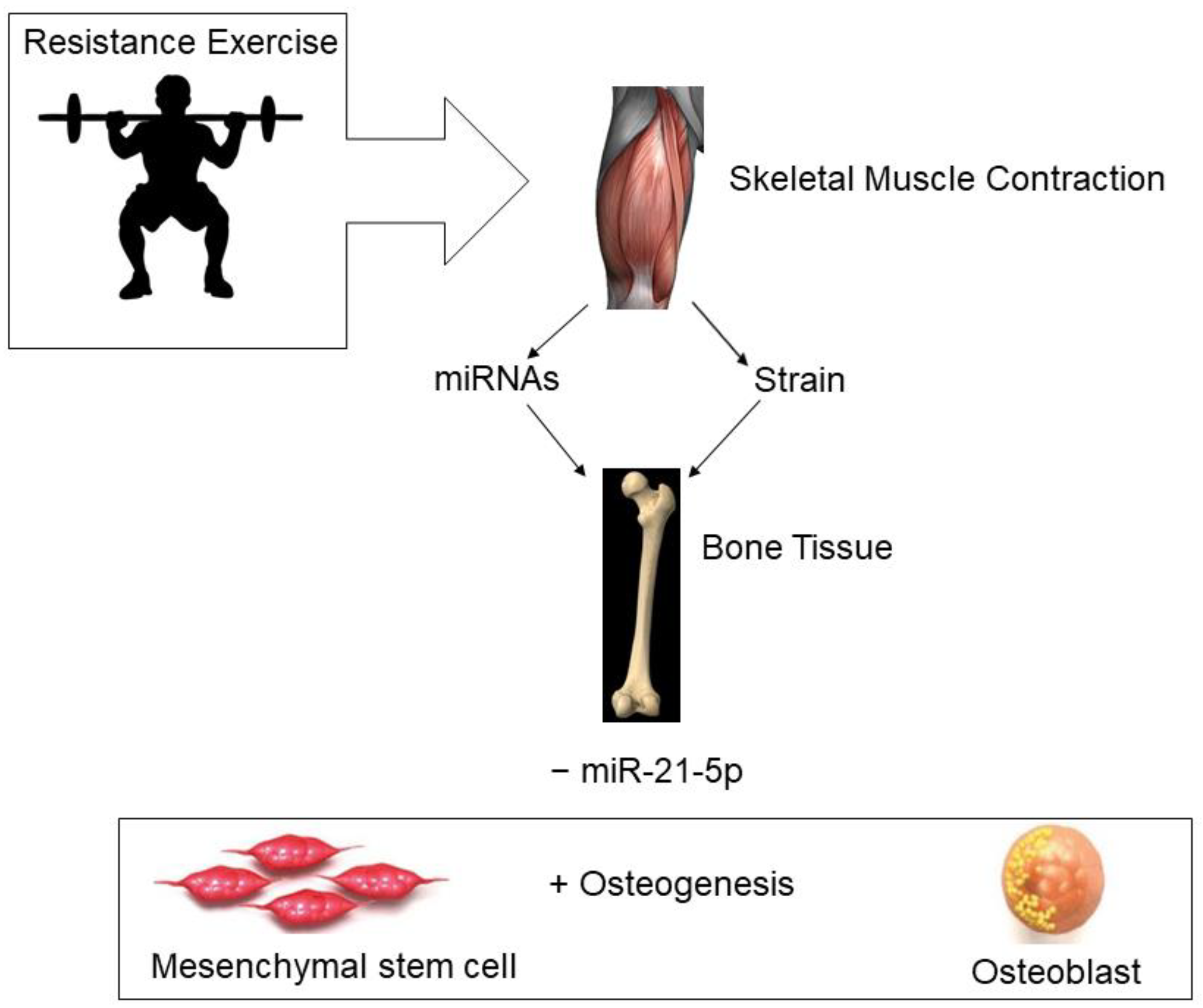
The assessment of miRNAs, particularly in the blood, is a rapidly growing area of research that may increase the understanding of the regulatory roles of miRNAs in disease pathology and in physiological responses of tissues to exercise. Figure 3 depicts a theoretical model for miRNA and bone formation responses to the mechanical strain induced by skeletal muscle contraction during resistance exercise. To date, resistance exercise studies have focused on miRNAs regulating muscle tissue, not bone tissue, and most of the studies have been conducted in men. The available data suggest that miRNAs are differentially expressed in young versus older participants in response to resistance exercise. There are gaps in the literature that need to be addressed, specifically more information is needed on bone-specific miRNA responses to resistance exercise, and more studies need to be conducted in women, especially postmenopausal women who are at greater risk for osteoporosis.
Author Contributions
D.A.B. wrote the first draft of the manuscript, created Figure 1 and Figure 3 and contributed to the literature review. Z.C. contributed to the literature review and article retrieval, edited and reviewed the manuscript and created Figure 2. S.R.B. contributed to the literature review and article retrieval and edited and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study by Buchanan [36] was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of the University of Oklahoma Health Sciences Center (IRB# 9569 approval date 14 August 2018).
Informed Consent Statement
Written informed consent was obtained from all subjects involved in the Buchanan study [36].
Data Availability Statement
The data used to support the findings of the Buchanan study [36] are restricted by the University of Oklahoma IRB in order to protect participant privacy. Data may be available in aggregate form from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BMD | bone mineral density |
| BMP | Bone Morphogenetic Protein |
| BMPR/BMPR2 BMP | receptor/type II |
| c-miRNA | circulating microRNA |
| CTX-I | C-telopeptide of Type I collagen cross-links |
| CXCL11 C-X-C | Motif Chemokine Ligand 11 |
| DXA | dual energy x-ray absorptiometry |
| eIF4E3 Eukaryotic | translation initiation factor 4E family member 3 |
| FASL | Fas Ligand |
| FC | fold change |
| KDM6B | Lysine Demethylase 6B |
| MAPK | Mitogen-Activated Protein Kinase |
| miRNA | microRNA |
| c-miRNA | circulating miRNA |
| MAFB | V-maf musculoaponeurotic fibrosarcoma oncogene homolog B |
| MCSA | Muscle cross-sectional area |
| MMP13 | Matrix Metalloproteinase 13 |
| nt | nucleotides |
| OSX | Osterix |
| PDCD4 | Programmed Cell Death Protein 4 |
| RE | resistance exercise |
| 1 RM 1 | Repetition Maximum |
| RNA | Ribonucleic Acid |
| RUNX2 | Runt-Related Transcription Factor 2 |
| SMAD1 | Small Mothers Against Decapentaplegic 1 |
| SMAD3 | Small Mothers Against Decapentaplegic 3 |
| SMAD7 | Small Mothers Against Decapentaplegic 7 |
| TGF-β | Transforming Growth Factor-Beta |
| TRAP5b | Tartrate-resistant acid phosphatase 5b |
| WBV | whole-body vibration |
| Wnt Wingless-Type MMTV Integration Site Family | |
References
- Van Wijnen, A.J.; van de Peppel, J.; van Leeuwen, J.P.; Lian, J.B.; Stein, G.S.; Westendorf, J.J.; Oursler, M.-J.; Im, H.-J.; Taipaleenmäki, H.; Hesse, E.; et al. MicroRNA functions in osteogenesis and dysfunctions in osteoporosis. Curr. Osteoporos. Rep. 2013, 11, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackl, M.; Heilmeier, U.; Weilner, S.; Grillari, J. Circulating microRNAs as novel biomarkers for bone diseases—Complex signatures for multifactorial diseases? Mol. Cell Endocrinol. 2016, 432, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Bellavia, D.; De Luca, A.; Carina, V.; Costa, V.; Raimondi, L.; Salamanna, F.; Alessandro, R.; Fini, M.; Giavaresi, G. Deregulated miRNAs in bone health: Epigenetic roles in osteoporosis. Bone 2019, 122, 52–75. [Google Scholar] [CrossRef]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeliger, C.; Karpinski, K.; Haug, A.T.; Vester, H.; Schmitt, A.; Bauer, J.S.; van Griensven, M. Five freely circulating miRNAs and bone tissue miRNAs are associated with osteoporotic fractures. J. Bone Miner. Res. 2014, 29, 1718–1728, Erratum in J. Bone. Miner. Res. 2015, 30, 195–196. [Google Scholar] [CrossRef]
- Panach, L.; Mifsut, D.; Tarín, J.J.; Cano, A.; García-Pérez, M.Á. Serum Circulating MicroRNAs as Biomarkers of Osteoporotic Fracture. Calcif. Tissue Int. 2015, 97, 495–505. [Google Scholar] [CrossRef]
- Kelch, S.; Balmayor, E.R.; Seeliger, C.; Vester, H.; Kirschke, J.S.; van Griensven, M. miRNAs in bone tissue correlate to bone mineral density and circulating miRNAs are gender independent in osteoporotic patients. Sci. Rep. 2017, 7, 15861. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Shen, G.; Ren, R.; Liang, D.; Yu, X.; Zhang, Z.; Huang, J.; Qiu, T.; Tang, J.; Shang, Q.; et al. Therapeutic potential of microRNAs in osteoporosis function by regulating the biology of cells related to bone homeostasis. J. Cell Physiol. 2018, 233, 9191–9208. [Google Scholar] [CrossRef]
- Sapp, R.M.; Shill, D.D.; Roth, S.M.; Hagberg, J.M. Circulating microRNAs in acute and chronic exercise: More than mere biomarkers. J. Appl. Physiol. 2017, 122, 702–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Sanjurjo, M.; de Gonzalo-Calvo, D.; Fernández-García, B.; Sergio, D.-R.; Ángel, M.-C.; Hugo, O.; Alberto, D.; Eduardo, I.-G. Circulating microRNA as Emerging Biomarkers of Exercise. Exerc. Sport Sci. Rev. 2018, 46, 160–171. [Google Scholar] [CrossRef]
- Shojaa, M.; von Stengel, S.; Kohl, M.; Schoene, D.; Kemmler, W. Effects of dynamic resistance exercise on bone mineral density in postmenopausal women: A systematic review and meta-analysis with special emphasis on exercise parameters. Osteoporos. Int. 2020, 31, 1427–1444. [Google Scholar] [CrossRef] [PubMed]
- Pasqualini, L.; Ministrini, S.; Lombardini, R.; Bagaglia, F.; Paltriccia, R.; Pippi, R.; Collebrusco, L.; Reginato, E.; Sbroma Tomaro, E.; Marini, E.; et al. Effects of a 3-month weight-bearing and resistance exercise training on circulating osteogenic cells and bone formation markers in postmenopausal women with low bone mass. Osteoporos. Int. 2019, 30, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Metazoan microRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, L.I.; Wallace, J.M. MicroRNAs and osteocytes. Bone 2021, 150, 115994. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.T.; Xiao, P. MiRNAs in bone diseases. Microrna 2013, 2, 20–31. [Google Scholar] [CrossRef]
- Hu, C.H.; Sui, B.D.; Du, F.Y.; Shuai, Y.; Zheng, C.X.; Zhao, P.; Yu, X.R.; Jin, Y. miR-21 deficiency inhibits osteoclast function and prevents bone loss in mice. Sci. Rep. 2017, 7, 43191. [Google Scholar] [CrossRef] [Green Version]
- Cheng, V.K.; Au, P.C.; Tan, K.C.; Cheung, C.L. MicroRNA and human bone health. JBMR Plus. 2018, 3, 2–13. [Google Scholar] [CrossRef]
- Sugatani, T.; Vacher, J.; Hruska, K.A. A microRNA expression signature of osteoclastogenesis. Blood 2011, 117, 3648–3657. [Google Scholar] [CrossRef]
- Gennari, L.; Bianciardi, S.; Merlotti, D. MicroRNAs in bone diseases. Osteoporos. Int. 2017, 28, 1191–1213. [Google Scholar] [CrossRef]
- Sugatani, T.; Hruska, K.A. Down-regulation of miR-21 biogenesis by estrogen action contributes to osteoclastic apoptosis. J. Cell Biochem. 2013, 114, 1217–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.Q.; Gordon, J.A.; Beloti, M.M.; Croce, C.M.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Liana, J.B. A network connecting Runx2, SATB2, and the miR-23a∼27a∼24-2 cluster regulates the osteoblast differentiation program. Proc. Natl. Acad. Sci. USA 2010, 107, 19879–19884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Bemben, M.G.; Bemben, D.A. Bone and muscle specific circulating microRNAs in postmenopausal women based on osteoporosis and sarcopenia status. Bone 2019, 120, 271–278. [Google Scholar] [CrossRef]
- Wang, J.; Cui, Q. Specific roles of microRNAs in their interactions with environmental factors. J. Nucleic Acids 2012, 2012, 978384. [Google Scholar] [CrossRef]
- Weilner, S.; Skalicky, S.; Salzer, B.; Keider, V.; Wagner, M.; Hildner, F.; Gabriel, C.; Dovjak, P.; Pietschmann, P.; Grillari-Voglauer, R.; et al. Differentially circulating miRNAs after recent osteoporotic fractures can influence osteogenic differentiation. Bone 2015, 79, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zhang, L.; Tong, X.; Zhang, M.; Zhao, Y.; Guo, J.; Lei, L.; Chen, X.; Tickner, J.; Xu, J.; et al. Mechanical stress regulates bone metabolism through microRNAs. J. Cell Physiol. 2017, 232, 1239–1245. [Google Scholar] [CrossRef]
- Wei, F.; Liu, D.; Feng, C.; Zhang, F.; Yang, S.; Hu, Y.; Ding, G.; Wang, S. MicroRNA-21 mediates stretch-induced osteogenic differentiation in human periodontal ligament stem cells. Stem. Cells Dev. 2015, 24, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Mai, Z.H.; Peng, Z.L.; Zhang, J.L.; Chen, L.; Liang, H.Y.; Cai, B.; Ai, H. miRNA expression profile during fluid shear stress-induced osteogenic differentiation in MC3T3-E1 cells. Chin. Med. J. (Eng) 2013, 126, 1544–1550. [Google Scholar]
- Drummond, M.J.; McCarthy, J.J.; Fry, C.S.; Esser, K.A.; Rasmussen, B.B. Aging differentially affects human skeletal muscle microRNA expression at rest and after an anabolic stimulus of resistance exercise and essential amino acids. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1333–E1340. [Google Scholar] [CrossRef] [Green Version]
- Rivas, D.A.; Lessard, S.J.; Rice, N.P.; Lustgarten, M.S.; So, K.; Goodyear, L.J.; Parnell, L.D.; Fielding, R.A. Diminished skeletal muscle microRNA expression with aging is associated with attenuated muscle plasticity and inhibition of IGF-1 signaling. FASEB J. 2014, 28, 4133–4147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telles, G.D.; Libardi, C.A.; Conceição, M.S.; Vechin, F.C.; Lixandrão, M.E.; DE Andrade, A.L.L.; Guedes, D.N.; Ugrinowitsch, C.; Camera, D.M. Time Course of skeletal muscle miRNA expression after resistance, high-intensity interval, and concurrent exercise. Med. Sci. Sports Exerc. 2021, 53, 1708–1718. [Google Scholar] [CrossRef] [PubMed]
- Sawada, S.; Kon, M.; Wada, S.; Ushida, T.; Suzuki, K.; Akimoto, T. Profiling of circulating microRNAs after a bout of acute resistance exercise in humans. PLoS ONE 2013, 8, e70823. [Google Scholar] [CrossRef] [PubMed]
- Margolis, L.M.; Lessard, S.J.; Ezzyat, Y.; Fielding, R.A.; Rivas, D.A. Circulating microRNA are predictive of aging and acute adaptive response to resistance exercise in men. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 1319–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, R.F.; Markworth, J.F.; Aasen, K.M.M.; Zeng, N.; Cameron-Smith, D.; Mitchell, C.J. Acute resistance exercise modulates microRNA expression profiles: Combined tissue and circulatory targeted analyses. PLoS ONE 2017, 12, e0181594. [Google Scholar] [CrossRef]
- Cui, S.; Sun, B.; Yin, X.; Guo, X.; Chao, D.; Zhang, C.; Zhang, C.Y.; Chen, X.; Ma, J. Time-course responses of circulating microRNAs to three resistance training protocols in healthy young men. Sci. Rep. 2017, 7, 2203. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, S.R. Alterations of c-miRNA Expression from Whole-body Vibration and Resistance Exercise in Postmenopausal Women. Doctoral Dissertation, University of Oklahoma, Norman, Oklahoma, 2019. SHAREOK repository. Available online: https://hdl.handle.net/11244/321121 (accessed on 22 February 2021).
Figure 1.
Functions of Circulating MicroRNAs (miRNAs).

Figure 2.
MiR-21-5p regulation of osteoclastogenesis (Panel A) and miR-23a-3p regulation of osteoblast differentiation (Panel B). PDCD4—Programmed Cell Death Protein 4; FASL—Fas Ligand; RUNX2—Runt-Related Transcription Factor 2.
Figure 2.
MiR-21-5p regulation of osteoclastogenesis (Panel A) and miR-23a-3p regulation of osteoblast differentiation (Panel B). PDCD4—Programmed Cell Death Protein 4; FASL—Fas Ligand; RUNX2—Runt-Related Transcription Factor 2.

Figure 3.
Theoretical model for resistance exercise effects on bone formation. miRNAs—microRNAs; − downregulation; + upregulation.
Figure 3.
Theoretical model for resistance exercise effects on bone formation. miRNAs—microRNAs; − downregulation; + upregulation.

Table 1.
Select MiRNAs with Bone Cell Regulatory Effects.
| MiRNA | Target Gene | Pathway/Enzyme | Biological Effect |
|---|---|---|---|
| miR-21-5p | SMAD7 PDCD4 | BMP, TGF-β c-Fos | Promote OB differentiation Promote OC differentiation |
| miR-23a-3p | RUNX2 SMAD3 | TGF-β | Suppress OB differentiation Suppress OB differentiation |
| miR-100-5p | BMPR2 SMAD1 | BMP BMP | Suppress OB differentiation Suppress OB differentiation |
| miR-125b-5p | BMPR OSX | BMP RUNX2 | Suppress OB differentiation Suppress OB differentiation |
| miR-126-3p | MMP13 | Matrix Degeneration | Suppress OC differentiation |
| miR-133a-3p | RUNX2 CXCL11 | Rank | Suppress OB differentiation Promote OC differentiation |
| miR-148a-3p | KDM6B MAFB | TGF-β Rank | Suppress OB differentiation Promote OC differentiation |
Modified from [3,5]. OB—osteoblast; OC—osteoclast; SMAD7—Small Mothers Against ecapentaplegic 7; BMP—Bone Morphogenetic Protein; TGF-β—Transforming Growth Factor-Beta; DCD4—Programmed Cell Death Protein 4; RUNX2—Runt-related Transcription Factor 2; SMAD3—Small Mothers Against Decapentaplegic 3; BMPR2—BMP receptor type II; BMPR—BMP receptor; SMAD1—Small Mothers Against Decapentaplegic 1; OSX—Osterix; MMP13—Matrix Metalloproteinase 13; CXCL11—C-X-C Motif Chemokine Ligand 11; KDM6B—Lysine Demethylase 6B; AFB—V-maf Musculoaponeurotic Fibrosarcoma Oncogene Homolog B.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bemben, D.A.; Chen, Z.; Buchanan, S.R. Bone-Regulating MicroRNAs and Resistance Exercise: A Mini-Review. Osteology 2022, 2, 11-20. https://doi.org/10.3390/osteology2010002
AMA Style
Bemben DA, Chen Z, Buchanan SR. Bone-Regulating MicroRNAs and Resistance Exercise: A Mini-Review. Osteology. 2022; 2(1):11-20. https://doi.org/10.3390/osteology2010002
Chicago/Turabian StyleBemben, Debra A., Zhaojing Chen, and Samuel R. Buchanan. 2022. "Bone-Regulating MicroRNAs and Resistance Exercise: A Mini-Review" Osteology 2, no. 1: 11-20. https://doi.org/10.3390/osteology2010002