Race as Social Construct


Department of Psychology, University of Aarhus, Asylvej, DK-8660 Aarhus, Denmark
†
(1968–2007, retired).
Psych 2019, 1(1), 139-165; https://doi.org/10.3390/psych1010011
Submission received: 15 March 2019
/
Revised: 24 April 2019
/
Accepted: 25 April 2019
/
Published: 29 April 2019
(This article belongs to the Special Issue Beyond Thirty Years of Research on Race Differences in Cognitive Ability)
Abstract
:It is often claimed that race is a social construct and that scientists studying race differences are disruptive racists. The recent April 2018 “Race Issue” of the widely distributed National Geographic Magazine (NG) provided its millions of readers with a particularly illustrative example of this position. As discussions of race issues often recur, in both scientific and lay literature, stir considerable polemics, and have political, societal and human implications, we found it of both scientific and general interest to identify and dissect the following partly overlapping key contentions of the NG race issue magazine: (1) Samuel Morton’s studies of brain size is reprehensible racism (2) Race does not relate to geographic location, (3) Races do not exist as we are all equals and Africans, (4) Admixture and displacement erase race differences as soon as they appear, and (5) Race is only skin color deep. Also examined is the claim that Race does not matter. When analyzed within syllogistic formalism, each of the claims is found theoretically and empirically unsustainable, as Morton’s continuously evolving race position is misrepresented, race relates significantly to geography, we are far from equals, races have definitely not been erased, and race, whether self-reported or defined by ancestry, lineage, ecotype, species, or genes, is much more than skin color deep. Race matters vitally for people and societies. We conclude that important research on existing population differences is hurt when widely respected institutions such as NG mobilize their full authority in a massively circulated attempt to betray its scientific and public readership by systematically misrepresenting historical sources and scientific positions, shaming past scientists, and by selectively suppressing unwanted or unacceptable results–acts included as examples of academic fraud by the National Academy of Sciences (US, 1986). Any unqualified a priori denial of the formative evolutionary aspects of individual and population differences threatens to impede the recent promising research on effects of genome wide allelic associations, which would lames us in the vital quest to develop rational solutions to associated globally pressing societal problems.
Keywords:
race differences; evolution; IQ; brain size; racialism; fertility; anti-racism; ecotypes; lineages1. Introduction
Research on individual and group differences in intelligence—particularly on sex and race differences—frequently generates controversy and harsh critique of researchers who document them.
Rushton and Jensen’s (2005) [1] comprehensive review on Thirty years of research on race differences in cognitive ability is no exception. It was accused, among many other things, of being pseudoscience, because race is an arbitrary social construction (Tate and Audette, 2001) [2]. There are countless historical and also more recent examples of race critique (Gottfredson, 2010; Nyborg, 2003 [3,4]; Rushton and Jensen, 2005 [1]; Scarr, 1987; Sesardic, 2010; Woodley, Dutton, Figueredo, Carl, et al., 2018, online [5,6,7]). Fortunately, these authors have already provided insightful responses to the controversy, so this introduction can be limited to adding only a few more significant examples.
United Nation’s declaration(s) (UNESCO Publication 1950: later repeatedly revised) [8] thus banned biological race research, in an otherwise fully understandable reaction to unacceptable Nazi misuse around the Second World War. Ashley Montagu published a popular book (1942/1945/1946) [9], arguing that the folk or ordinary concept of race is just a social notion with no biological or genetic basis. Montagu was a student of determined race critics Frantz Boas at Columbia University, USA, and was also influenced by Ruth Benedict. Both commented on an early version of Montagu’s book, and Montagu co-authored the first version of the UNESCO publication. Another line of race research critique is based on the fear that demonstration of individual and group differences in IQ will challenge ideals of equality and solidarity. This concern is regularly articulated by progressive left-oriented academics, who feel urged to condemn non-social group differences research (Gottfredson, 1994, 2000; Gould, 1981, 1996; Gross and Levitt, 1994/1998; Lewontin, Rose, and Kamin, 1984) [10,11,12,13,14,15].
A further obstruction to differential behavioral research is the widespread misrepresentation of facts on individual and group differences in intelligence, both in science, in the popular literature, and the media. So widespread and detrimental is this source of distortion that it recently got its own name—the “Gould Effect” (Woodley, Dutton, Figueredo, Carl, et al., 2018, online [7]). The effect is based on Steven Jay Gould’s still widely held assumption that research on intelligence differences is deeply corrupted by its inherent racist, sexist, and elitist motivation. No intellectual person, perhaps aside from Richard Lewontin, has been more successful than Gould in demonizing differential-psychological and behavior genetics studies of intelligence through dishonest misrepresentation of data and theory (Alcock, 1998 [16]; Nyborg, 2003, particularly pp. 461–468 [4]; Lewis, Degusta, Meyer, Monge, et al., 2011) [17].
The late Robert Parry (2006/2018) [18] was the first to use the term controversialization to summarize adverse strategies used against IQ, sex, and race difference researchers. Controversialization takes place when opponents for political, ethical, or other reasons make a view questionable to themselves more controversial that it really is, and when these opponents include appeals to some purported social harm such a view will imply. “Fact” is often conflated with “value” in the controversialization process (Cofnas, 2016) [19].
National Geographic’s April 2018 “Race Issue” (NG; Goldberg, 2018) [20] serves as an illustrative example of global-scale controversialization of race research. In its distribution to 6.5 million readers, NG not only questions the existence of race differences but accuses all who find them anyway of being racists, who harm minorities and pervert public politics.
The present study examines the scientific veracity of the NG claims by applying Occam’s razor in a step-by-step analysis of NG’s position in terms of verbatim pro-et-contra dissection of statements such as ‘There’s No Scientific Basis for Race’—‘It’s a Made-Up Label’… ‘Races do not exist because we are equals’, ‘the concept of race is not grounded in genetics’, etc.
We also examine the validity of NG’s accusation of purported social harm through claims that race has been used by racists ‘… to define and separate people for millennia’… to the effect that … ‘To a disturbing extent, race still determines people’s perceptions, their opportunities, and their experiences’ …. ‘To the victims of racism, it’s small consolation to say that the category has no scientific basis’.
We finally consider the implications of NG characterizing any scientist, who report what he/she a priori should have known are non-existent race differences, as a racist who: (1) Is blinded by the illusion that race as such determines vital human trait differences, (2) who uncritically infers racial superiority, and (3) who has caused inexcusably social harm throughout centuries to individuals, minorities, and societies.
The scientific merit(s) of NG’s many indictments of named racist researcher is analyzed within the framework of five short pro-et-contra syllogisms, each dealing with central, if partly overlapping, aspect of the NG’s general and specific critique. The societal implications of existing race differences are finally discussed.
2. Syllogistic Analyses
First syllogism: Samuel Morton is a reprehensible model racist with a fixed definition of race.
2.1. Major Premises
- Samuel Morton is the father of scientific racism.
- (We “know” that the father of scienctific racism has THE correct understanding of race).
- Morton thinks that races represent separate acts of creation.
- Morton thinks races are ranked in a divine hierarchy.
- Morton did not think that races were closely related.
- Morton thinks that races has distinct characters, which:
- (a)
- Are immutable or “fixed” across generations (i.e., no transmutation, aka evolution).
- (b)
- Are homogenous or “fixed” (in these senses of fixation) across individuals within races.
2.2. Ergo
Morton is wrong about 3–6, and thus represent the opposite of reality. We can then say, given 1–2 and 3–6, that races don’t exist.
2.3. Minor Premises
- 7.
- “He wasn’t choosy about his suppliers” … “A particularly large-headed Dutchman … helped inflate Morton’s estimate of Caucasian capacities.”
- 8.
- “Morton’s ‘craniometry’ showed, he claimed, that whites, or ‘Caucasians’ were the most intelligent of the races … then East Asians, Southeast Asians, native Americans, and at the bottom, blacks”.
- 9.
- “…what science actually has to tell us about race is just the opposite of what Morton contended.”
- 10.
- “So many of the horrors of the past few centuries can be traced to the idea that one race is inferior to another…”
2.4. Conclusion
Morton was wrong and “…we still live with Morton’s legacy: Racial distinctions continue to shape our politics, our neighborhoods, and our sense of self.”
2.5. Contras to Points 1–6
Ad 1: It is not obvious why NG chose Morton as the father of scientific racism. Morton was predominantly working within conventional, pre-evolutionary Rayian—Linnaean systematics (for details, see Müller-Ville and Rheinberger, 2012) [21] where species with uniform formal designs were made by the Creator. Members of a species, except in the case of “natural varieties” such as sexes, differed (i.e., Varieties) only due to the direct effects of the environment. This view dominated for much for the 17th century and the early part of the 18th century, with few dissenters.
So why does NG champion Morton as THE “father of scientific racism”? In fact, Aristotle, Linnaeus, Buffon, Locke, Kant, Gobineau, Malthus and many others could have been equal or better choices. Add to this that Morton’s works did not matter much outside the US, his most productive period is essentially limited to 1839 to 1851, which is about a hundred years after Buffon (1749–1789) [22] and many more used the term “race” in a natural history perspective.
It appears that NG wanted to construe a straw-concept of scientific racism by choosing Morton as the Father of racism, a convenient picking because he was already falsely accused of fabricating an opportune racial hierarchical brain size rank order (see later).
Ad 2: Morton certainly did not hold THE one “correct” definition of race. In fact, he did not explicitly define the term “race” in his major works. Rather, he defined “species,” and he used the term “race” to mean “lineage.” “Species,” being lineages, were “races”, but not all “races” were species. In fact, Morton did not consider human “races” to be separate species in his earlier works, as discussed below. In this he joined a longstanding debate, where some authors saw race in a technical sense as designating constant varieties of a species and most polygenists used race in a more general sense to mean lineage, which could describe both species and constant varieties.
Commenting in particular on the role of Morton’s American school in the debate, natural historian De Quatrefages (1889) [23] writes:
“—In Europe, all botanists, all zoologists, from Linnaeus to de Candolle, from Buffon to Cuvier, and Geoffroy Saint-Hilaire, have employed them to designate very different things. If some have designated race by the expression hereditary variety, this difference in words does not in any way affect ideas... The distinction which exists all facts considered is always translated into language. Yet it is this distinction that the American school seems to forget entirely here. For her, there are no more races or varieties in nature; there are only species”.[Our ital.]
So why does NG take Morton’s use of the term “race,” when describing species-lineages, to be the only true meaning of “race” (lineage)? One conclusion comes to mind: NG promoted a straw-concept while remaining ignorant of Morton’s own (changing) position(s). But what is NG’s view now on species? Does NG also think of species as a social construct? We do not know.
Ad 3: We already know by now that “race” was neither a distinct category for Morton nor for his colleagues. We also know that his American School clearly preferred “species”, “types”, “groups”. In Types of Mankind (1857, pg. 80–81) [24] Morton writes: “…Lineage … where species are a type, we now ask where species come from?”
Are species created or selected? In Josiah Clark Nott’s (1854) book [25] “Types of Mankind, we find Prichard quoted for saying:
“The meaning attached to species, in natural history is very definite and intelligible. It includes only the following condition: namely, separate origin and distinction of race, evinced by a constant transmission of some characteristic peculiarity of organization.”
This definition accords in fact with the dominant Rayian-Linnaean Systematics of the time. It is now clear that points 3, 4, and 6 pertain to Morton and his colleagues’ concept of species, which is the conventional pre-evolutionary one—While species could be races (i.e., lineages), not all races were specifically distinct, that is species.
But then Morton (1847) [26] redefines his concept of “species” as a “primordial organic form” by asking …“What constitutes a species?” and answering that “I now submit a definition, which I hope will obviate at least some of the objections to which I have alluded: SPECIES—a primordial organic form...” How do we best understand what it means that forms are primordial, Morton asks: “If they can be traced back into “the night of time” as dissimilar as we see them now, is it not more reasonable to regard them as aboriginal than to suppose them to be mere accidental derivations of an isolated patriarchal stem of which we know nothing?”
“… I do not use it to imply that all divisions are derived from a single pair; on the contrary, I believe that they have originated from several, perhaps even from many pairs, which were adapted, from the beginning, to the varied localities they were designated to occupy... [which]... does not imply a common origin.
Two conclusions come easily to mind here: (1) Morton began to regard human races (lineages) as species, meaning that if we are to follow NG’s logic, it rather claims that Morton’s species represent the opposite of reality and that the concept of species is “made up”, and (2) Morton began to regard species as having multiple localities of origin where they were pre-adapted to diverse conditions.
Ad 4: Morton thus came to think that species were pre-adapted to their environment, as opposed to differing only due to environmental influences or ranked by a divine hierarchy. In fact, already in his November 1st. 1842 lecture [28], before which he embraced the view that human races represented different species (are different creations) he wrote:
“Man, regarded in his general character, is the same in every zone; he possesses the same general confirmation, and notwithstanding some striking diversities of organization, the whole human family is to be regarded as a single species. Yet, notwithstanding this approximation of mankind in essential and specific characteristics, I firmly believe that they were originally, or, in other words, before their dispersion into different latitudes, endowed with those varied traits of mind and body which alone could adapt them to their various allotments on the face of the earth. The more I have reflected on those diversities, the more I am confirmed in the conclusion, that they have not resulted from physical causes acting on constitutions originally the same, but that, on the contrary, there has been a primeval difference among men; not an accidental occurrence, but a part of that all-pervading design which has adapted man, in common with animals and plants, to the diverse conditions which form a necessary part of the economy of creation... Is it not more probably that the same Infinite power that conducted them, before their dispersal, to the varied physical circumstances with which they were henceforward to contend?… I apprehend that without such adaption, the patriarchal germs of our species would have been utterly destroyed in the effect to contend with those pestilential influences which appear to be inherent in certain localities on the surface of the earth.”
It subsequent years Morton adopted the view that races were created in separate regions (Morton, 1850) [29], but the logic was the same. Morton’s “economy of nature”—not “divine hierarchy”—was part of the 18th century pre-evolutionary attempt by many naturalists to reconcile the logic of “natural selection” with creation. The difference was that Morton was willing to apply it to humans at a time when many of the authorities of the time would violently oppose to the bare suggestion that each species, whether of plant or animal, did not originate[d] in a single birthplace.
Ad 5: Insofar as races were species, they were, by definition, not genealogically related. However, many pre-evolutionary natural scientists, including Morton, allowed for differences in magnitude of relation—in the sense of similarity–between species. The differences between species of a Genera could be large or small. Morton distinguished between “remote” and “proximate” species and suggested that proximate species are similar enough to produce fertile offspring.
To the extent origin is a problem, it is for the “species” concept, not the “race” (lineage) concept.
Ad 6: Did Morton believe race characteristics were necessarily distinctive, homogenous, and immutable? Again, the answer to this question is already provided in the above discussion of “species”. It is: No!
2.6. Discussion of the 6 Major Premises in the First Syllogistic Analysis
The overall implication of NG’s first 6 primary premises for criticizing Morton’s position is that race and species are made-up concepts. This urges us to examine NG’s own scientific position on race (and, by implication, on lineage, ancestry, admixture, or genetic population differences).
We find it-in rather plain words-on the one page before its critique of Morton and race. NG here informs its readers about the content and value of its own down-loadable DNA ancestry tool-kit called “GENO 2.0 Your DNA [30], Your Story”. The kit is sold by the following description on its webpage:
“Through decades of research and reporting, National Geographic seeks to answer and share fundamental questions about our collective past: how our ancestors migrated from our African homeland, adapted, and populated the Earth. With your help, we are writing this ever-evolving story. The Geno 2.0 test examines a unique collection of nearly 300,000 DNA identifiers, called “markers,” that have been specifically selected to provide unprecedented ancestry-relevant information … In addition, for all participants, we analyze a collection of more than 250,000 other ancestry-informative markers from across your entire genome to reveal the regional affiliations of your ancestry, offering insights into your ancestors who are not on a direct maternal or paternal line”.
The science behind the DNA kit is also presented there:
“Different populations carry distinct mutation, or genetic markers. Identifying and following the markers back through generations reveals a relationship shared by all humans, best conceptualized in the form of a genetic tree. Today, thousands of diverse branches, corresponding to unique human groups, can be followed backward to their common African root more than 100 millennia ago…. Your results give you an unprecedented view of your lineage. You will discover the migration paths your ancient ancestors followed hundreds and even thousands of years ago. You will also learn the details of your unique ancestral makeup—the biological and geographical components that make up who you are. What are the ingredients, and how much of a mixture is your own DNA recipe?”
This scientific position challenges the substantial difference between NG’s “populations”, which represent “branches” of the “human genetic tree” and one’s “lineage”, and the “races” concepts of natural history from the mid-eighteenth century, when evolutionary theory was adopted right until the mid-nineteenth century, when “race-denialism” accelerated.
To be sure, the early conceptions differed by species and individual variation (“varieties”) and many natural historians in the middle of 17th. century thought of ‘race’ as ‘lineage’ (as in French, conf. Nugent’s French dictionary from the 1700s): Lignee, sf, lineage; issue; race)—one could speak of “noble de race” or “nobility by lineage”.
When ‘race’ is not formally defined in a natural historian’s work prior to the middle of 20th century, we can generally replace this term with ‘lineage’-understood as a sequence or succession of individuals. According to Buffon and Cuvier’s widely known definition of “species” (“A succession of similar individuals which re-produce themselves”), species are races (lineages) in a non-technical sense. But, sometimes ‘race’ was defined in a technical sense, to refer to infraspecific lineages (in particular, so-called “constant varieties”). Thus, we could speak of different races, or lineages, of a species of dogs, horses, etc. (Doron, 2016) [31]. Others see the similarity, too.
Add to this that the NG article immediately succeeding the DNA Test advertisement (Forget race, ancestry is the real story—and it’s much more interesting”) explains:
“These six [pictures of Afro-Caucasian individuals] had their DNA tested with National Geographic’s kit (see below). These results indicate essentially the same “racial” heritage, in the percentages [of biogeographic ancestry] shown above. But their experiences are unique. Brenda Yurkoski (lower left) knew before the test—which names ancestral populations, not individuals …”
We then understand that even within the same National Geographic “Race Issue” magazine, we find people thinking along similar scientific lines, such that race is identified with lineage and biogeographic ancestry.
Generally, it goes without saying that few natural scientists between the middle of the 17th century, when it was possible to speak about race as a natural scientific concept, would not have called differentially adapted the “branches” “of “the human genetic tree,” lineages, or “ancestral populations”, or “races”, at least if they were visibly distinct.
If we assume for the sake of argument that the DNA GENO 2.0 kit defines NG’s honest scientific position, then in which respects does it differ from that of its editor, Susan Goldberg, who motivated and launched the Race Issue attack? In fact, not much.
She too acknowledges that human variation exists by arguing “That race is a human construction doesn’t mean that we don’t fall into different groups or there’s no variation.” She further acknowledges the existence of variation between human lineage-populations or descent groups by saying that:
“Sometimes it’s clear that natural selection has favored a mutation, but it’s not clear why. Such is the case with a variant of a gene call EDAR (pronounced ee-dar). Most people of East Asian and Native American ancestry possess at least one copy of the variant, many possess two. But it’s rare among people of African and European descent.”
Interestingly, Goldberg also keeps in line with the typical 18th century conception, according to which lineages and descent groups were seen as branches, as can be seen when she says that:
“Studies of this genetic diversity have allowed scientists to reconstruct a kind of family tree of human populations… the Khoe-San, who now live in southern Africa, represent one of the oldest branches of the human family tree.”
Undoubtedly, Goldberg’s conception of groups as branches, which, by the way, are more discrete and thus more stereotypically racialist than most modern stances, must have left knowledgeable readers of her Race Issue in doubt about NG’s precise scientific point of view. Unfortunately, there are more reasons for confusion.
NG thus refers to Reich’s (2018) book [32], which discusses, among other important topics, the nature of phylogenic networks. Here Reich argues that:
“…while three is a good analogy for the relationships among species–because species rarely interbreed …. It is a dangerous analogy for human population …” because “… great mixtures of highly divergent population have occurred repeatedly. Instead of a tree, a better metaphor may be a trellis, branching and remixing far back into the past... This is greater than the separation times of the most distantly related human lineages today…”
We now see that, in contrast to Goldberg’s conception of stereotypically racialist branches, Reich prefers the term “lineage” and is in fact skeptical of her tree model.
But then again, Reich’s use of the term lineage lines up with the customary translation of lineage: race. Of course, races (lineages) were conceptualized as phylogenetic networks too, as illustrated by Buffon (1749–1789) [22] and others. Now, if Reich and Goldberg agree with what most people up until the middle of the 20th century called “races”, does this mean that they do not think races differ? Not at all. In addition to NG’s scientific position for selling its DNA kit, Reich writes in his 2018 book [32]:
“But “ancestry” is not a euphemism, nor is it synonymous with “race.” Instead, the term is born of an urgent need to come up with a precise language to discuss genetic differences among people at a time when scientific developments have finally provided the tools to detect them. It is now undeniable that there are nontrivial average genetic differences across populations in multiple traits, and the race vocabulary is too ill-defined and too loaded with historical baggage to be helpful. If we continue to use it, we will not be able to escape the current debate, which is mired in an argument between two indefensible positions. On the one side there are beliefs about the nature of the differences that are grounded in bigotry and have little basis in reality. On the other side there is the idea that any biological differences among populations are so modest that as a matter of social policy they can be ignored and papered over. It is time to move on from this paralyzing false dichotomy and to figure out what the genome is actually telling us... But such a statement is wrongheaded as if we were to randomly pick two people living in the world today, we would find that many of the population lineages contributing to them have been isolated from each other for long enough that there has been ample opportunity for substantial average biological differences to arise between them.”
The critical reader may now also begin to wonder whether replacing a term—because it is “too loaded with historical baggage”—is not a euphemism, regardless of what is otherwise claimed? Confusion rules again as to the precise nature of the argument. Does the term “race” means/meant something radically or substantially different? If not, the reader witnesses an attempt to control the language and exclude— “cutting off the baggage”—the dense nomological network surrounding the term “race”. Too many options are still left open.
2.7. Contras to Points 7–10
Ad 7: Neither do accusations of racism account as scientifically valid counter-arguments against Morton’s empirical brain measures, nor can indirect guilt by association to slavery be accepted as a scientifically valid counterargument. They are primarily vilifications.
Ad 8: Morton aimed at still larger numbers of still more diverse crania, because this improves the power of analysis. It is no error to include large sized (Dutch or other) crania unless this reflects intended sample bias. In fact, Morton had the largest and most diverse skull collection available at the time (Morton, 1845) [24]. Add to this that Rushton (1992) [33] later found similar significant cranial differences and broke them down by sex and race (also see Rushton and Rushton, 2003; Symmons, 1942 [34,35]; and below).
Ad 9 and 10: Morton’s early racial ranking of crania has a long history of recent re-examinations, including being criticized by Steven Jay Gould (1978 [36]; 1981/1996 [12,13]) and defended independently by Lewis, Degusta, Meyer, Monge, et al. (2011) [17] and by Wade (2011) [37]. The latter two studies were in turn questioned in an Editorial (2011) [38], and then Weisberg (2014) [39] partially supported Gould’s accusations.
NG fails to call attention to these selected examples of reasoned doubt either way, and instead turns one-sidedly against a defenseless scientist such as Samuel Morton—long gone.
2.8. Discussion of the 4 Minor Premises in the First Syllogistic Analysis
NG fails to refer to independent available data confirming that North East Asians have—on average—the largest skull average of all races studied (Lynn, 2006/2015) [40]. NG fails to inform the reader about documented geographic differences in prehistoric skull sizes, including an estimated increase in skull volume of 2.5 cm3 with each degree latitude (Smith and Beals, 1990) [41], after measuring close to 20.000 crania from 87 populations, categorizing them into 10 races, and finding significant race differences (Beals, Smith, and Dodd, 1984) [42]. NG finally fails to call attention to Cold Winter theory (Lynn, 2006/2015, 2008) [40,43], which on theoretical ground predicts, and in practice finds, racial brain size and IQ differences as a reflection of climate, i.e., the colder, the larger or higher.
Unqualified racial slur has, without doubt, brought havoc through centuries and have caused some individuals and groups to believe in their own higher standing, but to negate empirically based racial hierarchies or embrace all race difference researcher as guilty raises the need for spotting the difference between slur and reality.
2.9. Summary of the First Syllogistic Analysis
NG has created a straw-concept by singling out Morton as THE father of scientific racism. Many other eminent historical researchers would qualify as well or better, given NG’s premises, and most of them reached beyond the limited impact Morton had outside the US. NG muddies the already controversial concept of “race” in several ways, including being unclear about technical (e.g., hereditary varieties) and more general usages (e.g., lineage) and more general usages (e.g., hereditary variant, species, types, groups). NG pays no attention to the fact that race was not a distinct category for Morton, who used "race" to refer to lineage, include both infraspecific and species-lineages, and who repeatedly revised his definition of species.
Still worse, even NG editor Goldberg herself admits to human genetic variation in a stereotypically enhanced racialist way, and Reich, while criticizing race, fails to see that his favored lineage easily translates into race.
Obviously, NG’s claim—that Morton’s race differences in brain size (skull size, brain capacity, volume, or however defined) do not exists—is misleading. Morton’s findings are in fact supported by empirical evidence from such varied sources as (1) early fossil findings (Beals, Smith and Dodd, 1984) [42]; Smith and Beals, (1990) [41], and (2) Rushton’s current findings (1988a, 1988b [44,45], 1992 [33], 1997, 2000 [46,47]; Rushton and Osborne, 1995 [48]). No doubt, Morton misclassified the “cold” North East Asian average skull size by ranking it lower than less cold European ecotypes, perhaps due to incomplete series of specimens. The effect of temperature refers here to the observation that ecotype skulls found in prehistorically colder econiches are in general larger than those found in warm econiches (Smith and Beals, 1990 [41]).
We are already at this early stage of analysis led to the conclusion that a more suitable title for NG’s whole Race Issue paper would be: “National Geographic’s Mischaracterization of Morton”, even without calling for a full testimony of all aspects of the critique and Morton’s many letters, lecture notes, and entire works, spanning the period 1839–1857.
Doing so is obviously beyond the scope of the present paper, but the above evidence suffices to show that NG systematically misrepresents Morton’s and other’s historical and current sources of differences in brain size and selectively suppresses unwanted or unacceptable results. Moreover, by making Samuel Morton responsible for centuries of horror after publishing his empirical data on race differences in skull size, NG provides examples of ad hominem accusation and guilt by association, based on hearsay and feelings of white shame (see later). NG fails in providing the needed scientific counter-evidence in this context.
In a word, NG’s many accusations—including that Morton is a racist harming minorities and public policy—are scientifically unbecoming.
3. Second Syllogism: Race does not Relate to Geographic Location
NG claims (with Reich, 2018 [32]) that geographic race differences do not exist, because “What the genetics shows is that mixture and displacement have happened again and again”.
Where the first syllogistic analysis disproved the existential version of this claim in term of easily observable race or lineage differences, the second syllogistic analysis aims to illustrate that there are pivotal geographic race/lineage differences with profound societal effects. Just one example suffices to prove this.
3.1. Main Premise
- 1.
- “There are no fixed traits associated with specific geographic locations …” because …
3.1.1. Minor Premise
- 2.
- “… as often as isolation has created differences among populations, migration and mixing have blurred or erased them.”
3.1.2. Ergo
- 3.
- “… our pictures of past ‘racial structures’ are almost always wrong” and harmful.
3.2. Contras
Ad 1. Neither of these claims are true. Let’s take intelligence (IQ) as example, because: (1) It is relatively fixed, (i.e., difficult to change rapidly by environmental means, (2) It is individually and generationally stable over time, (3) It—and many of its covariant traits—differs significantly among populations (e.g., Average Income and Democracy Index), now and then (Becker, 2019 [49]; Lynn, 2006/2015 [40], 2008 [43]; Lynn and Vanhanen, 2002, 2012 [50,51]). Please note that the short IQ term represents the latent general intelligence factor g, pointing to the ability to deal with complexity irrespective of the indicator (For details, see Spearman, 1927; Jensen, 1998 [52,53]).
Figure 1 illustrates historic geographical differences in IQ.
The colors in Figure 1 mirrors the estimated global distribution of IQ for indigenous populations around the 14th Century (Lynn, 2006/2015 [40]) and conform by and large to predictions from Cold Winter theory, that is, the colder temperature, the higher the IQ.
This early IQ distribution differ in several ways from the modern IQ distribution illustrated in Figure 2.
Marked IQ color differences between maps thus confirm the observation that massive displacements have taken place historically, reflecting, among other things, the substantial North European migration into previously low-IQ geographic areas such as Australia, North America, and EuAsia. There are two major points to take home from this: (1) Marked geographical IQ differences are still clearly identifiable on a global scale despite massive displacement and admixture, (2) The about 30 IQ point difference between central Africa and Northern Europe still persists after many centuries.
With respect to co-variates, average ecological population differences in IQ have been shown to relate to vital average differences in societal sophistication (e.g., Lynn and Vanhanen, 2002, 2012 [50,51]). The suggested partway is as follows: The cold econiche conditions selected harshly for evolution of the general intelligence hierarchical factor g—defined as a complex latent brain factor. This g-factor appears as the key node variable in the center of a factor matrix, where it is (one-way causally?) related to multiple individual and societal educational, economic, and societal behavioral outcome measures (Herrnstein and Murray, 1994; Jensen, 1998 [53,54]).
The repeatedly confirmed existence of globally distributed significant IQ differences and the examination of the many significant correlational network relations in the hierarchical factor matrix model (e.g., Gottfredson, 1997a, 1997b, 2000 [11,55,56]) negate NG’s claim of no significant global race/lineage differences in IQ and societal sophistication.
Ad 2. The claim that migration and mixing has blurred, or erased, all race differences is accordingly patently false. This applies to the vital case of IQ but extends to race differences in medicine and nutritional response, and to other areas. Admittedly, draught, natural catastrophes, mountains, rivers, seas, and migration (the latter either based on curiosity, aggression, hunger, or other conditions) might temporarily or permanently have separated or shuffled groups of people around at a geographical scale, as suggested by the color differences between Figure 1 and Figure 2. But these events, combined with admixture and displacement, have evidently not been sufficient for eradicating north-south differences in, say, brain size at a scale covering about 100 cm3, and a latitudinal IQ difference scaling about 30 points (Lynn, 2006/2015; Rushton, 2000 [40,47]).
Moreover, effects of this selective latitudinal geo-bio-climatic gradient may reach far back in prehistoric times, as suggested by several lines of evidence. Behavior genetic studies consistently document that IQ is a brain-size related trait (r = 0.30–0.45), with a variation in age-dependent heritability estimates ranging from about 20% in childhood, to more than 50% in adolescence, and even rising to about 80+% in adulthood, the so-called Wilson effect (Bouchard, 2013; Plomin, DeFries, Knopik, and Neiderhiser, 2016 [57,58]). Given that skull size correlates moderately with IQ and increases by 2.5 cm3 per degree latitude, both in fossilized 10,000–1,200,000 years old crania (Beals, Smith and Dodd, 1984; Smith and Beals, 1990 [41,42]), and in contemporary crania, we have reason to expect—by analogy and in accordance with Cold Winter theory (Lynn, 2006/2015 [40])—that these latitudinal trait differences have existed throughout this period. However, they did not originally give rise to higher levels of pre-historic democracy or civilization such as the modern forms (Murray, 2003 [59]), most likely because early populations were too small and scattered to establish such complex collective societal phenomena, which require extended specialization of functions within society.
Another reason is that the groups of people migrating out of high IQ Europe centuries ago (to the US, Canada, Australia etc.) even today retain the average of their country-of-origin IQ—across multiple generations and over widely varying conditions. Adaption to new econiche conditions appears to take millennia in order to establish new genotypic and phenotypic normal distributions.
Ad 3. The above-mentioned evidence makes NG losing the license for blaming “our pictures of past racial structures’” for harmful consequences of racial trait distribution. Nobody is to be blamed for Darwinian adaptation to Eons of unadorned evolutionary econiche selection.
3.3. Discussion and Conclusions of Syllogism 2 Analysis
Ample direct and indirect evidence confirm that NG’s claim that considerable genetic displacement and mixture has eradicated geographical race differences is not sustainable.
Important differences associate significantly to geographic locations, confirming that migration and mixing have neither blurred nor erased, but certainly changed some of them. It follows that our pictures of past ‘racial structures’ are not always wrong and harmful.
It has recently become economically and technically possible to do genome wide association screenings of individuals and large groups to identify, among many other things, positive or negative structural gene variation for physiological traits such as IQ (e.g., Plomin and von Stumm, 2018 [60]). This scientific break-through lets us expect that it is only a matter of time before geographically differentiated patterned distributions of coupled gene variants will be identified to further support the existence of biologically based individual and group differences. We now know that populations differ in frequency of SNPs (single nuclear proteonids) associated with intelligence (e.g., Piffer, 2013, 2015 [61,62]), in types of y-DNA chromosomal haplogroups (Rindermann, Woodley and Strafford, 2012 [63]), and in genetic distance measures (León and Burga-León, 2015; Becker and Rindermann, 2016 [64,65]). Research in these areas virtually explodes right now, and the specific and generalized results will most likely not only constitute ultimate counter-arguments against claim of gene-free population non-variance for important phenotypic and societal traits, but also change our view radically on the molecular basis for many individual and population difference (Nyborg, 1998 [66]).
4. Third Syllogism: Races do not Exist: We are Equals and Africans
The ideal of sex-and-race-equality has long dominated in wide academic circles (Gottfredson, 1994, 1997, 2000 [10,11,55,56]), along with widely acknowledged theoretical cultural-Marxist social-constructivist positions (Gross and Levitt, 1994/1998 [14]; Nyborg, 2003, 2011 [4,67]), according to whom Man is made by society
NG is no exception to defending ideas of equality, even in their strong anti-genetic form. It thus refers to the “fact” that “… genetic research has revealed two deep truths about people”.
4.1. Major Premises
- “…all humans are closely related.”
- “In a very real sense, all people alive today are Africans.”
- “Genetic diversity in Africa is much larger than outside this continent.”
- “Because they [migrants] were just a small subset of Africa’s population, the migrants took with them only a fraction of its genetic diversity.”
4.2. Minor Premises
- 5.
- Admittedly, “… the longer two groups are separated, the more distinctive tweaks [mutations] they will acquire”, BUT …
- 6.
- “The concept of race has no genetic or scientific basis.” (NG here refers to a Craig Venter statement at a White House meeting, June 2000; see later).
4.3. Ergo
- 7.
- “Science tells us there is no genetic or scientific basis for race. Races do not exist because we are [all] equals.”
4.4. Proet Contra
Ad 1–6. Ample empirical evidence supports NG’s statements that all humans are closely related, that all alive today most likely are of African origin, that Africans differ more genetically than those outside that continent, that migrants left Africa with only a fraction of the large genetic diversity found there, due to genetic Bottle-necks and serial founder effect (e.g., Ramachandran, Deshpande, Roseman, Rosenberg, et al. 2005 [68]). It is also true that the longer groups are separated, the larger will be the number of distinctive mutations—even if this speaks in favor of increased population/racial/lineage differentiation and, in fact, suggests that race has a genetic basis.
But the crucial counter-point here is that neither can any of the single premises 1-6, nor any of their combinations, be accepted as scientific proof that race differences have no genetic basis. Neither do any of them imply that we all—or the races—are equal(s). Moreover, the previous illustrations of the existence of more or less permanent, systematic, geo-bio-climatic Eco-type trait differentials will be hard to explain except in terms of some underlying genetic population variation.
We here find another of NG’s more serious problems: It has fallen prey to the classical Lewontinean Fallacy. Lewontin (1972) [69], a geneticist from Harvard University, thus maintained that human racial classification “…is of no social value, destructive, and virtually of no genetic or taxonomic significance. He suggested that “…All things considered, then, the 6.3% of human diversity assignable to race is about right … but ‘The largest part by far of human variation [is] being accounted for by the differences between individuals‘ … so … ‘no justification can be offered‘ … for ‘human racial classification.”
To this Edwards (2003) [70] dryly responded that if ever so small genetic variation correlates significantly with some race difference, then it certainly have taxonomic significance.
Moreover, despite the fact that we in an abstract historical sense all are Africans, a more pertinent question is how closely this statement relates to degree of genetic admixture, in the sense of how many important gene variants and related phenotypic traits we do share with our remote African forefathers. It is also worth noting that most of the dwindling out-of-Africa genetic variation concerns non-causal variants, also called Junk DNA.
The final blow to NG’s Lewontinean reasoning is delivered by the recent observations of average differences in patterned polygenic based traits between individual and populations. The human genome contains close to 3.1 billion base pairs that do not differ from one person to another, that is, make no expressive difference, but then there remains about 56 million base pair variants, including some very rare forms.
Let us for the sake of argument accept Lewontin’s own estimate that about 6.3% of race differences have a genetic basis. This leaves us with some 3.5 million gene variants which could potentially differ among peoples/races/ancestries/lineages/species/ecotypes. A window that size provides ample genetic space for explaining even large differences. Moreover, variation in just one base pair can shift the entire DNA reading frame and consequently make a world of phenotypic differences. In addition, polygenic variation may make population traits differ significantly as a function of the allelic balance among plus- and minus variants. Obviously, Lewontin and others at his time did not know that most of the genetic information that makes for (race) differences are discernible only in the correlational structure of widely distributed allelic effects (e.g., Edwards, 2003 [70]).
The new genetics is beginning to harvest this vital analytic information and will without reasonably doubt change our view on the role of genes in individual and population differences. Frequency differences in polygene traits have already been shown to relate to significant geographic phenotypic differences in the mean value of physical traits such as height and body mass (Robinson, Hemani, Medina-Gomez, Mezzavilla, et al., 2015 [71]). These researchers looked for genetic variance for body height and BMI in 250,000 persons across 14 European countries and found that captured additive variance accounted for 24% variation in body height and 8% in Body Mass. Novembre, Johnson, Bryc, Kutalik, et al. (2008) [72] noted that mutations differ as a function of geographic and population circumstances. They warned that this raises the need to control for population stratification, but the jury is still out on this. Okbay, Jonathan, Beauchamp, Fontana, et al. (2016) [73] mapped the educational level of 400,000 mostly European individuals, (over?)-controlled for socio-economic factors, and found an overrepresentation of 74 gene variants affecting neuronal development, in well-educated Europeans as compared to fewer in individual with less education. Educability and IQ are arguable physiological (Spearman, 1927) [52] brain size related traits with a stability throughout life equal to those for body height and weight (Jensen, 1998) [53]. Even one gene on the Y chromosome can make a world of differences: It is just 886 base pair long and counts for only about 0.0016% of the total genome, but nevertheless largely determines whether male or female development unfold.
Ad 7. Both old and new scientific evidence thus proves that NG misinforms its readers when it claims that science “tells us” that there is no genetic or scientific basis for race and that we are all equals”.
The ensuing conclusion is inevitable: NG, either out of carelessness or deliberately (the latter is more likely, see later) suppresses or misrepresents significant evidence confirming that physical race differences do exist and proves beyond reasonable doubt that most of us are far from equals (Gottfredson, 1994, 2000 [10,11]). Peoples north of Sahara are admittedly all Africans, but only in a remote prehistorical sense; they certainly differ genetically and phenotypically from their African forefathers in any meaningful sense of that word.
4.5. Discussion of Syllogism 3
We may learn from NG’s critique, that Morton developed inaccurate “made up” concepts for self-serving reasons, that Morton’s thinking was the opposite of reality and a product of his bias, that people such as Morton intentionally developed the “science of race” to suit their own purposes, and that all this add up to considering Morton and scientists like him as self-servingly biased, and since they were all wrong, we can say that race was “made up” or “invented”.
To answer this critique fairly, we have to place Morton in a historical perspective. He was a conventional pre-evolutionary scientist but did not apply this framework unconditionally like most other contemporaries. Obviously, this implied creationism, which is wrong, but it also saw species as generative units. He considered the possibility of natural selection and the need for adaptation to the “…pestilential influences which appear to be inherent in certain localities on the surface of the earth.” which would otherwise have destroyed our species’ patriarchal germs. Morton acknowledged the intergenerationally stability (constancy) of races as they had existed since the dawn of time. In contradistinction to many moral natural historians of the time, Morton neither believed that the physical environment had produced race differences directly, nor were of a different nature than those which cause recognized specific differences.
Importantly, Morton’s thinking evolved over time, as described by his protégés Nott and Gliddon (1854) [25]. To them, Morton at first accepted the unity doctrine and taught it for long in good faith but was then led by the conspicuous wide ethnic diversities to question the popular doctrine and the notion of a common origin of all races. He seems to have abandoned the idea that ordinary physical influences could explain existing diversities within such short time frame, not by denying entirely the possibility as some of his contemporaries did, but he eventually came to the conclusion that “… the organic characters of the people themselves, through all their endless ramifications of tribes and nations, prove them to belong to one and the same race, and that this race is distinct from all others.” (1842, 1844, p. 35.) [28].”
In other words, Morton came to think that “… differences between certain human races was specific (of a species type)”, and he applied natural historian rules to humans in an unbiased way, contra many of his contemporaries. He neither distorted data nor made up unprincipled exceptions based on questionable claims, nor advanced positions for moral, ideological, or other reasons.
This is illustrated in a reply to Rev. John Bachman (Morton, 1850) [29] where he confesses that:
“I have never felt the slightest hesitation in investigating the facts of Nature—well knowing that “truth will never conflict with itself,” no matter how diversified so ever may be the points in which we view it. I am far, however, from desiring to make startling propositions to ignorant minds; but, as I address myself, in this, as in former instances, to educated persons, I cannot conceive that evil consequences will any more result than would follow scientific investigations in astronomy, geology and chronology — each one of which has, in its turn, contended against the inveterate repositions, not only of the ignorant, but of many otherwise learned and enlightened individuals … I have never swerved, viz.: that the diversities existing among the different human families have not been acquired; or, in other words, are not the result of climate, locality, food, and other physical agents, but have existed aborigine: and, in the early period of my investigation, I was content, as elsewhere expressed, to suppose that the distinctive characteristics of the several races might have been marked upon the immediate family of Adam. More light on this interesting question has compelled me to change my opinion. I was not aware, however, that this was so great a dereliction of propriety as Dr. Bachman considers it.”
Now is a good time for the learned NG reader ask: Whom are the ideologically biased ones?
5. Fourth Syllogism: Admixture and Displacement Have Erased All Race Differences
5.1. Major Premises
- (Race implies unadmixed groups between which there are fixed—“fix”, in the sense of fixation index—traits.
- (From Reich (2018) [32] race implies “primeval” groups...separated tens of thousands of years ago”.
- Genetics shows that mixture and displacement have happened again and again”... and … as a result “Differences have been blurred or erased”.
- Thus, “there are no fixed traits associated with specific geographic locations…”
- And “…our pictures of past ‘racial structures’ are almost always wrong” and harmful.
Ergo
Since human descent groups are mixed and do not exhibit fixed trait differences and since there are no 10-thousand-year-old primeval groups, there are no races.
5.2. Contras
Ad 1. The idea of unadmixed races with fixed differences comes from pre-evolutionary primary species thinking and contrasts modern views.
Ad 2. NH cites Reich for defining races as primeval groups separated long ago, so we better examine the arguments for this in his recent 2018 book [32]:
“Today, many people assume that humans can be grouped biologically into “primeval” groups, corresponding to our notion of “races,” whose origins are populations that separated tens of thousands of years ago. But this long-held view about “race” has just in the last few years been proven wrong—and the critique of concepts of race that the new data provide is very different from the classic one that has been developed by anthropologists over the last hundred years … Most of today’s populations are not exclusive descendants of the populations that lived in the same locations ten thousand years ago.”
So, the idea is that race implied relatively immobile, 10-thousand-year-old groups.18th century natural historians were quite well aware of this migration and admixture. As seen in Morton’s work and the work of his colleagues, the American school was no less so. This was one of the reasons Morton spent so much time researching hybridity among species.
Here is a typical passage Reich chose to adapt from Morton’s Types of Mankind (p. 316) [24]:
“Europe was successfully invaded by the Celtic, Teutonic, and Slavonic races. The Celtic migration is of extreme antiquity, yet there can be no question that they displaced pre- existing tribes. Among the latter may be mentioned the Iberians of Spain, what are represented by a fragment of their race—the Basque or Euskaldunes of Biscany” … The Indostanic family.—No part of the world presents a greater diversity of human races than the country which bears the collective name of India. Exotic nations have repeatedly conquered that unfortunate region, and to a certain degree amalgamated with its primitive inhabitants... That the peninsular India was originally peopled, at least, in part by races of very dark and even black complexion, is beyond question. These people are stigmatized as Barbarians by their conquerors, the Ayras.”
This traditionally account of “racial structure” seemingly differs little from Reich’s own model of dark-skinned Dravidians being invaded from the north. As for Reich’s claim that the “long-held view” was that races were tens of thousands of years separated, we suspect that the young earth creationists, who dominated the pre-evolutionary discourse on human race, would have vigorously disagreed. The point being here is that there was no “long-held view,” but rather an ever shifting one on the time of origin and divergence.
Ad 3. Since Morton is the “bad-man-out” in NG’s race quarrel (i.e., he don’t see that mixture and displacement have blurred or erased all race differences), we better quote his own discussion on human races (at this time still conceptualized as varieties of a species), expressed in On the varieties of the human species of Crania Americana (1839) [74]:
“Such appear to have been the primitive distinctions among men: but hostile invasions, the migratory habits of some tribes, and the casual dispersions of others into remote localities, have a constant tendency to confound these peculiarities; and the proximity of two races has uniformly given rise to an intermediate variety, partaking of the characteristics of both, without being identical with either: these are called mixed races.”
Despite its relevance, Reich chose not to take up this passage.
Ad 4. The prevalent idea during the 17th and 18th centuries were that human races did not qualify as species with their Fixed traits, an idea that has transferred relatively unchanged to current Behavior Genetics, which often relies on Blumenbach’s early 1779 5-race partitioning.
The early naturalists differed more on the question of the formation of races and which lineages coalesced into which, and using this as a cardinal point that “population lineages” differ totally from “races” makes little sense.
Ad 5. It is indeed ironic to find that NG’s claim, that our pictures of past racial structures are wrong and harmful, is long ago drowned in massive previous counter-evidence, but still sails strong in NG’s uncompromising anti-racialist boat.
5.3. Discussion of the Fourth Syllogistic Analysis
NG refers to Craig Venter’s international authority in a straightforward way, something like: We must accept what authority Venter says and he says that “The concept of race has no genetic or scientific basis”. Ergo, Craig Venter said race has no genetic or scientific basis, so it has not!
However, falsifiable hypotheses, sound methodology, and solid data should always trump authority and one-liners in science. Given these premises, we better examine the science behind Venter’s brief statement at the White House meeting.
Venter (2000) [75] explains:
“The method used by Celera has determined the genetic code of five individuals. We have sequenced the genome of three females and two males, who have identified themselves as Hispanic, Asian, Caucasian or African American. We did this sampling not in an exclusionary way, but out of respect for the diversity that is America, and to help illustrate that the concept of race has no genetic or scientific basis. In the five Celera genomes, there is no way to tell one ethnicity from another. Society and medicine treat us all as members of populations, where as individuals we are all unique, and population statistics do not apply.”
These statements raise several questions, all begging for an answer. First, given that race has no genetic or scientific basis, we apparently still can tell members of different self- identified racial and ethnic groups apart? Second, given a sample of just 5 individuals (Hispanic, Caucasian, Asian, and African American) we can distinguish and safely conclude in general, with a sufficiently high degree of probability, that race has no genetic or scientific basis? But does self-identified race not correspond to actual genealogy here?
With no proper answers to these questions, we may conclude that NG cited Venter’s statement more out of authority than based on science, as a very large number of huge-N samples contradict Venter’s statement flatly.
Moreover, Venter actually stated that “In the five Celera genomes, there is no way to tell one ethnicity from another”. Tal (2013) [76] has the explanation: “The paradoxical results is most likely an artifact of the high error rate and low coverage in the Watson’s SNP calling.” In fact, the probability of finding genetic race distance differences among people in Venter’s sample with N = 5 is extremely low, and we know today that this probability increases with the number of genetic markers noted and sample size (e.g., Becker and Rindermann, 2016 [65]; Plomin and v. Stumm, 2018 [60]; Visscher, Wray, Zhang, Sklar, et al., 2017) [77].
As before, NG not only appeals to authority but also neglects available solid data contradicting its claim that race and genetic differences do not exist.
We can further trace NG’s many more convoluted ways to unfold it race denial thesis. Truly we are all African, it says, but if race is real, we share a common descent, because we descend from Africa. Then again, if races exist, they would be “homogenous”, but there is no homogenous African race. We are led to the conclusion that since lineage populations share a common descent and are not homogenous, they are not “races”, but this is correct only if “race” means species in the pre-evolutionary sense of the term. Then again, according to NG’s logic, species is made up concept. This is low level confusion.
With respect to the recurring question of the origin of races, the most common traditional idea was that we all descend from the Middle East—as Caucasians—with biblical reference to the inferred location of the Garden of Eden. However, few, if any, today would consider races as non-hybridized pre-evolutionary species. A modern scientist would rather find it easier to subscribe to the fact that African lineages are more closely related to one another than to members of north-of-Saharan lineages, due mainly to a marked loss of genetic variance as relatively small samples with African genotypes migrated out of African (Founder effects), but also because of admixture.
6. Fifth Syllogism: Race is only Skin Color Deep
The results of the previous formal syllogistic analyses suggest that race/lineage/ecotype is more skin-color-deep, than NG claims. How deep, then?
6.1. Major Premises
- “When people speak about race, usually they seem to be referring to skin color and, at the same time, to something more than skin color.”
- “This is the legacy of people such as Morton, who developed the “science” of race to suit his own prejudices and got the actual science totally wrong.”
- “Science today tells us that the visible differences between peoples are accidents of history. They reflect how our ancestors dealt with sun exposure, and not much else.”
- There is no homogenous African race.
6.2. Ergo
Since race is only based on skin color, it is made up by racists.
6.3. Contra
Ad 1. What could be “deeper” than skin color? “Morph” and “Variant” form” are certainly not “made up” concepts, but they could admittedly be taken to mean interpopulation variants in single traits (e.g., a color morph). Then again, few scientists today would interpret available data to suggest that “race” just meant “morph”, or “variant”, or “form”.
Ad 2. If race just meant skin color, why would anybody then refer to race (meaning lineage)? Why would Buffon and other early naturalist include albinos within the “white” race, if all they wanted to illustrate is what race is not?
A final question: Why does NG tout “Forget race, ancestry is the real story—and it’s much more interesting” when advertising its DNA Test, when it at the same time acknowledges that “racial heritage” and ancestral population heritage are essentially the same thing?
This convoluted way of arguing reflects incoherent thinking apt to camouflage the empirical world of facts.
Ad 3. Are population differences just historical or random accidents? The fact that Morton, Buffon, Blumenbach, Cuvier, Darwin and many others grouped peoples by gestalt similarity suggest otherwise and scientists such as Morton strived hard to establish a “natural” system. This is for example reflected in Morton’s Crania Americana (1839) [74], where he grouped “mankind by physical and ethnographic methods“…“into great divisions characterized by similarity of exterior conformation”.
Ad 4. It was the conventional monogenist position that racial differences were caused by sun exposure, which evoked something akin to epigenetics to account for the “inheritance” of differences.
6.4. General Discussion of the Syllogistic Analyses
Historic and current evidence both indicate that skin color associates significantly with much more than just accidental sun exposure. Already the classical Greek and Roman scholars discussed the significant associations between behavior and the color of peoples’ skin, mostly, however, at the phenomenological level. Then Blumenbach’s (1779) [78] systematization of humankind into five physiological types began to inspire much later research, even if his categorization was eventually revised numerically and otherwise several times (conf. the previous discussion). Buffon, Cuvier, Morton, Darwin, and many others strived to create “natural classifications” by Gestalt Similarity.
This fruitful line of research was effectively torpedoed by UNESCO in 1950, when it published its previously mentioned, ideologically based “The Race Question” statement, in an otherwise fully understandable attempt to counter the blatant Nazi misuse of racial thinking around the second world war. The UNESCO statement suggested in fact that, for historical reasons, research on race with biological connotations ought to be substituted by studies of ethnic groups in a cultural context. Many later biologists, cultural anthropologists, sociologists, and their academic organizations faithfully copied this unscientific and essentially politically motivated course, and some even began around 1950 to take an active and consequential stance against colleagues, whom they began to characterize as fraudulent race researcher (e.g., Woodley, Dutton, Figueredo, Carl, et al. 2018 [7]; for an earlier review, see Nyborg, 2003 [4]).
These critics found no inspiration in animal analog studies showing that behavioral differences related to melanin-based coloration. Ducrest, Keller, and Rouling, 2008) [79] thus observed that darker color is associated with greater aggressiveness in 10 mammal species, three kinds of birds, and more Lizard forms entirely evaded them. They condemned the color analogue with respect to humans, and reacted forcefully when Rushton and Templer (2009) [80] drew data from no less than 113 countries and found that “… murder, rape, and serious assault were associated with darker skin color, lower IQ, higher birth rate, higher infant mortality, higher HIV/AIDS rate, lower life expectancy, and lower income” (also see Templer, 2015) [81]. They disregard, as does NG today, that Census in the United States labels two of its five self-designated race categories by color (white and black) and then finds that even such a loose subjective classification of one’s own race relates significantly to pervasive medical, developmental, physical, economic, political, criminal, and other behavioral differences associated with skin color (Herrnstein and Murray, 1994) [54]. Neither do critics pay much attention to the fact that average IQ correlates ecologically (r = 0.93) with average skin color (Templer and Arikawa, 2006 [82]) (the correlation to IQ between individuals within a population is much lower but still significant).
It should be noted that the two variables are most likely genetically separate, so that correlated alleles for skin color do not guide IQ development, nor do correlated alleles for IQ guide skin color. The two most likely just co-vary along a latitudinal econiche gradient, IQ due to strong selection for increased brain power to survive under cold harsh conditions, and away from black skin color, which acted as a useful protective adaption in heavy Sun exposure tropics, but needed to gradually adapt into whitish to increasingly compensate during northbound migration for still lower production of vital D-vitamin in colder areas.
It is disquieting to find that prominent social scientists have used the IQ-skin color racial covariation to explain the establishment of so-called pigmentocracies or colored societal stratification within societies, but without referring with one word to the well-known IQ-skin color covariation. It is thought-provoking to think that political correctness or the firm social-constructivist persuasion of most of these researcher virtually warms/forbids them to infer co-variant race differences in IQ and color as a function of genetic differences. A straightforward explanation is at hand: Their results can be safely cited by anti-racists and their scholarly position would remain intact within currently politically and academically admissible lines; otherwise they would be sacked (Nyborg, 2003 [4]; Woodley, Dutton, Figueredo, Carl, et al., 2018 [7]).
To take an example, the anthropologist Alejandro Lipschutz (1944) [83] was the first to coin the term “Pigmentocracy” to characterize easily observable hierarchies or ethnoracial categories related to skin color. An ensuring PERLA in-depth survey in four countries thus separated the effects of ethnoracial categorization from skin color estimates (interviewers rated the color of the skin by comparing it to a color chart; Telles, 2014, pp. 3–4; Telles, Flores, and Urrea-Giraldo, 2015) [84,85]. They then found that “…skin color is a central axis of social stratification in at least several Latin American countries, though it is often ignored”. They also noted that ethnoracial classification “do[es] not consistently support expectations of pigmentocracy (Categorial Pigmentocracy)”. They arrived at three conclusions: (1) “Skin color is a more consistent but overlooked dimension of ethnoracial inequality in Latin America (Continuous Pigmentocracy”; (2) “Individuals with darker skin tone have significantly lower levels of educational attainment and occupational status”; and (3) “they are more likely to live in poverty and less likely to be affluent, even after controlling for other individual characteristics” (ibid., p 652) [86].
It generates a noteworthy reflection on the current forbidding academic climate at many modern universities that both Telles (2014, p 229–230) [84] and Villarreal (2010, p. 652) [86] limited themselves to keep the explanation of their remarkable pigmentocratic ranking within the accepted social-constructivist framework, by referring to discrimination, colonialism, low level social mobility, and other environmental repressive or structural factors, much in line with NG’s politically correct anti-racialist position. Aside from the possibility of stressful silent self-censure, another part of the explanation for this may be found in the fact that the PERLA team was manned exclusively by sociologists, anthropologists, linguists, and historians, housing not one scientist with a professional background in psychometrics, behavioral genetics, evolution, differential psychology, physiology, or physical anthropology.
This common neglect of easily available, but controversial, scientific evidence is peculiar, because Cold Winter theory and ample empirical evidence actually forecast pigmentocracy, not just based on “historical accidents”, unpremeditated exposure to the Sun, or white suppression, but on IQ.
Pigmentocratic stratification of society is, in fact, a predictable co-variant consequence of prehistoric migration through still more Sun power poor, harsh, northern areas, favoring selection of multiple functionally unrelated or related allele structures for skin color, brain size, intellectual power, and altruism, the effects of which are traceable today not only at a global geographic north-south scale, but also at the narrower within-country perspective.
Most likely, major latitudinal deviations from perfect co-variant north-south regression lines in skin color, brain size, IQ, altruism, and the above mentioned within-society pigmentocracies, reflect mainly local reverse migratory effects caused by natural disasters, sudden hostile local replacements, or other disturbances to the long-time residing interbreeding ecotype demes in various econiches.
However, the two main point relating to NG’s claims that race is caused by historical accidents and casual sun exposure are that—despite admittedly considerable historic replacements—we can still to this day trace rudiments of the phenotypic and genetic adaptations that took place during the prehistoric south-to-north migration along a latitudinal gradient, even within modern societies.
7. The Race doesn’t Matter Argument
Even a brief examination of the luckily now defunct brutal South African apartheid system and the unfortunately still current black-white tensions in the US suggest that race plays a prominent societal role in many peoples’ life.
Quantitatively, perhaps nothing better than the demographics of reproduction underlines the vital importance of the existence of race/lineage/ecotype differences. The current situation in this area proves that NG’s anti-racialist position is not only meaningless but actually harmful for our deeper understanding of the survival of old and modern societies. The reason for this is simple: Fertility differentials determine the presence or disappearance of races, lineages, ecotypes or what goes under similar names, from the evolutionary scene. Figure 3 illustrates this point.
The figure suggests that southern ecotypes adapted to survival in warm econiches will over time numerically outcompete cold ecotypes (everything else equal, which may not be).
The demographic situation has not always been this. Pre-historically speaking, the more intelligent probably had more surviving children than the less intelligent, as being able to deal rationally with the complexities of existence increases the probability of survival, successful reproduction, and protection of offspring, according to classical evolutionary theory. About 275.000 years after the Exodus from Africa, average brain size and intelligence had increased some 100 cm3 and 30 IQ points during increasingly harsher north-bound migration (Lynn, 2006/2015 [40]; Rushton, 2000 [47]). The costly re-shuffling of the internally fixed metabolic energy budget during migration had to be traded-off by allocating less energy to fertility endeavors, as part and parcel of the adaptation to survival in energy-poor northern areas with low caring capacity (e.g., Nyborg, 1994, 2013a) [88,89].
But then, around 1850 this pre-historic picture changed. For possibly the first time in history, the less gifted races/lineages/ecotypes situated at the lower left side of the IQ population distribution began to out-reproduce intelligent individuals found at the higher right side, due to improved hygiene, better nutrition, and social welfare increases in general. As the chances of survival increased more for the poor and uneducated (in Africa and in modern industrialized countries) than for the educated and rich (Lynn, 2006/2015 [40]), the proportion of gifted began to recede proportionally—worldwide and locally—together with a declining total world fertility rate.
No doubt, a brief low-reproduction period, where parents have fewer than 2.05 children on average, may actually benefit not only Africa and the Middle East, but also the most densely populated modern western societies, since fewer individuals spend less energy and leave less pollution; the formation of agriculture and Industrialization succeeded with fewer people than living today. But as soon as ample resources began to accumulate from large-scale industrialization, they began to favor the less educated more than the productive high-IQ groups (Lynn, 2006/2015 [40]). This opens up for the somber perspective that modern welfare societies gradually build up a self-destructive genetic potential, where the average IQ declines and with it the collective productivity factor that originally build it up and carried it on (Nyborg, 2012) [90].
Figure 2 illustrated how gifted ecotypes/races/lineages/species turn into evolutionary losers when they over time have fewer than 2.05 surviving children per parents, whereas fertile less gifted warm ecotypes slowly take over the evolutionary scene. This reminds us of Shakespeare’s eternal words: “To be or not to be, that is the question”. This somber scenario opens up with the UN World Population Prospect: 2015 Revision (2015) [91], confirming that despite a worldwide reduced fertility, the highest rate of population growth is for warm middle Eastern and African ecotypes with their relatively young population profile, relative to Europe’s. In fact, these econiches are expected to be the home for more than half the entire world’s population growth from now onto 2050, while European ecotypes have long decreased in number (Nyborg, 2012) [90].
In addition to the somber environmental, economic, societal, human, and political challenges inherent in these quantitative demographic changes, Table 1 illustrates a serious qualitative evolutionary consequence of the current race/lineage/ecotype difference in fertility, IQ, and brain size.
The fertility data in Table 1 is from CIA (2017) [92], standardized, and broken down into five equidistant geo-bio-climatic categories of a distribution (derived by Principal Component factoring of three variables: (1) Average country latitudinal distance from Equator in degree, (2) Temperature in centigrade, and (3) Skin Color grade. Stipulated brain size refers to the result of applying the mathematical model by Smith and Beals (1990) [41], according to which—based on multiple empirical pre-historic skull measures—we should add 2.5 cm3 brain mass per degree latitude gained over the African average of 1297 cm3, the latter established by Rushton (1992, 2000; Rushton and Rushton, 2003) [33,34,47].
The data pattern in Table 1 supports the hypothesis of a dynamic intra-systemic metabolic trade-off between average ecologic fertility and brain size and that this trade-off differs by race/lineage/ecotype. Now, given that large-brained high-IQ individuals currently fade away over time due to their relative infertility (Figure 2), and that small-brained fertile individuals increase in number, the world brain size average drops, as does the world average IQ (Becker, 2019) [49]. In particular, for high-IQ/low-fertile Europe, the recent significant immigration from southern non-Western low-IQ immigration will, if unopposed, relatively fast reduce its average IQ level (Nyborg, 2012) [90].
These vast demographic transitions represent a challenge to the sustainability of democracy and Enlightenment in high-IQ areas, according to Lynn (2006/2015 [40]; Vanhanen, 2009 [93]). NG’s ignorance or denial of the importance of race differences for such prospective demographic scenarios, may leave us blind to such austere evolutionary realities and to the dynamic changes currently taking place in a modern highly mobile globalized world. Anti-racism reduces the possibility of dealing rationally and prospectively with these problems in the name of the common good. This is the ultimate sin of people and institutions blind—or blinding us—to the importance of race/lineage/ecotype differences. Modern ignorance may mean the difference between the rise and fall of modern civilizations (Nyborg, 2012) [90].
8. General Discussion
The history of differential psychology is sprinkled with examples of sex and race difference researchers being challenged on their skills and moral by their universities, in the media, and in court (see review by Nyborg, 2003) [4].
It is within this anti-scientific movement we find the April 2018 Race Issue of National Geographic (NG) attack on race difference research(ers), exemplary for its unforgiving denunciation of named scientist(s) by questioning their motives and downright accusing them of deliberately harming minority identity and politics.
NG’s empirical misrepresentations may not be the largest scientific problem here, because it is so easy to counter by solid scientific data, accumulated over centuries by multiple researchers in such different areas as medicine, law, psychometrics, criminology, behavioral genetics, and governance. Given that the discussion of the existence, nature, size, and direction of existing race/lineage/ecotype differences is kept close to an objective level, it is ideally just a matter of time before scientifically sound (and humane) knowledge of such difference can be turned into good use for individuals, minority groups, and society in general. The real issue here is not even that NG singles out individual researchers for naming and shaming.
The real danger is that widely respected institutions such as NG mobilizes its full authority in a massively circulated attempt to betray its scientific and public readership by systematically misrepresenting historical sources and selectively suppressing unwanted or unacceptable results—acts included as examples of academic fraud by the National Academy of Sciences (US; 1986) [94]. NG deliberately bypasses ample counter-evidence and misrepresents the evidence it sees.
In this way NG not only amplifies the historical harm done to psychometric and demographic research in matters of considerable interest to society, but it further reinforces the general injury through its worldwide circulation in about 40 languages to about 6.5 million readers. Being such a central source of information about science, nature, and peoples to millions of professionals and lay people alike, NG carries a serious responsibility. Failure to live up to this obligation is bound to exert a pervasive negative impact on future studies in sensible areas such as individual, minority, and population differences.
The force of NG’s anti-racist stance begs the question of whether the magazine had a priori imperative motive(s) for launching its devastating attack on race research(ers)? The answer is in fact already provided by the current editor of NG, Susan Goldberg.
On her recent appointment, she found that NG had historically treated groups of people around the world in a racist way. She consequently “… decided to re-examine its coverage to mark 50 years since civil rights leader Martin Luther King was murdered. “Let’s confront today’s shameful use of racism as a political strategy and prove we are better than this”, she said on BBC News (13 March 2018). In an Editorial she followed up by admitting that “… some of the magazine’s archive material left her “speechless”, including a 1916 photo of Australian Aborigines with the caption “South Australian Blackfellows: These savages rank lowest in intelligence of all human beings”.
Editorial statements such as these leave little doubt that the purpose, content, and extend of the “Race Issue” was motivated by, (1) A deep-felt wish to “repair” previous “shameful use of racism as a political strategy”, (2) An urge to excuse the unacceptable ignorance of non-white Americans, and (3) A bleeding culpability over decades of “…promoting racial clichés.” In brief: White shame motivated NG’s attack on predominantly white race research(ers).
This raises several interesting ethical questions. First, does admitting to white shame, ignorance of minorities, racial clichés, and alleged minority harm justify sweeping misrepresentations of current race research, selective omission of unwanted results, and questioning the motives and findings of scientific race researcher—all in the face of millions of readers worldwide? We think not.
Second, race critics often demand of race difference researcher to adhere to extra high scientific standards because of the potential harm their findings might do, but ironically, not all such critics seem themselves overly concerned with living up to such elevated scientific standards (Gottfredson, 1994, 2000 [10,11]; Gross and Levitt,1994/1998 [14]; Nyborg, 2003 [4]; Sesardic, 2010 [6]; Woodley, Dutton, Figueredo, and Carl, 2018 [7]). NG’s April 2018 “Race Issue” exemplifies in reality a largely data-table free series of innuendos, misrepresentations, and unfair accusations, based on editorial guilt and the idea that from race science comes only evil knowledge and social harm.
Ethics finally comes to mind when stringent methodology, impeccable theory, and standard experimental psychology methods are misused in the examination of effects of racial bias and racial discrimination, resulting in unreliable results, scapegoating of serious scientists, and even wrongly altering the direction in race training programs. Francis (2015) [95] thus used TES (a general method to test for excessive success when relating experimental findings to theoretical conclusions (for details, see Francis, 2013 [96]), to analyze three related papers finding that racial bias changes perception and decision making in training programs and elsewhere (Eberhardt, Goff, Purdie, and Davies, 2004; Goff, Eberhardt, Williams, and Jackson, 2008; Williams and Eberhardt, 2008 [97,98,99]; also see MacArthur Foundation, 2014 [100]). Using the TES method, Francis found that all three studies had “Excess success”, and very low probability of proper replication. He concluded that they “…should not be used to guide policies related to racial bias …” and that it seems “… premature to apply these ideas to policy decisions or to interventions that might reduce the presence or impact of racial bias.”
Judged in the global perspective of ever-existing global cultural and economic race/lineage/ecotype conflicts it seems of little hope that such “too successful” studies and NG’s mistreatment of race difference research—will ever lead to anything good. Any attempt to gloss over or deny pre-historic or current ecologic realities might prevent us from ever being able to develop and apply rational means to solve current global or internal societal conflicts on a rational basis because we know not of their true nature.
Geographic variation in genetic, phenotypic, and demographic characteristics is of real, significant, and considerable societal importance, and too central for the development of fair policies, peace processes, welfare and family programs, democracy, and a civilized life for all, to be ignored, misrepresented, or shamed.
The recent explosion in Genome Wide Association Studies (GWAS) promises to soon lay bare the molecular DNA foundation of evolutionary and developmental differences in terms of identifying correlated allelic DNA structures guiding the development of individual, sex, family, and population phenotypic similarities and differences. It may in fact hold the key to more insightful future engineering of traits for a better world for all, and there is admittedly ample space for improvements.
However, we prefer to conclude on a more modest note by suggesting that until then we might cautiously use racial categorization as a rough preliminary solution until the arrival of much more sophisticated classifications of individuals and their differentially aggregated ecotypes, combining metabolic traces of the particular selective evolutionary forces which ruled in their different long-term econiches with the often weaker environmental impacts. A deeper politically motive-free scientific understanding of “molecular man in a molecular world” (Nyborg, 2001) [67] may in the long run increase our understanding and promotion of lived harmony among democratic peoples. Future generations deserve it.
Funding
This research received no external funding.
Acknowledgments
The many constructive and critical comments from the anonymous reviewers are gratefully acknowledged.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rushton, J.P.; Jensen, A.R. Thirty years of research on race differences in cognitive ability. Psychol. Public Policy Law 2005, 11, 235–294. [Google Scholar] [CrossRef]
- Tate, C.; Audette, D. Theory and Research on “Race” as a Natural Kind Variable. Theory Psychol. 2001, 4. [Google Scholar] [CrossRef]
- Gottfredson, L. Lessons in academic freedom as lived experience. Pers. Individ. Differ. 2010, 49, 272–280. [Google Scholar] [CrossRef]
- Nyborg, H. The sociology of psychometric and biobehavioral sciences: A case study of destructive social reductionism and collective fraud in 20th century academia. In The Scientific Study of General Intelligence: Tribute to Arthur R. Jensen; Nyborg, H., Ed.; Pergamon/Elsevier Science: Amsterdam, The Netherland, 2003; pp. 441–501. [Google Scholar]
- Scarr, S. Three cheers for behavior genetics: Winning the war and losing our identity. Behav. Genet. 1987, 17, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, N. Race: A social destruction of a biological concept. Biol. Philos. 2010, 25, 143–162. [Google Scholar] [CrossRef]
- Dutton, E.; Figueredo, A.J.; Carl, N.; Debes, F.; Hertler, S.; Irwing, P.; Kura, K.; Lynn, R.; Madison, G.; Meisenberg, G.; Miller, E. Communicating intelligence research: Media misrepresentation, the Gould Effect and unexpected forces. Intelligence 2018, 70, 80–87. [Google Scholar]
- UNESCO Publication. UNESCO and Its Programme, III, The Race Question; UNESCO Digital Library: Paris, France, 1950; p. 791. [Google Scholar]
- Montagu, A. Man’s Most Dangerous Myth: The Fallacy of Race; Columbia University Press: New York, NY, USA, 1999; pp. 496–498. [Google Scholar]
- Gottfredson, L. Egalitarian fiction and collective fraud. Society 1994, 31, 53–59. [Google Scholar] [CrossRef]
- Gottfredson, L. Equal potential: A collective fraud. Society 2000, 37, 19–28. [Google Scholar] [CrossRef]
- Gould, S.J. The Mismeasure of Man; W.W. Norton & Co.: New York, NY, USA, 1981. [Google Scholar]
- Gould, S.J. The Mismeasure of Man (Revised and Expanded 2nd edition); W.W. Norton & Co.: New York, NY, USA, 1996. [Google Scholar]
- Gross, P.R.; Levitt, N. Higher Superstition: The Academic Left and Its Quarrels with Science; Johns Hopkins University Press: Baltimore, MD, USA, 1997. [Google Scholar]
- Lewontin, R.; Rose, S.; Kamin, L. Not in Our Genes; Penguin Books: New York, NY, USA, 1984. [Google Scholar]
- Alcock, J. Unpunctuated equilibrium in the Natural History Essays of Stephen Jay Gould. Evol. Hum. Behav. 1998, 19, 321–336. [Google Scholar]
- Lewis, J.E.; Degusta, D.; Meyer, M.R.; Monge, J.M.; Mann, A.E.; Holloway, R.L. The mismeasure of science: Stephen Jay Gould versus Samuel George Morton on skulls and bias. PLoS Biol. 2011, 9, e1001071. [Google Scholar] [CrossRef]
- Parry, R. All Investigative Journalists Do What Julian Assange Did. Available online: https://consortiumnews.com/ (accessed on 28 April 2019).
- Cofnas, N. Science is not always “self-correcting”: Fact-value conflation and the study of intelligence. Found. Sci. 2016, 21, 477–492. [Google Scholar] [CrossRef]
- Goldberg, S. National Geographic. Race Issue. April 2018. Available online: https://www.nationalgeographic.com/magazine/2018/04/race-genetics-science-africa/ (accessed on 28 April 2019).
- Müller-Ville, S.; Rheinberger, H.-J. A Cultural History of Heredity; The University of Chicago Press Books: Chicago, IL, USA, 2012. [Google Scholar]
- Buffon. Histoire Naturelle, Générale et Particulière, Avec la Description du Cabinet du Roi, Tome Quatorzième (« Histoire Naturelle des Quadrupèdes » XI); Imprimerie Royale: Paris, France, 1766; pp. 1749–1789. [Google Scholar]
- De Quatrefages, A. Histoire Générale des Races Humaines: Introduction a L’Étude des Races Humaines; A Hennuyer, Imprimeur-Éditeur: Paris, France, 1889. [Google Scholar]
- Morton, G.M. Types of Mankind: Or, Ethnological Researches Based Upon Their Natural, Geographical, Philological, and Biblical History, 8th ed.; J.B. Lippincott & Co.: Philadelphia, PA, USA, 1857; Available online: https://archive.org/details/typesofmankindo00nott/page/n10 (accessed on 28 April 2019).
- Nott, J.C.; Gliddon, G.R. Types of Mankind: Or, Ethnological Researches Based upon Their Natural, Geographical, Philological, and Biblical History; Lippincott, Grambo & Co.: Philadelphia, PA, USA; Trübner & Co.: London, UK, 1854; Available online: https://archive.org/details/typesofmankindo00nott/page/42 (accessed on 28 April 2019).
- Morton, S.G. Hybridity in Animals and Plants, Considered in Reference to the Question of the Unity of the Human Species: Read Before the Academy of Natural Sciences of Philadelphia; Stanford Library: Stanford, CA, USA, 1847. [Google Scholar]
- Morton, G.M. Dr. Morton’s Craniological Collection: On the Varieties of the Human Species (Introduction); American Ethnological Society: New York, NY, USA, 1845; p. 245. Available online: https://archive.org/details/transactionsame00socigoog/page/n22. (accessed on 28 April 2019).
- Morton, G.M. Brief Remarks on the Diversities of the Human Species: And on Some Kindred Subjects: Being an Introductory Lecture Delivered Before the Class of Pennsylvania Medical College; Merrihew & Tompson, Printers: Philadelphia, PA, USA, 1842. [Google Scholar]
- Morton, S.G. To the Rev. John Bachman, D.D., on the Question of Hybridity in Animals, Considered in Reference to the Unity of the Human Species; Steam-Power Press of Walker & James: Charleston, SC, USA, 1850; Available online: https://babel.hathitrust.org/cgi/pt?id=njp.32101075715951;view=1up;seq=5 (accessed on 28 April 2019).
- GENO 2.0 Your DNA. Your Story. Available online: https://genographic.nationalgeographic.com/about/ (accessed on 28 April 2019).
- Doron, C.-O. The Birth of Race (L’Homme Altéré. Race et Dégénérescence); Champ Vallon: Paris, France, 2016. [Google Scholar]
- Reich, D. Who We Are and How We Got Here: Ancient DNA and the New Science of the Human Past; Oxford University Pres: Oxford, UK, 2018. [Google Scholar]
- Rushton, J.P. Cranial capacity related to sex, rank and race differences in a stratifies sample of 6325 U.S. military personnel. Intelligence 1992, 16, 401–413. [Google Scholar] [CrossRef]
- Rushton, J.P.; Rushton, E. Brain size, IQ, and racial-group differences: Evidence from musculoskeletal traits. Intelligence 2003, 31, 139–155. [Google Scholar] [CrossRef]
- Symmons, K. Cranial capacities by both plastic and water techniques with cranial linear measurements of the Reserve Collection: White and Negro. Hum. Biol. 1942, 14, 473–498. [Google Scholar]
- Gould, S.J. Morton’s ranking of races by cranial capacity. Science 1978, 200, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Wade, N. Scientists Measure the Accuracy of a Racism Claim. The New York Times, 14 June 2011. [Google Scholar]
- A Critique of the Work of Stephen Jay Gould Should Serve as Encouragement to Scrutinize the Celebrated While They Are Still Alive. Available online: https://www.nature.com/articles/474419a (accessed on 28 April 2019).
- Weisberg, M. Remeasuring man. Evol. Dev. 2014, 16, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Lynn, R. Race Differences in Intelligence: An Evolutionary Analysis (Second Revised Edition); Washington Summit Publishers: Augusta, GA, USA, 2015. [Google Scholar]
- Smith, C.L.; Beals, K.L. Cultural correlates with cranial capacity. Am. Anthr. 1990, 92, 193–200. [Google Scholar] [CrossRef]
- Beals, K.L.; Smith, C.L.; Dodd, S.M. Brain Size, Cranial Morphology, Climate, and Time Machines. Curr. Anthr. 1984, 3, 301–330. [Google Scholar] [CrossRef]
- Lynn, R. The Global Bell Curve: Race, IQ, and Inequality Worldwide; Washington Summit Publishers: Augusta, GA, USA, 2008. [Google Scholar]
- Rushton, J.P. Race differences in behaviour: A review and evolutionary analysis. Pers. Individ. Differ. 1988, 9, 1009–1024. [Google Scholar] [CrossRef]
- Rushton, J.P. Sex and race differences in cranial capacity from International Labor Office data. Intelligence 1988, 19, 281–294. [Google Scholar] [CrossRef]
- Rushton, J.P. Cranial size and IQ in Asian Americans from birth to seven. Intelligence 1997, 25, 7–20. [Google Scholar] [CrossRef]
- Rushton, J.P. Race, Evolution and Behaviour; Charles Darwin Research Institute: Port Huron, MI, USA, 2000. [Google Scholar]
- Rushton, J.P.; Osborne, R.T. Genetic and environmental contributions to cranial capacity in black and white adolescents. Intelligence 1995, 20, 1–13. [Google Scholar] [CrossRef]
- Becker, D. The NIQ-DATASET (V1.3.2.). 2019. Available online: https://www.researchgate.net/project/Worlds-IQ (accessed on 28 April 2019).
- Lynn, R.; Vanhanen, T. IQ and the Wealth of Nations; Praeger: Westport, CT, USA, 2002. [Google Scholar]
- Lynn, R.; Vanhanen, T. Intelligence: A Unifying Construct for the Social Sciences; Ulster Institute for Social Research: London, UK, 2012. [Google Scholar]
- Gottfredson, C. The Abilities of Man: Their Nature and Measurement; Macmillan: New York, NY, USA, 1927. [Google Scholar]
- Jensen, A.R. The g Factor: The Science of Mental Ability; Praeger: Westport, CT, USA, 1998. [Google Scholar]
- Herrnstein, R.J.; Murray, C. The Bell Curve: Intelligence and Class Structure in American Life; Free Press: New York, NY, USA, 1994. [Google Scholar]
- Gottfredson, L. Mainstream science on intelligence (editorial). Intelligence 1997, 24, 13–23. [Google Scholar] [CrossRef]
- Gottfredson, L.S. Why g matters: The complexity of everyday life. Intelligence 1997, 24, 79–132. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, T.J. The Wilson effect: The increase in heritability of IQ with age. Twin Res. Hum. Genet. 2013, 16, 923–930. [Google Scholar] [CrossRef]
- Plomin, R.; DeFries, J.C.; Knopik, V.S.; Neiderhiser, J.M. Behavioral Genetics, 7th ed.; Worth Publishers: New York, NY, USA, 2016. [Google Scholar]
- Murray, C. Human Accomplishment: The Pursuit of Excellence in the Arts and Sciences; Harper Collins: New York, NY, USA, 2003. [Google Scholar]
- Plomin, R.; von Stumm, S. The new genetics of intelligence. Nat. Rev. Genet. 2018, 19, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Piffer, D. Factor analysis of population allele frequencies as a simple, novel method of detecting signals of recent polygenic selection: The example of educational attainment and IQ. Mank. Q. 2013, 54, 168–200. [Google Scholar]
- Piffer, D. A review of intelligence GWAS hits: Their relationship to country IQ and the issue of spatial autocorrelation. Intelligence 2015, 53, 43–50. [Google Scholar] [CrossRef]
- Rindermann, H.; Woodley, M.A.; Stratford, J. Haplogroups as evolutionary markers of cognitive ability. Intelligence 2012, 40, 362–375. [Google Scholar] [CrossRef]
- León, F.R.; Burga-León, A. How geography influences complex cognitive ability. Intelligence 2015, 50, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Becker, D.; Rindermann, H. The relationship between cross-national genetic distances and IQ-differences. Pers. Individ. Differ. 2016, 98, 300–310. [Google Scholar] [CrossRef]
- Nyborg, H. Molecular man in a molecular world: Applied physicology. Psyche Logos 1998, 18, 457–474. [Google Scholar]
- Nyborg, H. The greatest collective scientific fraud of the 20th Century: The demolition of differential psychology and eugenics. Mank. Q. 2011, 51, 241–268. [Google Scholar]
- Ramachandran, S.; Deshpande, O.; Roseman, C.C.; Rosenberg, N.A.; Feldman, M.W.; Cavalli-Sforza, L.L. Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. Proc. Natl. Acad. Sci. USA 2005, 102, 15942–15947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewontin, R. The apportionment of human diversity. Evol. Biol. 1972, 6, 391–398. [Google Scholar]
- Edwards, A.W.F. Human genetic diversity: Lewontin’s fallacy. BioEssays 2003, 25, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.R.; Hemani, G.; Medina-Gomez, C.; Mezzavilla, M.; Esko, T.; Shakhbazov, K.; Powell, J.; Vinkhuyzen, A.; Berndt, S.I.; Gustafsson, S.; et al. Population genetic differentiation of height and body mass index across Europe. Nat. Genet. 2015, 47, 1357–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novembre, J.; Johnson, T.; Bryc, K.; Kutalik, Z.; Boyko, A.R.; Auton, A.; Indap, A.; King, K.S.; Bergmann, S.; Nelson, M.R.; et al. Genes mirror geography within Europe. Nature 2008, 456, 98–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okbay, A.; Beauchamp, J.P.; Fontana, M.A.; Lee, J.J.; Pers, T.H.; Rietveld, C.A.; Turley, P.; Chen, G.B.; Emilsson, V.; Meddens, S.F.; et al. Benjamin Genome-wide association study identifies 74 loci associated with educational attainment. Nature 2016, 533, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Morton, G.M. Crania Americana; or a Comparative View of the Skulls of Various Aboriginal Nations of North and South America: To Which Is Prefixed an Essay on the Varieties of the Human Species; J. Dobson: Philadelphia, PA, USA, 1839; Available online: https://archive.org/details/cihm_38366/page/n5 (accessed on 28 April 2019).
- Venter, C. 2000. Available online: https://www.genome.gov/10001356/june-2000-white-house-event/ (accessed on 28 April 2019).
- Tal, O. Two complementary perspectives on inter-individual genetic distance. Biosystems 2013, 111, 18–36. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS Discovery: Biology, Function, and Translation. Am. J. Hum. Genet. 2017, 101, 5–22. [Google Scholar] [CrossRef] [Green Version]
- Blumenbach, J.F. Handbuch der Naturgeschichte. 1779. Available online: http://www.deutschestextarchiv.de/book/view/blumenbach_naturgeschichte_1779?p=85 (accessed on 28 April 2019).
- Ducrest, A.-L.; Keller, L.; Roulin, A. Pleiotropy in the melanocortin system, coloration, and behavioural syndromes. Trends Ecol. Evol. 2008, 23, 502–510. [Google Scholar] [CrossRef]
- Rushton, J.P.; Templer, D.I. National differences in intelligence, crime, income, and skin color. Intelligence 2009, 37, 341–346. [Google Scholar] [CrossRef]
- Templer, D.I. Rushton: The great theoretician and his contribution to personality. In The Life History Approach to Human Differences: A Tribute to J. Phillippe Rushton; Nyborg, H., Ed.; Ulster Institute for Social Science: London, UK, 2016; Chapter 6; pp. 213–232. [Google Scholar]
- Templer, D.I.; Arikawa, H. Temperature, skin color, per capita income, and IQ. Intelligence 2006, 34, 121–128. [Google Scholar] [CrossRef]
- Lipschutz, A. El Indoamericamismo y el Problema Racial en las Américas; Nascimento: Santiago, Chile, 1944. [Google Scholar]
- Telles, E.E. Pigmentocracies: Ethnicity, Race, and Color in Latin America, and the Project on Ethnicity and Race in Latin America (PERLA); The University of North Carolina Press: Chapel Hill, NA, USA, 2014. [Google Scholar]
- Telles, E.; Flores, R.D.; Urrea-Giraldo, F. Pigmentocracies: Educational inequality, skin color and census identification in eight Latin American Countries. Res. Soc. Strat. Mobil. 2015, 40, 39–58. [Google Scholar] [CrossRef]
- Villarreal, A. Stratification by skin color in contemporary Mexico. Am. Sociol. Rev. 2010, 75, 652–678. [Google Scholar] [CrossRef]
- Nyborg, H. What made Europe great and what could destroy it. In Proceedings of the 2017 American Renaissance Conference “Turning the Tide”, Nashville, TN, USA, 28–30 July 2017; Available online: https://www.youtube.com/watch?v=02vvYDxXQ3w (accessed on 28 April 2019).
- Nyborg, H. Hormones, Sex, and Society: The Science of Physicology; Greenwood Publishing Group: Westport, CT, USA, 1994; Available online: http://helmuthnyborg.dk/wp-content/uploads/2016/07/Publ_1994_Sex-Hormones-and-Society_Book.pdf (accessed on 28 April 2019).
- Nyborg, H. Migratory selection for inversely related covariant T-, and IQ-Nexus traits: Testing the IQ/T-Geo-Climatic-Origin theory by the General Trait Covariance model. Pers. Individ. Differ. 2013, 55, 267–272. [Google Scholar] [CrossRef]
- Nyborg, H. The decay of Western Civilization: Double relaxed Darwinian selection. Pers. Individ. Differ. 2012, 53, 118–125. [Google Scholar] [CrossRef]
- UN World Population Prospect: 2015 Revision. 2015. Available online: https://www.un.org/en/development/desa/publications/world-population-prospects-2015-revision.html (accessed on 28 April 2019).
- Central Intelligence Agency (CIA). The World Fact Book; Central Intelligence Agency: Langley, VA, USA, 2017. Available online: https://www.cia.gov/library/publications/the-world-factbook/ (accessed on 28 April 2019).
- Vanhanen, T. The Limits of Democratization: Climate, Intelligence, and Resource Distribution; Washington Summit Publishers: Washington, DC, USA, 2009. [Google Scholar]
- National Academy of Sciences. National Academy of Engineering (US) and Institute of Medicine (US) Panel on Scientific Responsibility and the Conduct of Research. Responsible Science: Ensuring the Integrity of the Research Process: Volume II; Procedures for Investigating Academic Fraud; National Academies Press (US): Washington, DC, USA, 1993; p. 18. Available online: https://www.ncbi.nlm.nih.gov/books/NBK236197/ (accessed on 28 April 2019).
- Francis, G. Excess success for three related papers on racial bias. Front. Psychol. 2015, 6, 512. [Google Scholar] [CrossRef] [PubMed]
- Francis, G. Replication, statistical consistency, and publication bias. J. Math. Psychol. 2013, 57, 153–169. [Google Scholar] [CrossRef]
- Eberhardt, J.L.; Goff, P.A.; Purdie, V.J.; Davies, P.G. Seeing black: Race, crime, and visual processing. J. Pers. Soc. Psychol. 2004, 87, 876–893. [Google Scholar] [CrossRef] [PubMed]
- Goff, P.A.; Eberhardt, J.L.; Williams, M.J.; Jackson, M.C. Not yet human: Knowledge, historical dehumanization, and contemporary consequences. J. Pers. Soc. Psychol. 2008, 94, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.; Eberhardt, J.L. Biological conceptions of race and motivation to cross racial boundaries. J. Pers. Soc. Psychol. 2008, 94, 1033–1047. [Google Scholar] [CrossRef] [PubMed]
- MacArthur Foundation. MacArthur Fellows: Jennifer L. Eberhardt. 2014. Available online: http://www.macfound.org/fellows/913/ (accessed on 28 April 2019).
Figure 1.
The latitudinal distribution of IQ by indigenous peoples (Lynn, 2006 [35]; white areas mean no data).
Figure 1.
The latitudinal distribution of IQ by indigenous peoples (Lynn, 2006 [35]; white areas mean no data).
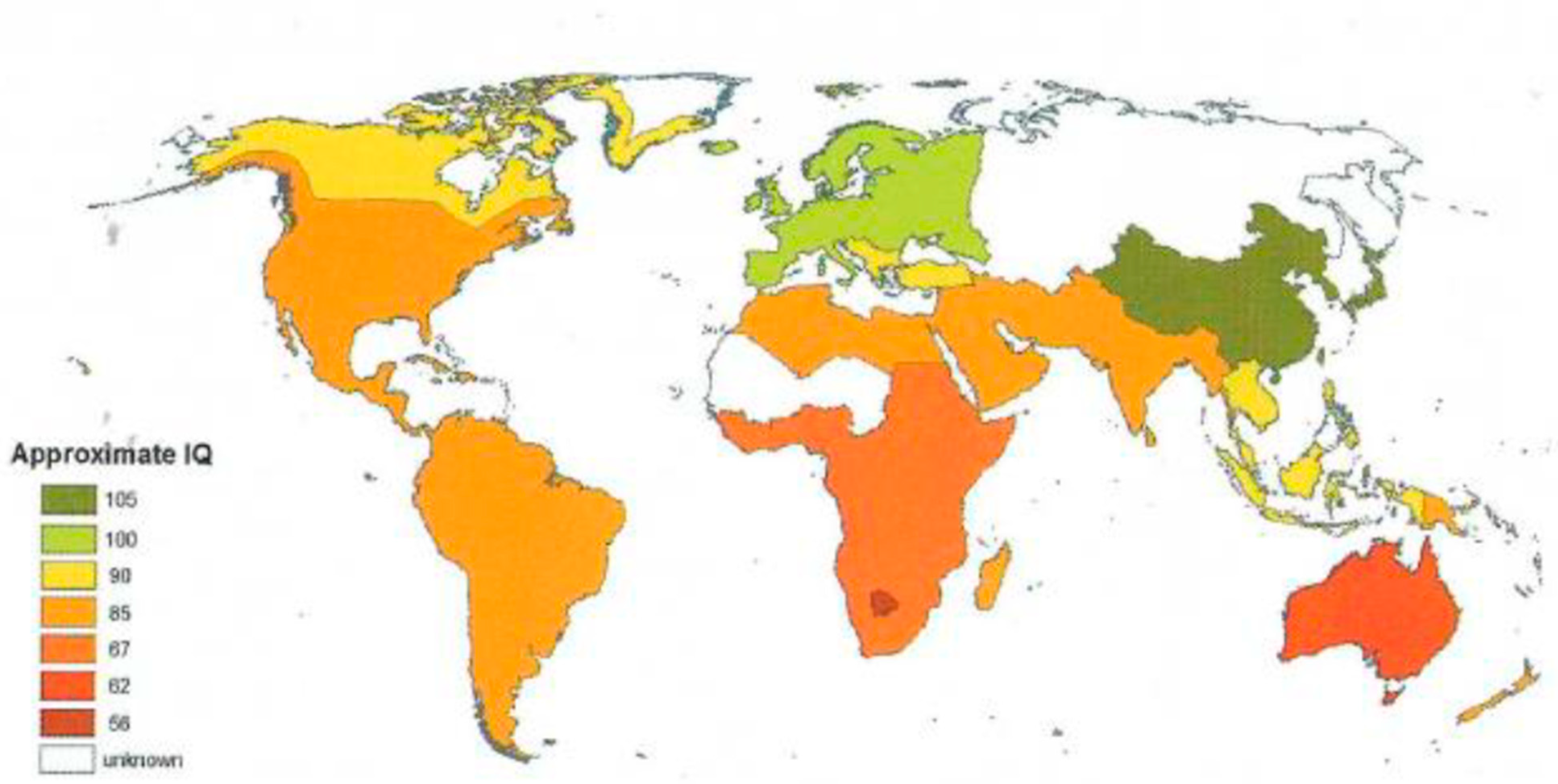
Figure 2.
Current latitudinal distribution of world IQs (Becker, 2019) [49].
Figure 2.
Current latitudinal distribution of world IQs (Becker, 2019) [49].
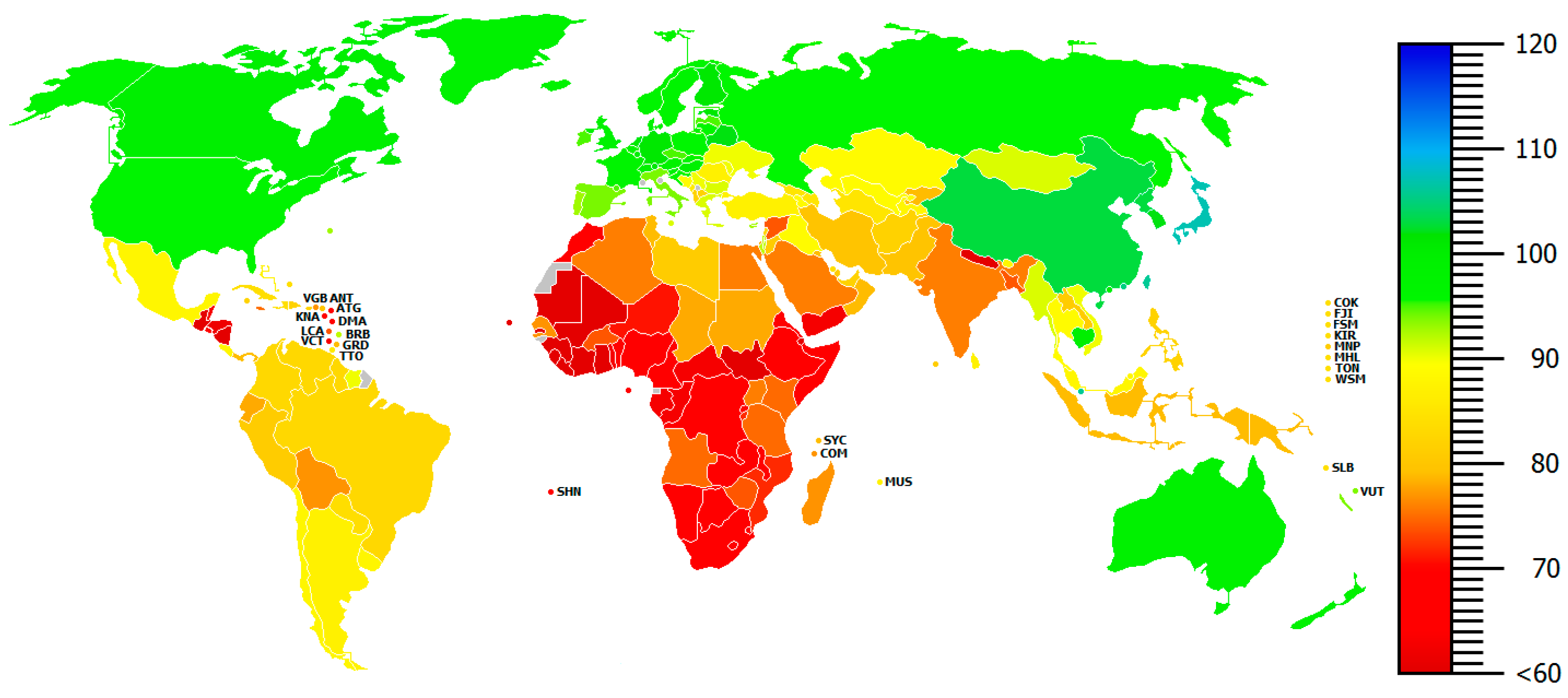
Figure 3.
Fertility rates—however measured—are differentially distributed along a five-category geo-bio-climatic north-south econiche gradient. These ecotype categories largely reflect the five major races (Nyborg, 2017) [87].
Figure 3.
Fertility rates—however measured—are differentially distributed along a five-category geo-bio-climatic north-south econiche gradient. These ecotype categories largely reflect the five major races (Nyborg, 2017) [87].
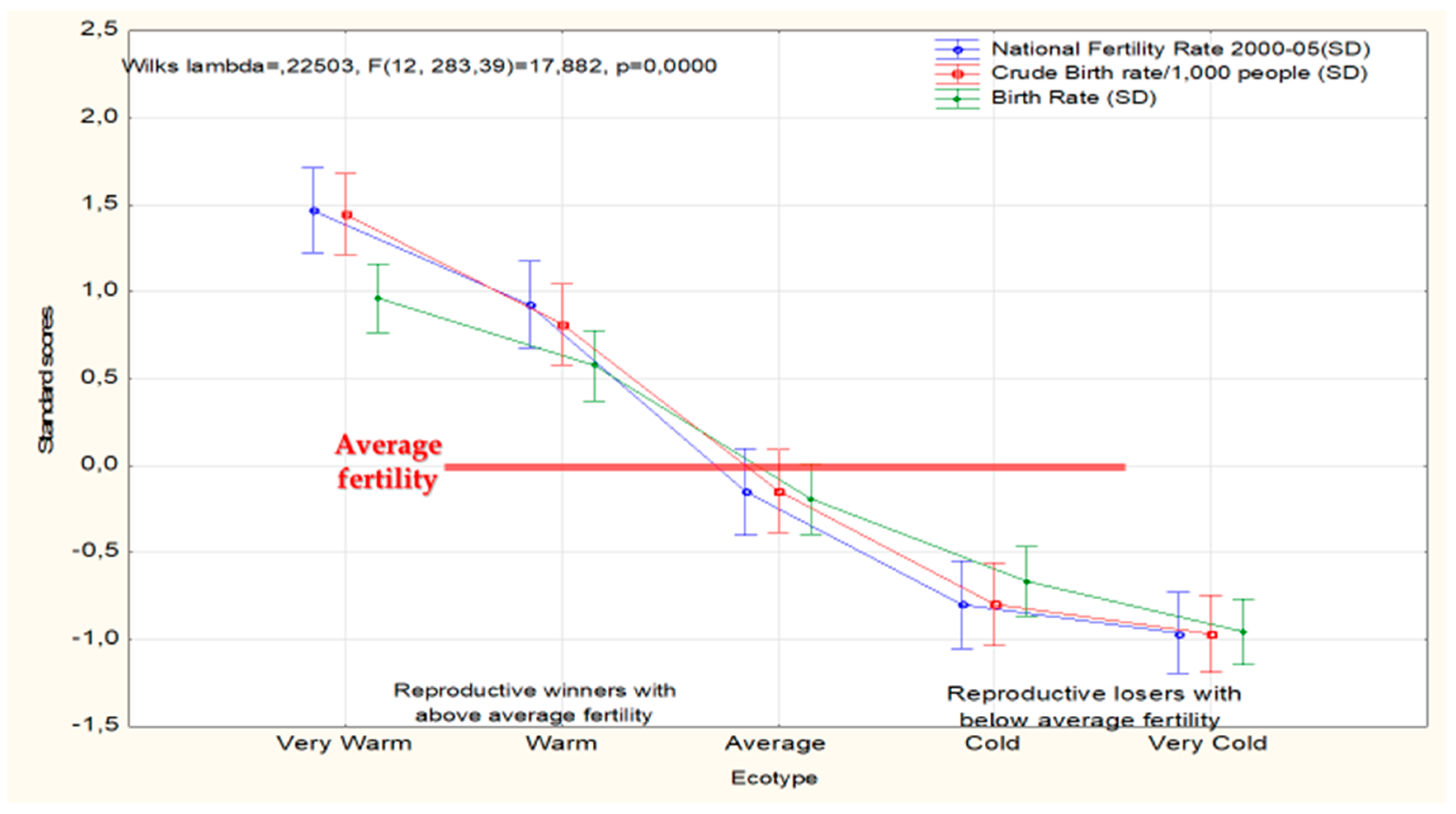

{kind=link}
{kind=link}
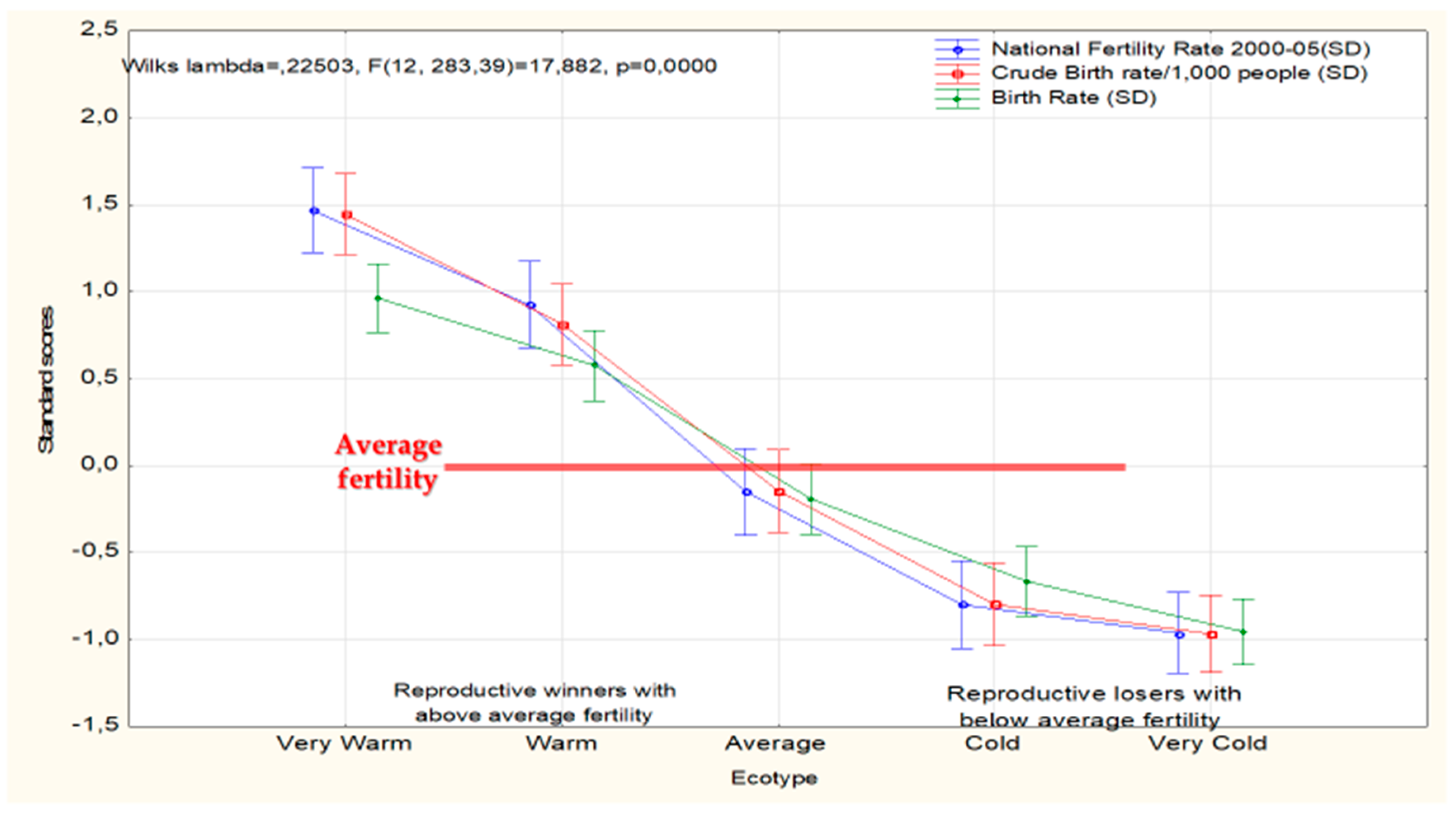{kind=link}
Table 1.
The inverse ecologic relationship among average fertility and stipulated brain size (Nyborg, 2017) [27].
Table 1.
The inverse ecologic relationship among average fertility and stipulated brain size (Nyborg, 2017) [27].
| Ecotype | Fertility | Brain Size |
|---|---|---|
| Very Warm | 4.66 | 1297 |
| Warm | 3.79 | 1312 |
| Average | 2.30 | 1350 |
| Cold | 1.88 | 1375 |
| Very Cold | 1.64 | 1399 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nyborg, H. Race as Social Construct. Psych 2019, 1, 139-165. https://doi.org/10.3390/psych1010011
AMA Style
Nyborg H. Race as Social Construct. Psych. 2019; 1(1):139-165. https://doi.org/10.3390/psych1010011
Chicago/Turabian StyleNyborg, Helmuth. 2019. "Race as Social Construct" Psych 1, no. 1: 139-165. https://doi.org/10.3390/psych1010011