
High-Resolution Multiproxy Record of Environmental Changes and Anthropogenic Activities at Unguja Ukuu, Zanzibar, Tanzania during the Last 5000 Years


 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.1.1. Environmental Setting
2.1.2. Archaeological Context
2.2. Sediment Core Sampling
2.3. Palaeoecological Analyses
2.3.1. Pollen and Spore Analysis
2.3.2. Phytolith Analysis
2.3.3. Microcharcoal Analysis
2.3.4. Trace Element Analysis
2.3.5. Particle Size Analysis
2.3.6. Chronology
3. Results
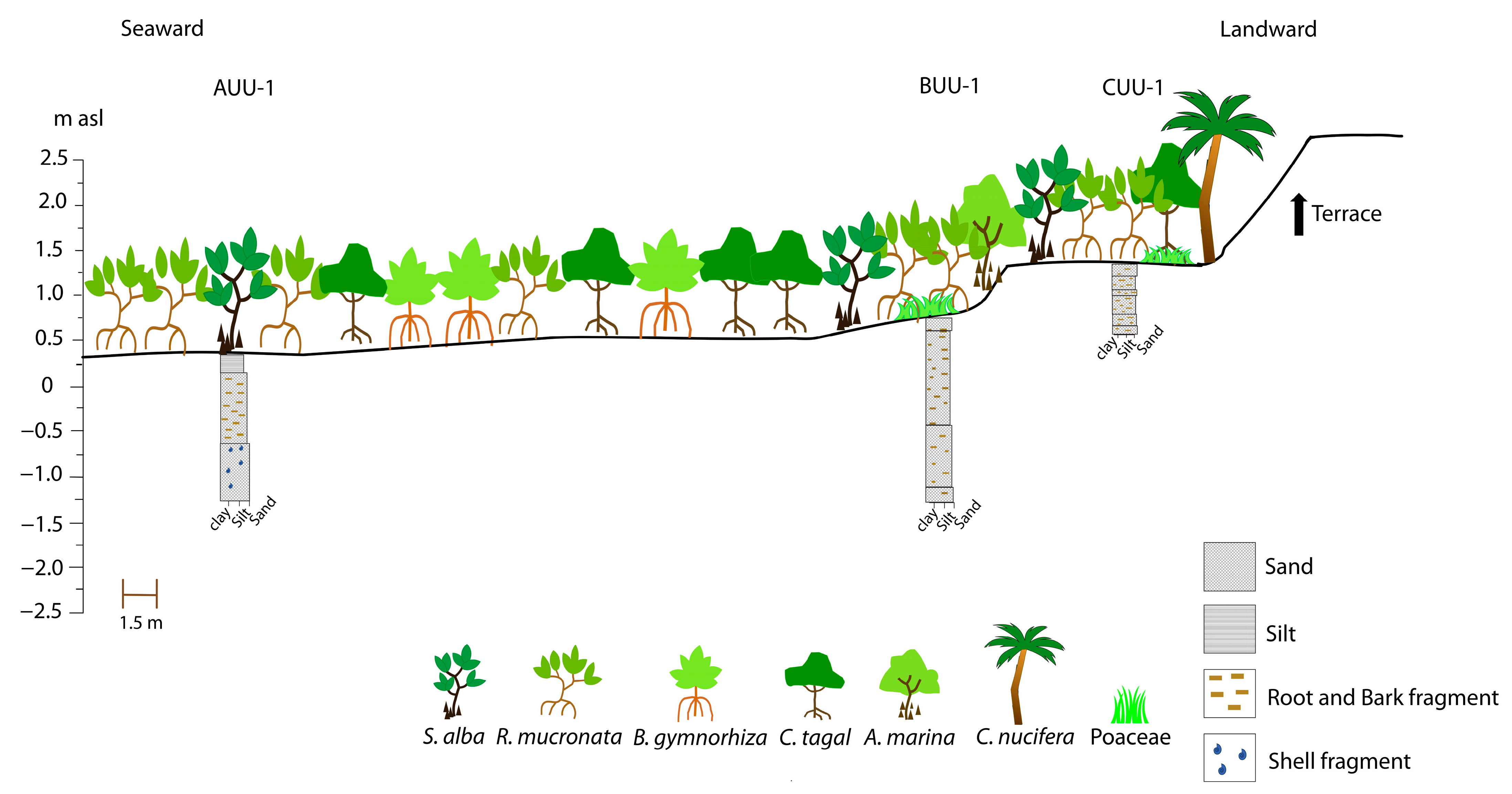
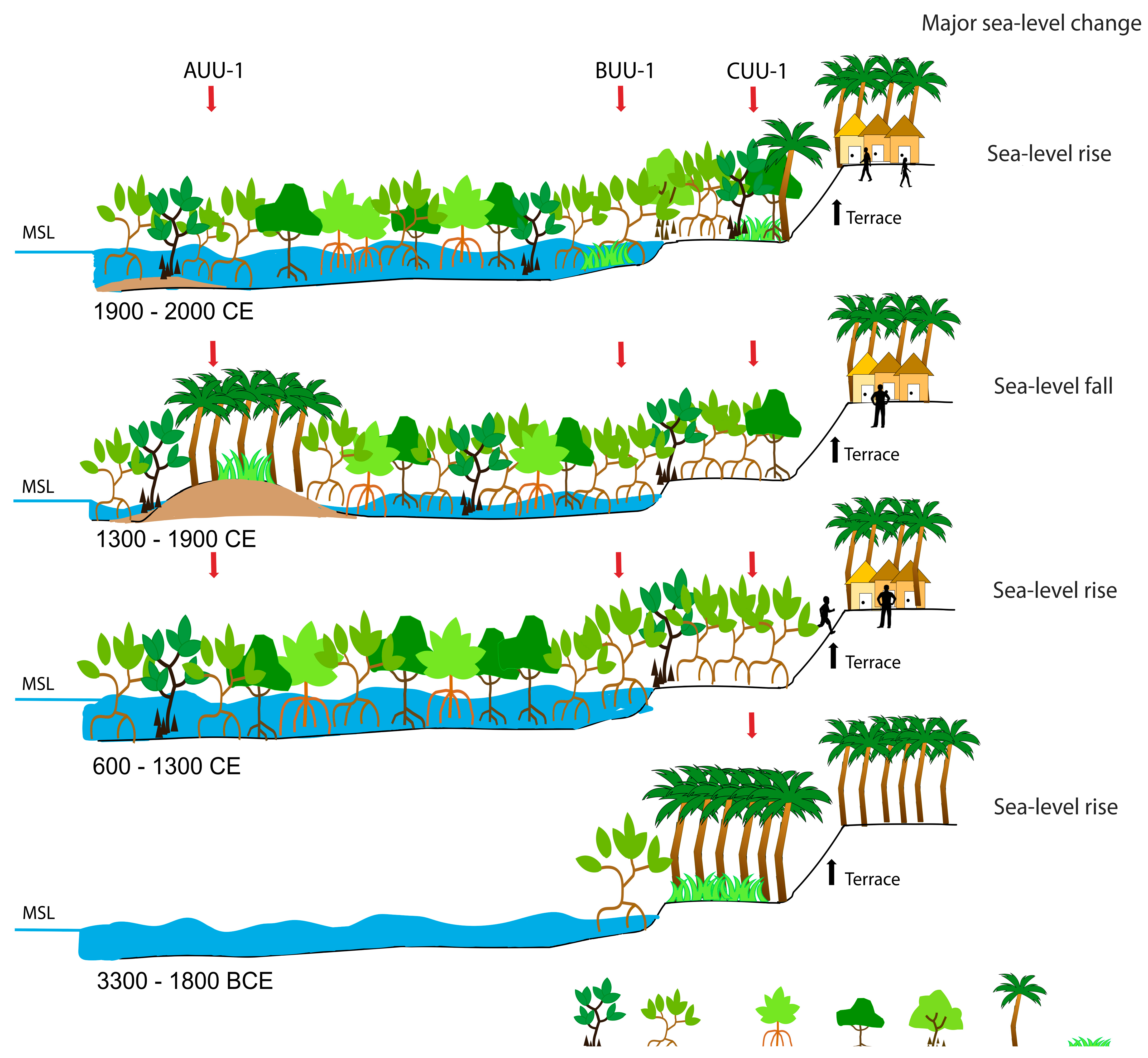
3.1. Vegetation Survey and Mangrove Zonation in Unguja Ukuu, Zanzibar Island
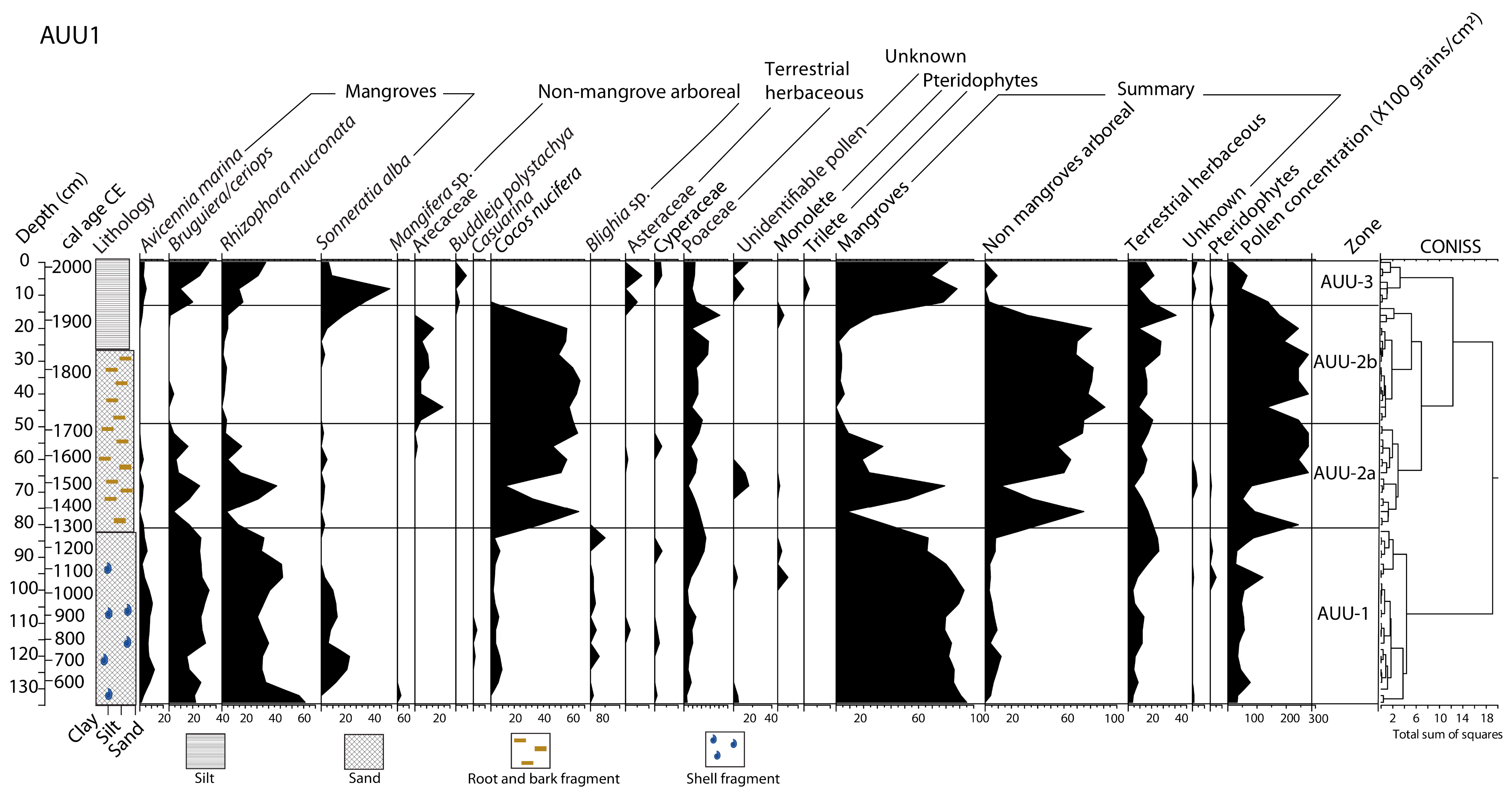
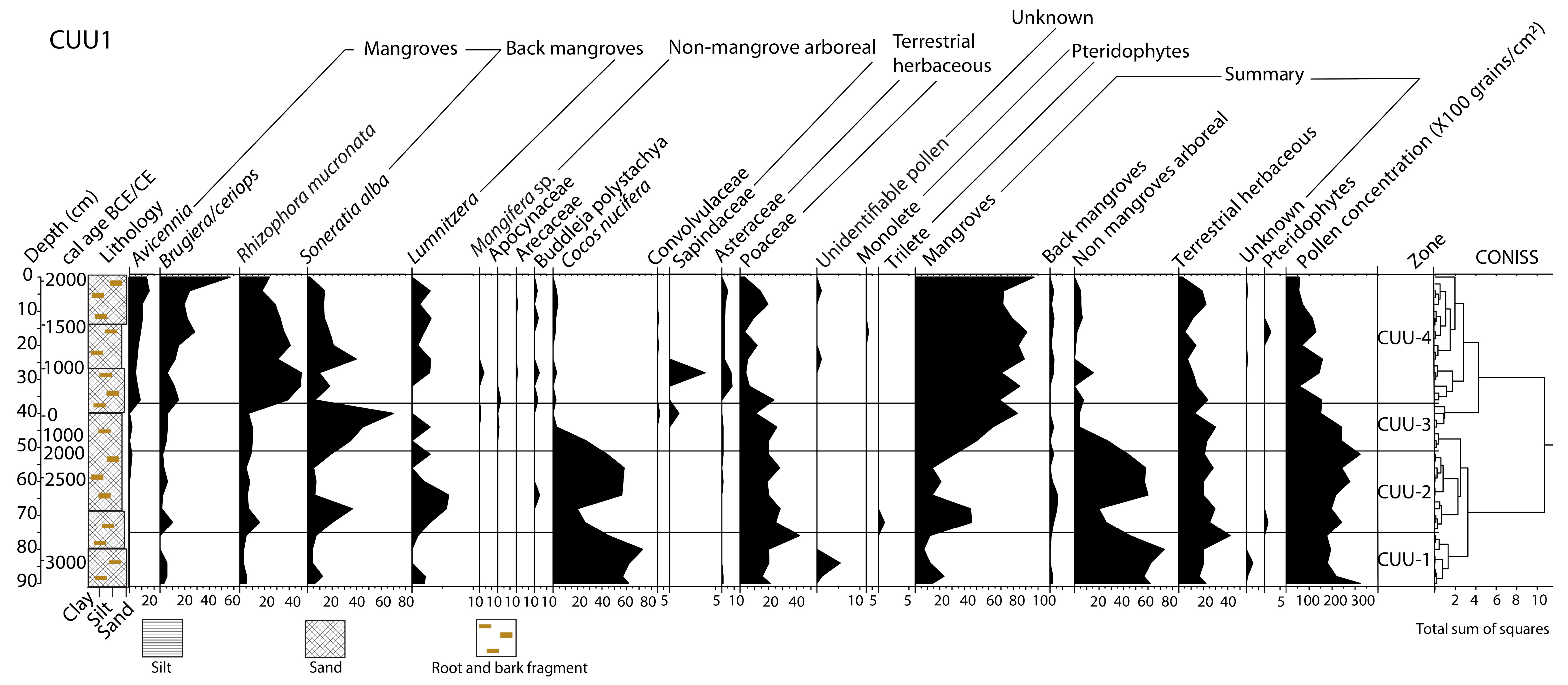
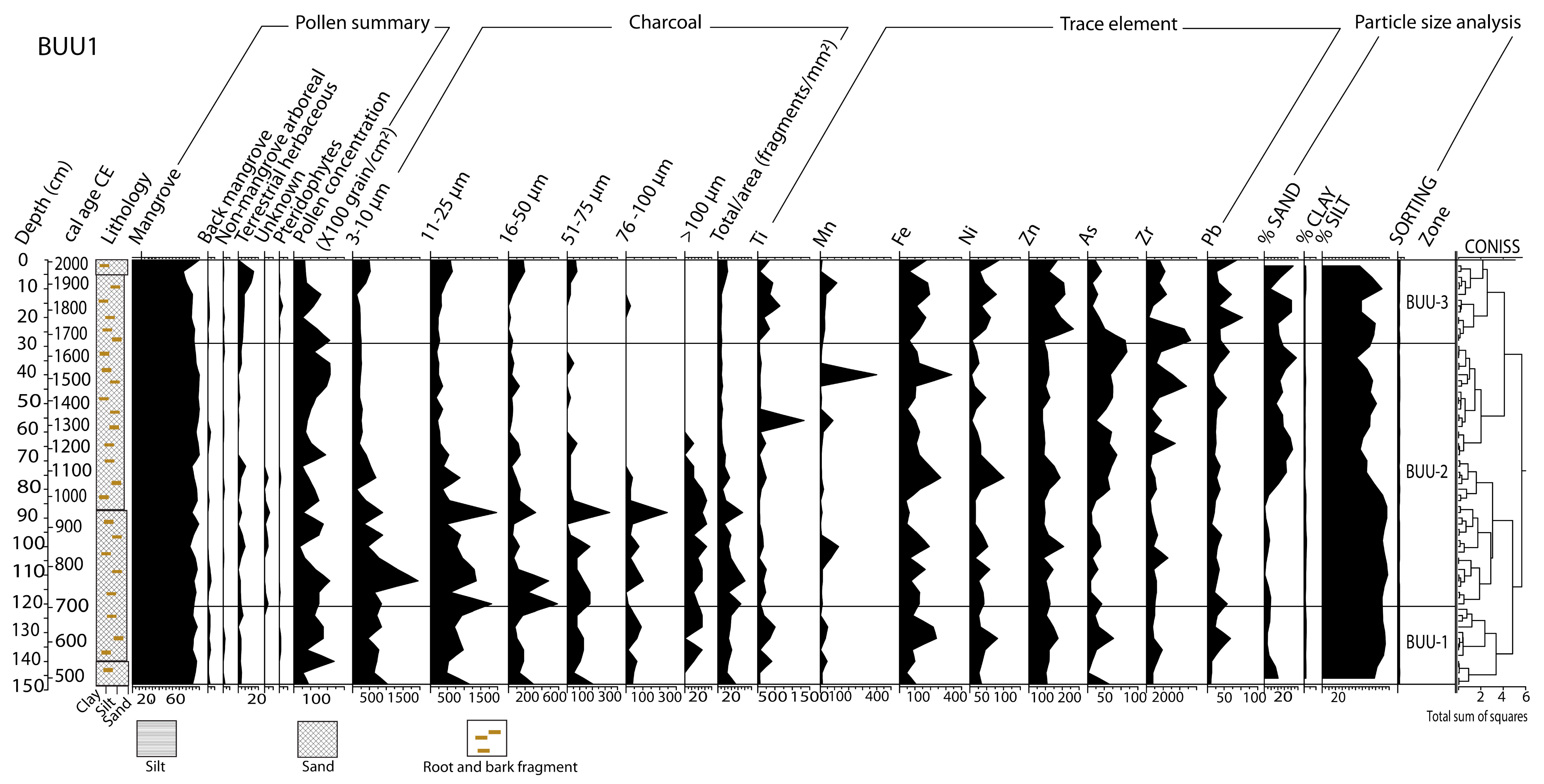
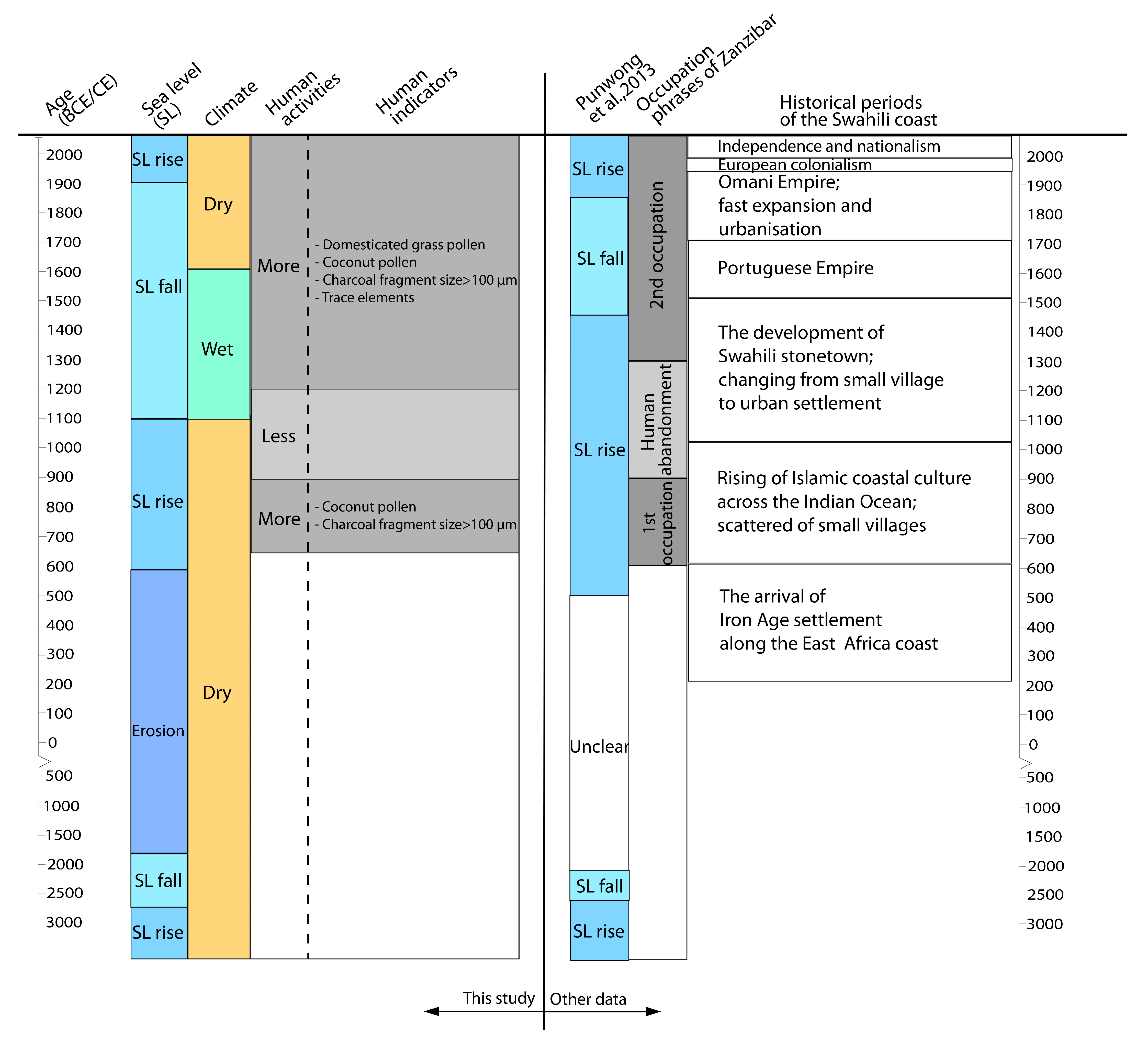
3.2. Core Chronological Results and Sedimentation Rates
3.3. Stratigraphy, Particle-Size and Trace Elements Analyses
3.4. Pollen Records
3.5. Microcharcoal Records
4. Interpretation and Discussion
4.1. ~3300 BCE–600 CE (5250–1350 cal yr BP)
4.2. ~600–950 CE (1350–1000 cal yr BP)
4.3. ~950–1300 CE (1000–650 cal yr BP)
4.4. ~1300 CE to the Present Day
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spalding, M. World Atlas of Mangroves; Routledge: London, UK, 2010; p. 336. [Google Scholar]
- Marchio, D.; Savarese, M.; Bovard, B.; Mitsch, W. Carbon Sequestration and Sedimentation in Mangrove Swamps Influenced by Hydrogeomorphic Conditions and Urbanization in Southwest Florida. Forests 2016, 7, 116. [Google Scholar] [CrossRef]
- Matos, C.R.L.; Berrêdo, J.F.; Machado, W.; Sanders, C.J.; Metzger, E.; Cohen, M.C.L. Carbon and Nutrient Accumulation in Tropical Mangrove Creeks, Amazon Region. Mar. Geol. 2020, 429, 106317. [Google Scholar] [CrossRef]
- de Jong Cleyndert, G.; Cuni-Sanchez, A.; Seki, H.A.; Shirima, D.D.; Munishi, P.K.T.; Burgess, N.; Calders, K.; Marchant, R. The Effects of Seaward Distance on above and below Ground Carbon Stocks in Estuarine Mangrove Ecosystems. Carbon Balance Manag. 2020, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Ellison, J.C. Vulnerability Assessment of Mangroves to Climate Change and Sea-Level Rise Impacts. Wetl. Ecol. Manag. 2015, 23, 115–137. [Google Scholar] [CrossRef]
- Ellison, A.M.; Alexander, J.; Daniel, A. Mangrove rehabilitation and restoration as experimental adaptive management. Front. Mar. Sci. 2020, 7, 327. [Google Scholar] [CrossRef]
- Ellison, J.C. Long-Term Retrospection on Mangrove Development Using Sediment Cores and Pollen Analysis: A Review. Aquat. Bot. 2008, 89, 93–104. [Google Scholar] [CrossRef]
- Cohen, M.C.L.; Figueiredo, B.L.; Oliveira, N.N.; Fontes, N.A.; França, M.C.; Pessenda, L.C.R.; Souza, A.V.; Macario, K.; Giannini, P.C.F.; Bendassolli, J.A.; et al. Impacts of Holocene and Modern Sea-level Changes on Estuarine Mangroves from Northeastern Brazil. Earth Surf. Process. Landf. 2019, 45, 375–392. [Google Scholar] [CrossRef]
- Punwong, P.; Marchant, R.; Selby, K. Holocene Mangrove Dynamics from Unguja Ukuu, Zanzibar. Quat. Int. 2013, 298, 4–19. [Google Scholar] [CrossRef]
- Punwong, P.; Marchant, R.; Selby, K. Holocene Mangrove Dynamics in Makoba Bay, Zanzibar. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 379–380, 54–67. [Google Scholar] [CrossRef]
- Punwong, P.; Marchant, R.; Selby, K. Holocene Mangrove Dynamics and Environmental Change in the Rufiji Delta, Tanzania. Veget. Hist. Archaeobot. 2013, 22, 381–396. [Google Scholar] [CrossRef]
- Gilman, E.L.; Ellison, J.; Duke, N.C.; Field, C. Threats to Mangroves from Climate Change and Adaptation Options: A Review. Aquat. Bot. 2008, 89, 237–250. [Google Scholar] [CrossRef]
- Grinsted, A.; Moore, J.C.; Jevrejeva, S. Reconstructing Sea Level from Paleo and Projected Temperatures 200 to 2100 Ad. Clim. Dyn. 2010, 34, 461–472. [Google Scholar] [CrossRef]
- Horton, B.P.; Khan, N.S.; Cahill, N.; Lee, J.S.H.; Shaw, T.A.; Garner, A.J.; Kemp, A.C.; Engelhart, S.E.; Rahmstorf, S. Estimating Global Mean Sea-Level Rise and Its Uncertainties by 2100 and 2300 from an Expert Survey. NPJ Clim. Atmos. Sci. 2020, 3, 18. [Google Scholar] [CrossRef]
- Jaritz, W. Das Quartär im Küstengebiet von Moçambique und seine Schwermineralführung. Geologisches Jahrbuch B 1977, 26, 3–93. [Google Scholar]
- Ramsay, P.J. 9000 years of sea-level change along the southern African coastline. Quat. Int. 1996, 31, 71–75. [Google Scholar] [CrossRef]
- Compton, J.S. Holocene Sea-Level Fluctuations Inferred from the Evolution of Depositional Environments of the Southern Langebaan Lagoon Salt Marsh, South Africa. Holocene 2001, 11, 395–405. [Google Scholar] [CrossRef]
- Ramsay, P.J.; Cooper, J.A.G. Late Quaternary Sea—Level Change in South Africa. Quat. Res. 2002, 57, 82–90. [Google Scholar] [CrossRef]
- Camoin, G.F.; Montaggioni, L.F.; Braithwaite, C.J.R. Late Glacial to Post Glacial Sea Levels in the Western Indian Ocean. Mar. Geol. 2004, 206, 119–146. [Google Scholar] [CrossRef]
- Muzuka, A.N.N.; Nyandwi, N. Preliminary Investigation on the Pleistocene/Holocene Sea-Level Changes along the Coastline of Tanzania, with Reference to Unguja and Pemba Islands. Boletim Geológico 2004, 43, 8–13. [Google Scholar]
- Strachan, K.L.; Finch, J.M.; Hill, T.; Barnett, R.L. A Late Holocene Sea-Level Curve for the East Coast of South Africa. S. Afr. J. Sci. 2014, 110, 1–9. [Google Scholar] [CrossRef]
- Woodroffe, S.A.; Long, A.J.; Milne, G.A.; Bryant, C.L.; Thomas, A.L. New Constraints on Late Holocene Eustatic Sea-Level Changes from Mahé, Seychelles. Quat. Sci. Rev. 2015, 115, 1–16. [Google Scholar] [CrossRef]
- Ellis, E.C. Anthropogenic Transformation of the Terrestrial Biosphere. Philos. Trans. Royal Soc. A 2011, 369, 1010–1035. [Google Scholar] [CrossRef]
- Crutzen, P.J. Geology of Mankind. In A Pioneer on Atmospheric Chemistry and Climate Change in the Anthropocene; Crutzen, P.J., Brauch, H., Crutzen, P.J., Eds.; Springer: Cham, Switzerland, 2016; Volume 50, pp. 211–215. [Google Scholar]
- Codron, D.; Lee-Thorp, J.A.; Sponheimer, M.; Codron, J. Nutritional Content of Savanna Plant Foods: Implications for Browser/Grazer Models of Ungulate Diversification. Eur. J. Wildl. Res. 2007, 53, 100–111. [Google Scholar] [CrossRef]
- Henry, A.G.; Hutschenreuther, A.; Paine, O.C.C.; Leichleiter, J.; Codron, D.; Codron, J.; Loudon, J.; Adolph, S.; Sponheimer, M. Influences on Plant Nutritional Variation and Their Potential Effects on Hominin Diet Selection. Rev. Palaeobot. Palynol. 2019, 261, 18–30. [Google Scholar] [CrossRef]
- Revelles, J.; Allué, E.; Alcolea, M.; Antolín, F.; Berihuete-Azorín, M.; Expósito, I.; Garay, B.; Mas, B.; Piqué, R.; Obea, L. Site Formation Processes, Human Activities and Palaeoenvironmental Reconstructions from Archaeobotanical Records in Cave and Rock-Shelter Sites in NE Iberia. Rev. Palaeobot. Palynol. 2022, 299, 104612. [Google Scholar] [CrossRef]
- Marchant, R. East Africa’s Human Environment Interactions: Historical Perspectives for a Sustainable Future; Springer Nature: Cham, Switzerland, 2021. [Google Scholar]
- Church, J.A.; Clark, P.U.; Cazenave, A.; Gregory, J.M.; Jevrejeva, S.; Levermann, A.; Merrifield, M.A.; Milne, G.A.; Nerem, R.S.; Nunn, P.D.; et al. Chapter 13: Sea Level Change. In Climate Change 2013: The Physical Science Basis: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; 1535p. [Google Scholar]
- Khojasteh, D.; Glamore, W.; Heimhuber, V.; Felder, S. Sea level rise impacts on estuarine dynamics: A review. Sci. Total Environ. 2021, 780, 146470. [Google Scholar] [CrossRef]
- Juma, A. Unguja Ukuu on Zanzibar: An Archaeological Study of Early Urbanism. Ph.D. Thesis, Uppsala University, Uppsala, Sweden, 2004. [Google Scholar]
- Hardy, A.; Mageni, Z.; Dongus, S.; Killeen, G.; Macklin, M.G.; Majambare, S.; Ali, A.; Msellem, M.; Al-Mafazy, A.-W.; Smith, M.; et al. Mapping Hotspots of Malaria Transmission from Pre-Existing Hydrology, Geology and Geomorphology Data in the Pre-Elimination Context of Zanzibar, United Republic of Tanzania. Parasites Vectors 2015, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Knopp, S.; Mohammed, K.A.; Khamis, I.S.; Mgeni, A.F.; Stothard, J.R.; Rollinson, D.; Marti, H.; Utzinger, J. Spatial Distribution of Soil-Transmitted Helminths, Including Strongyloides Stercoralis, among Children in Zanzibar. Geospat. Health 2008, 3, 47–56. [Google Scholar] [CrossRef]
- Mwandya, A.W.; Gullström, M.; Andersson, M.H.; Öhman, M.C.; Mgaya, Y.D.; Bryceson, I. Spatial and Seasonal Variations of Fish Assemblages in Mangrove Creek Systems in Zanzibar (Tanzania). Estuar. Coast. Shelf Sci. 2010, 89, 277–286. [Google Scholar] [CrossRef]
- Mkumbukwa, A.R. The History of Use and Conservation of Marine Resources in Zanzibar: Nineteenth Century to the Present. Ph.D. Thesis, Universitaet Bayreuth, Bayreuth, Germany, 2017. [Google Scholar]
- Sulas, F.; Kristiansen, S.M.; Wynne-Jones, S. Soil geochemistry, phytoliths and artefacts from an early Swahili daub house, Unguja Ukuu, Zanzibar. J. Archaeol. Sci. 2019, 103, 32–45. [Google Scholar] [CrossRef]
- Shunula, J. Public Awareness, Key to Mangrove Management and Conservation: The Case of Zanzibar. Trees 2002, 16, 209–212. [Google Scholar] [CrossRef]
- Horton, M.C.; Clark, C.M. Archaeological Survey of Zanzibar. Azania Archaeol. Res. Afr. 1985, 20, 167–171. [Google Scholar] [CrossRef]
- Crowther, A.; Veall, M.-A.; Boivin, N.; Horton, M.; Kotarba-Morley, A.; Fuller, D.Q.; Fenn, T.; Haji, O.; Matheson, C.D. Use of Zanzibar Copal (Hymenaea Verrucosa Gaertn.) as Incense at Unguja Ukuu, Tanzania in the 7–8th Century CE: Chemical Insights into Trade and Indian Ocean Interactions. J. Archaeol. Sci. 2015, 53, 374–390. [Google Scholar] [CrossRef]
- Fitton, T.; Wynne-Jones, S. Understanding the Layout of Early Coastal Settlement at Unguja Ukuu, Zanzibar. Antiquity 2017, 91, 1268–1284. [Google Scholar] [CrossRef]
- Walshaw, S.C. Converting to Rice: Urbanization, Islamization and Crops on Pemba Island, Tanzania, AD 700–1500. World Archaeol. 2010, 42, 137–154. [Google Scholar] [CrossRef]
- Wood, M.; Panighello, S.; Orsega, E.F.; Robertshaw, P.; van Elteren, J.T.; Crowther, A.; Horton, M.; Boivin, N. Zanzibar and Indian Ocean trade in the first millennium CE: The glass bead evidence. Archaeol. Anthropol. Sci. 2017, 9, 879–901. [Google Scholar] [CrossRef]
- Kessy, E. Iron Age Settlement Patterns and Economic Change on Zanzibar and Pemba Islands. In East African Archaeology: Foragers, Potters, Smiths, and Traders; Kusimba, C.M., Kusimba, S.B., Eds.; University of Pennsylvania Press: Philadelphia, PA, USA, 2003; pp. 117–132. [Google Scholar]
- Troels-Smith, J. Characterization of Unconsolidated Sediments. In CA Reitzels Forlag, Axel Sandal: Geological Survey of Denmark IV; CA Reitzels Forlag: Copenhagen, Denmark, 1955; Volume 3, pp. 38–71. [Google Scholar]
- Kershaw, A.P. A modification of the Troels-Smith system of sediment description and portrayal. Quat. Int. 1997, 15, 63–68. [Google Scholar]
- Erdtman, G. Handbook of Palynology: Morphology, Taxonomy, Ecology; Hafner: New York, NY, USA, 1969. [Google Scholar]
- Faegri, K.; Iversen, J. Text Book of Pollen Analysis; John Wiley & Sons Ltd.: Chichester, UK, 1989. [Google Scholar]
- Hunt, C.O. Recent advances in pollen extraction techniques: A brief review. Palaeobiol. Investig. 1985, 266, 181–187. [Google Scholar]
- Bonny, A.P. A method for determining absolute pollen frequencies in lake sediments. New Phytol. 1972, 72, 393–405. [Google Scholar] [CrossRef]
- Smol, J.P.; Birks, H.J.; Last, W.M. Tracking Environmental Change Using Lake Sediments Volume 3: Terrestrial, Algal, and Siliceous Indicators; Kluwer Academic: New York, NY, USA; Boston, MA, USA; Dordrecht, The Netherlands; London, UK; Moscow, Russia, 2001; Volume 3. [Google Scholar]
- Watson, J. Mangrove forests of the Malay Peninsula. Malay For. Rec. 1928, 6, 1–275. [Google Scholar]
- Santisuk, T. Taxonomy and distribution of terrestrial trees and shrubs in the mangrove formations in Thailand. Nat. Hist. Bull. Siam. Soc. 1983, 31, 63–91. [Google Scholar]
- Grimm, E.C. TGView 2.1.1; Illinois State Museum: Springfield, IL, USA, 2018. [Google Scholar]
- Madella, M.; Powers-Jones, A.H.; Jones, M.K. A Simple Method of Extraction of Opal Phytoliths from Sediments Using a Non-Toxic Heavy Liquid. J. Archaeol. Sci. 1998, 25, 801–803. [Google Scholar] [CrossRef]
- Tinner, W.; Hu, F.S. Size Parameters, Size-Class Distribution and Area-Number Relationship of Microscopic Charcoal: Relevance for Fire Reconstruction. Holocene 2003, 13, 499–505. [Google Scholar] [CrossRef]
- Rucina, S.M.; Muiruri, V.M.; Kinyanjui, R.N.; McGuiness, K.; Marchant, R. Late Quaternary Vegetation and Fire Dynamics on Mount Kenya. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 283, 1–14. [Google Scholar] [CrossRef]
- Clark, J.S. Particle motion and the theory of stratigraphic charcoal analysis: Source area, transport, deposition, and sampling. Quat. Res. 1988, 30, 67–80. [Google Scholar] [CrossRef]
- Solé, V.A.; Papillon, E.; Cotte, M.; Walter, P.; Susini, J. A Multiplatform Code for the Analysis of Energy-Dispersive X-Ray Fluorescence Spectra. Spectrochim. Acta Part B At. Spectrosc. 2007, 62, 63–68. [Google Scholar] [CrossRef]
- Brock, F.; Higham, T.; Ditchfield, P.; Ramsey, C.B. Current Pretreatment Methods for AMS Radiocarbon Dating at the Oxford Radiocarbon Accelerator Unit (Orau). Radiocarbon 2010, 52, 103–112. [Google Scholar] [CrossRef]
- Blaauw, M.J.; Christen, A.; Aquino-Lopez, M.A. rbacon: Age-Depth Modelling Using Bayesian Statistics. R Package Version 2.5.5. 2011. Available online: https://CRAN.R-project.org/package=rbacon (accessed on 20 January 2022).
- Blaauw, M.; Christen, J.A. Bacon Manual v2. 3.3, Queens University: Belfast, UK, 2013.
- R Core Team. A Language and Environment for Statistical Computing (Version 3.0. 1). R Foundation for Statistical Computing. 2015. Available online: http://www.R-project.org (accessed on 20 January 2022).
- Hogg, A.G.; Heaton, T.J.; Hua, Q.; Palmer, J.G.; Turney, C.S.; Southon, J.; Bayliss, A.; Blackwell, P.G.; Boswijk, G.; Bronk Ramsey, C.; et al. SHCal20 Southern Hemisphere Calibration, 0–55,000 Years Cal BP. Radiocarbon 2020, 62, 759–778. [Google Scholar] [CrossRef]
- Somboon, J.R.P. Palynological study of mangrove and marine sediments of the Gulf of Thailand. J. Southeast Asian Earth Sci. 1990, 4, 85–97. [Google Scholar] [CrossRef]
- Van Campo, E.; Bengo, M.D. Mangrove Palynology in Recent Marine Sediments off Cameroon. Mar. Geol. 2004, 208, 315–330. [Google Scholar] [CrossRef]
- Englong, A.; Punwong, P.; Selby, K.; Marchant, R.; Traiperm, P.; Pumijumnong, N. Mangrove Dynamics and Environmental Changes on Koh Chang, Thailand during the Last Millennium. Quat. Int. 2019, 500, 128–138. [Google Scholar] [CrossRef]
- Herzschuh, U.; Birks, H.J.B. Evaluating the Indicator Value of Tibetan Pollen Taxa for Modern Vegetation and Climate. Rev. Palaeobot. Palynol. 2010, 160, 197–208. [Google Scholar] [CrossRef]
- Cao, X.; Tian, F.; Li, K.; Ni, J.; Yu, X.; Liu, L.; Wang, N. Lake Surface-Sediment Pollen Dataset for the Alpine Meadow Vegetation Type from the Eastern Tibetan Plateau and Its Potential in Past Climate Reconstructions. Earth Syst. Sci. 2021, 13, 3525–3537. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Liu, L.; Wang, N.; Cao, X. No Evidence of Human Disturbance to Vegetation in the Zoige Region (North-Eastern Tibetan Plateau) in the Last Millennium until Recent Decades. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 589, 110843. [Google Scholar] [CrossRef]
- Biagioni, S.; Haberzettl, T.; Wang, L.-C.; St-Onge, G.; Behling, H. Unravelling the Past 1000 Years of History of Human–Climate–Landscape Interactions at the Lindu Plain, Sulawesi, Indonesia. Veget. Hist. Archaeobot. 2016, 25, 1–17. [Google Scholar] [CrossRef]
- Githumbi, E.N.; Kariuki, R.; Shoemaker, A.; Courtney-Mustaphi, C.J.; Chuhilla, M.; Richer, S.; Lane, P.; Marchant, R. Pollen, People and Place: Multidisciplinary Perspectives on Ecosystem Change at Amboseli, Kenya. Front. Earth Sci. 2018, 5, 113. [Google Scholar] [CrossRef]
- Van der Plas, G.W.; Rucina, S.M.; Hemp, A.; Marchant, R.A.; Hooghiemstra, H.; Schüler, L.; Verschuren, D. Climate-Human-Landscape Interaction in the Eastern Foothills of Mt. Kilimanjaro (Equatorial East Africa) during the Last Two Millennia. Holocene 2021, 31, 556–569. [Google Scholar] [CrossRef]
- Punwong, P.; Promplin, S.; Lomchantrasilp, C.; Soonthornampaipong, P.; Englong, A.; Marchant, R.; Selby, K.; Chirawatkul, P. Documenting a Thousand Years of Environmental and Anthropogenic Changes on Mangroves on the Bangkok Coast, the Upper Gulf of Thailand. Veget. Hist. Archaeobot. 2023, 32, 17–34. [Google Scholar] [CrossRef]
- Eskola, T.; Kontio, R.; Lunkka, J.P. Comparison between modified LST Fastfloat and conventional HF methods for pollen preparation in highly minerogenic sediments. Bull. Geol. Soc. Finl. 2021, 93, 5–8. [Google Scholar] [CrossRef]
- González, C.; Urrego, L.E.; Martínez, J.I.; Polanía, J.; Yokoyama, Y. Mangrove Dynamics in the Southwestern Caribbean since the ‘Little Ice Age’: A History of Human and Natural Disturbances. Holocene 2010, 20, 849–861. [Google Scholar] [CrossRef]
- Harries, H.; Baudouin, L.; Cardena, R. Floating, Boating and Introgression: Molecular Techniques and the Ancestry of Coconut Palm Populations on Pacific Islands. Ethnobot. Res. App. 2004, 2, 037–053. [Google Scholar] [CrossRef]
- Athens, J.S.; Ward, J.V.; Blinn, D.W. Vegetation History of Laysan Island, Northwestern Hawaiian Islands. Pac. Sci. 2007, 61, 17–37. [Google Scholar] [CrossRef]
- Ellison, J.C. Palaeo-lake and Swamp Stratigraphic Records of Holocene Vegetation and Sea-level Changes, Mangaia, Cook Islands. Pac. Sci. 1994, 48, 1–15. [Google Scholar]
- Athens, J.S.; Ward, J.V. Paleoenvironment of the Orote Peninsula, Guam. Micronesica 1995, 28, 205–223. [Google Scholar]
- Marchant, R.; Richer, S.; Boles, O.; Capitani, C.; Courtney-Mustaphi, C.J.; Lane, P.; Prendergast, M.E.; Stump, D.; de Cort, G.; Kaplan, J.O.; et al. Drivers and trajectories of land cover change in East Africa: Human and environmental interactions from 6000 years ago to present. Earth-Sci. Rev. 2018, 178, 322–378. [Google Scholar] [CrossRef]
- Chritz, K.L.; Cerling, T.E.; Freeman, K.H.; Hildebrand, E.A.; Janzen, A.; Prendergast, M.E. Climate, Ecology, and the Spread of Herding in Eastern Africa. Quat. Sci. Rev. 2019, 204, 119–132. [Google Scholar] [CrossRef]
- Opiyo, B.; Gebregiorgis, D.; Cheruiyot, V.C.; Deocampo, D.M.; Kiage, L.M. Late Quaternary Paleoenvironmental Changes in Tropical Eastern Africa Revealed by Multi–Proxy Records from the Cherangani Hills, Kenya. Quat. Sci. Rev. 2019, 222, 105907. [Google Scholar] [CrossRef]
- Punwong, P.; Selby, K.; Marchant, R. Holocene Mangrove Dynamics and Relative Sea-Level Changes along the Tanzanian Coast, East Africa. Estuar. Coast. Shelf Sci. 2018, 212, 105–117. [Google Scholar] [CrossRef]
- Whitlock, C.; Larsen, C. Charcoal as a Fire Proxy. In Tracking Environmental Change Using Lake Sediments; Smol, J.P., Birks, H.J.B., Last, W.M., Bradley, R.S., Alverson, K., Eds.; Developments in Paleoenvironmental Research; Springer Netherlands: Dordrecht, The Netherlands, 2002; Volume 3, pp. 75–97. [Google Scholar]
- Msaky, E.S.; Livingstone, D.; Davis, O.K. Paleolimnological Investigations of Anthropogenic Environmental Change in Lake Tanganyika: V. Palynological Evidence for Deforestation and Increased Erosion. J. Paleolimnol. 2005, 34, 73–83. [Google Scholar] [CrossRef]
- Bloszies, C.; Forman, S.L.; Wright, D.K. Water Level History for Lake Turkana, Kenya in the Past 15,000years and a Variable Transition from the African Humid Period to Holocene Aridity. Glob. Planet Change 2015, 132, 64–76. [Google Scholar] [CrossRef]
- Tomlinson, P.B. The Botany of Mangroves, 2nd ed.; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Miller, D.E.; Yates, R.J.; Jerardino, A.; Parkington, J.E. Late Holocene Coastal Change in the Southwestern Cape, South Africa. Quat. Int. 1995, 29–30, 3–10. [Google Scholar] [CrossRef]
- Khan, N.S.; Ashe, E.; Shaw, T.A.; Vacchi, M.; Walker, J.; Peltier, W.R.; Kopp, R.E.; Horton, B.P. Holocene relative sea-level changes from near-, intermediate-, and far-field locations. Curr. Clim. Change Rep. 2015, 1, 247–262. [Google Scholar] [CrossRef]
- Liu, X.; Rendle-Bühring, R.; Henrich, R. Geochemical Composition of Tanzanian Shelf Sediments Indicates Holocene Climatic and Sea-Level Changes. Quat. Res. 2017, 87, 442–454. [Google Scholar] [CrossRef]
- Njagi, D.M.; Routh, J.; Olago, D.; Gayantha, K. A Multi-Proxy Reconstruction of the Late Holocene Climate Evolution in the Kapsabet Swamp, Kenya (East Africa). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 574, 110475. [Google Scholar] [CrossRef]
- Chami, F.A. Roman Beads from the Rufiji Delta, Tanzania: First Incontrovertible Archaeological Link with the Periplus. Curr. Anthropol. 1999, 40, 237–242. [Google Scholar] [CrossRef]
- Wynne-Jones, E.S.; LaViolette, A. The Swahili World; Routledge: London, UK; New York, NY, USA, 2018; p. 702. [Google Scholar]
- Allibert, C. Austronesian Migration and the Establishment of the Malagasy Civilization: Contrasted Readings in Linguistics, Archaeology, Genetics and Cultural Anthropology. Diogenes 2008, 55, 7–16. [Google Scholar] [CrossRef]
- Beaujard, P. The First Migrants to Madagascar and Their Introduction of Plants: Linguistic and Ethnological Evidence. Azania Archaeol. Res. Afr. 2011, 46, 169–189. [Google Scholar] [CrossRef]
- Boivin, N.; Crowther, A.; Helm, R.; Fuller, D.Q. East Africa and Madagascar in the Indian Ocean World. J. World Prehist. 2013, 26, 213–281. [Google Scholar] [CrossRef]
- Prendergast, M.E.; Rouby, H.; Punnwong, P.; Marchant, R.; Crowther, A.; Kourampas, N.; Shipton, C.; Walsh, M.; Lambeck, K.; Boivin, N.L. Continental Island Formation and the Archaeology of Defaunation on Zanzibar, Eastern Africa. PLoS ONE 2016, 11, e0149565. [Google Scholar] [CrossRef]
- Faulkner, P.; Harris, M.; Ali, A.K.; Haji, O.; Crowther, A.; Horton, M.C.; Boivin, N.L. Characterising Marine Mollusc Exploitation in the Eastern African Iron Age: Archaeomalacological Evidence from Unguja Ukuu and Fukuchani, Zanzibar. Quat. Int. 2018, 471, 66–80. [Google Scholar] [CrossRef]
- Kotarba-Morley, A.M.; Kourampas, N.; Morley, M.W.; MacAdams, C.; Crowther, A.; Faulkner, P.; Horton, M.; Boivin, N. Coastal Landscape Changes at Unguja Ukuu, Zanzibar: Contextualizing the Archaeology of an Early Islamic Port of Trade. J. Island Coast. Archaeol. 2022, 1–35. [Google Scholar] [CrossRef]
- Wynne-Jones, S.; Sulas, F.; Out, W.A.; Kristiansen, S.M.; Fitton, T.; Ali, A.K.; Olsen, J. Urban Chronology at a Human Scale on the Coast of East Africa in the 1st Millennium A. D. J. Field Archaeol. 2021, 46, 21–35. [Google Scholar] [CrossRef]
- Chami, F.A. Further Archaeological Research on Mafia Island. Azania Archaeol. Res. Afr. 2000, 35, 208–214. [Google Scholar] [CrossRef]
- Pollard, E. Settlement Adaptation to a Changing Coastline: Archaeological Evidence from Tanzania, during the First and Second Millennia AD. J. Island Coast. Archaeol. 2009, 4, 82–107. [Google Scholar] [CrossRef]
- Verschuren, D.; Laird, K.R.; Cumming, B.F. Rainfall and Drought in Equatorial East Africa during the Past 1100 Years. Nature 2000, 403, 410–414. [Google Scholar] [CrossRef]
- Åse, L. Studies of Shores and Shore Displacement on the Southern Coast of Kenya. Especially in Kilifi District. Phys. Geogr. 1981, 63, 303–310. [Google Scholar]
- Andreucci, S.; Pascucci, V.; Murray, A.S.; Clemmensen, L.B. Late Pleistocene Coastal Evolution of San Giovanni Di Sinis, West Sardinia (Western Mediterranean). Sediment Geol. 2009, 216, 104–116. [Google Scholar] [CrossRef]
- IPCC. Contribution of working groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2014: Synthesis Report; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Szwitzerland, 2014; p. 151. [Google Scholar]
- Dangendorf, S.; Marcos, M.; Wöppelmann, G.; Conrad, C.P.; Frederikse, T.; Riva, R. Reassessment of 20th Century Global Mean Sea Level Rise. Proc. Natl. Acad. Sci. USA 2017, 114, 5946–5951. [Google Scholar] [CrossRef]
- Nerem, R.S.; Beckley, B.D.; Fasullo, J.T.; Hamlington, B.D.; Masters, D.; Mitchum, G.T. Climate-Change–Driven Accelerated Sea-Level Rise Detected in the Altimeter Era. Proc. Natl. Acad. Sci. USA 2018, 115, 2022–2025. [Google Scholar] [CrossRef]
- Kiage, L.M.; Liu, K. Late Quaternary Paleoenvironmental Changes in East Africa: A Review of Multiproxy Evidence from Palynology, Lake Sediments, and Associated Records. Prog. Phys. Geogr. 2006, 30, 633–658. [Google Scholar] [CrossRef]
- Semesi, A.K. Mangrove Management and Utilization in Eastern Africa. Ambio 1998, 27, 620–626. [Google Scholar]
- Ngoile, M.A.K.; Shunula, J.P. Status and Exploitation of the Mangrove and Associated Fishery Resources in Zanzibar. Hydrobiologia 1992, 247, 229–234. [Google Scholar] [CrossRef]
- Crowther, A.; Faulkner, P.; Prendergast, M.E.; Quintana Morales, E.M.; Horton, M.; Wilmsen, E.; Kotarba-Morley, A.M.; Christie, A.; Petek, N.; Tibesasa, R.; et al. Coastal Subsistence, Maritime Trade, and the Colonization of Small Offshore Islands in Eastern African Prehistory. J. Island Coast. Archaeol. 2016, 11, 211–237. [Google Scholar] [CrossRef]
- Joly, C.; Barillé, L.; Barreau, M.; Mancheron, A.; Visset, L. Grain and Annulus Diameter as Criteria for Distinguishing Pollen Grains of Cereals from Wild Grasses. Rev. Palaeobot. Palynol. 2007, 146, 221–233. [Google Scholar] [CrossRef]
- Quamar, M.F. Late Holocene Vegetation Dynamics and Monsoonal Climatic Changes in Jammu, India. Acta Palaeobot. 2022, 62, 36–49. [Google Scholar] [CrossRef]
- Crowther, A.; Prendergast, M.E.; Fuller, D.Q.; Boivin, N. Subsistence Mosaics, Forager-Farmer Interactions, and the Transition to Food Production in Eastern Africa. Quat. Int. 2018, 489, 101–120. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and the Environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar]
- Shahid, M.; Ferrand, E.; Schreck, E.; Dumat, C. Behavior and Impact of Zirconium in the Soil–Plant System: Plant Uptake and Phytotoxicity. Rev. Environ. Contam. Toxicol. 2013, 221, 107–127. [Google Scholar] [PubMed]
- Masalu, D.C.P. Coastal and Marine Resource Use Conflicts and Sustainable Development in Tanzania. Ocean Coast. Manag. 2000, 43, 475–494. [Google Scholar] [CrossRef]
- He, Q.; Silliman, B.R. Climate Change, Human Impacts, and Coastal Ecosystems in the Anthropocene. Curr. Biol. 2019, 29, R1021–R1035. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Adame, M.F.; Bennion, V.; Hayes, M.; Reef, R.; Santini, N.; Cahoon, D.R. Sea Level and Turbidity Controls on Mangrove Soil Surface Elevation Change. Estuar. Coast. Shelf Sci. 2015, 153, 1–9. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Core | Depth (cm) | Altitude (m a.m.s.l.) | Codes | Sample Type | 14C Age | Min-Max Age Range (BCE/CE) | Median Age (BCE/CE) | Median Age (Cal yr BP) | Source |
|---|---|---|---|---|---|---|---|---|---|
| AUU1 | 56 | −0.52 | D-AMS 045311 | Uncharred plant | 218 ± 18 | 1507-1759 CE | 1673 CE | 277 | This study |
| 91 * | −0.81 | UBA-16626 | Bulk sediment | 169 ± 22 | 1672-1954 CE | 1813 CE | 137 | [9] | |
| 128 | −1.24 | D-AMS 045312 | Uncharred plant | 1469 ± 20 | 756-635 CE | 635 CE | 1315 | This study | |
| BUU1 | 88 | −0.71 | D-AMS 045313 | Charcoal | 1182 ± 20 | 883-999 CE | 949 CE | 1001 | This study |
| 138 | −1.21 | UBA-16627 | Bulk sediment | 1534 ± 23 | 441-631 CE | 565 CE | 1385 | [9] | |
| CUU1 | 36 | 1.02 | D-AMS 045314 | Uncharred plant | 1458 ± 20 | 545-758 CE | 64 CE | 1309 | This study |
| 55 | 0.83 | D-AMS 046339 | Charcoal, bark | 4158 ± 35 | 2664-1461 BCE | 2389 BCE | 4339 | This study | |
| 78 | 0.6 | UBA-16628 | Bulk sediment | 4211 ± 25 | 3190-2641 BCE | 2823 BCE | 4773 | [9] | |
| * 88 | 0.5 | D-AMS 045315 | Uncharred plant | 1506 ± 20 | 550-643 CE | - | - | This study |
| Group | Families, Genera or Species | AUU1 | BUU1 | CUU1 |
|---|---|---|---|---|
| Mangroves | Avicennia marina | + | + | + |
| Bruguiera/Ceriops | + | + | + | |
| Rhizophora mucronata | + | + | + | |
| Sonneratia alba | + | + | + | |
| Back mangroves | Acrostichum sp. | + | ||
| Lumnitzera sp. | + | + | ||
| Non-mangrove arboreal | Apocynaceae | + | ||
| Arecaceae | + | + | ||
| Blighia sp. | + | + | ||
| Buddleja polystachya | + | + | + | |
| Casuarina sp. | + | + | ||
| Cocos nucifera | + | + | ||
| Convolvulaceae | + | + | ||
| Mangifera sp. | + | + | ||
| Sapindaceae | + | + | + | |
| Tiliaceae | + | + | ||
| Terrestrial herbaceous | Asteraceae | + | + | + |
| Cichorioideae | + | |||
| Cyperaceae | + | + | + | |
| Poaceae | + | + | + | |
| Poaceae, >40–60 µm | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Englong, A.; Punwong, P.; Marchant, R.; Seelanan, T.; Wynne-Jones, S.; Chirawatkul, P. High-Resolution Multiproxy Record of Environmental Changes and Anthropogenic Activities at Unguja Ukuu, Zanzibar, Tanzania during the Last 5000 Years. Quaternary 2023, 6, 21. https://doi.org/10.3390/quat6010021
Englong A, Punwong P, Marchant R, Seelanan T, Wynne-Jones S, Chirawatkul P. High-Resolution Multiproxy Record of Environmental Changes and Anthropogenic Activities at Unguja Ukuu, Zanzibar, Tanzania during the Last 5000 Years. Quaternary. 2023; 6(1):21. https://doi.org/10.3390/quat6010021
Chicago/Turabian StyleEnglong, Apichaya, Paramita Punwong, Rob Marchant, Tosak Seelanan, Stephanie Wynne-Jones, and Prae Chirawatkul. 2023. "High-Resolution Multiproxy Record of Environmental Changes and Anthropogenic Activities at Unguja Ukuu, Zanzibar, Tanzania during the Last 5000 Years" Quaternary 6, no. 1: 21. https://doi.org/10.3390/quat6010021
APA StyleEnglong, A., Punwong, P., Marchant, R., Seelanan, T., Wynne-Jones, S., & Chirawatkul, P. (2023). High-Resolution Multiproxy Record of Environmental Changes and Anthropogenic Activities at Unguja Ukuu, Zanzibar, Tanzania during the Last 5000 Years. Quaternary, 6(1), 21. https://doi.org/10.3390/quat6010021