
Protein Extract of Tobacco Expressing Solanum torvum PP5-Encoding Gene Inhibits Verticillium dahliae Proliferation


College of Life Sciences, Nanjing Agricultural University, Nanjing 210095, China
*
Authors to whom correspondence should be addressed.
Horticulturae 2022, 8(3), 252; https://doi.org/10.3390/horticulturae8030252
Submission received: 29 January 2022
/
Revised: 14 March 2022
/
Accepted: 15 March 2022
/
Published: 16 March 2022
(This article belongs to the Collection Genetic Engineering and Quality Improvement in Vegetable Crops)
Abstract
:Verticillium wilt, a soilborne disease caused by Verticillium dahliae (V. dahliae), can severely affect the yields of Solanaceae crops. In a previous study, it was observed in Solanum torvum (S. torvum) that protein phosphatase 5 (PP5) was induced by V. dahliae infection. To elucidate the function of PP5 more clearly, this study cloned an StPP5 cDNA from S. torvum by PCR. The cDNA contained an ORF of 1458 bp long encoding a putative protein of 485 amino acid residues with a predicted molecular mass of 54.63 kDa and a theoretical isoelectric point of 5.66. StPP5 protein contained a conserved PP domain and showed high similarity to other homologous members of the PP5 family from various plant species. The expression of StPP5 gene was upregulated after V. infection and reached its maximum value at 24 h in leaves. In order to clarify the role of StPP5, four transgenic tobacco plants expressing StPP5 were generated through Agrobacterium-mediated transformation and identified by PCR. In vitro culture assay showed that the growth of V. dahliae in PDA medium containing proteins extracted from the leaves of transgenic tobacco line P6 was inhibited, whose inhibition rate was 55.1%, higher than the non-transgenic control. These results indicated that StPP5 might be involved in plant defense against V. dahliae infection.
1. Introduction
Eggplant (Solanum melongena L.), also known as aubergine or brinjal, has been widely cultivated in the world for centuries and is currently an important crop species [1]. However, the yields of eggplant are severely affected by certain soilborne diseases such as Verticillium wilt (VW), which is caused by the soil-inhabiting fungus Verticillium dahliae Kleb (V. dahliae) that has a worldwide distribution [2]. Once the plants are infected with V. dahliae, there are no effective available fungicides currently [3]. The effective methods are introducing genes from resistant germplasm into the eggplant cultivars to develop tolerant plants by breeding [4]. Therefore, transgenesis is utilized to achieve plants resistant to V. dahliae, but the effects have been varied [5,6]. Presently, some genes related to resistance to VW have been isolated in plants, but only a few genes have been analyzed in function. Ve1 and Ve2 are two resistance genes first isolated from tomato (Solanum lycopersicum L.) [7,8,9], but only Ve1 was valuable for application in breeding according to its resistance [8]. StoVe1, the homolog of Ve1, is a gene with higher resistance, which was cloned from Solanum torvum (Solanaceae), and the expression of the gene in potato enhanced resistance to V. dahliae [2,10]. In recent years, cotton transformed respectively with hpa1Xoo or GbVe genes conferred an improved resistance to V. dahlia [11,12].
Earlier studies discovered that alteration of the serine/threonine dephosphorylation state has significant effects on plant growth, which is induced by different types of mitogens [13]. Consequently, researchers cloned the gene encoding protein phosphatase from many plants such as Arabidopsis and Alfalfa, and certain proteins were purified [14,15]. In comparison with protein kinases, there are fewer studies about protein phosphatases of plants, and most of the cloned protein phosphatase genes have not been studied extensively. Protein phosphatase 5 (PP5), a member of the serine/threonine-specific protein phosphatase family, is widely distributed and highly conserved among eukaryotes. Subcellular localization showed that PP5 is expressed in both the nucleus and the cytoplasm [16].
Wild eggplant has a rich genetic resource of genes with high resistance to VW [17]. In the previous study, we established a V. dahliae-induced S. torvum cDNA library, including 118 related defense transcript-derived fragments (TDFs) [18]. The gene StPP5 was selected from this cDNA library. Here, we cloned and analyzed the StPP5 sequence from S. torvum and the inhibitory effect of the protein extract of tobacco, including StPP5 to V. dahliae proliferation, which indicates that StPP5 might be involved in plant defense against V. dahliae infection.
2. Materials and Methods
2.1. Plant Materials
S. torvum was grown in a greenhouse without temperature and light control. Young leaves and stems were collected from well-developed plants, frozen in liquid nitrogen immediately, and stored at −70 °C for gene cloning and expression analysis.
The seedlings of tobacco (Nicotiana tabacum L. cv. NC89) were produced by germinating sterilized seeds on Murashige and Skoog (MS) medium under conditions previously reported [19].
2.2. Isolation of Full-Length cDNA of StPP5
Total RNA was extracted from 0.1 g of fresh leaves with total RNA isolation reagent following the manufacturer’s instructions. First-strand cDNA was synthesized with M-MLV-reverse transcriptase from TaKaRa Biotech Company (Dalian, China) according to the manufacturer’s instructions.
To clone the conserved region of StPP5 cDNA, a pair of primers, P1 (5′-AAAGATGCACTCAAGGATTTTCAACAGGT-3′) and P2 (5′-GTTTACTAGGTCCTCTACCAGGCTGAGG-3′) (Figure 1A), were designed according to the conserved region of PP5 gene from other plants using the DNAssist 2.0 software. Polymerase chain reaction (PCR) was carried out according to the following program: firstly, incubation at 94 °C for 4 min, and then followed by 35 cycles (94 °C for 40 s, 60 °C for 40 s, 72 °C for 1 min), with a final extension step at 72 °C for 10 min. The PCR product was about 800 bp long.
To obtain 5′ end sequence, a primer P3 (5′-ATGCCCGGTATGGAAGCTGAG-3′) was designed according to 5′ end sequence of PP5 (accession no. AY182777) (Figure 1A). The PCR was carried out using P3 and P2 under the following PCR condition: firstly, incubation at 94 °C for 4 min, then followed by 35 cycles (94 °C for 30 s, 62 °C for 50 s, 72 °C for 40 s) and by extension at 72 °C for 10 min.
To amplify the 3′ end of StPP5, a primer P4 (5′-TTAAGAAAACATGCGAAGGAAGTTGTTGG-3′) was designed according to 3′ end sequence of PP5 (accession no. AY182777) (Figure 1A). The PCR was carried out using P1 and P4 under the following PCR condition: 94 °C for 4 min, followed by 35 cycles (94 °C for 30 s, 60 °C for 30 s, 72 °C for 30 s) and by extension at 72 °C for 10 min.
The full length of the gene cDNA was amplified with gene-specific primers P3 and P4 after assembling the full-length sequence of StPP5 (Figure 1A). PCR conditions were: 94 °C for 5 min, followed by 30 cycles (94 °C for 50 s, 60 °C for 40 s, 72 °C for 40 s) and with a final extension step at 72 °C for 10 min.
The volume of PCR amplification reactions was 25 μL, containing 10 × PCR Buffer, 2.5 μL; 10 mmol/L dNTP, 1 μL; forward and reverse primer, 1 μL; cDNA template, 0.5 μL; 5 U/μL Taq polymerase, 0.25 μL; double-distilled water, 16.75 μL.
PCR products were separated on 1% agarose gels and recovered by gel extraction. Then, the target products were introduced into pMD18-T vector from TaKaRa Biotech, and finally transformed into competent cells of Escherichia coli strain DH5α. The positive colonies were identified by PCR and sequenced.
2.3. Bioinformatics Analysis
The protein similarity of StPP5 was analyzed using DNAssist program. GenBank ORF finder (http://www.ncbi.nih.gov/gorf/gorf.html, accessed on 15 May 2019) was used for prediction of the open reading frame (ORF) of the sequence; phylogenetic analysis was performed using Mega 5.0. ProtScale (http://us.expasy.org/cgi-bin/protscale.pl, accessed on 27 January 2022) of ExPASy was used for prediction of protein hydrophobicity, and transmembrane domain was analyzed by TMHMM-2.0 (http://www.cbs.dtu.dk/services/TMHMM-2.0/, accessed on 27 January 2022).
2.4. Expression Analysis of StPP5 Gene
2.4.1. RT-PCR Analysis of StPP5 Expression
To study the expression of StPP5 gene in S. torvum under the stress of V. dahliae infection, S. torvum plants were treated with V. dahliae under greenhouse conditions. For V. dahliae infection test, we collected spore suspension and a fungus slice and then mixed them. After mixing, we filtered out the debris and infected the plant material with the filtrate. The stems and leaves of plants treated were harvested at 0 h, 12 h, 24 h, 48 h, and 72 h after treatment. The materials were frozen immediately using liquid nitrogen and stored at −70 °C.
Total RNA was extracted, and cDNA was synthesized using the same method as described above. RT-PCR was performed using specific primers: PP5-F (forward primer, 5′-TCAAATAGTACTGCAAACAAGA-3′) and PP5-R (reverse primer, 5′-ATG CACTTGAAGGCAAATAATG-3′). The GAPDH (glyceraldehyde-3-phosphate dehydrogenase) gene was used as an internal control with specific primers (forward primer, CAAGGACTGGAGAGGTGG; reverse primer, TTCACTCGTT GTCGTACC). The reaction system consisted of a 5-min denaturation at 94 °C, followed by 26 cycles of 40 s at 94 °C, 40 s at 53 °C, and 40 s at 72 °C.
2.4.2. RT-qPCR Analysis of StPP5 Expression
The program included 94 °C for 1 min, 40 cycles at 95 °C for 15 s, and 54 °C for 20 s according to the reaction condition: 10 µL 2 × SYBR premix Ex TaqTM (TaKaRa Biotech), 0.5 µL forward primer (10 μM), 0.5 µL reverse primer (10 μM), 2 µL cDNA, and 7 µL nuclease-free water.
2.5. Construction of Overexpression Vector
The target gene cDNA fragment was added with NcoI and SpeI restriction sites at its ends. Then, it was connected with vector pCAMBIA1304, double-digested, and the connected product was transformed into E. coli DH5α.
2.6. Genetic Transformation and Identification of Transgenic Plants
The Agrobacterium-mediated method was used for tobacco (N. tabacum ‘NC89’) transformation. Fully expanded leaves from sterile plants were cut into 0.5 cm2 segments in Petri dishes. A. tumefaciens strain LBA4404 carrying recombinant plasmid was grown at 28 °C to OD600 = 0.5~0.7 in LB medium with kanamycin (50 mg/L). The leaf segments were immersed in bacterial suspension of LBA4404 for 10 min. The leaf segments were blotted dry on aseptic filter paper and transferred to a solidified Murashige and Skoog medium (MS) for 2 days of cocultivation (MS + 1.0 mg/L 6-benzylaminopurine (6-BA) + 0.1 mg/Lα-naphthalene acetic acid (NAA)). After that, the leaf segments were transferred to the medium (MS + 2 mg/L 6-BA + 0.5 mg/L NAA + 250 mg/L Cefotaxime (Cef) + 20 mg/L Hygromycin B (Hyg B)). When shoots appeared in callus parts, the explants were placed on shoot differentiation medium (MS + 3 mg/L 6-BA + 0.2 mg/L NAA + 250 mg/L Cef + 20 mg/L Hyg B). Approximately 1 month later, shoots (1 cm long) were chosen and rooted on MS medium supplemented with 2 mg/L NAA, 250 mg/L Cef, and 25 mg/L Hyg B. Surviving plantlets were propagated on MS medium.
After the screening, the resistant ones were then PCR-identified. The PCR positive transformants were grown in tissue culture room for rooting at 25 ± 1 °C with 16 h light/8 h dark photoperiods.
2.7. Antifungal Assay
One gram of tobacco leaf was ground in phosphate buffer solution (PBS). After centrifugation at 12,000× g for 15 min, the supernatant liquid-containing proteins were collected as crude protein solution. Then, the concentrations of the crude protein were measured by the Coomassie brilliant blue method, and 36 μg crude protein of each sample was used for the following antifungal assay in vitro.
Cake with the fungus method [5,20] was applied for analyzing the inhibitory activity of transgenic tobacco protein extracts to V. dahliae in the in vitro antifungal assay. During the experiment, V. dahliae was cultured at 25 °C for 7~10 days. The experiment consisted of three treatments, blank control (water), wild-type tobacco, and transgenic plants, with three biological repetitions, respectively. Inhibition zone diameter was measured, and inhibitory rate was calculated using the following formula: Inhibitory rate = (control colony diameter (mm) − treatment colony diameter (mm))/control colony diameter (mm) × 100%. Difference in the significance between treatments was determined by t-test.
3. Results
3.1. Isolation of Full-Length cDNA of StPP5 Gene
A full-length cDNA of StPP5 was isolated from S. torvum by PCR with a pair of specific primers designed according to the assembled sequence of three fragment sequences of the gene. The cDNA fragment was about 1500 bp long (Figure 1).
DNAssist analysis showed that the StPP5 gene contained an open reading frame of 1458 bp long and encoded a protein of 485 amino acids (Figure 2). The cDNA sequence was submitted to the NCBI; the accession number is JF421126. Alignment analysis showed that StPP5 shared with Solanum lycopersicum, Zea. mays, Arabidopsis thaliana, and Oryza sativa Japonica Group had a higher homology of 84%, 82%, 81%, and 80%, respectively (Figure 2).
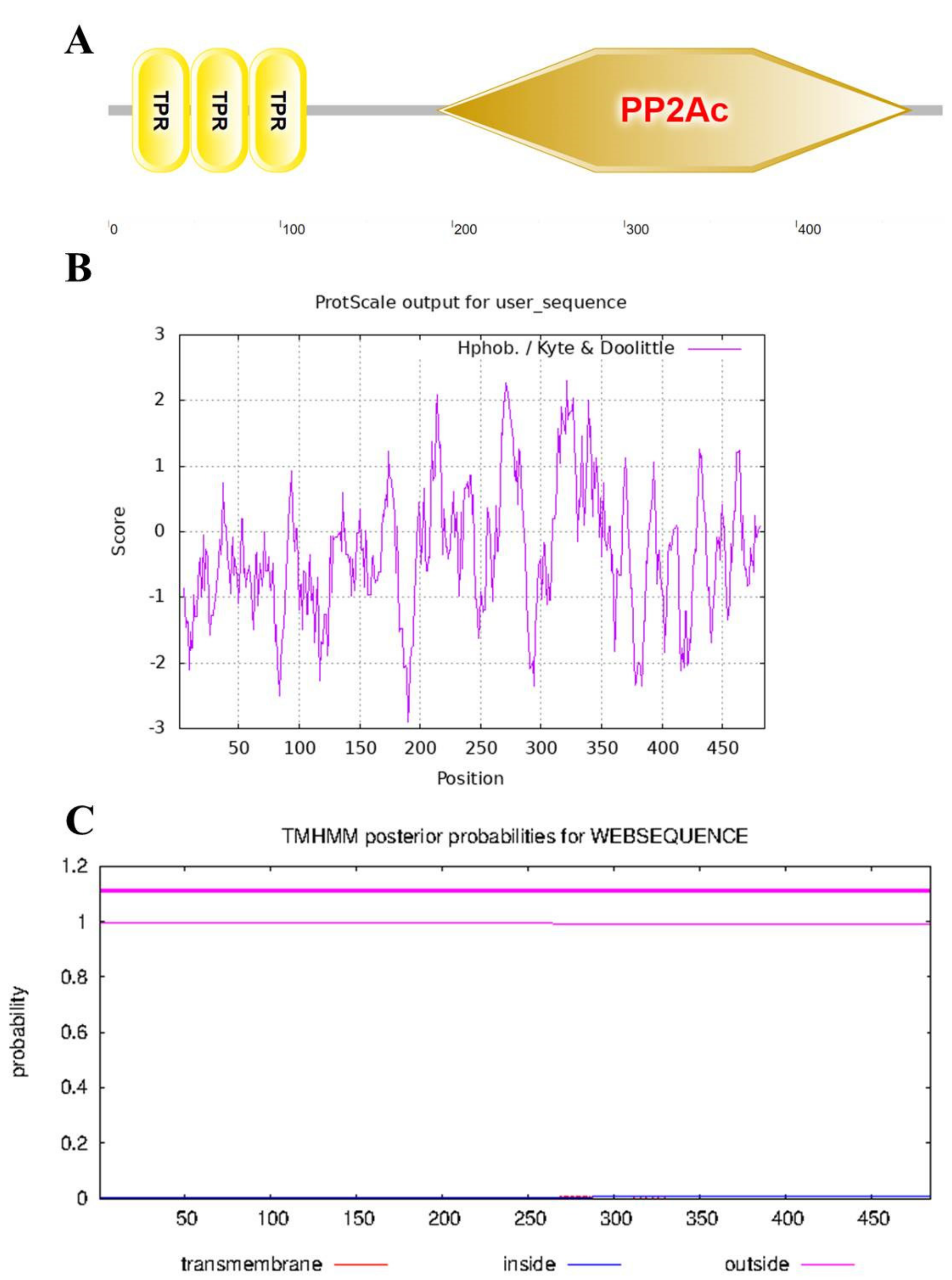
The StPP5 protein had a TPR domain in the N terminal and a PP-type specific domain in the C terminal (Figure 3A). Furthermore, the hydrophobicity of the StPP5 protein was predicted using the ProtScale program. The result showed that the maximum value of hydrophobicity was 2.300, and the minimum value was −2.900. Hydrophobic amino acids were distributed in the entire polypeptide chain, and the result suggests that StPP5 is a hydrophobic protein (Figure 3B). In addition, the protein transmembrane domain was predicted with the TMHMM Server v.2.0., and the result indicated that StPP5 was not a transmembrane protein (Figure 3C).
3.2. Phylogenetic Analysis of StPP5
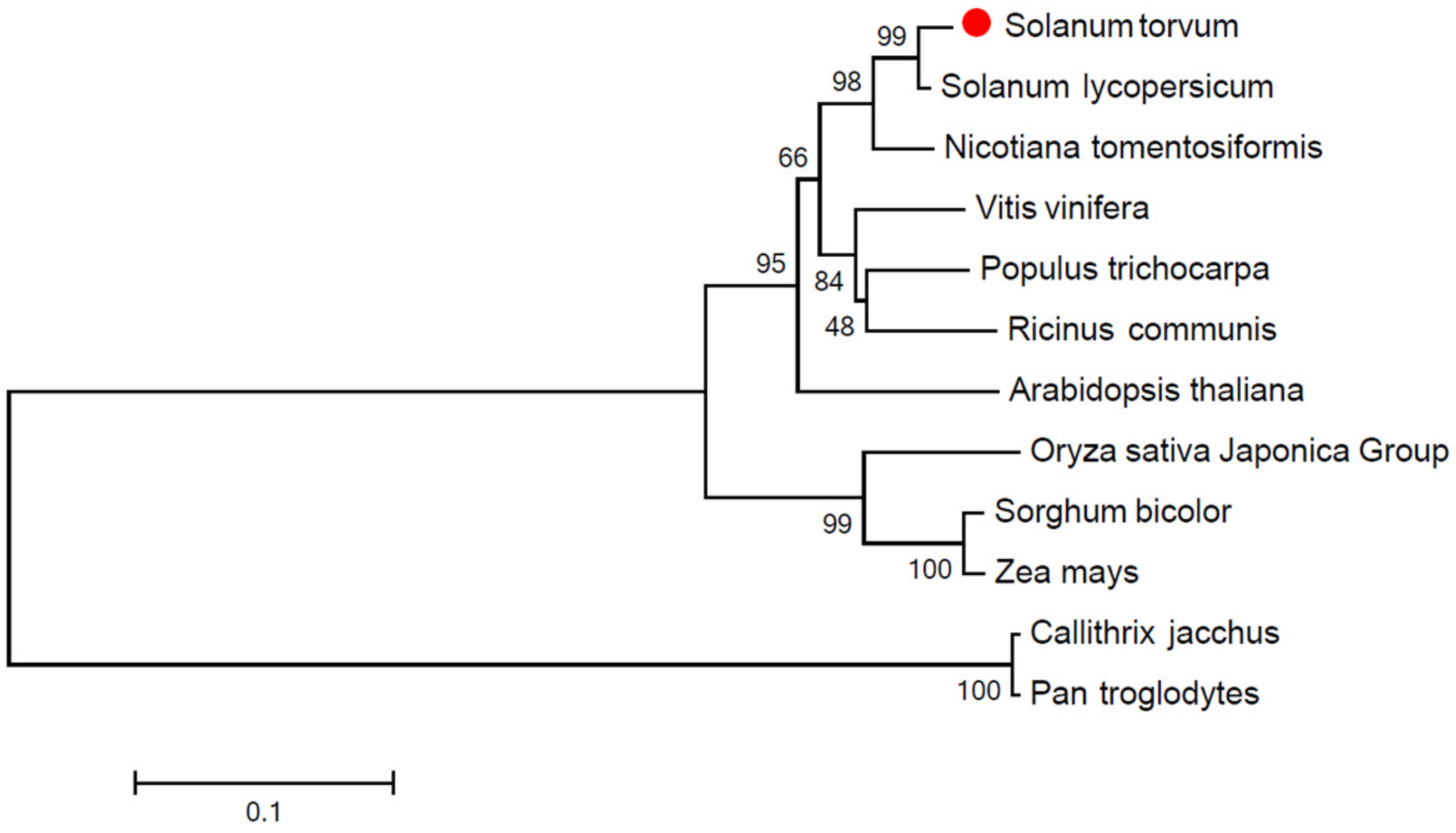
To determine the phylogenetic relationship of StPP5 with those from other species, a phylogenetic tree was constructed using the sequences of PP5 proteins from different species downloaded from GenBank. The phylogenetic tree analysis revealed that StPP5 was clustered to SlPP5 (Figure 4).
3.3. Expression of StPP5 Gene under V. dahliae Infection
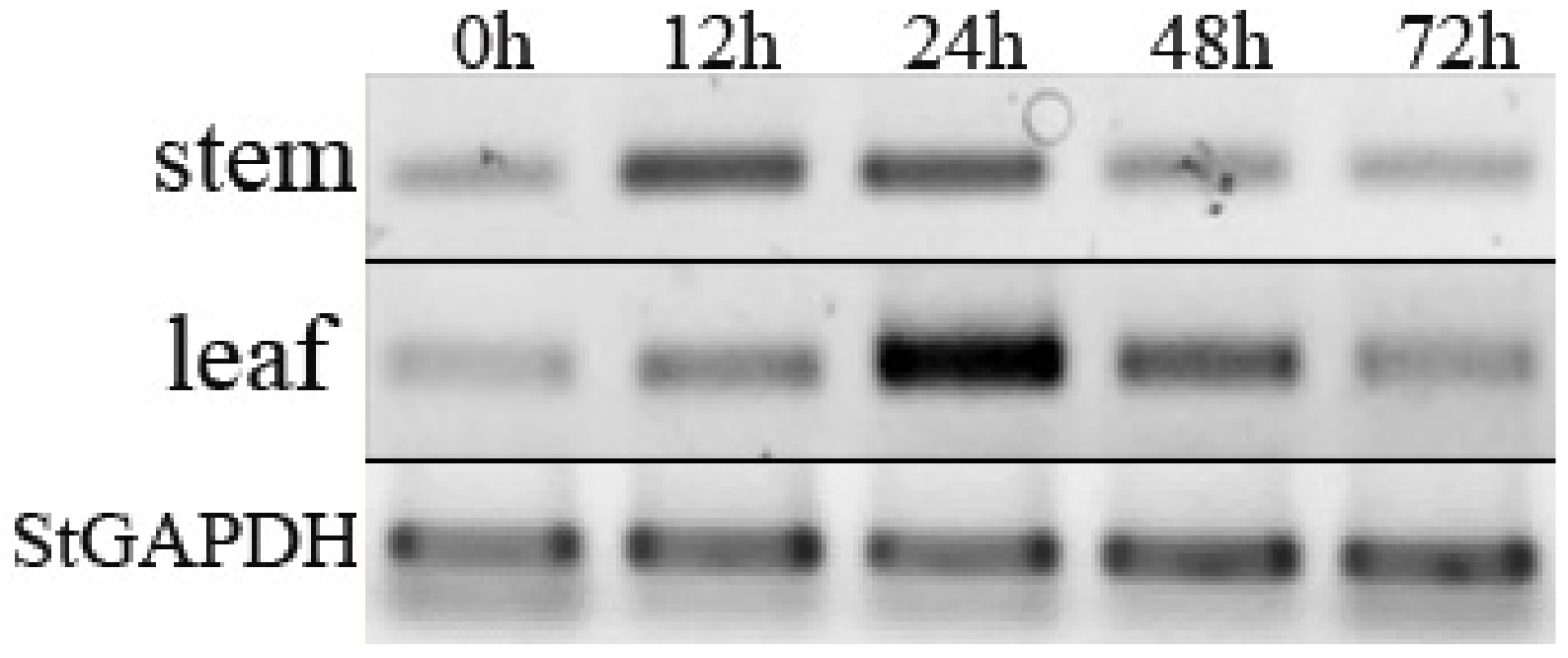
The expression of StPP5 under V. dahliae infection is shown in Figure 5. The expression patterns were different between stem and leaf. In stem, the maximal value of transcript appeared 12 h after V. dahliae inoculation and was maintained up to 24 h. In leaf, the maximal value of transcript was observed 24 h after inoculation; even the expression was raised from 12 h after infection.
3.4. Genetic Transformation and Plant Regeneration
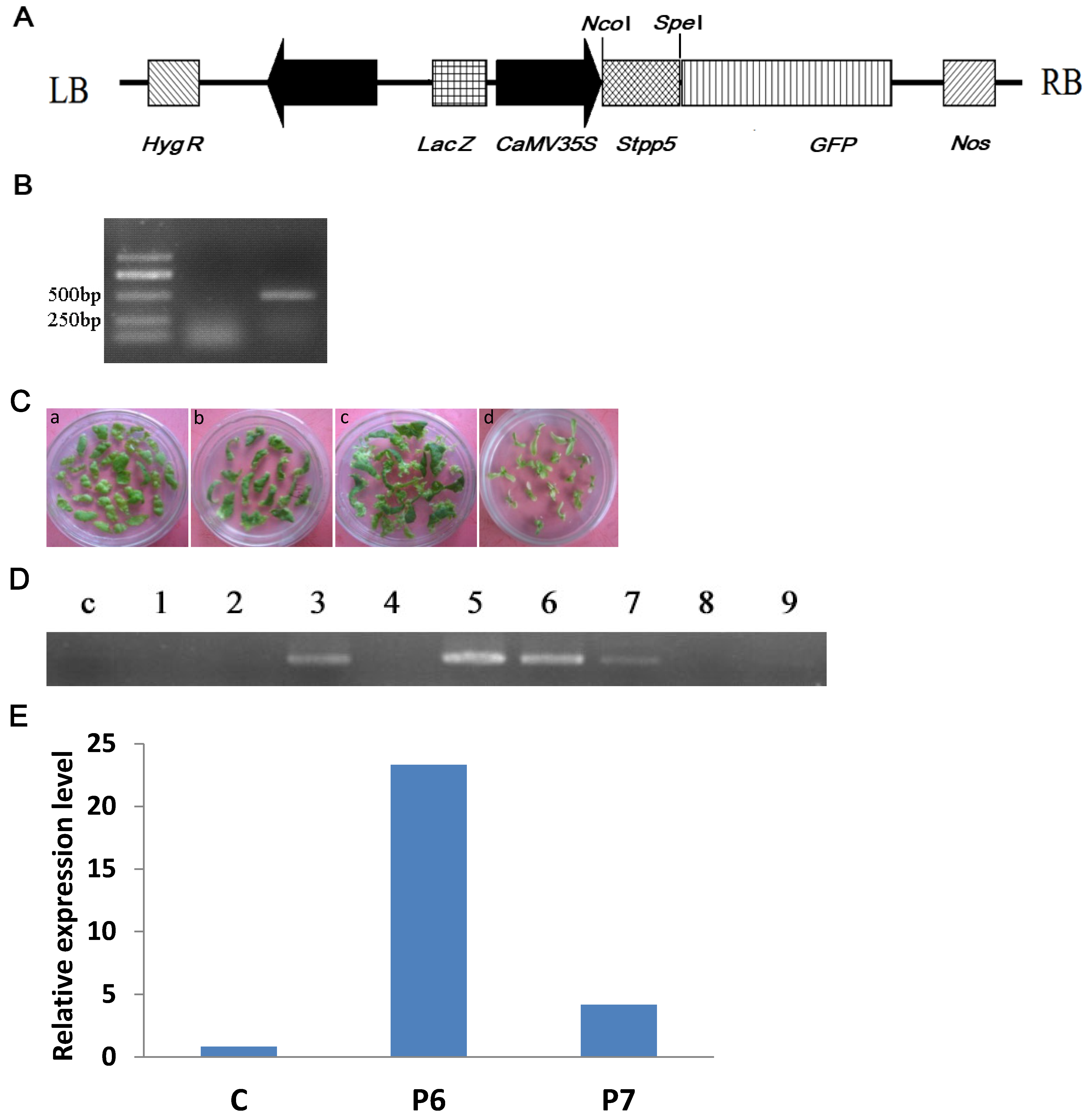
For transformation, pCAMBIA1304-StPP5 plasmid (Figure 6A) was transferred to competent Agrobacterium GV3101. PCR amplification produced a sequence of about 500 bp, which was consistent with the expected size (Figure 6B).
The transformation of tobacco was performed by the Agrobacterium-mediated method. Leaf fragment explants were cultured on the medium for callus induction, and the callus appeared 10 days later, after which it was transferred to the shoot induction medium. Shoots emerged from about 20 days of culture on the shoot induction medium (Figure 6C). Next, the shoots were excised and rooted on the rooting medium containing antibiotics for selection (Figure 6C). Surviving plantlets were propagated on the MS medium. In total, nine Hyg B positive plants were obtained and four transgenic lines were identified by RT-PCR, which are P3, P5, P6, and P7 (Figure 6D).
In order to confirm the expression of the StPP5 gene in transgenic lines, the lines P6 and P7 were selected for RT-qPCR analysis. The results showed that the level of StPP5 mRNA in lines P6 and P7 was much higher than the control, and P6 was 23 times of the control (Figure 6E).
3.5. Inhibitory Effect of Transgenic Tobacco Protein Extracts on V. dahliae
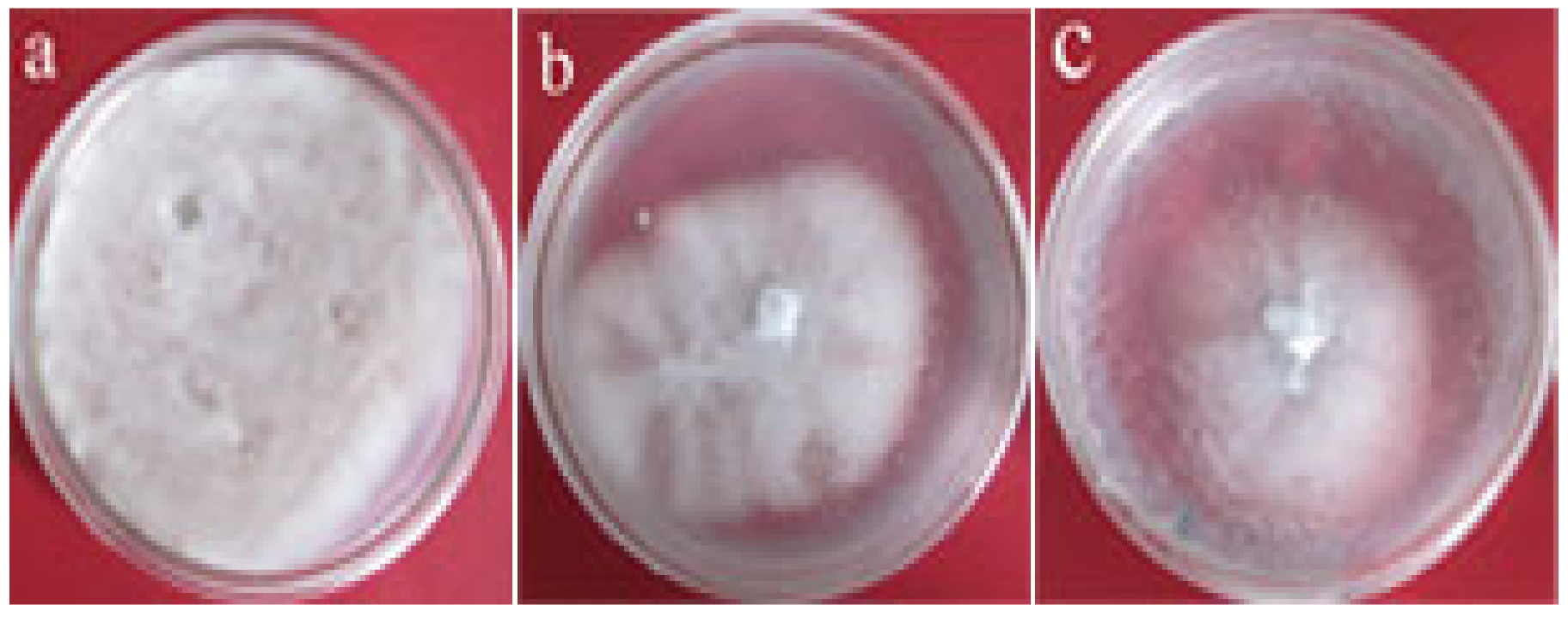
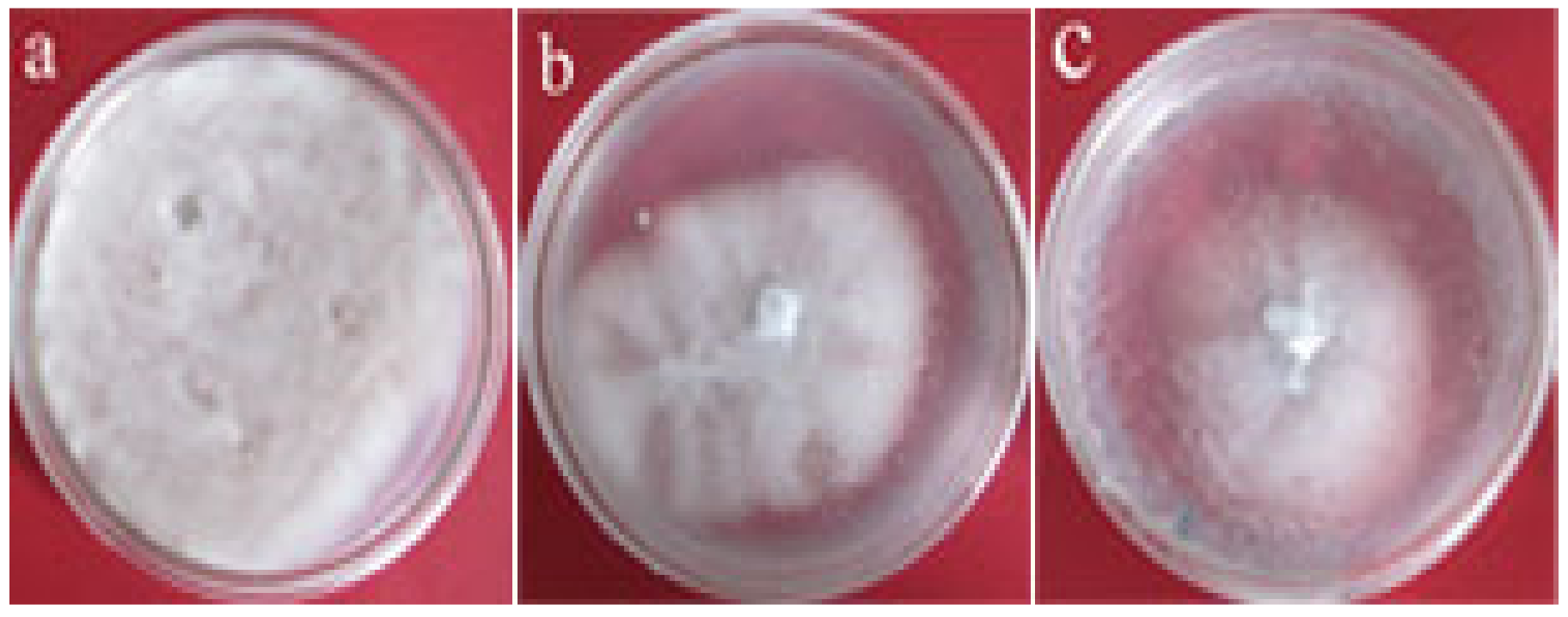
To reveal the function of the StPP5 gene, transgenic tobacco protein crude extract was added to the PDA medium for V. dahliae culture. The result showed that the growth of V. dahliae was significantly inhibited in the medium containing proteins extracted from the leaves of transgenic line P6 compared with wild-type tobacco. The average inhibition rate of line P6 was 55.2%, and wild-type tobacco was 31.1% (Figure 7, Table 1). These results showed that StPP5 had an inhibitory effect on V. dahliae proliferation.
4. Discussion
VW is widespread throughout the world and an important factor in most agricultural economies [21,22]. The plant defense response to VW is a complex process. Based on similarities in the genetic pathway, some VW resistance-related genes, including Ve1, Ve2, and PGIP (polygalacturonase-inhibiting protein), have been tested and found to be expressed after V. dahliae infection [7,23]. In the previous study in our laboratory, 118 TDFs were identified from eggplant S. torvum before and after inoculation of V. dahliae using cDNA-AFLP technology, in which a TDF was closer to serine/threonine-protein phosphatase and named StPP5 [18]. Protein phosphatases that cleave phosphate from phosphorylated serine and threonine in proteins are crucial for the regulation of most cellular processes [24,25]. We isolated an StPP5 gene from S. torvum, which belongs to a kind of PPPs family. The encoded amino acid sequence contains the typical domain structures common to most plant PPPs. The blastp comparison showed that StPP5 is highly homologous to other Solanaceaes, including a consistency with tomato of up to 96%. Smart online tools with protein molecules of StPP5 domain analysis revealed that its N terminal domain belonged to the TPR protein family, which is likely the domain interacting with other proteins, and the C terminal has a PP-type protein [26,27].
Serine/threonine-protein phosphatases are involved in cell metabolism, DNA replication, gene expression, signal transduction, cell cycle, cell differentiation, and apoptosis [28,29]. In this study, StPP5 was induced by V. dahliae infection. In vitro inhibitory assays showed that the inhibition rates of transgenic tobacco protein extracts to V. dahliae reached 55.2%, which was higher than that of wild-type tobacco. Quantitative RT-PCR analysis result indicated that StPP5 among transgenic tobacco lines was more highly expressed than wild-type tobacco and P6 was 23 times the wild-type control, which was consistent with the result of the inhibitory assay, suggesting that StPP5 might regulate the expression of downstream resistance genes through dephosphorylation, thus regulating the resistance of plants to V. dahliae. Similar results were reflected in StoVe1: the antifungal assay revealed that transgenic tobacco lines had higher inhibition rates of 35.5%, 29.4%, and 27.3% than the control plants, and the inhibition rates of transgenic potato protein extracts to V. dahliae ranged from 39.4 to 45.5% [2,30]. Previous studies discovered that the PP2C-type protein phosphatases gene operated at the junction of drought, heat shock, and oxidative stress [31], and Xb15 encoding a PP2C negatively regulated the XA21-mediated innate immune response in rice [32], consistent with our experimental results about the change of protein phosphatases in stress.
5. Conclusions
In summary, we cloned an StPP5 gene from S. torvum and revealed an inhibitory effect of StPP5 transgenic tobacco protein extracts to V. dahliae, thereby demonstrating the importance of StPP5 in response to VW and possibly providing a potential gene for the breeding of Solanaceae resistant to VW.
Author Contributions
Conceptualization, M.C. and C.X.; methodology, C.X.; validation, C.X.; formal analysis, M.C.; investigation, M.C. and C.X.; resources, M.C.; data curation, M.W.; writing—original draft, M.C. and C.X.; writing—review and editing, W.Z. and Q.Y.; visualization, M.C., W.Z. and Q.Y.; supervision, Q.Y.; project administration, Q.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was found by the National Natural Science Foundation of China (31901582), the Jiangsu Agricultural Science and Technology Innovation Found (cx (11)1020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original data sets described in the study are included in the article. Further inquiries can be addressed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weese, T.L.; Bohs, L. Eggplant origins: Out of Africa, into the Orient. Taxon 2010, 59, 49–56. [Google Scholar] [CrossRef]
- Liu, S.P.; Zhu, Y.P.; Xie, C.; Jue, D.W.; Hong, Y.B.; Chen, M.; Hubdar, A.K.; Yang, Q. Transgenic potato plants expressing Sto Ve1 exhibit enhanced resistance to Verticillium dahliae. Plant Mol. Biol. Rep. 2012, 30, 1032–1039. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. alboatrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.A. Mechanisms of disease resistance in Gossypium species and variation in Verticillium dahliae. Proc. World Cotton Res. Conf. 1994, 1, 225–235. [Google Scholar]
- Liu, H.; Zhang, N.; Yin, C.H.; Wang, J.H.; Ma, Z.H.; Zhou, L.G. Comparison between radial growth assay and liquid suspension culture assay for screening of antifungal natural compounds. Nat. Prod. Res. Dev. 2009, 21, 577–580. [Google Scholar]
- Parkhi, V.; Kumar, V.; Campbell, L.M.; Bell, A.A.; Shah, J.; Rathore, K.S. Resistance against various fungal pathogens and reniform nematode in transgenic cotton plants expressing Arabidopsis NPR1. Transgenic Res. 2010, 19, 959–975. [Google Scholar] [CrossRef] [PubMed]
- Kawchuk, L.M.; Hachey, J.; Lynch, D.R.; Kulcsar, F.; van Rooijen, G.; Waterer, D.R.; Robertson, A.; Kokko, E.; Byers, R.; Howard, R.J.; et al. Tomato Ve disease resistance genes encode cell surface-like receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 6511–6515. [Google Scholar] [CrossRef] [Green Version]
- Fradin, E.F.; Zhang, Z.; Juarez Ayala, J.C.; Castroverde, C.D.; Nazar, R.N.; Robb, J.; Liu, C.M.; Thomma, B.P. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiol. 2009, 150, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Fradin, E.F.; Abd-El-Haliem, A.; Masini, L.; van den Berg, G.C.; Joosten, M.H.; Thomma, B.P. Interfamily transfer of tomato Ve1 mediates Verticillium resistance in Arabidopsis. Plant Physiol. 2011, 156, 2255–2265. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.J.; Yin, Y.; Wang, Z.; Chen, M.; Huang, Q.S.; Huang, L.P.; Yang, Q. Cloning and analysis of full-length cDNA of Sto Ve1 gene from Solanum torvum. Plant Physiol. Commun. 2006, 42, 638–642. [Google Scholar]
- Miao, W.; Wang, X.; Li, M.; Song, C.; Wang, Y.; Hu, D.; Wang, J.S. Genetic transformation of cotton with a harpin-encoding gene hpaXoo confers an enhanced defense response against different pathogens through a priming mechanism. BMC Plant Biol. 2010, 10, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, X.; Yang, S.; Chi, J.; Zhang, G.; Ma, Z. Cloning and characterization of a Verticillium wilt resistance gene from Gossypium barbadense and functional analysis in Arabidopsis thaliana. Plant Cell Rep. 2011, 30, 2085–2096. [Google Scholar] [CrossRef]
- Stone, J.M.; Collinge, M.A.; Smith, R.D.; Horn, M.A.; Walker, J.C. Interaction of a protein phosphatase with an Arabidopsis serine-threonine receptor kinase. Science 1994, 266, 793–795. [Google Scholar] [CrossRef]
- Meskiene, I.; Bögre, L.; Glaser, W.; Balog, J.; Brandstötter, M.; Zwerger, K.; Ammerer, G.; Hirt, H. MP2C, a plant protein phosphatase 2C, functions as a negative regulator of mitogen-activated protein kinase pathways in yeast and plants. Proc. Natl. Acad. Sci. USA 1998, 95, 1938–1943. [Google Scholar] [CrossRef] [Green Version]
- Kerk, D.; Bulgrien, J.; Smith, D.W.; Barsam, B.; Veretnik, S.; Gribskov, M. The complement of protein phosphatase catalytic subunits encoded in the genome of Arabidopsis. Plant Physiol. 2002, 129, 908–925. [Google Scholar] [CrossRef] [Green Version]
- Mumby, M.C.; Walter, G. Protein-serine threonine phosphatases-structure, regulation, and functions in cell-growth. Physiol. Rev. 1993, 73, 673–699. [Google Scholar] [CrossRef]
- Gousset, C.; Collonnier, C.; Mulya, K.; Mariska, I.; Rotino, G.L.; Besse, P.; Servaes, A.; Sihachakr, D. Solanum torvum, as a useful source of resistance against bacterial and fungal diseases for improvement of eggplant (S. melongena L.). Plant Sci. 2005, 168, 319–327. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, J.L.; Zhang, F.; Huang, Q.S.; Huang, L.P.; Yang, Q. Differential expression analysis by cDNA-AFLP of Solanum torvum upon Verticillium dahliae infection. Russ. J. Plant Physiol. 2010, 57, 676–684. [Google Scholar] [CrossRef]
- Meng, X.; Li, F.; Liu, C.; Zhang, C.; Wu, Z.; Chen, Y. Isolation and characterization of an ERF Transcription factor gene from cotton (Gossypium barbadense L.). Plant Mol. Biol. Rep. 2010, 28, 176–183. [Google Scholar] [CrossRef]
- Quiroga, E.N.; Sampietro, A.R.; Vattuone, M.A. Screening antifungal activities of selected medicinal plants. J. Ethnopharmacol. 2001, 74, 89–96. [Google Scholar] [CrossRef]
- Wu, B.M.; Subbarao, K.V. A model for multiseasonal spread of verticillium wilt of lettuce. Phytopathology 2014, 104, 908–917. [Google Scholar] [CrossRef]
- Short, D.P.G.; Gurung, S.; Koike, S.T.; Klosterman, S.J.; Subbarao, K.V. Frequency of Verticillium species in commercial spinach fields and transmission of V. dahliae from spinach to subsequent Lettuce crops. Phytopathology 2015, 105, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Aguero, C.B.; Uratsu, S.L.; Greve, C.; Powell, A.L.; Labavitch, J.M.; Meredith, C.P.; Dandekar, A.M. Evaluation of tolerance to pierce′s disease and botrytis in transgenic plants of Vitis vinifera L. expressing the pear PGIP gene. Mol. Plant Pathol. 2005, 6, 43–51. [Google Scholar] [CrossRef]
- Cohen, P.T.W. Overview of protein serine/threonine phosphatases. In Protein Phosphatases; Springer: Berlin/Heidelberg, Germany, 2004; pp. 1–20. [Google Scholar]
- Mochida, S.; Maslen, S.L.; Skehel, M.; Hunt, T. Greatwall phosphorylates an inhibitor of protein phosphatase 2A that is essential for mitosis. Science 2010, 330, 1670–1673. [Google Scholar] [CrossRef]
- Das, I.; Dennis, J.E. Normal-boundary Intersection: A new method for generating the pareto surface in nonlinear multicriteria optimization problems. SIAM J. Optim. 1998, 8, 631–657. [Google Scholar] [CrossRef] [Green Version]
- Li, F.H.; Fu, F.L.; Sha, L.N.; Liang, H.; Li, W.C. Differential expression of serine/threonine protein phosphatase type-2c under drought stress in Maize. Plant Mol. Biol. Rep. 2009, 27, 29–37. [Google Scholar] [CrossRef]
- Sontag, E. Protein phosphatase 2A: The Trojan Horse of cellular signaling. Cell Signal. 2001, 13, 7–16. [Google Scholar] [CrossRef]
- Janssens, V.; Goris, J. Protein phosphatase 2A: A highly regulated family of serine/threonine phosphatases implicated in cell growth and signalling. Biochem. J. 2001, 353, 417–439. [Google Scholar] [CrossRef]
- Liu, S.P.; Hong, Y.B.; Wu, Z.; Ma, Y.S.; Jue, D.W.; Xie, C.; Zhu, Y.P.; Chen, M.; Yang, Q. Protein extract of tobacco expressing Sto Ve1 gene inhibits Verticillium dahliae proliferation. Czech J. Genet. Plant Breed. 2013, 49, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Vranová, E.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Camp, W.V. Oxidative stress, heat shock and drought differentially affect expression of a tobacco protein phosphatase 2C. J. Exp. Bot. 2000, 51, 1763–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.J.; Peng, Y.; Chen, X.; Dardick, C.; Ronald, P.C. Rice XB15, a Protein Phosphatase 2C, Negatively Regulates Cell Death and XA21-Mediated Innate Immunity. PLoS Biol. 2008, 6, 1910–1926. [Google Scholar]
Figure 1.
The schematic diagram (A) and agarose gel electrophoresis (B) of PCR amplification products of StPP5: P1,P2,P3,P4, the primers used for StPP5 cloning; M, DL 2000 marker; a, middle fragment; b, 3′ end fragment; c, 5′ end fragment; full-length, full-length cDNA of StPP5.
Figure 1.
The schematic diagram (A) and agarose gel electrophoresis (B) of PCR amplification products of StPP5: P1,P2,P3,P4, the primers used for StPP5 cloning; M, DL 2000 marker; a, middle fragment; b, 3′ end fragment; c, 5′ end fragment; full-length, full-length cDNA of StPP5.

Figure 2.
Alignment of the amino acid sequence of StPP5 with that of other homologous: LePP5 (AAN64317), AtPP5 (NP-565985), ZmPP5 (NP-001150042), and OsJGPP5 (NP-001054892).
Figure 2.
Alignment of the amino acid sequence of StPP5 with that of other homologous: LePP5 (AAN64317), AtPP5 (NP-565985), ZmPP5 (NP-001150042), and OsJGPP5 (NP-001054892).

Figure 3.
Structural analysis of StPP5: (A) structural domain; (B) hydrophobicity analysis of StPP5; (C) transmembrane domain prediction of StPP5.
Figure 3.
Structural analysis of StPP5: (A) structural domain; (B) hydrophobicity analysis of StPP5; (C) transmembrane domain prediction of StPP5.

Figure 4.
Phylogenetic analysis of StPP5. The Stpp5 were highlighted with the red circle. Source and accession number of PP5: Vitis vinifera (XP_002280111), Zea mays (NP_001150042), Arabidopsis thaliana (NP_001031534), Callithrix jacchus (XP_002762302), Pan troglodytes (XP_512768), Ricinus communis (XP_002520400), Populus trichocarpa (XP_024441018), Sorghum bicolor (XP_002439438), Oryza sativa Japonica Group (XP_015637335), Nicotiana tomentosiformis (XP_009603851), Solanum lycopersicum (AAN64317), and Solanum torvum (AEM45799).
Figure 4.
Phylogenetic analysis of StPP5. The Stpp5 were highlighted with the red circle. Source and accession number of PP5: Vitis vinifera (XP_002280111), Zea mays (NP_001150042), Arabidopsis thaliana (NP_001031534), Callithrix jacchus (XP_002762302), Pan troglodytes (XP_512768), Ricinus communis (XP_002520400), Populus trichocarpa (XP_024441018), Sorghum bicolor (XP_002439438), Oryza sativa Japonica Group (XP_015637335), Nicotiana tomentosiformis (XP_009603851), Solanum lycopersicum (AAN64317), and Solanum torvum (AEM45799).

Figure 5.
Expression analysis of StPP5 at different times after V. dahliae infection.
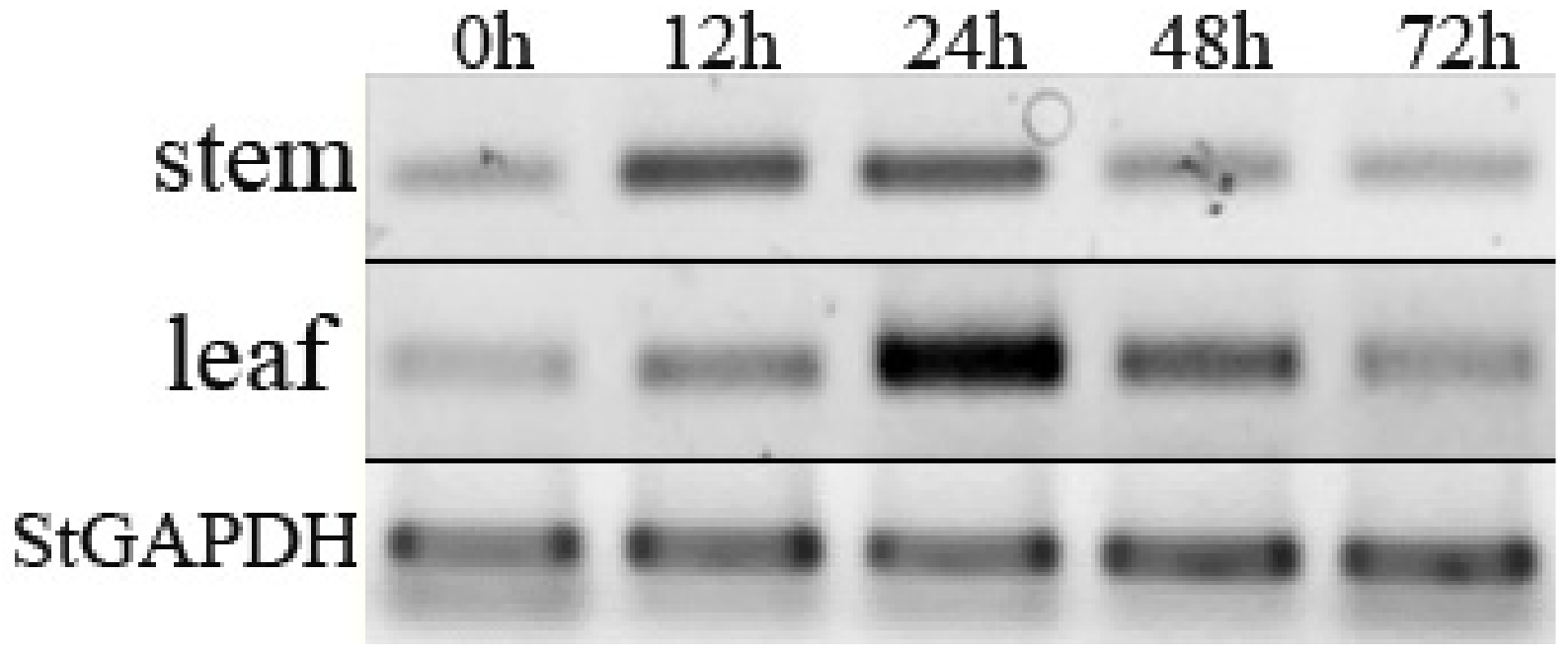
Figure 6.
Genetic transformation and regeneration of transgenic StPP5 tobacco plants: (A) Schematic representation of the binary vector pCMBIA1304-StPP5: LB, T-DNA left border; Hyg R, hygromycin resistance; lacZ, lacZ lapha; CaMV 35S, promoter; GFP, green fluorescent protein gene; nos, Nos(nopaline synthase) polyA signal, terminator; RB, T-DNA right border; (B) PCR identification of pCAMBIA1304-StPP5 recombinant plasmid in transformed A. tumefaciens: M, Marker; c, non-transformed control; cz, recombinant plasmid pCAMBIA1304-StPP5; (C) regeneration of transgenic tobacco plants: a, Explants; b, adventitious bud formation; c, the roots selection of 25 mg/L Hyg B; d, transgenic tobacco regeneration; (D) identification of StPP5 transgenic tobacco lines by PCR; (E) RT-qPCR analysis of StPP5 expression in transgenic tobacco lines: c, non-transgenic plant; P6 and P7, transgenic line.
Figure 6.
Genetic transformation and regeneration of transgenic StPP5 tobacco plants: (A) Schematic representation of the binary vector pCMBIA1304-StPP5: LB, T-DNA left border; Hyg R, hygromycin resistance; lacZ, lacZ lapha; CaMV 35S, promoter; GFP, green fluorescent protein gene; nos, Nos(nopaline synthase) polyA signal, terminator; RB, T-DNA right border; (B) PCR identification of pCAMBIA1304-StPP5 recombinant plasmid in transformed A. tumefaciens: M, Marker; c, non-transformed control; cz, recombinant plasmid pCAMBIA1304-StPP5; (C) regeneration of transgenic tobacco plants: a, Explants; b, adventitious bud formation; c, the roots selection of 25 mg/L Hyg B; d, transgenic tobacco regeneration; (D) identification of StPP5 transgenic tobacco lines by PCR; (E) RT-qPCR analysis of StPP5 expression in transgenic tobacco lines: c, non-transgenic plant; P6 and P7, transgenic line.

Figure 7.
Growth and propagation of V. dahliae in the media with different tobacco protein extracts: (a) water control; (b) non-transgenic line; (c) transgenic line P6.
Figure 7.
Growth and propagation of V. dahliae in the media with different tobacco protein extracts: (a) water control; (b) non-transgenic line; (c) transgenic line P6.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Inhibition rate of the protein extract of transgenic and non-transgenic tobacco to V. dahliae.
Table 1.
Inhibition rate of the protein extract of transgenic and non-transgenic tobacco to V. dahliae.
| Treatment | Average Inhibition Zone Diameter (mm) | Inhibition Rate (%) |
|---|---|---|
| Wild-type tobacco Transgenic line P6 | 62.0 40.3 | 31.1 55.2 * |
*: p < 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, M.; Xie, C.; Yang, Q.; Zhu, W.; Wang, M. Protein Extract of Tobacco Expressing Solanum torvum PP5-Encoding Gene Inhibits Verticillium dahliae Proliferation. Horticulturae 2022, 8, 252. https://doi.org/10.3390/horticulturae8030252
AMA Style
Chen M, Xie C, Yang Q, Zhu W, Wang M. Protein Extract of Tobacco Expressing Solanum torvum PP5-Encoding Gene Inhibits Verticillium dahliae Proliferation. Horticulturae. 2022; 8(3):252. https://doi.org/10.3390/horticulturae8030252
Chicago/Turabian StyleChen, Min, Chao Xie, Qing Yang, Wenjiao Zhu, and Man Wang. 2022. "Protein Extract of Tobacco Expressing Solanum torvum PP5-Encoding Gene Inhibits Verticillium dahliae Proliferation" Horticulturae 8, no. 3: 252. https://doi.org/10.3390/horticulturae8030252
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.