1. Introduction
Cadophora luteo-olivacea is a fungus mainly associated with decay symptoms in vines as a vascular pathogen and in kiwifruit (
Actinidia chinensis var.
deliciosa (A. Chev) cv. Hayward) as a rotting fruit agent [
1]. Khol et al. [
2] described
C. luteo-olivacea as a pathogen able to infect developing fruits during the growing season, remaining quiescent and then appearing during the post-harvest phase in cold storage. However, its incidence can vary over time and is strictly connected to field conditions [
3]. Even if
Botrytis cinerea represents the most critical post-harvest pathogen of kiwifruit [
4], skin pitting symptoms after different storage months (on average 4) in most Italian packaging houses were detected.
Kiwifruit cv. Hayward can be stored for 3–6 months at −1 °C in normal refrigeration (NR) or in a controlled atmosphere (CA) (2% O
2; 4.5% CO
2). Nevertheless, CA conditions seem to stimulate gray mold development and less stimulate biocontrol antagonists’ mechanisms of action, as reported by Di Francesco et al. [
5]. In this context, biocontrol agents (BCAs) and natural substances represented non-conventional methods to control post-harvest fungal diseases.
Among the various alternative approaches, biological control utilizing antagonistic microorganisms has been shown to effectively manage different post-harvest diseases of kiwifruit [
5,
6,
7]. Along with antagonistic microorganisms,
A. pullulans was often tested in the postharvest phase to control different fungal pathogens and also against the gray mold of kiwifruit [
5], showing good efficacy. Even in the case of
T. harzianum, the BCA is often recognized due to its effectiveness in controlling several pathogens in diverse fruits, including kiwifruit [
8]. The efficacy of both BCAs is always related to several mechanisms of action reported like competition, antibiosis, parasitism (involving lytic enzymes), and the induction of plant defenses.
However, in recent years, the interest in natural substances (essential oils, aromatic compounds, and hydrolytic reaction products) has increased, and numerous studies on their biocidal activity have been conducted. Among these, several compounds can be artificially added by fumigation in cold storage, such as glucosinolates, a large class of approximately 100 compounds produced by
Cruciferae [
9]. The antifungal activity of allylisothiocyanate (AITC), a naturally occurring flavor compound in mustard and horseradish, successfully demonstrated its efficiency on different fruit species [
10,
11,
12]. AITC is ‘generally recognized as safe’ (GRAS) by the Food and Drug Administration (FDA) in the United States since 2006, authorized as a natural food preservative in Japan, and evaluated as a safe additive by EFSA (European Food Safety Authority, 2010). Nevertheless, fruit fungicides dipping is still used for post-harvest applications [
13], especially fludioxonil, registered for post-harvest applications against fungal diseases. Generally, fludioxonil is considered a fungicide with low toxicity and a broad spectrum, but it remains unclear whether its residues in food have potential harm to humans.
In addition, it is well known that some inorganic salts as CaCl
2 (calcium chloride) can increase the storage quality of many fruits [
11], probably determining a higher resistance to fungal pathogens attack. In fact, calcium alters intracellular and extracellular processes, which retard ripening [
13] but at high concentration could cause severe pitting on fruits [
11].
To the best of our knowledge, not so many studies have been conducted to explore the efficacy of utilizing BCAs and natural substances to control
C. luteo-olivacea of kiwifruit [
14].
This study’s objective was to evaluate traditional (fungicides) and unconventional (BCAs, GRAS) treatments efficacy to control skin pitting symptoms of kiwifruit through in vitro and in vivo assays. Different strategies have been adopted: (a) testing the efficacy of Aureobasidium pullulans and Trichoderma harzianum strains by dual culture, VOCs, and N-VOCs assays; (b) evaluating the EC50 values of CaCl2 and fludioxonil (Scholar) against C. luteo-olivacea CFU and mycelial growth (c) and the effect of AITC on fungal isolates growth. (d) The same above-mentioned treatments were tested on kiwifruits to verify their effectiveness; (e) the effect of AITC biofumigation on kiwifruit was biochemically evaluated through FT-IR (Fourier-Transform Infrared) spectroscopy.
2. Materials and Methods
2.1. Fruit
Hayward kiwifruits (Actinidia chinensis var. deliciosa (A. Chev.), C.F. Liang & A.R. Ferguson) obtained from an orchard located in Faenza (Italy) were used. Fruits harvested at the time of collection (6.5° Brix) were picked from trees. Fruits were stored at 0 °C, and immediately before the experiment were wounded with a sterile nail (3 × 3 × 3 mm).
2.2. Biocontrol Agents and Pathogens
Aureobasidium pullulans strains L1 and L8 were molecularly characterized by Di Francesco et al. [
15] and maintained on nutrient yeast dextrose agar (NYDA: 8 g of nutrient broth, 5 g of yeast extract, 10 g of dextrose, and 15 g of agar in 1 L of distilled water) at 4 °C until use [
16]. Two days before trials, each antagonist was grown on NYDA at 25 °C, and the yeast cells were collected in sterile distilled water containing 0.05% (
v/v) Tween 80 and quantified for a final concentration of 10
8 cells mL
−1 by counting spore suspension on a hemocytometer cell.
Trichoderma harzianum (Th1) isolate derived from Criof fungal collection was maintained on potato dextrose agar (PDA, 39 g in 1 L of distilled water) (Oxoid, Basingstoke, UK) at 4 °C until use. Four days before trials, the fungal isolate was grown on new PDA at 20 °C, and conidia were collected in sterile distilled water containing 0.05% (
v/v) Tween 80 and quantified for a final concentration of 10
5 conidia mL
−1, as above reported.
Cadophora luteo-olivacea isolates (CadA, CadB, CadC) belonged to Criof fungal collection. Each isolate was grown on PDA and incubated at 20 °C. Conidial suspensions were prepared from 20-days-old colonies by scraping and suspending conidia in sterile distilled water with 0.05% (
v/v) Tween 80 and adjusted to the final concentration required (10
5 conidia mL
−1) with a hemocytometer.
2.3. Chemical and Natural Substances
Regarding the chemical fungicide, Scholar® (fludioxonil) (Syngenta, Basil, Switzerland), registered for post-harvest applications against diseases of fruits in different European countries, was tested at different concentrations (0.001, 0.025, 0.05, 0.10, 0.17, 0.25, 0.5, 1.0, and 2.0 g L−1) by in vitro (amended medium) and in vivo (dipping) assays. CaCl2 (Sigma Aldrich) was used at different concentrations (0.1, 0.2, 0.4, and 0.8 g L−1) for the in vitro (amended medium) and in vivo (dipping) experiments. Brassica nigra defatted meals were prepared by shaking ground seeds (Jungepflanzen, Forchheim, Germany) with hexane (1:10, w/v) overnight. After filtration through filter paper, the residual solvent was removed under vacuum. The ground seeds meal was mixed with sterile distilled water at a rate of 3:1 (w/v) (grams) before each treatment.
2.4. In Vitro Assays with Antagonists
In the co-culturing assay,
C. luteo-olivacea isolates CadA, CadB, CadC were separately co-cultured with L1 and L8 yeast strains, and Th1 on PDA (Oxoid, Basingstoke, United Kingdom) plates. In each plate, a mycelial plug (6 mm in diameter) of
C. luteo-olivacea isolates CadA, CadB, CadC derived from a 20-days-old colony was inoculated 30 mm from the plate edge, and
A. pullulans cells, from a 48-hour-old colony grown on NYDA and a mycelial plug (6 mm in diameter) of Th1 from 7-days-old colony grown on PDA, were respectively streaked by a sterile loop and positioned 30 mm from the fungal plug [
15]. In control plates, only
C. luteo-olivacea isolates CadA, CadB, CadC plugs were inoculated. Plates were incubated at 20 °C in the dark for 15 days; the fungal colony’s radius was measured on a line from the plug’s center to the yeast swipe.
For the N-VOCs assay, cells (100 μL; 108 cells mL−1) of each Aureobasidium strain and conidia suspensions (100 μL; 105 conidia mL−1) obtained respectively from 2-days-old colonies grown on NYDA and 5-days-old colonies grown on PDA were spread on sterile cellophane layer (Safta, Piacenza, Italy) previously positioned on PDA plates. In control plates, 100 μL of sterile distilled water (SDW) were used.
After 48 h of incubation at 25 °C and 20 °C, respectively, cellophane was removed, and a mycelial plug (6 mm diameter) of each C. luteo-olivacea isolate (CadA, CadB, CadC) was inoculated in the center of the plate. Plates were incubated at 20 °C, and the colony diameter was measured after 15 days from the inoculation. The VOCs produced by L1, L8, and Th1 strains were tested against the mycelial growth of C. luteo-olivacea isolates. For this purpose, NYDA and PDA plates were respectively inoculated spreading 100 μL of L1 and L8 cells suspension (108 cells mL−1) and 100 μL of Th1 conidia suspension (105 cells mL−1). The plate’s lid was replaced, after 48 h of incubation at 25 °C and 20 °C, by a base plate of PDA inoculated with a mycelial plug (6 mm diameter) of each pathogen. The two base plates were sealed immediately with a double layer of parafilm and incubated at 20 °C for 7 days. In all experiments, five plates (replicates) were used for each combination and the controls. The experiments were conducted twice.
2.5. In Vitro Assays: Amended Medium and Brassica nigra Defatted Meal Biofumigation
To determine the sensitivity of CFU and mycelial growth of C. luteo-olivacea isolates to fludioxonil and CaCl2, PDA was autoclaved, and the products incorporated after cooling (50 °C) at final concentrations of 0.001, 0.025, 0.05, 0.10, 0.17, 0.25, 0.5, 1.0, 2.0 g L−1 and 0.1, 0.2, 0.4, 0.8 g L−1, respectively. A mycelium plug (6 mm in diameter) and conidial suspensions (103 conidia mL−1; 100 µL), derived from 15-days-old actively growing colonies on PDA of each C. luteo-olivacea isolate, were transferred to each amended PDA dish and incubated at 20 °C for 15 days. Five replicates (plates) were used for each isolate at different product concentrations. No amended PDA plates were inoculated with each isolate and used as a control. The experiment was performed twice. EC50 values were calculated as the concentration (g L−1) that inhibits mycelial and CFU growth by 50% compared with the control.
For the biofumigation assay, Petri dishes (90 mm diameter) with two compartments were used. One compartment was filled with 10 mL of PDA, suddenly inoculated by C. luteo-olivacea isolates mycelial plugs (6 mm diameter), and in the other plate compartment, 3 g of B. nigra defatted meal soaked with 9 mL of SDW were inserted. The plates were sealed immediately with a double layer of parafilm and incubated at 20 °C for two weeks. Plates without B. nigra meal were considered as control. Five replicates (plates) were used for each isolate, and the experiment was performed twice.
2.6. In Vivo Assays
In vivo assays were conducted testing all the above-cited in vitro treatments: L1 and L8 yeast strains, Th1 isolate, CaCl2, fludioxonil, and B. nigra defatted meal. For the BCAs assay, kiwifruits were wounded by a sterile nail (3 × 3 × 3 mm) at the equator (one wound per fruit) and then inoculated with 20 μL of each isolate BCAs suspension (108 cells mL−1 and 105 conidia mL−1, respectively for L1, L8, and Th1). After one hour from BCAs inoculation, 20 μL of CadA, CadB, CadC conidia suspension (105 conidia mL−1) were inoculated.
Regarding the treatments with fludioxonil and CaCl2, kiwifruits were dipped in solutions concentrated 1 g L−1 and 0.8 g L−1, respectively, for each product.
The fruits were dried at room temperature and suddenly artificially inoculated with pathogen conidial suspension, as above described. About the kiwifruit biofumigation with B. nigra defatted meal, 10 g of meal soaked with 30 mL of SDW were placed inside a storage cabinet (0.1 m3) together with fruits for 24 h at 0 °C. Biofumigated kiwifruits were artificially inoculated with 20 μL of CadA, CadB, CadC conidia suspensions (105 conidia mL−1).
Kiwifruits were stored at 0 °C for five months and then at 20 °C for another seven days (shelf-life). The sample unit was represented by three replicates (baskets) of 25 fruits each for all the treatments. The experiments were performed twice. In all experiments, control fruits were represented by untreated fruits placed in the same conditions. Results were collected by recording the percentage of fruits with symptoms of rot in each treatment and the lesion’s diameter (mm) by using a ruler.
2.7. FT-IR Spectroscopy
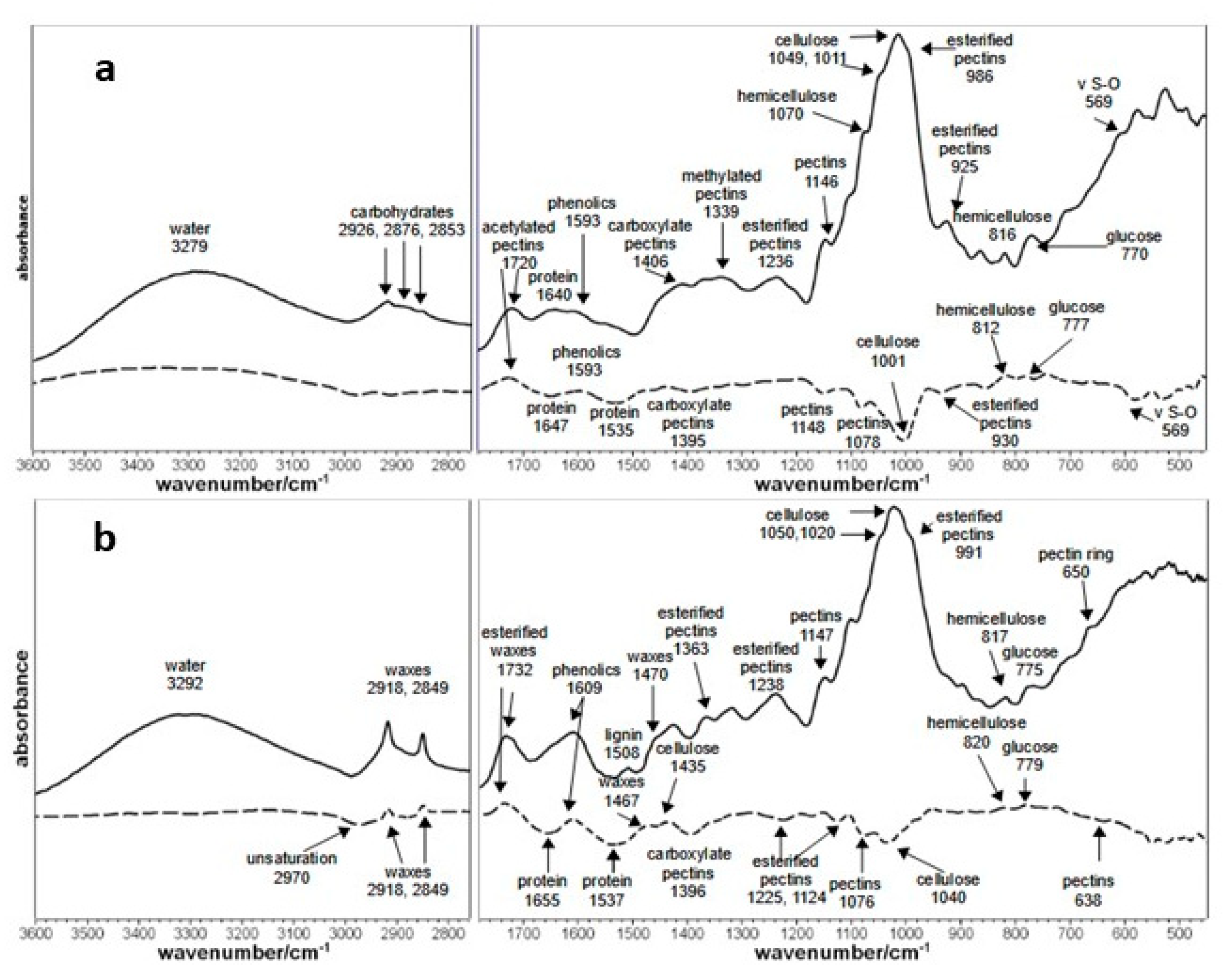
Kiwifruit samples (50 g) treated 24 h with B. nigra defatted meal volatiles and untreated were collected in sterile tubes (2 mL), stored at −80 °C and lyophilized. Fruit samples were analyzed by FT-IR spectroscopy to obtain a rapid and non-destructive characterization of their main molecular components. Infrared spectra were recorded with a Bruker ALPHA series FT-IR spectrophotometer (Bruker, Ettlingen, Germany) equipped with an apparatus for attenuated total reflectance (ATR-Diamond crystal). The spectra were collected from 4000 to 400 cm−1 and averaged over 100 scans (resolution = 4 cm−1): four spectra were measured for each sample. Spectra were registered both on fruit skin and fruit flesh to study their different biochemical compounds and their variations.
2.8. Statistical Analysis
Data were statistically handled by one-way analysis of variance (ANOVA). Statistical comparison of means was carried out to reveal the differences between treatments using Tukey’s HSD Test (α = 0.05). All analyses were performed with MiniTab 16 software. The EC50 of each substance was calculated using the probit analysis applied to mycelial growth inhibition percentage.
4. Discussion
Alternative methods for kiwifruit decay control have always been studied and tested with encouraging results [
5,
19] but always against the most common post-harvest pathogens such as
B. cinerea,
Penicillium expansum,
Alternaria alternata, Rhizopus stolonifer [
13]. A pathogen such as
C. luteo-olivacea represents a critical problem in these last years, even if its incidence in the post-harvest phase seems to be strictly connected to field conditions, making it less manageable. Spadaro et al. [
3] demonstrated that kiwifruit, which showed a low level of dry matter and high nitrogen content, could be more susceptible to
C. luteo-olivacea infection.
Little is known about the natural products and biological systems to control this pathogen. The study presented some interesting results about the possibility of preserving kiwifruit quality using alternative methods to fungicides against the skin pitting causal agent. Despite this, it is often rare to find alternatives that give a level of control comparable with that obtained using synthetic fungicides [
20]. Furthermore, our study showed interesting results where
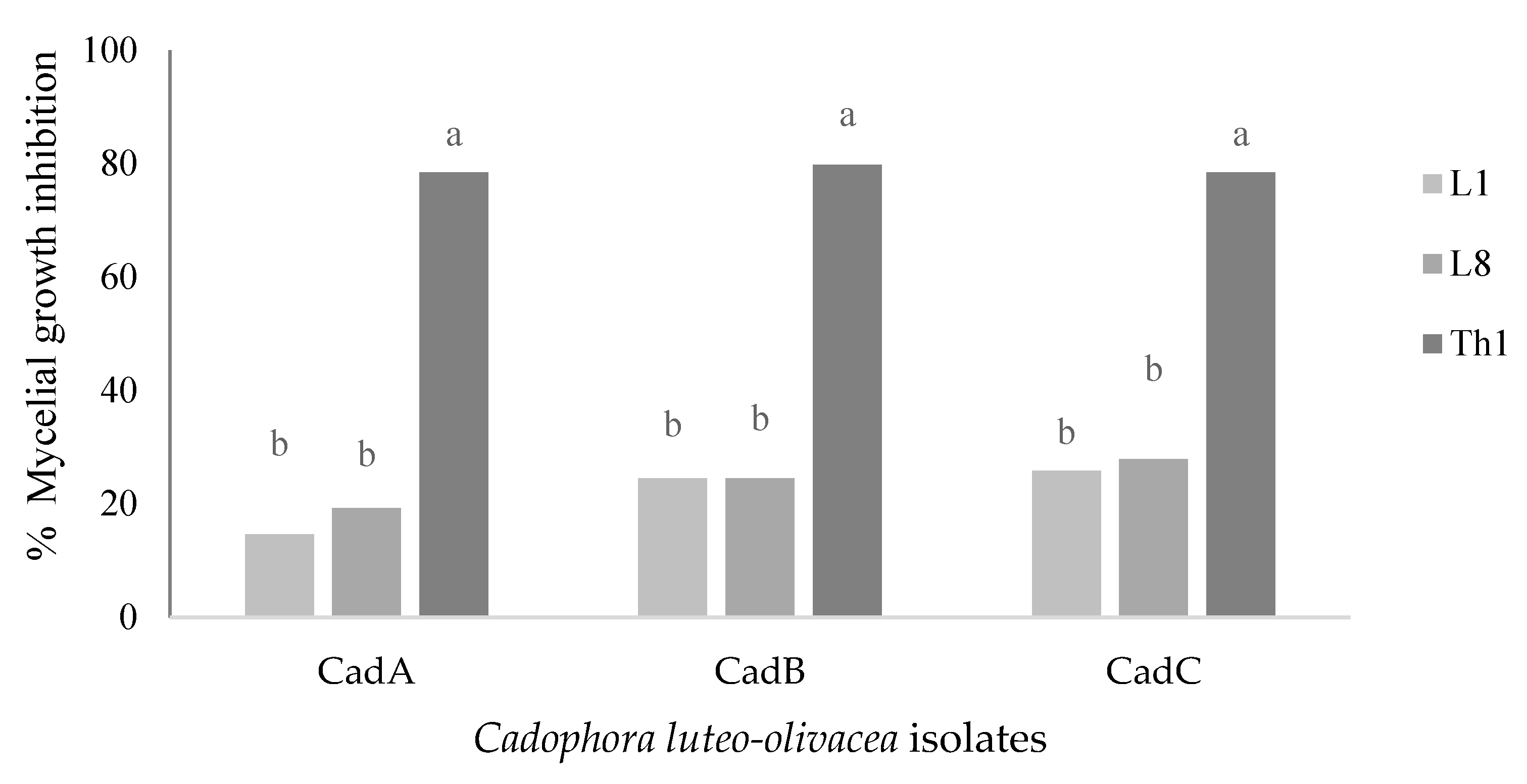
T. harzianum showed the highest inhibitory activity against the three tested
C. luteo-olivacea isolates (CadA, CadB, CadC) by volatile, non-volatile, and by dual culture assay, displaying high percentages of inhibition such as on average, respectively, by 90%, 70.6%, and 78.8%. The biocontrol activity exerted by
A. pullulans L1 and L8 strains was less efficient; consequently, the pathogen has been challenging to control on most of the carried-out treatments. By in vitro experiments, both strains reduced on average by VOCs, no-VOCs, and dual culture the pathogen isolates respectively by 23.3% and 25.8%, 50% and 34.7%, and 22.5 and 23.6%. It is known that the antifungal activity of microorganisms by in vitro and in vivo conditions can vary depending on pathogen aggressiveness and growth behavior [
5,
21]. In fact, L1 and L8 yeasts against
B. cinerea of kiwifruit showed an excellent efficacy by competing for nutrients [
5] through in vitro and in vivo experiments. Against
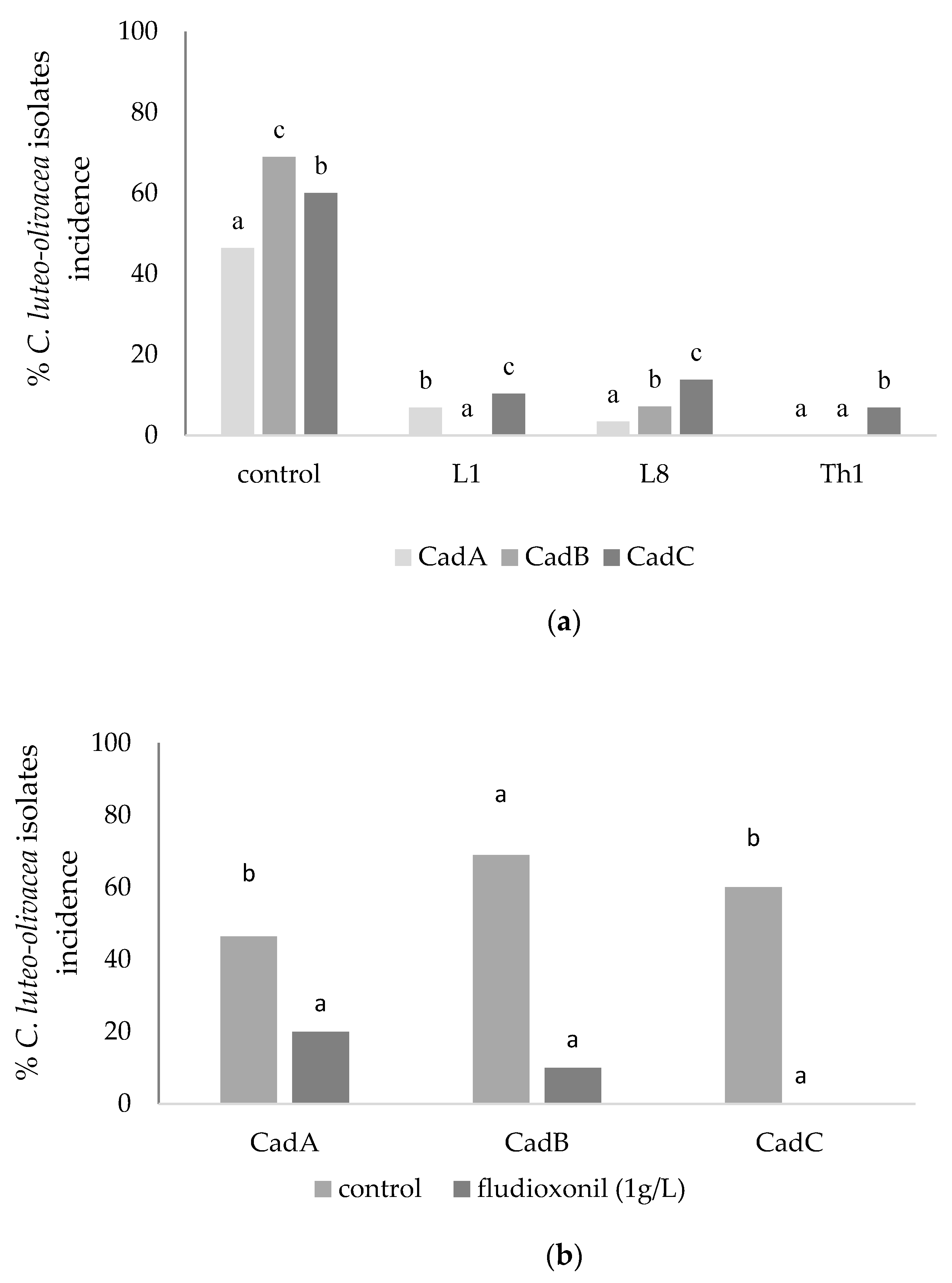
C. luteo-olivacea regarding the in vivo experiments, the tested BCAs showed better efficacy than in vitro to significantly reduce the skin pitting symptoms, especially for L1 and L8 strains that probably activated a resistance response on kiwifruits [
16,
18].
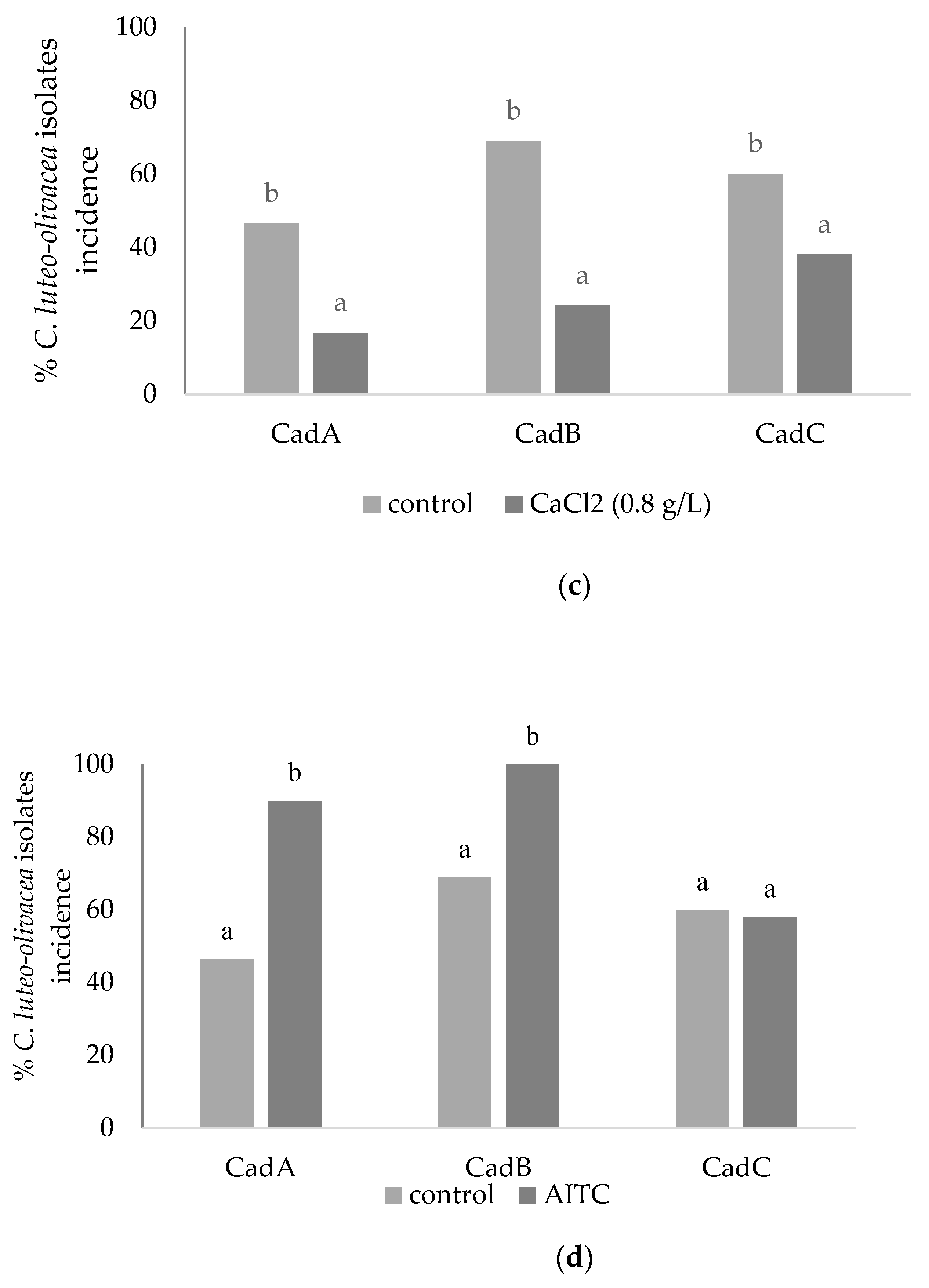
Between the alternative strategies tested to control
C. luteo-olivacea isolates, CaCl
2 activity was verified before by in vitro assay to define the most active concentration to use during the in vivo phase. CaCl
2 concentration of 0.8 g L
−1 resulted in the most efficient, although the EC50 values were averagely high (22.5 g L
−1) against mycelial growth. It has been reported that pre and post-harvest calcium application in fruit tissues delays softening rate and ripening by retarding the disintegration of cell walls [
22]. Antunes et al. [
21] reported that fruits dipped in 1% CaCl
2 softened slower than fruits not treated. In the present study, in vivo assay fruits were not affected by CaCl
2 treatment; on the contrary, they showed reasonable control of the skin pitting symptoms (on average 53% of fungal incidence inhibition), displaying the benefits of this treatment on the storage life of kiwifruit.
The study included as reference treatment the use of a fungicide with fludioxonil as an active compound. The product (Scholar) was used first in in vitro assay to define the most active concentration against
C. luteo-olivacea isolates mycelial growth and defining its EC50 value (on average, 0.36 and 0.92 g L
−1, respectively for CFU and mycelial growth). Our results showed that fludioxonil displayed a good efficacy starting from 1 g L
−1 of concentration, a reduced amount with respect to the kiwifruit reference dose (1.3 g L
−1) for
B. cinerea post-harvest inhibition, also probably reducing the residue levels. Nevertheless, the tested fungicide cannot be used in kiwifruits to be marketed within 5 months of its application unless fruits stayed for a more extended period in storage [
13]. However, the debate on the comparative safety of synthetic and natural pesticides is still open.
This study also showed that kiwifruit dipping in the most active fludioxonil solution (1 g L
−1) before the cold storage effectively reduced the post-harvest skin pitting symptoms after 4 months of storage, as reported for
B. cinerea by Kim et al. [
23] and Lolas-Caneo et al. [
24] also on different products.
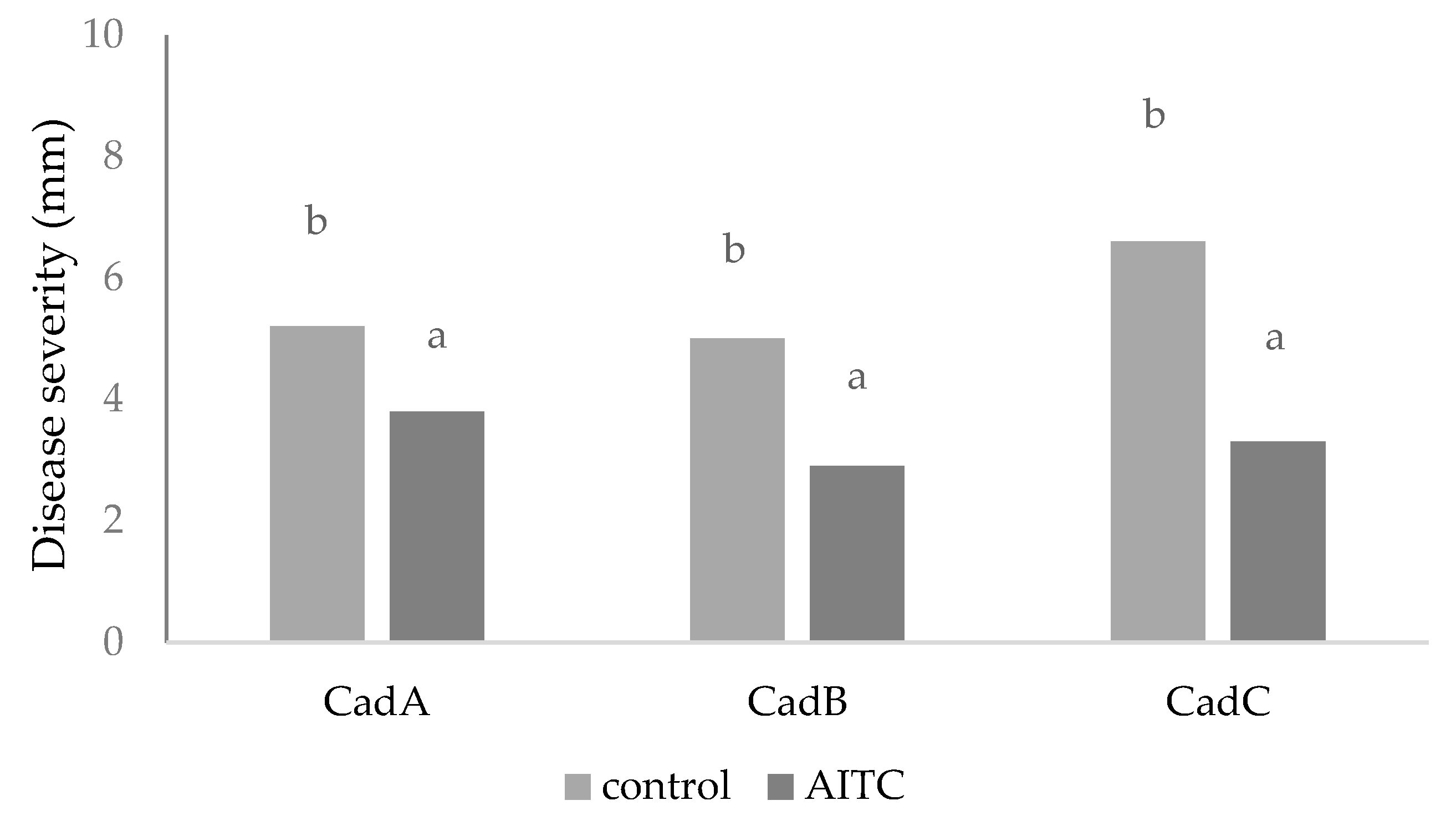
As an alternative treatment, the biofumigation in cold storage with AITC was carried out, always after testing its potential efficacy against
C. luteo-olivacea isolates mycelial growth that was total (100% of inhibition). The use of AITC needs a better understanding of its mode of action and toxicological effect to establish its safety [
25]. In Japan, synthetic AITC is registered as a food additive, and preliminary data show that food preserved with AITC vapor contains a very low residue of this compound [
26]. The biofumigation treatment determined a decrease of pectins amount and their chemical variation (increased of ester-acetylation) and a partial degradation of polysaccharides components (hemicellulose and free glucose), probably influencing the fungal behavior as monitored by FT-IR analysis. It is well known that different degrees of esterification can affect plant susceptibility to pathogens [
27,
28,
29,
30]. The treatment affected the fruit resistance, making it more susceptible to the fungal attack: an increase in protein content was a consequence of the volatile compounds produced by
A. pullulans in stone fruit [
18], which helped to control brown rot better.
On the contrary, the increases of waxes and phenolics, already observed in stone fruits [
18], is a typical response of the plant to fungal attack: waxes are responsible for the thickening of cell walls, while phenolics compounds enhance both the antioxidant activity and membrane integrity.
At the same time, the in vivo assay showed that the pathogens were stimulated to grow faster through an external sporification with respect to the control. In this case, the treatment showed two different modes of action if directly applied against fungal growth or on fruits. However, in vivo, the treatment reduced the fungal severity but not the incidence. The potential use of volatile fungicides to control post-harvest diseases requires a detailed examination of their biological activity and dispersion in fruit tissues and developing a formula that inhibits pathogens’ growth without producing phytotoxic effects on fruits.
Although the management of post-harvest diseases by employing BCAs or natural substances has been successfully evaluated on many species and against many pathogens, their efficacy could be increased by adopting an integrated strategy that combines both strategies and including antagonists and physical methods, including sometimes the use of fungicides at low doses [
31].










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}