RNA Helicase Mediates Competitive Fitness of Listeria monocytogenes on the Surface of Cantaloupe


Department of Food, Bioprocessing and Nutrition Sciences, North Carolina State University, Raleigh, NC 27604, USA
*
Author to whom correspondence should be addressed.
Horticulturae 2018, 4(4), 40; https://doi.org/10.3390/horticulturae4040040
Submission received: 19 October 2018
/
Revised: 5 November 2018
/
Accepted: 8 November 2018
/
Published: 14 November 2018
(This article belongs to the Special Issue Food Safety Pertinent to Fresh Produce)
Abstract
:Listeria monocytogenes is a foodborne pathogen that is implicated in numerous outbreaks of disease (listeriosis) via fresh produce. The genetic features of L. monocytogenes that allow adherence and growth on produce remain largely uncharacterized. In this study, two non-motile transposon mutants were characterized for attachment, growth, and survival on the surface of cantaloupe rind. One of the mutants, L1E4, harbored a single transposon insertion in a DEAD-box RNA helicase gene (lmo0866 homolog), while the other, M1A5, harbored an insertion in a gene from a flagellum biosynthesis and chemotaxis gene cluster (lmo0694 homolog). When inoculated alone, neither mutant was significantly impaired in growth or survival on the surface of cantaloupe at either 25 or 37 °C. However, when co-inoculated with the wildtype parental strain, the RNA helicase mutant L1E4 had a clear competitive disadvantage, while the relative fitness of M1A5 was not noticeably impacted. Genetic complementation of L1E4 with the intact RNA helicase gene restored relative fitness on cantaloupe. The findings suggest that the DEAD-box RNA helicase encoded by the lmo0866 homolog is critical for relative fitness of L. monocytogenes on cantaloupe. Mutant L1E4 was pleiotropic, being not only non-motile but also cold-sensitive and with reduced hemolytic activity, warranting further studies to elucidate the role of this helicase in the competitive fitness of L. monocytogenes on produce.
1. Introduction
Listeria monocytogenes is a facultative intracellular pathogen that causes the disease listeriosis, which is associated with severe symptoms and high mortality in susceptible humans. While serotypes 1/2a, 1/2b, and 4b account for the majority of human listeriosis cases, serotype 4b is most commonly linked to outbreaks. However, in 2011, a major multistate listeriosis outbreak in the United States associated with whole cantaloupe was found to be associated with strains of serotype 1/2a and 1/2b. This was the largest listeriosis outbreak to date in the United States and the first to involve cantaloupe [1].
The genetic features of L. monocytogenes that allow it to adhere and grow on the surface of produce remain largely uncharacterized. One feature that was examined in this study was the potential impact of motility. In L. monocytogenes, there are four to six flagella per cell with the production of flagella being temperature-dependent. At 37 °C, most L. monocytogenes strains do not produce flagella and are non-motile due to MogR repression of flagellin gene transcription, while at 30 °C and below L. monocytogenes is motile due to the inhibition of MogR by GmaR [2].
The impact of motility on the ability of L. monocytogenes to adhere to and grow on surfaces and form biofilms remains poorly understood. L. monocytogenes is able to contaminate food during processing partly because of its ability to adhere, grow, and persist on food contact surfaces [3,4,5]. While previous studies have investigated the relation between flagellar motility and biofilm formation [2,6], limited information is currently available on the potential role of motility in colonization of fresh produce.
When stored at 4 and 20 °C for 15 days, L. monocytogenes was able to survive on whole cantaloupe, but exhibited a 1 to 2 log10 CFU/cm2 decline in one study [7], while it was able to grow on the rind of cantaloupe in another investigation [8]. Previous work implicated both the formation of flagella and motility in the ability of L. monocytogenes to adhere to produce [9,10]. The objective of this study was to characterize two mariner-based non-motile mutants of L. monocytogenes 2011L-2858, a serotype 1/2b strain from the 2011 cantaloupe outbreak [1,8], regarding their colonization potential and fitness on cantaloupe rind.
2. Results and Discussion
Screening of approx. 1900 transposon mutants on soft agar identified nine that were motility-impaired. All nine mutants were confirmed to harbor single-copy transposon insertions. Of these nine, seven contained insertions in genes that are associated with flagella biosynthesis, and one mutant from this group, M1A5, was chosen for further testing. M1A5 harbored an insertion in the coding region of the lmo0694 homolog of strain 2858 (nt 161), a member of a motility and chemotaxis gene cluster. Of the two mutants to harbor a transposon insertion outside of flagellar biosynthesis genes, one, L1E4, harbored an insertion in the lmo0866 homolog (nt 981), encoding a DEAD-box RNA helicase, while the other mutant exhibited growth deficiencies that made it unfit for assessment in produce growth assays. Under phase contrast microscopy, the parental strain had tumbling motility characteristic of L. monocytogenes, while the mutant strains lacked motility. The lack of motility was also shown by loss of the capacity to swarm on soft agar at 25 °C (Figure 1 and data not shown).
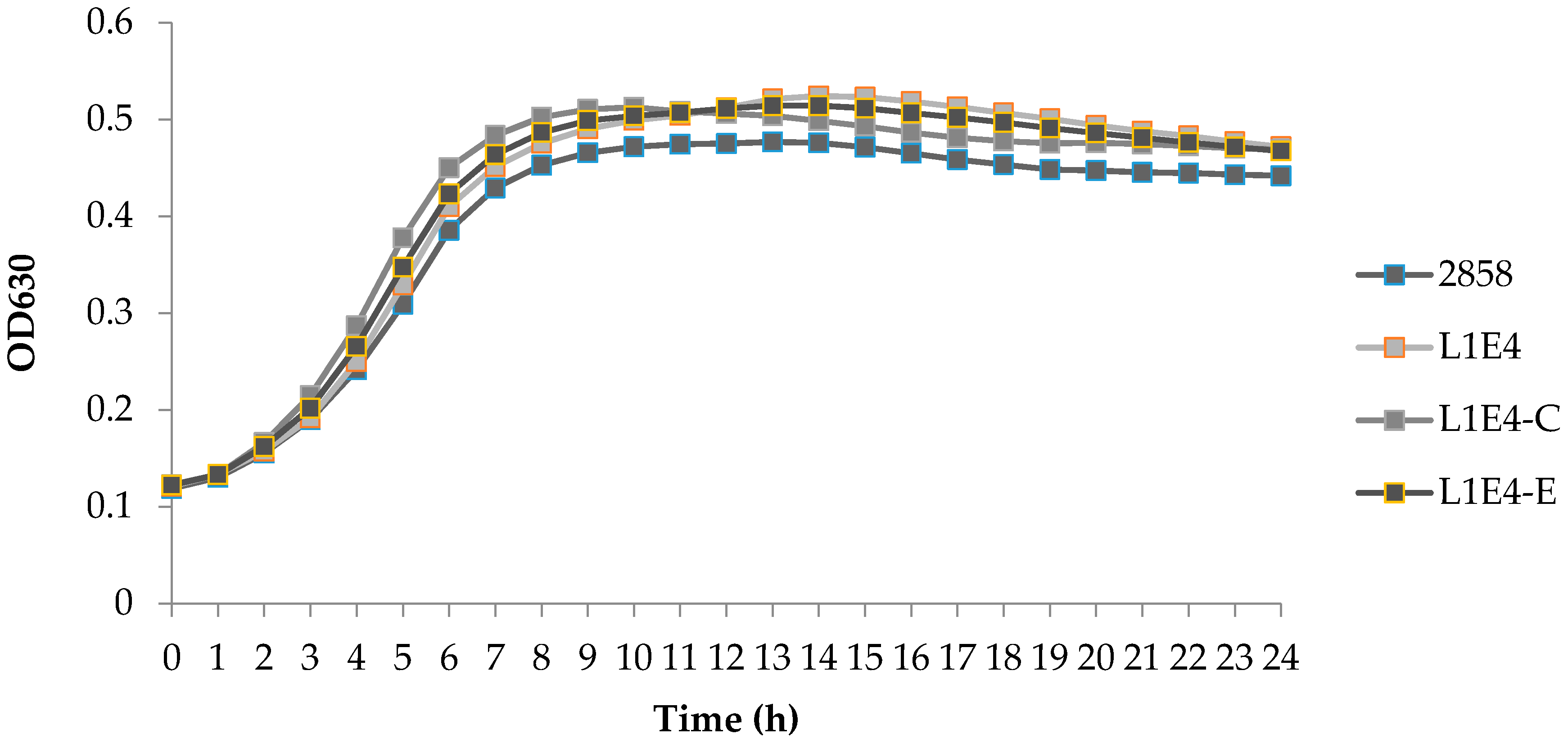
At 37 °C each of the mutants grew similarly to the parental strain in liquid medium (brain heart infusion, BHI) (Figure 2). Growth rates for 2858 and L1E4 in BHI at 37 °C were 0.103 and 0.109, respectively. On agar (Brain heart infusion agar, BHIA), the colonies formed by L1E4 were smaller than those of the parental strain or M1A5, and also had a slightly concaved appearance (data not shown). This difference in colony size may reflect a surface-dependent impairment in growth, since no differences were noted in growth in liquid medium (BHI). Growth rates for 2858 and L1E4 in BHI at 28 °C were 0.241 and 0.246, respectively.
A pronounced difference was noted between L1E4 and M1A5 in capacity to grow at low temperatures. The helicase mutant L1E4 was cold-sensitive, completely unable to grow in liquid or on agar media at 4 or 10 °C, as previously shown for other helicase mutants [11,12], in contrast to M1A5, which grew at these low temperatures similarly to strain 2858. Lastly, L1E4 was less hemolytic on blood agar plates than the parent strain or mutant M1A5, as also reported by others for helicase mutants of L. monocytogenes [13]. The pleiotropic phenotype of L1E4 is in agreement with the requirement for functional RNA helicases in diverse phenotypes of L. monocytogenes [11,12,13,14,15]. Four DEAD-box RNA helicase genes (lmo0866, lmo1246, lmo1450, lmo1722) have been identified in L. monocytogenes EGD-e [14,15,16]. However, the precise mechanisms mediated the impact of the different RNA helicases of L. monocytogenes in capacity to grow at low temperature, motility, and other phenotypes remain poorly understood [14,15,16].
2.1. Impact of the Mutations on Growth and Competitive Fitness of L. monocytogenes on Cantaloupe Rind
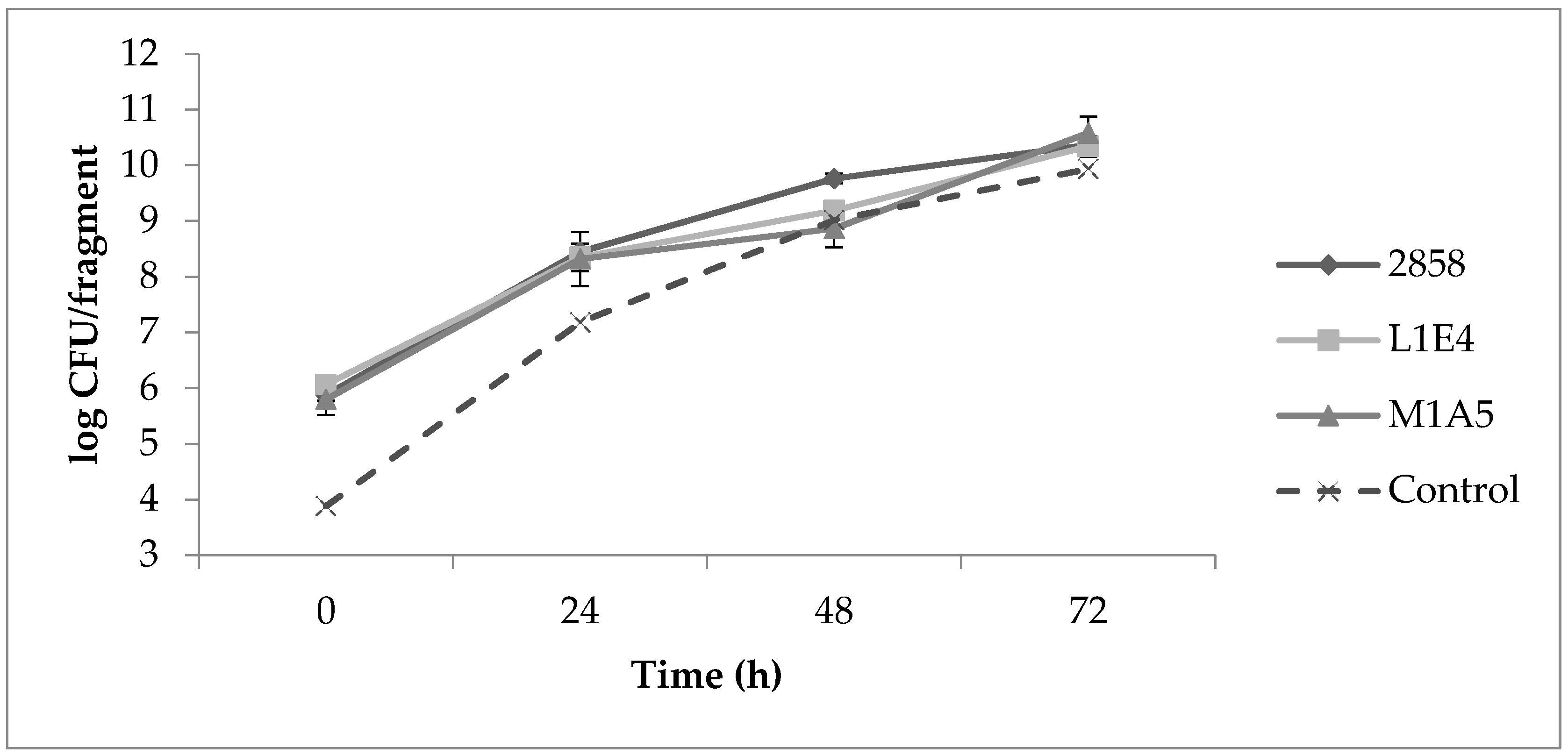
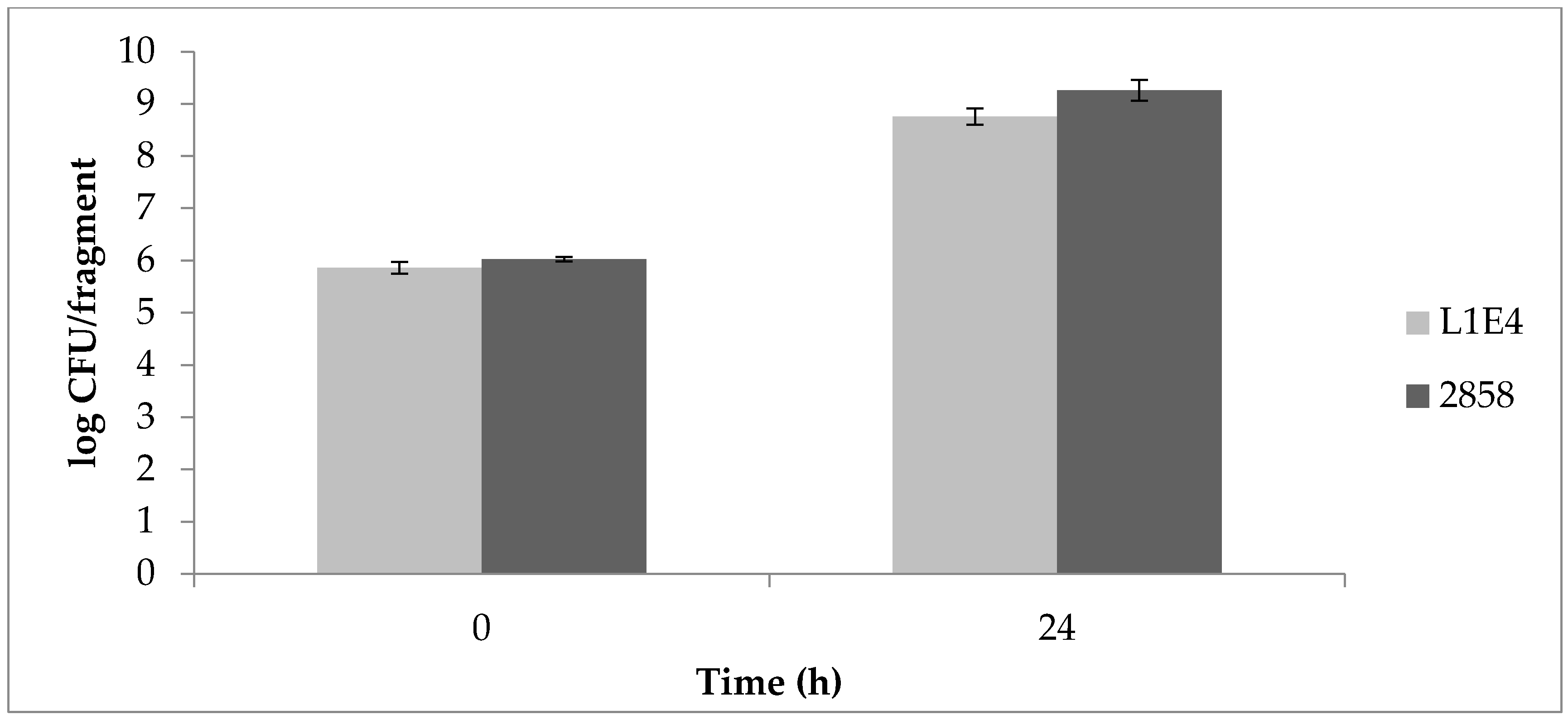
Uninoculated control fragments were negative for Listeria throughout the study. Determinations of aerobic plate counts (APCs) on uninoculated cantaloupe fragments indicated on average 105 CFU/fragment prior to inoculation with a 4-log increase after 24 h at 25 °C, followed by further increases after 48 and 72 h (Figure 3). Initial Listeria counts on MOX were approx. 106 CFU/fragment for each of the strains individually as well as for the mixed cultures. The two mutant strains grew similarly to the parental strain 2858. After 24 h, Listeria CFU/fragment increased by approx. 2 logs, with further increases after 48 and 72 h (Figure 3). At 37 °C, Listeria populations increased by approx. 3 logs after 24 h (Figure 4).
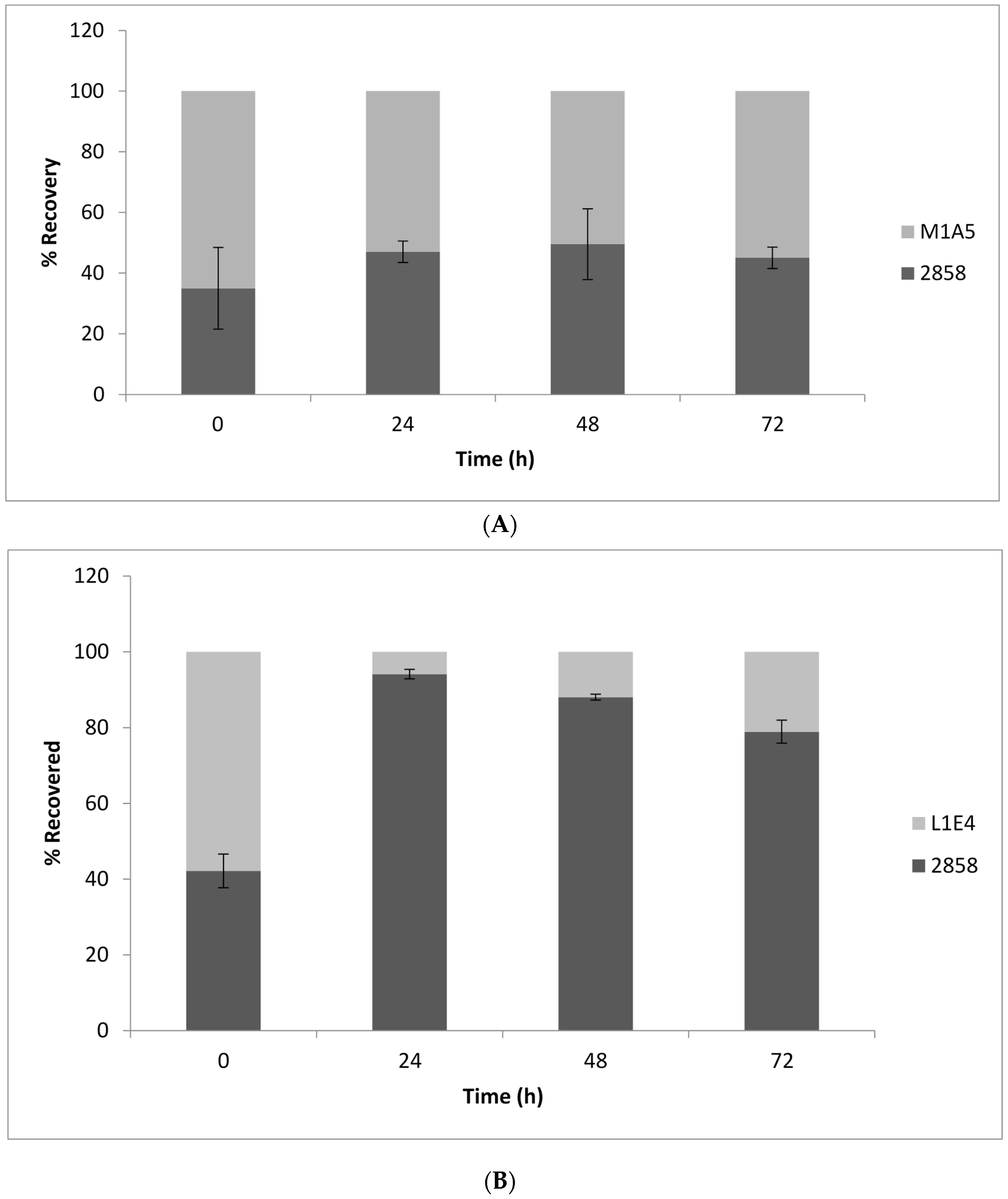
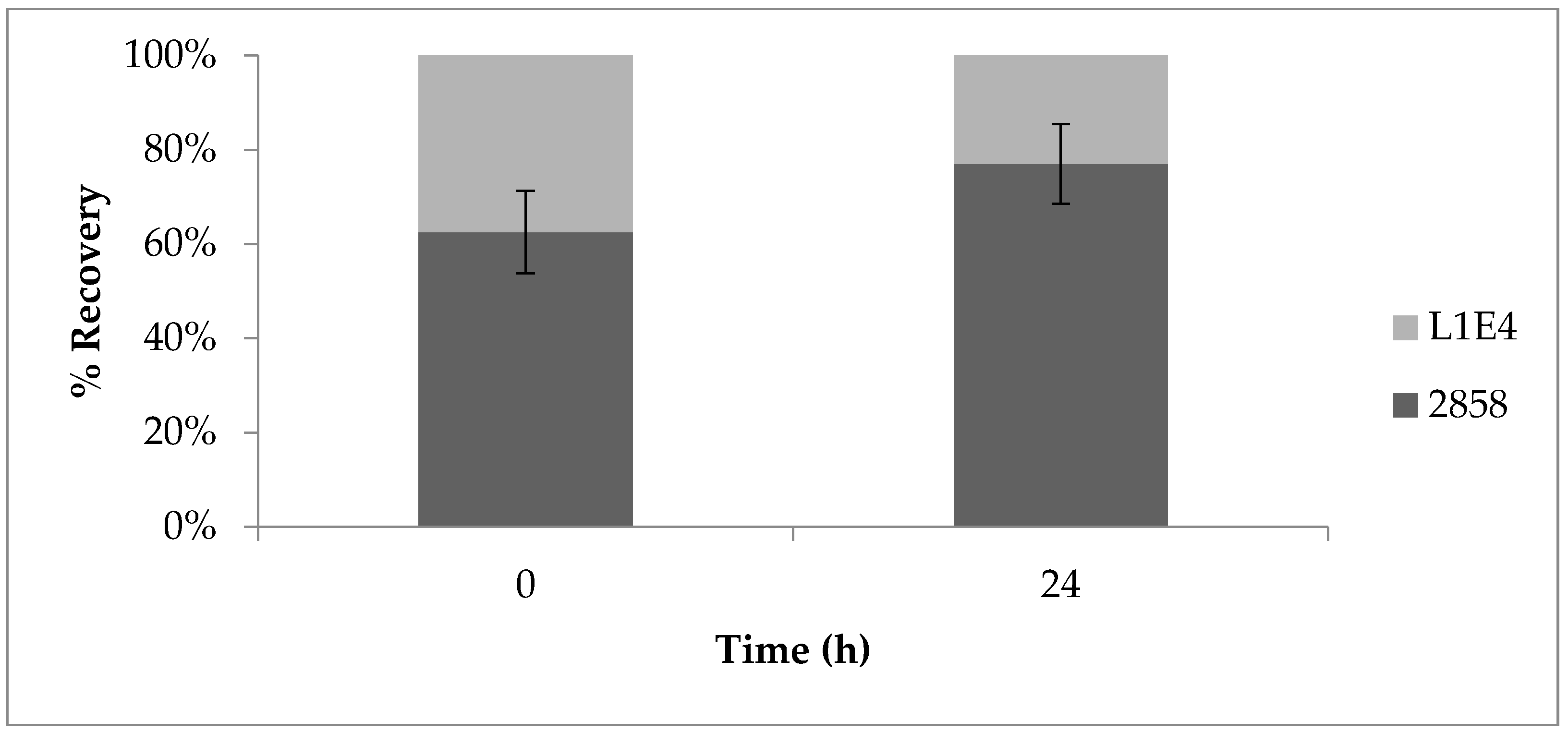
To determine potential impacts of the mutations on the competitive fitness of the bacteria on cantaloupe, the rind was inoculated with 1:1 mixtures of the parental strain and each of the mutants and incubated at 25 or 37 °C. Screening of L. monocytogenes colonies from the MOX plates for growth on BHIA with erythromycin (BHI-Em5) revealed that, for M1A5, the ratio of parental strain to mutant remained around 50% at all tested time points (Figure 5A), suggesting that the relative fitness of this mutant did not differ significantly from that of the wild-type parental strain. However, L1E4 was significantly impaired (p < 0.0001) in its recovery post-inoculation when in combination with the wild-type parental strain (Figure 5B). Similar trends were observed at 37 °C (Figure 6); after 24 h, L1E4 was less commonly recovered from the cantaloupe fragments than the wildtype parental strain.
2.2. Genetic Complementation of Mutant L1E4 Confirms Impact of the DEAD-Box RNA Helicase Encoded by lmo0866 in Competitive Fitness on Cantaloupe
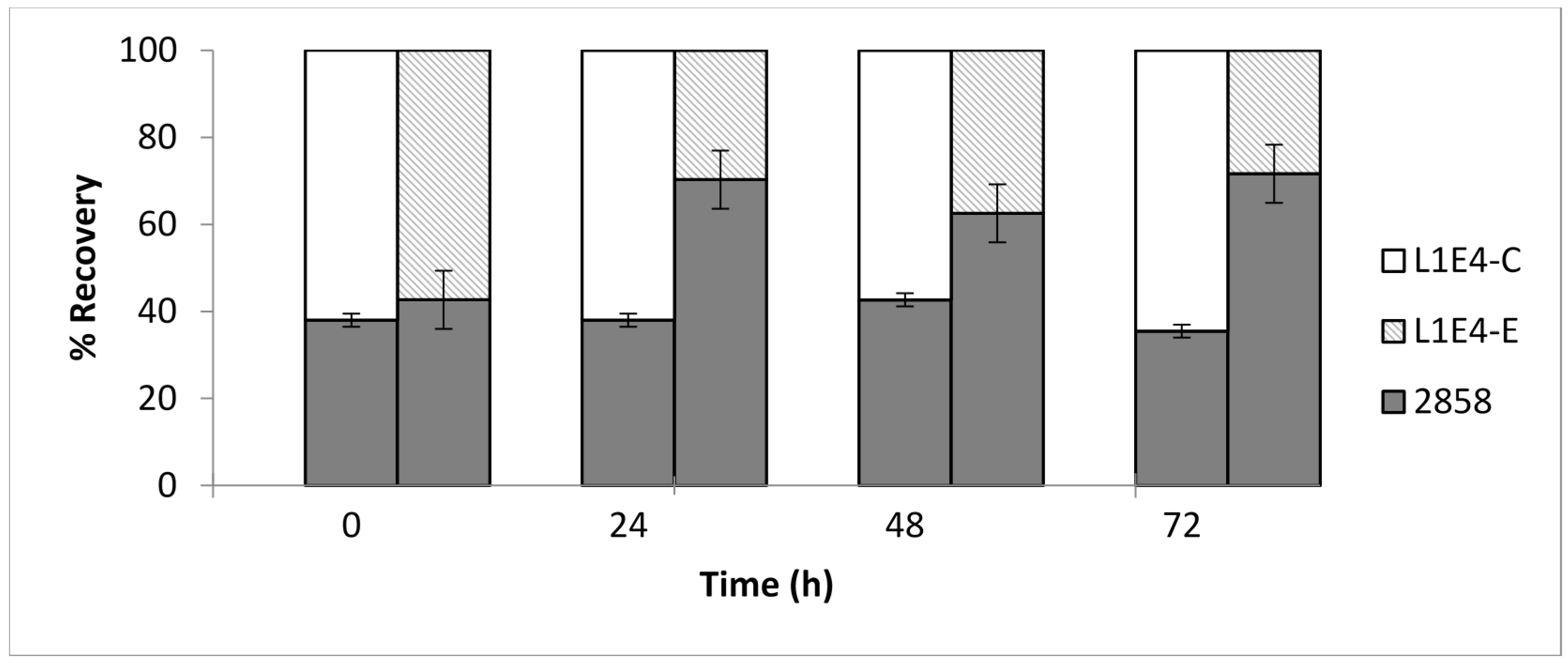
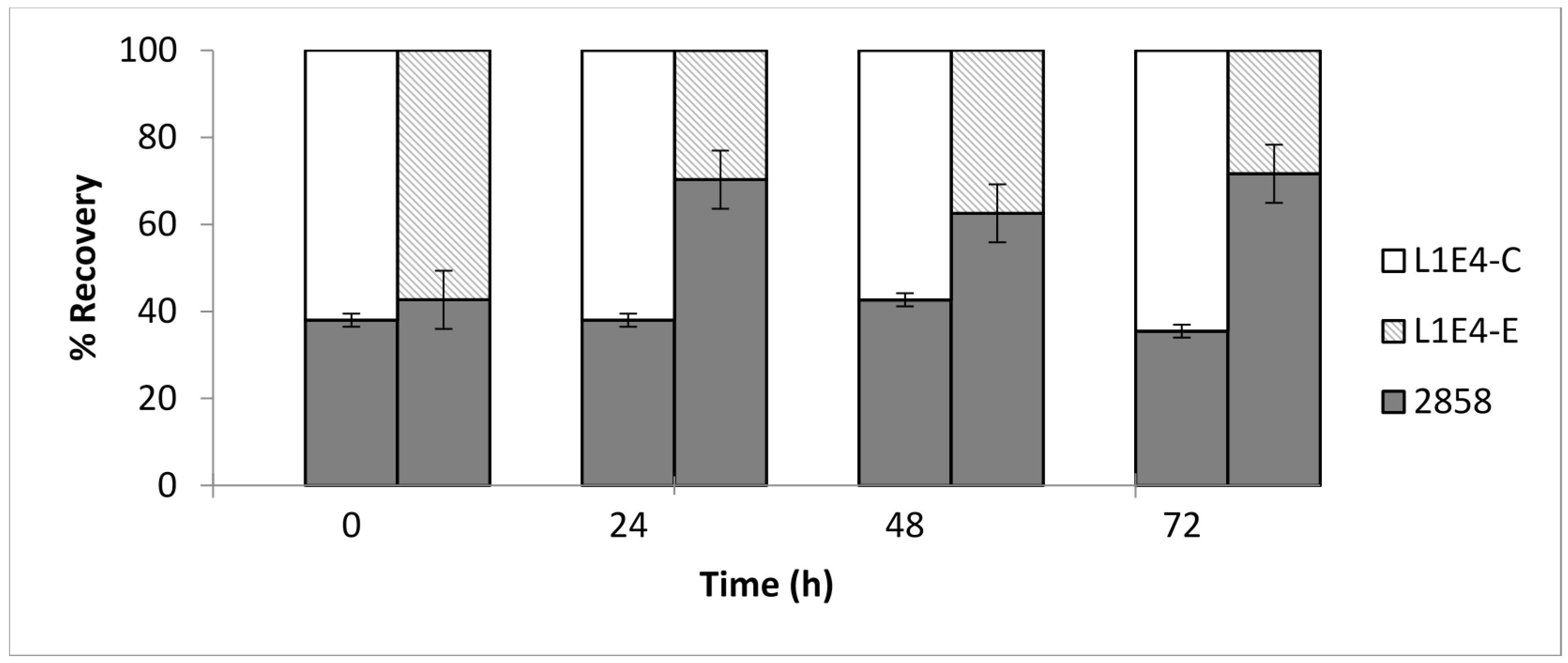
Mixed-culture testing of L1E4-C, a genetically complemented derivative of L1E4 harboring an intact copy of the DEAD-box RNA helicase gene lmo0866 from L. monocytogenes 2858, revealed the restoration of competitive fitness on cantaloupe to levels similar to those of the wild type parental strain. The ratio of the genetically complemented derivative L1E4-C to the parental strain 2858 remained stable over 72 h at 25 °C (Figure 7). In contrast, L1E4-E, which harbors the empty vector pPL2, was less likely to be recovered from the inoculated fragments than the parental strain (p = 0.0064) (Figure 7), similarly to what was described above for L1E4.
Similarly to other helicase mutants of L. monocytogenes [11,12,13,14,15], L1E4 was impaired in multiple phenotypes. Originally isolated on the basis of its impaired motility, L1E4 was in addition cold-sensitive, had reduced colony size on agar media, and it was less hemolytic on blood agar plates than its parental strain. Since mutant M1A5 was not impaired in competitive fitness on cantaloupe even though it was non-motile, attributes other than the loss of motility might be responsible for the impaired competitive fitness of the helicase mutant L1E4.
The roles of RNA helicases are complex, as evidenced by the pleiotropic nature of the L1E4 mutant in which the lmo0866 homolog was inactivated by the insertion of the mariner-based transposon. It is possible, for instance, that this helicase is essential for the expression, or RNA maturation of, key genes controlling motility, capacity to grow at low temperature, and hemolytic activity, among other phenotypes. Given that this family of genes has been shown to be of importance in diverse stress scenarios [14], competition in a nutrient poor environment and in the presence of other microbiota could make them particularly necessary in competitive growth on the surface of produce. Further studies are needed to elucidate such attributes and characterize the mechanisms that may impact competitive fitness of L. monocytogenes on cantaloupe and other types of fresh produce. Elucidation of such mechanisms will be valuable in the design of novel tools and strategies to reduce the contribution of fresh produce to human listeriosis.
3. Materials and Methods
3.1. Bacterial Strains and Growth Conditions
Bacterial strains that were used in this study are listed in Table 1. Mutant libraries were constructed as described previously [17,18] in the serotype 1/2b strain 2011L-2858 (hereafter designated as strain 2858), isolated from cantaloupe that were implicated in the 2011 cantaloupe outbreak [1,8]. Listeria monocytogenes was routinely grown in brain heart infusion broth (BHI) (Becton, Dickinson & Co., Sparks, MD, USA) or on brain heart infusion agar (BHIA) with BHI supplemented with 1.2% agar (Bacto-agar, Becton, Dickinson & Co.). When needed, erythromycin (Em, MP Biomedicals, Solon, OH, USA) was added at 5 µg/mL in BHI (BHI-Em5) or in BHIA (BHIA-Em5) and kanamycin (Km, Fisher Scientific, Fair Lawn, NJ, USA) was added at 10 µg/mL to BHI (BHI-Km10) and BHIA (BHIA-Km10). L. monocytogenes strains were routinely grown in BHI at 37 °C in standing cultures and preserved at −80 °C in BHI with 20% glycerol.
3.2. Cantaloupe Adherence, Growth and Survival Assay
Strain 2858 and the motility-impaired mutants M1A5 and L1E4 were grown in 5 mL of BHI at 37 °C for approx. 16 h to a concentration of approx. 108 CFU/mL. The overnight cultures were then centrifuged for three min at 16,000 rpm at room temperature. The supernatant was discarded and the bacteria were washed twice in 5 mL of sterile deionized water. The inoculum was prepared by diluting 50 µL of the washed cell suspension into 5 mL of BHI, resulting in a suspension with approx. 106 CFU/mL.
Cantaloupes that were used in this study were purchased from local grocery stores in Raleigh, NC. The majority of the cantaloupes were imported from the Honduras or Guatemala during the winter months and were from North Carolina, Georgia, and California during summer months. At retail, cantaloupes were displayed at room temperature and they were kept at room temperature during transportation to the laboratory where they were used within 2 h of purchase. Rind fragments (5 × 5 × 1 cm) were prepared as described [8] and placed rind side up in groups of four into labeled, sterile plastic petri dishes (100-mm diameter, Fisher Scientific, Pittsburgh, PA, USA). Cantaloupes were subject to selective enrichments to rule out native contaminating L. monocytogenes, as described in Azizoglu et al. (2014) [19]. Each fragment was spot-inoculated as described [8] in 10 evenly separated droplets of 10 µL each (total inoculum 100 µL, corresponding to approx. 105 CFU/fragment); two fragments were inoculated for each time point, and several fragments similarly spotted with sterile water as controls. The petri dishes were sealed with parafilm and stored at either room temperature (25 °C) or 37 °C. This temperature (37 °C) was chosen as it alleviates potential differences in motility, since flagellum production is repressed in L. monocytogenes at this temperature [20,21].
Bacteria on the fragments were enumerated 30 min after inoculation (time 0), and at 24, 48, and 72 h for fragments stored at room temperature or at 30 min and 24 h after inoculation for fragments incubated at 37 °C. For each time point, two inoculated fragments per strain were individually placed in 50-mL Falcon™ conical centrifuge tubes (Corning Life Sciences, Corning, NY, USA) and vortexed at top speed (Vortex Genie 2, Diagger, Vernon Hills, IL, USA) in 10 mL of sterile deionized water for two min, and enumerations of total aerobic plate counts (APCs) and Listeria were done on BHIA or on modified Oxford Listeria selective agar (MOX, Oxoid, Hampshire, UK), respectively, as described [8]. Each mutant/wild type comparison for adherence and growth was done in at least three independent trials.
3.3. Competitive Fitness Assessments
Mixed-strain inoculations were prepared by inoculating 25 µL of strain 2858 and 25 µL of each strain (approx. 105 CFU each) into 5 mL of sterile deionized water. This diluted mixture was then used to inoculate the surface of cantaloupe fragments, as previously described. To enumerate the mutant populations from cantaloupe fragments inoculated with these 1:1 mixtures, colonies from the MOX plates were inoculated into 96-well plates containing 200 µL of BHI and incubated for approx. 16 h. The 96-well cultures were then stamped in batches of 48 using a sterile microplate replicator onto agar media with and without erythromycin, and growth was assessed following incubation at 37 °C for 48 h. Each competitive fitness assay was done in at least two independent trials.
3.4. Genetic Complementation of L1E4
Genetic complementation of L1E4 was carried out utilizing Listeria shuttle vector pPL2, as previously described [18,22]. A 2012-bp fragment was PCR- amplified using primers L1E4compF (5′-CTAAGGTACCTTCGATAATAGCAAGGAAAGAT-3′) and L1E4compR (5′-GTAACCCGGGCCACACTCCCGTATCCTTAA-3′), and purified using the QiaQuick PCR Clean Up kit (Qiagen, Valencia, CA, USA). This region began 341 bp upstream of the start codon of LMOf2365_0884, and included putative promoter sequences identified by Softberry BPROM software (http://www.softberry.com/berry.phtml?topic=bprom&group=programs&subgroup=gfindb). Primers L1E4compF and L1E4compR were designed with restriction sites for endonucleases KpnI and XmaI, respectively, whose cut sites are underlined in the primer sequences. The purified fragment was then digested with KpnI and XmaI and ligated into similarly digested vector pPL2 using T4 DNA ligase (Promega, Madison, WI, USA). The recombinant plasmid was electroporated into E. coli strain S17-1, which enabled its conjugative transfer into mutant L1E4, as described previously [18]. Chromosomal insertion was confirmed using primers NC16 (5′-GTCAAAACATACGCTCTTATC-3′), located on the chromosome of L. monocytogenes in the tRNA Arg gene, and catR (5′-GACAATTGGAAGAGAAAAGAG-3′), located in the chloramphenicol resistance gene on the backbone of the vector. The same methodology was used for the insertion and confirmation of the empty vector pPL2 into L1E4.
3.5. In Vitro Growth Comparison of Strain 2858 and Motility Impaired Mutants
Overnight cultures of the parental strain 2858 and the two motility mutants used in this study, L1E4 and M1A5, were grown in 5 mL of BHI at 37 °C for approx. 16 h to a concentration of approx. 108 CFU/mL. The strains (4 µL) were inoculated in 200 µL of BHI in individual wells of a 96-well plate (Corning Inc., Corning, NY, USA) with each strain being tested in triplicate. The plate was then placed into a BioTek microplate reader (BioTek Instrument, Inc., Winooski, VT, USA) and incubated for 24 h at either 28 °C or 37 °C with optical density at 630 nm (OD630) measurements taken at 1-h intervals with 10 s of shaking before each measurement. Growth rates for each of the motility mutants and the parental strain 2858 were reported relative to the optical density of sterile BHI broth, used as a control. To determine growth rates, individual growth curves were processed in sequential sets of n data values, as previously described by Breidt et al. in 1994 [23].
3.6. Statistical Analysis
The optical densities of each of the strains were entered in to a custom algorithm created by F. Breidt using Matlab software (The Mathworks, Inc., Natick, MA, USA) to derive the relative growth rates and the standard errors for each strain. The growth rates of the individual strains were fit into a General Linear Model (GLM) of SAS (SAS Institute, Cary, NC, USA) to compute the analysis of variance. A general linear model was also fit to the CFU/fragment values for each of the individual strains that were inoculated on cantaloupe as well as the mixed-culture ratios from the surface of the cantaloupe.
4. Conclusions
The current study suggests that the DEAD box RNA helicase encoded by the lmo0866 homolog of a Listeria monocytogenes strain from the 2011 cantaloupe outbreak is important to ensure competitive fitness of L. monocytogenes on cantaloupe rind. A transposon mutant in this gene, originally identified based on its loss of motility, exhibited significantly impaired fitness on cantaloupe rind in comparison to the wildtype parental strain, and fitness was restored upon genetic complementation. Our findings with a different non-motile mutant harboring an insertion in an unrelated gene suggest that the impaired fitness of the RNA helicase mutant was not due to lack of motility. The RNA helicase encoded by lmo0866 is essential for multiple phenotypes of L. monocytogenes, including motility, capacity to grow at refrigeration temperature, and wild-type levels of hemolytic activity. This necessitates additional studies to elucidate the mechanism(s) via which this RNA helicase impacts fitness of L. monocytogenes on fresh produce.
Author Contributions
R.P. carried out experiments involving growth and survival on produce and original manuscript preparation; C.P. performed mutant library creation, mutant phenotypic assays and manuscript revision; S.K. contributed experimental design and manuscript revision.
Funding
This material is based upon work that was partially supported by the National Institute of Food and Agriculture, U.S. Department of Agriculture, under USDA NIFA award number 2012-67017-30218.
Acknowledgments
We thank Fred Breidt for feedback and access to equipment and software utilized in this study. We appreciate the feedback and support of Mira Rakic-Martinez and thank Ben Costollo for assistance in screening the mutant library.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McCollum, J.T.; Cronquist, A.B.; Silk, B.J.; Jackson, K.A.; O’Connor, K.A.; Cosgrove, S.; Gossack, J.P.; Parachini, S.S.; Jain, N.S.; Ettestad, P.; et al. Multistate outbreak of listeriosis associated with cantaloupe. N. Engl. J. Med. 2013, 369, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Lemon, K.P.; Higgins, D.E.; Kolter, R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J. Bacteriol. 2007, 189, 4418–4424. [Google Scholar] [CrossRef] [PubMed]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [PubMed]
- Chae, M.S.; Schraft, H. Comparative evaluation of adhesion and biofilm formation of different Listeria monocytogenes strains. Int. J. Food Microbiol. 2000, 62, 103–111. [Google Scholar] [CrossRef]
- Chae, M.S.; Schraft, H.; Truelstrup Hansen, L.; Mackereth, R. Effects of physicochemical surface characteristics of Listeria monocytogenes strains on attachment to glass. Food Microbiol. 2006, 23, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Todhanakasem, T.; Young, G.M. Loss of flagellum-based motility by Listeria monocytogenes results in formation of hyperbiofilms. J. Bacteriol. 2008, 190, 6030–6034. [Google Scholar] [CrossRef] [PubMed]
- Ukuku, D.O.; Fett, W. Behavior of Listeria monocytogenes Inoculated on cantaloupe surfaces and efficacy of washing treatments to reduce transfer from rind to fresh-cut pieces. J. Food Prot. 2002, 65, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Rakic-Martinez, M.; Osborne, J.; Jayeola, V.O.; Katic, V.; Kathariou, S. Capacity of Listeria monocytogenes strains from the 2011 cantaloupe outbreak to adhere, survive, and grow on cantaloupe. J. Food Prot. 2016, 79, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Gorski, L.; Duhé, J.M.; Flaherty, D. The use of flagella and motility for plant colonization and fitness by different strains of the foodborne pathogen Listeria monocytogenes. PLoS ONE 2009, 4, e5142. [Google Scholar] [CrossRef] [PubMed]
- Gorski, L.; Palumbo, J.D.; Mandrell, R.E. Attachment of Listeria monocytogenes to radish tissue is dependent upon temperature and flagellar motility. Appl. Environ. Microbiol. 2003, 69, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Markkula, A.; Mattila, M.; Lindström, M.; Korkeala, H. Genes encoding putative DEAD-box RNA helicases in Listeria monocytogenes EGD-e are needed for growth and motility at 3 °C. Environ. Microbiol. 2012, 14, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Azizoglu, R.O.; Kathariou, S. Inactivation of a cold-induced putative rna helicase gene of Listeria monocytogenes is accompanied by failure to grow at low temperatures but does not affect freeze-thaw tolerance. J. Food Prot. 2010, 73, 1474–1479. [Google Scholar] [CrossRef] [PubMed]
- Netterling, S.; Bäreclev, C.; Vaitkevicius, K.; Johansson, J. RNA Helicase important for Listeria monocytogenes hemolytic activity and virulence factor expression. Infect. Immun. 2016, 84, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Markkula, A.; Lindström, M.; Johansson, P.; Björkroth, J.; Korkeala, H. Roles of four putative DEAD-box RNA helicase genes in growth of Listeria monocytogenes EGD-e under heat, pH, osmotic, ethanol, and oxidative stress conditions. Appl. Environ. Microbiol. 2012, 78, 6875–6882. [Google Scholar] [CrossRef] [PubMed]
- Bäreclev, C.; Vaitkevicius, K.; Netterling, S.; Johansson, J. DExD-box RNA-helicases in Listeria monocytogenes are important for growth, ribosomal maturation, rRNA processing and virulence factor expression. RNA Biol. 2014, 11, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- Glaser, P.; Frangeul, L.; Buchrieser, C.; Rusniok, C.; Amend, A.; Baquero, F.; Berche, P.; Bloecker, H.; Brandt, P.; Chakraborty, T.; et al. Comparative genomics of Listeria species. Science 2001, 294, 849–852. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Bitar, A.P.; Marquis, H. A mariner-based transposition system for Listeria monocytogenes. Appl. Environ. Microbiol. 2007, 73, 2758–2761. [Google Scholar] [CrossRef] [PubMed]
- Parsons, C.; Lee, S.; Jayeola, V.; Kathariou, S. Novel cadmium resistance determinant in Listeria monocytogenes. Appl. Environ. Microbiol. 2017, 83, e02580-16. [Google Scholar] [CrossRef] [PubMed]
- Azizoglu, R.O.; Gorski, L.; Kathariou, S. Isolation of Listeria monocytogenes from food and water: Official and experimental protocols. In Current Protocols in Microbiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; Volume 33, pp. 9B.5.1–9B.5.19. [Google Scholar]
- Peel, M.; Donachie, W.; Shaw, A. Temperature-dependent expression of flagella of Listeria monocytogenes studied by electron microscopy, SDS-PAGE and western blotting. J. Gen. Microbiol. 1988, 134, 2171–2178. [Google Scholar] [CrossRef] [PubMed]
- Way, S.S.; Thompson, L.J.; Lopes, J.E.; Hajjar, A.M.; Kollmann, T.R.; Freitag, N.E.; Wilson, C.B. Characterization of flagellin expression and its role in Listeria monocytogenes infection and immunity. Cell. Microbiol. 2004, 6, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Lauer, P.; Chow, M.Y.; Loessner, M.J.; Portnoy, D.A.; Calendar, R. Construction, characterization, and use of two Listeria monocytogenes site-specific phage integration vectors. J. Bacteriol. 2002, 184, 4177–4186. [Google Scholar] [CrossRef] [PubMed]
- Breidt, F.; Romick, T.L.; Fleming, H.P. A rapid method for the determination of bacterial growth kinetics. J. Rapid Methods Autom. Microbiol. 1994, 3, 59–68. [Google Scholar] [CrossRef]
Figure 1.
Swarming of (A) 2858, (B) L1E4, (C) genetically complemented mutant L1E4-C, and (D) L1E4-E with empty vector pPL2. Each strain was spotted in duplicate on a TSA soft agar plate which was then incubated at 25 °C for 48 h, as described in Materials and Methods.
Figure 1.
Swarming of (A) 2858, (B) L1E4, (C) genetically complemented mutant L1E4-C, and (D) L1E4-E with empty vector pPL2. Each strain was spotted in duplicate on a TSA soft agar plate which was then incubated at 25 °C for 48 h, as described in Materials and Methods.
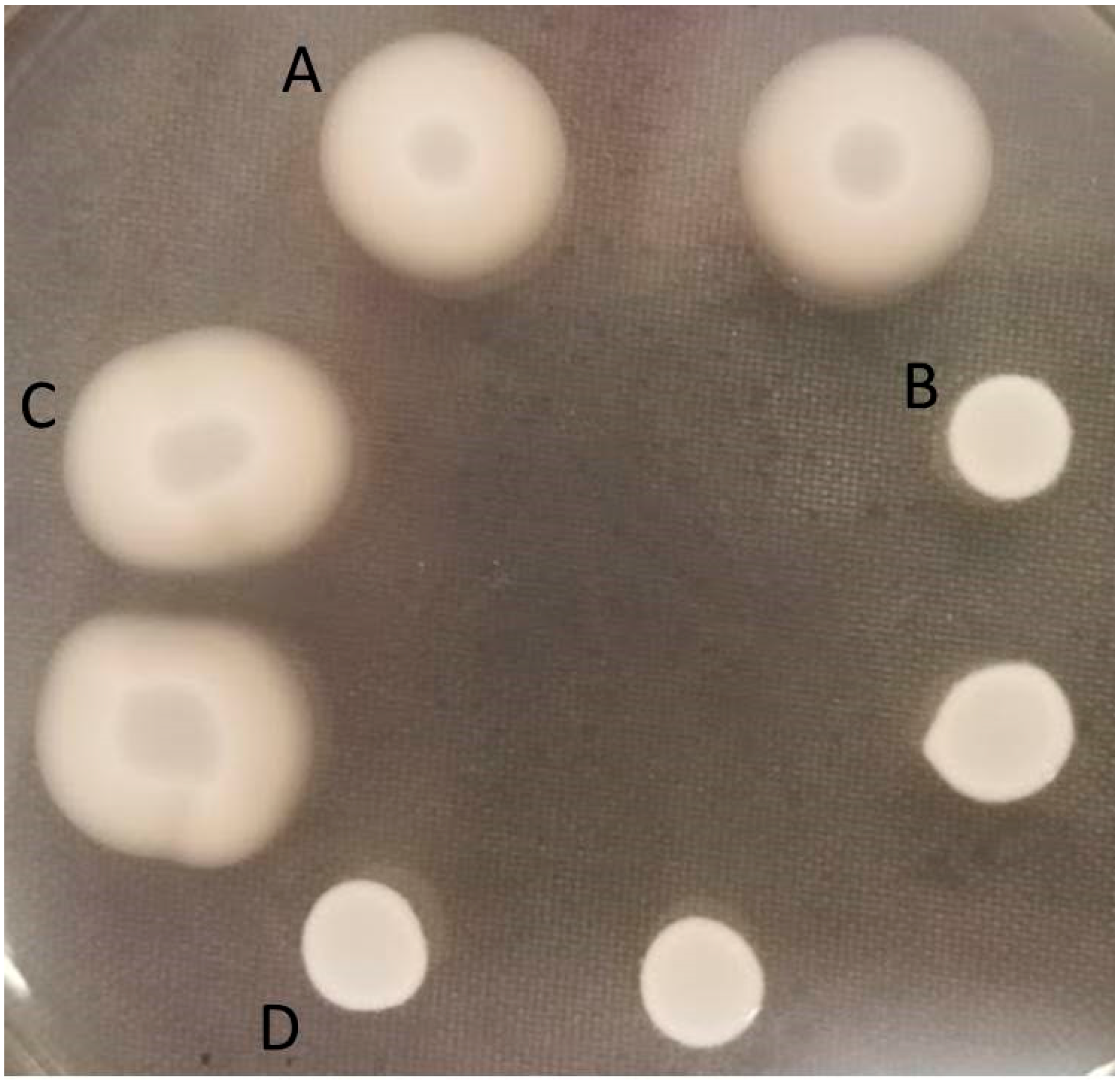
Figure 2.
Growth of L1E4, genetically complemented derivative and parental strain 2858 at 37 °C. L1E4-C and L1E4-E indicate the genetically complemented L1E4 and L1E4 harboring just the empty vector, respectively. Cultures were incubated in BHI at 37 °C and A630 was measured at hourly intervals using a BioTek microplate reader, as described in Materials and Methods. Results are the average of three wells assessed in one trial.
Figure 2.
Growth of L1E4, genetically complemented derivative and parental strain 2858 at 37 °C. L1E4-C and L1E4-E indicate the genetically complemented L1E4 and L1E4 harboring just the empty vector, respectively. Cultures were incubated in BHI at 37 °C and A630 was measured at hourly intervals using a BioTek microplate reader, as described in Materials and Methods. Results are the average of three wells assessed in one trial.
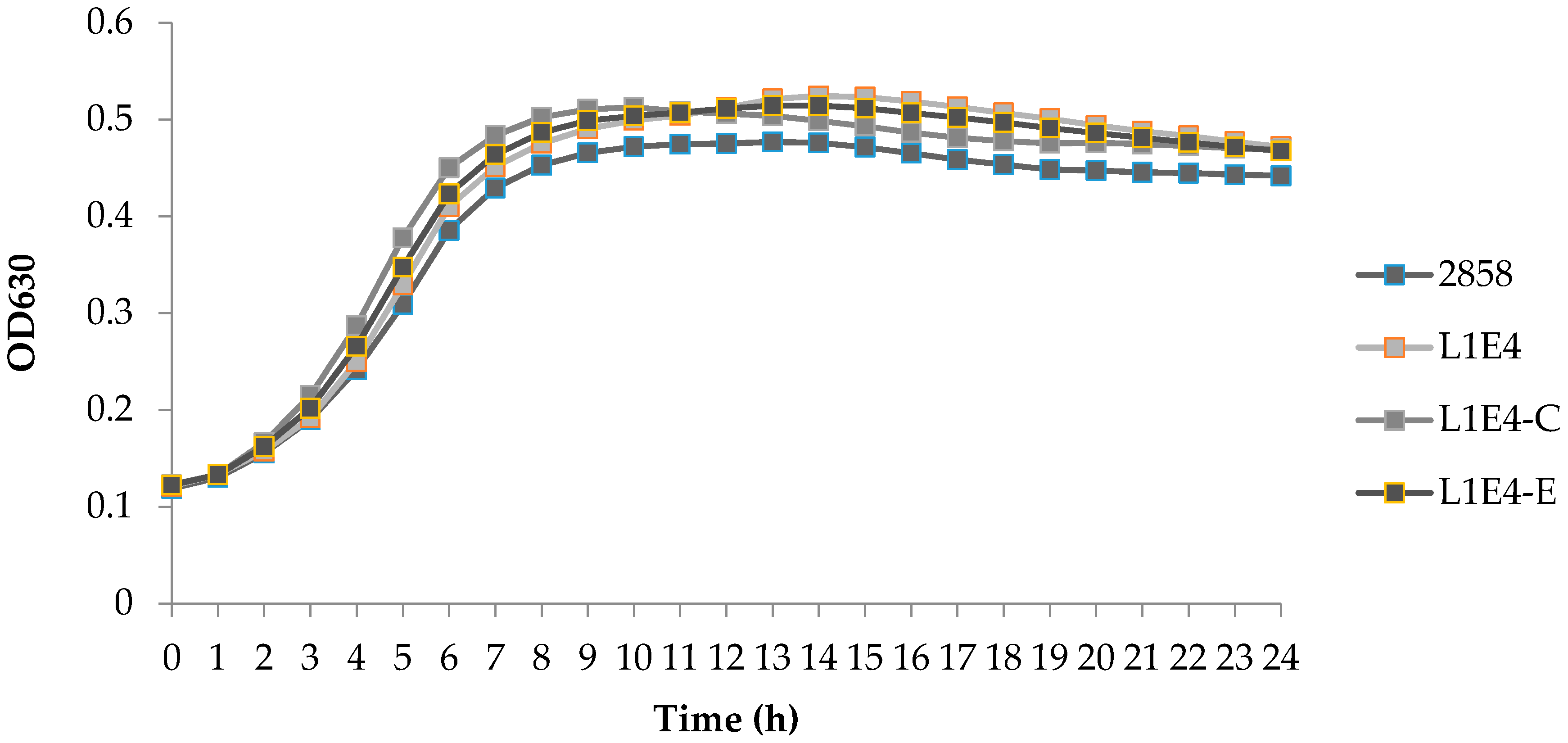
Figure 3.
Increases in aerobic plate counts (APC) and Listeria populations on cantaloupe. Cantaloupe rind fragments were prepared and inoculated as described in Materials and Methods, with time 0 corresponding to 30 min post-inoculation, and subsequently incubated at 25 °C over 72 h. APCs from uninoculated fragments (“control”) were determined on BHI, while Listeria populations from inoculated fragments were enumerated on modified Oxford medium (MOX), as described in Materials and Methods.
Figure 3.
Increases in aerobic plate counts (APC) and Listeria populations on cantaloupe. Cantaloupe rind fragments were prepared and inoculated as described in Materials and Methods, with time 0 corresponding to 30 min post-inoculation, and subsequently incubated at 25 °C over 72 h. APCs from uninoculated fragments (“control”) were determined on BHI, while Listeria populations from inoculated fragments were enumerated on modified Oxford medium (MOX), as described in Materials and Methods.
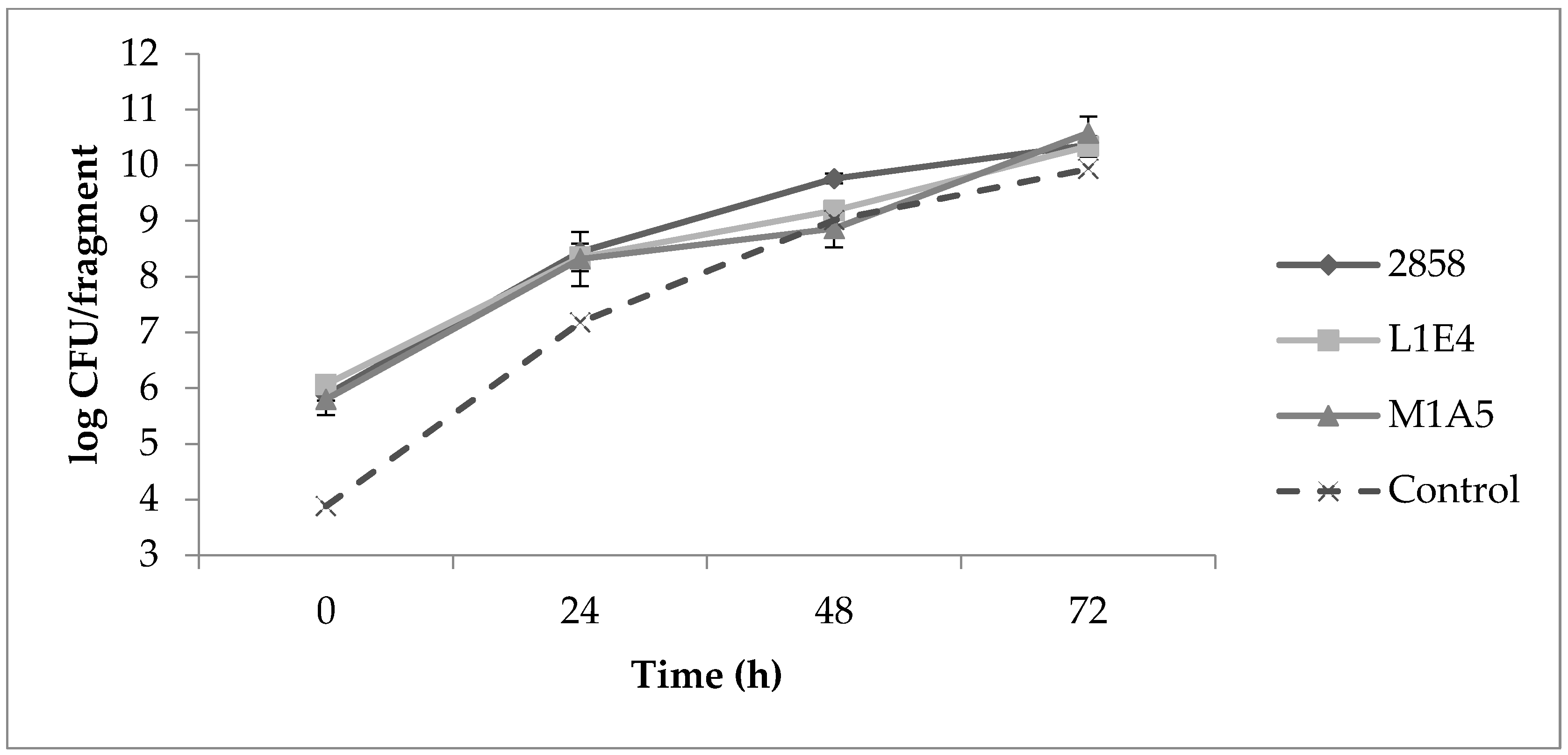
Figure 4.
Increases in Listeria counts on cantaloupe at 37 °C. Cantaloupe rind fragments were prepared and inoculated as described in Materials and Methods, with time 0 corresponding to 30 min post-inoculation, and subsequently incubated for 24 h at 37 °C. Populations were enumerated on MOX, as described in Materials and Methods.
Figure 4.
Increases in Listeria counts on cantaloupe at 37 °C. Cantaloupe rind fragments were prepared and inoculated as described in Materials and Methods, with time 0 corresponding to 30 min post-inoculation, and subsequently incubated for 24 h at 37 °C. Populations were enumerated on MOX, as described in Materials and Methods.
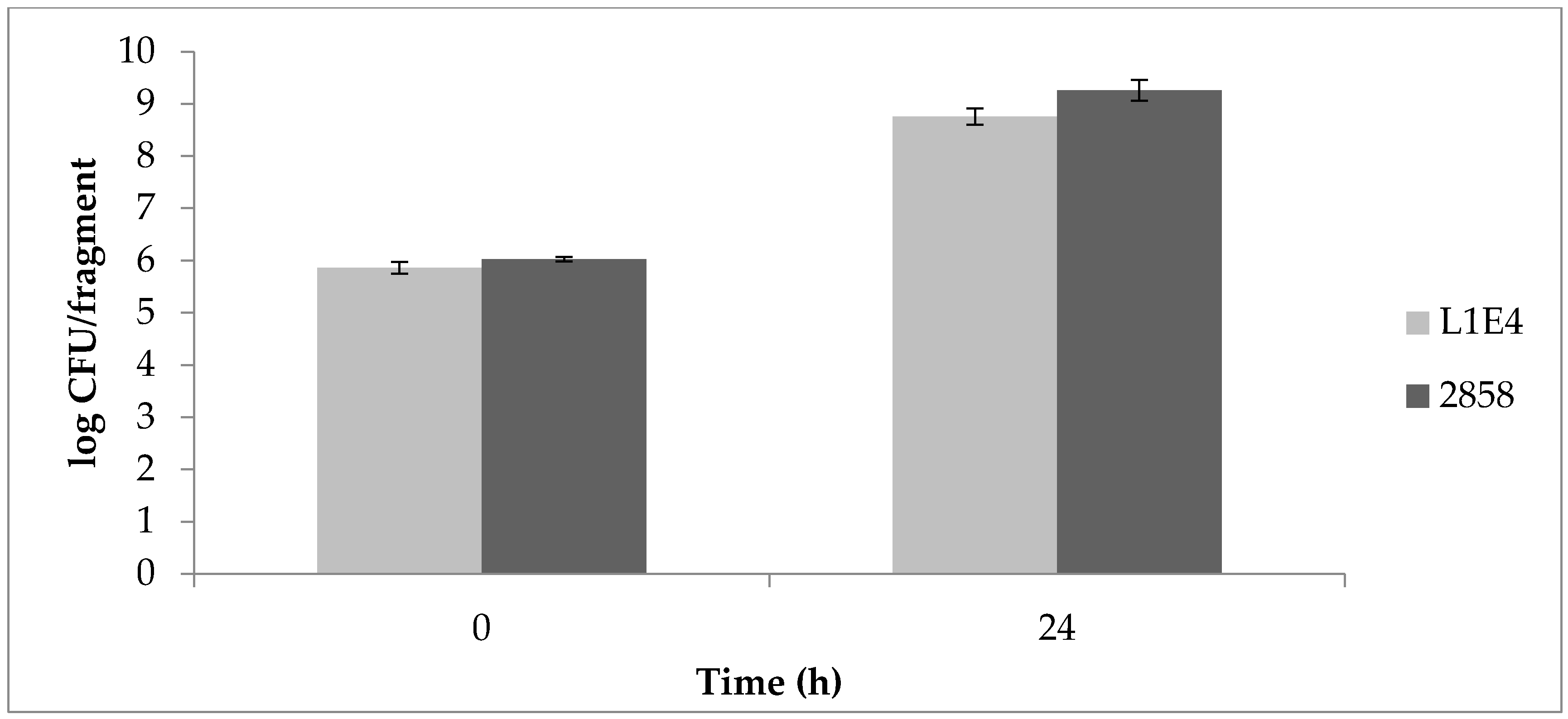
Figure 5.
Competitive fitness of parental strain 2858 vs. non-motile mutant (A) M1A5 and (B) L1E4 on cantaloupe fragments incubated at 25 °C. Time 0 corresponds to 30 min post-inoculation. Ratios were determined by inoculating in individual wells of 96-well plates, which were then plated on media with and without erythromycin, as described in Materials and Methods. Results are from at least two independent trials.
Figure 5.
Competitive fitness of parental strain 2858 vs. non-motile mutant (A) M1A5 and (B) L1E4 on cantaloupe fragments incubated at 25 °C. Time 0 corresponds to 30 min post-inoculation. Ratios were determined by inoculating in individual wells of 96-well plates, which were then plated on media with and without erythromycin, as described in Materials and Methods. Results are from at least two independent trials.
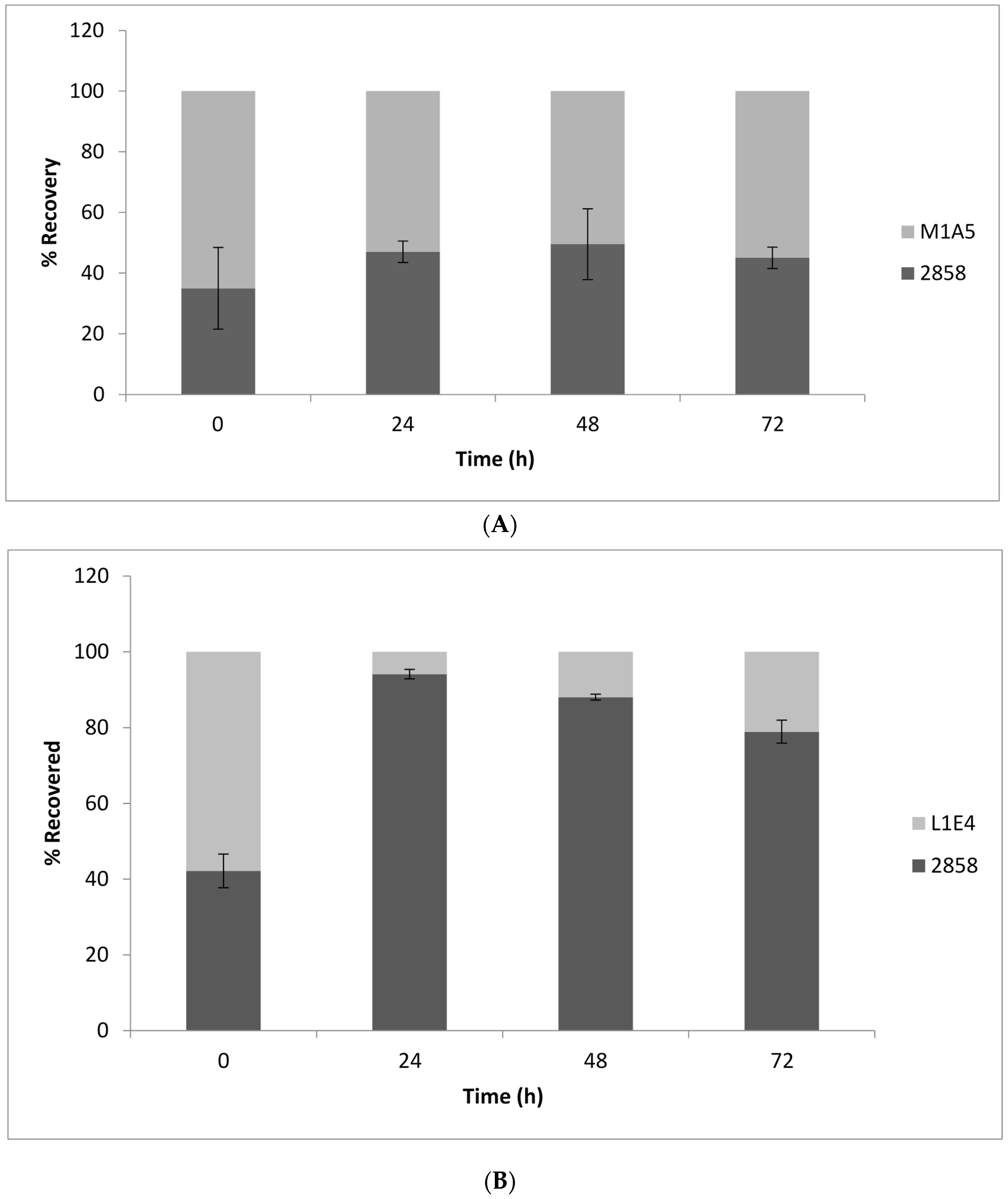
Figure 6.
Competitive fitness of parental strain 2858 vs. helicase mutant L1E4 on cantaloupe fragments incubated at 37 °C over 24 h. Colonies from MOX plates were subcultured in individual wells of 96-well plates which were then plated on media with and without erythromycin, as described in Materials and Methods.
Figure 6.
Competitive fitness of parental strain 2858 vs. helicase mutant L1E4 on cantaloupe fragments incubated at 37 °C over 24 h. Colonies from MOX plates were subcultured in individual wells of 96-well plates which were then plated on media with and without erythromycin, as described in Materials and Methods.
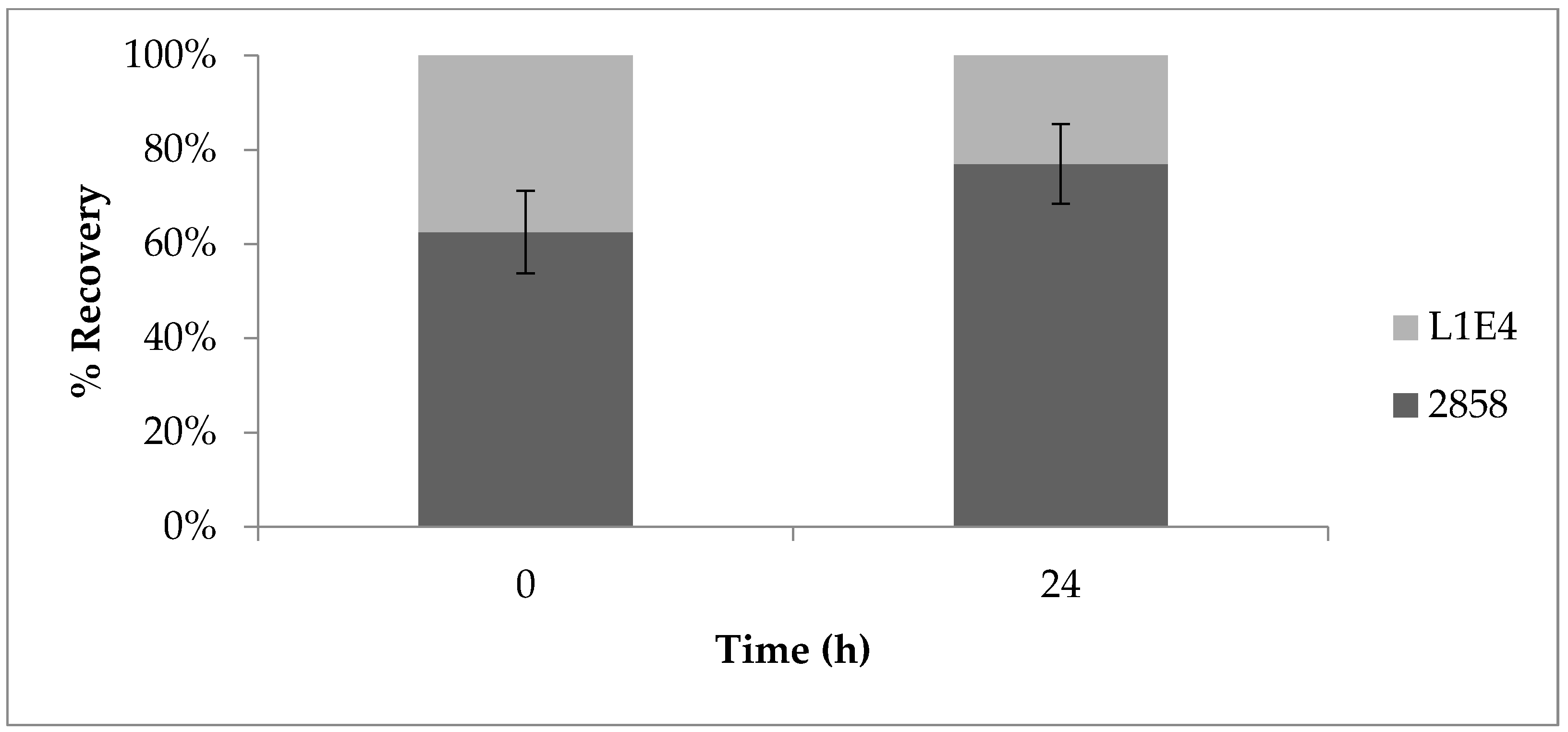
Figure 7.
Restoration of competitive fitness of L1E4 on cantaloupe rind at 25 °C following genetic complementation. Shown are averages of ratios between the parental strain 2858 (gray) vs. L1E4 with empty vector alone (L1E4-E; diagonal lines) and strain 2858 vs. the complemented L1E4 (L1E4-C; white) on cantaloupe fragments incubated at 25 °C over 72 h, with time 0 corresponding to 30 min post-inoculation. Ratios were determined as in legend of Figure 6.
Figure 7.
Restoration of competitive fitness of L1E4 on cantaloupe rind at 25 °C following genetic complementation. Shown are averages of ratios between the parental strain 2858 (gray) vs. L1E4 with empty vector alone (L1E4-E; diagonal lines) and strain 2858 vs. the complemented L1E4 (L1E4-C; white) on cantaloupe fragments incubated at 25 °C over 72 h, with time 0 corresponding to 30 min post-inoculation. Ratios were determined as in legend of Figure 6.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Listeria monocytogenes strains used in this study.
| Strains | Genotype and Features | Phenotype |
|---|---|---|
| 2858 | Wild-type strain from 2011 cantaloupe outbreak | EmS (Erythromycin susceptible) |
| L1E4 | Transposon mutant of 2858 DEAD-box RNA helicase in lmo0866 homolog (nt 981) | Non-motile, cold sensitive, EmR (Erythromycin resistant), reduced hemolysis on blood agar plates |
| M1A5 | Transposon mutant of 2858 in lmo0694 homolog (nt 161) (flagella biosynthesis and chemotaxis gene cluster) | Non-motile, EmR |
| L1E4-C | Complemented L1E4 mutant | Motile, cold growth proficient; EmR, CmR (chloramphenicol resistant) |
| L1E4-E | L1E4 mutant with empty vector (pPL2) | Non-motile, cold-sensitive; EmR, CmR |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Price, R.; Parsons, C.; Kathariou, S. RNA Helicase Mediates Competitive Fitness of Listeria monocytogenes on the Surface of Cantaloupe. Horticulturae 2018, 4, 40. https://doi.org/10.3390/horticulturae4040040
AMA Style
Price R, Parsons C, Kathariou S. RNA Helicase Mediates Competitive Fitness of Listeria monocytogenes on the Surface of Cantaloupe. Horticulturae. 2018; 4(4):40. https://doi.org/10.3390/horticulturae4040040
Chicago/Turabian StylePrice, Robert, Cameron Parsons, and Sophia Kathariou. 2018. "RNA Helicase Mediates Competitive Fitness of Listeria monocytogenes on the Surface of Cantaloupe" Horticulturae 4, no. 4: 40. https://doi.org/10.3390/horticulturae4040040
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.