Abstract
During an investigation of the macrofungal flora in the Huanglong Mountains of the Loess Plateau, northwest China, a unique gomphoid fungus was discovered and collected. After morphological identification and molecular phylogenetic analyses, a new genus named Luteodorsum and its type species, L. huanglongense, were proposed. Phylogenetic analyses were conducted using datasets of nuclear ribosomal DNA 28S large subunit (LSU), mitochondrial (mt) adenosine triphosphatase (ATPase) subunit 6 (atp6), and mt small-subunit rDNA (mtSSU). The results confirmed that L. huanglongense forms an independent clade within Gomphales, with full maximum likelihood bootstrap support (MLBS), maximum parsimony bootstrap support (MPBS), and Bayesian posterior probability (BPP). L. huanglongense is characterized by its sandy-brown, orange-brown, or coffee-brown color; clavate to infundibuliform shape; wrinkled and ridged hymenophore; ellipsoid to obovoid warted basidiospores; cylindrical to clavate flexuous pleurocystidia; and crystal basal mycelium. Overall, this study contributes to the growing body of knowledge on the diversity and evolution of Gomphales and provides valuable insights into the unique fungal flora found in the Huanglong Mountains.
1. Introduction
The order Gomphales Jülich (Agaricomycetes, Basidiomycota) is regarded as a monophyletic group that is closely related to the Geastrales, Phallales, Gloeophyllales, and Hysterangiales orders [1,2,3]. It comprises over 410 species within 3 families, namely Clavariadelphaceae Corner, Lentariaceae Jülich, and Gomphaceae Donk [3,4]. These species are widely distributed worldwide, particularly in the northern hemisphere, and are mostly mycorrhizal or saprotrophic, playing important roles in fungal diversity and forest ecology [2,3,5].
However, the three families of Gomphales exhibit significant macromorphological differentiation. Clavariadelphaceae are typified by club-shaped (clavaroid) basidiomes, such as Clavariadelphus Donk, or stalked basidiomes with teeth underneath the cap, such as the genus Beenakia D.A. Reid [6,7,8]. Lentariaceae are characterized by stalked clavarioid basidiomes, such as Lentaria Corner, or resupinate-hydnoid basidiomes, such as Hydnocristella R.H. Petersen and Kavinia Pilát [9,10,11]. Gomphaceae, which encompass 15 genera, exhibit the most differentiated morphologies among Gomphales, with hypogeous or epigeous; solitary or gregarious; infundibuliform; coralliform; and clavate or irregularly branched forms [3,4,12]. For instance, Ramaria Fr. ex Bonord. spp. are coral fungi, while Gloeocantharellus Singer and Gomphocantharellus L. Fan, Y.Y. Xu, Zhu L. Yang, and S.P. Jian spp. are gilled mushrooms, and Gautieria Vittad. spp. are false truffles. Furthermore, Gomphus Pers., Phaeoclavulina Brinkmann, and Turbinellus Earle spp. are cantharelloid–gomphoid [1,4].
In addition to ecological, molecular phylogenetic, and macromorphological evidence, microscopic and ultramicroscopic features are of great taxonomic value. For Gomphales, comparative descriptions of the different types of spore ornamentation and hilar appendices have helped taxonomists propose relationships between the species, genus, and family levels over recent decades [4,8,13,14,15,16,17,18]. The special gloeocystidia in the hymenium make Gloeocantharellus a recognizable genus of Gomphaceae [19]. Moreover, variations in the basal mycelium, rhizomorphs, and crystals on the surface are also helpful in distinguishing several Clavariadelphus, Lentariaceae, Gomphocantharellus, and Phaeoclavulina species of Gomphales [4,8,9,10,20].
During a survey of the macrofungi flora in the Huanglong Mountains Crossoptilon mantchuricum National Nature Reserve located in the Loess Plateau of northwest China, basidiomes forming a conspicuous fairy ring on litter in a mixed broadleaf–conifer forest were noticed. Although this species resembled a gomphoid mushroom, it was distinct from any known species. After elaborative morphological observations and phylogenetic analyses, a new genus of Luteodorsum and its type species L. huanglongense sp. nov. were proposed.
2. Materials and Methods
2.1. Morphological Studies
Specimens were collected and photographed from the Huanglong Mountains Crossoptilon mantchuricum National Nature Reserve in Shaanxi Province, China. After being dried, voucher specimens were deposited at Herbarium Mycologicum Academiae Sinicae (HMAS), Institute of Microbiology, Chinese Academy of Sciences, Beijing, China, and the Mycological Herbarium of the Forestry College, Northwest A & F University (HMNWAFU-CF), Shaanxi Province, China. Macroscopic characteristics were recorded from both fresh and dried specimens, and standardized color-code designations matching the color of the description were taken from Color-hex (https://www.color-hex.com/, accessed on 3 May 2023). Microscopic observations followed Xu et al. [4]. Fungal histological sections of dried specimens were mounted in 3% KOH, Congo red, Melzer’s reagent [21], and 0.1% (w/v) Cotton blue in lactic acid and observed under an Olympus CX41RF microscope (Tokyo, Japan). The notation “[n/m/p]” indicates n basidiospores from m basidiomes of p collections. The dimensions of the basidiospores are presented using notation of the form (a–)b–c(–d). The range b–c contains a minimum of 90% of the measured values. Extreme values, i.e., a and d, are presented in parentheses. Lm and Wm indicate the average basidiospore length and width (±standard deviation) for the measured basidiospores, respectively. Q represents the mean length/width ratio of a basidiospore from the side view, and Qm represents the average Q of all specimens ± sample standard deviation. Hand-drawn illustrations of the microscopic features were produced using a digital pen tablet (GAOMON WH850) and Adobe Photoshop 2022 software, as previously described [22]. To observe the ultrastructure, basidiospores and basal mycelium scraped from dried specimens were mounted on a scanning electron microscopy (SEM) stub with doubled-sided carbon tape, coated with platinum film using a Shinkuu MSP-1S ion-sputter coater (Mito, Japan), and examined and photographed with a Hitachi S-4800 SEM (Tokyo, Japan). Qualitative X-ray microanalyses were performed on crystals using an energy-dispersive X-ray spectrometry (EDS) microprobe that was fitted on the same SEM and processed using EDAX Genesis Spectrum v6.29 software.
2.2. DNA Extraction, PCR Amplification, and DNA Sequencing
Small amounts of dried basidiome tissues were collected to extract the total genomic DNA using a rapid fungi genomic DNA isolation kit (Sangon Biotech, Shanghai, China). Polymerase chain reaction (PCR) amplification was performed for mitochondrial (mt) adenosine triphosphatase (ATPase) subunit 6 (atp6) using primers ATP6-1/ATP6-2, mt ribosomal DNA small subunit (mtSSU) using primers MS1/MS2, nuclear ribosomal DNA internal transcribed spacer (ITS) region using primers ITS1/ITS4, and nuclear ribosomal DNA 28S large subunit (LSU) using primers LR0R/LR3 [23,24,25].
PCR was performed in a 20 μL reaction volume comprising 2 μL of the DNA template; 1 μL of each primer (10 μM); 10 μL 2× Taq PCR Master Mix (Cowin Biotech, Taizhou, China); and 6 µL ddH2O. PCR amplification procedures were performed using a GeneAmp PCR TC-96 (Bioer Technology, Hangzhou, China) according to the following conditions: for atp6, an initial denaturation stage at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 43 °C for 45 s, extension at 72 °C for 1 min, and a final extension at 72 °C for 10 min, with the procedure ending at 4 °C; for mtSSU, ITS, and LSU, the annealing temperatures were 45 °C, 57 °C, and 50 °C, separately. PCR products were separated via electrophoresis on a 1% agarose gel in a 1× TAE buffer (Solarbio, Beijing, China) and then sequenced by Sangon Biotech Co., Ltd. (Shanghai, China).
2.3. Phylogenetic Analyses
To determine the phylogenetic position of the new genus and species within Gomphales, phylogenetic analyses were conducted using three independent loci of LSU, atp6, and mtSSU based on the maximum likelihood (ML), maximum parsimony (MP), and Bayesian inference (BI) [1,2,4]. The LSU, atp6, and mtSSU sequences of the reference taxa were aligned using the MUSCLE algorithm and manually modified in MEGA-X, respectively, and then combined in SequenceMatrix 1.8 [26]. Three taxa of Mutinus elegans (Mont.) E. Fisch., Phallus impudicus L., and Pseudocolus fusiformis (E. Fisch.) Lloyd were selected as outgroups, as in previous studies [2,4,27].
The ML analysis was conducted in raxmlGUI 2.0 [28] using a GTRGAMMAI model, with all other parameters set to default. A total of 1000 bootstrap replicates were computed using a rapid bootstrap analysis and search for the best-scoring ML tree. The MP analysis was carried out in PAUP* 4.0a169, and bootstrap values were generated with 1000 replicate searches on all parsimony-informative characteristics using 100 random sequence addition replications [29]. Tree bisection reconnection (TBR) branch-swapping algorithms were employed. Tree length (TL), the consistency index (CI), the retention index (RI), the rescaled consistency index (RC), and the homoplasy index (HI) were also calculated. BI analysis was performed in MrBayes 3.1.2 using a partitioned mixed model with LSU, atp6, and mtSSU sequences defined as three independent partitions [30]. Each gene was modeled separately with different parameters. The best-fitting substitution model for each gene was GTR + I + G according to MrModeltest 2.3 software. Four Markov chain Monte Carlo (MCMC) models were run for an initial 5,000,000 generations under the default settings, and continued with analysis until the average standard deviations of the split frequency (ASDSF) values were lower than 0.01 at the end of the runs. Trees were sampled every 100 generations after burn-in (25% of trees were discarded as the burn-in phase of the analyses, set up well after convergence), and 50% majority-rule consensus trees were constructed. Clades with bootstrap support (MLBS and MPBS) ≥ 70% and a Bayesian posterior probability (BPP) ≥ 0.95 were considered significantly supported [31,32]. Phylogenetic trees were viewed with FigTree v1.4.3 and manually annotated using Adobe Illustrator 2022 software.
3. Results
3.1. Phylogenetic Analyses
A total of 238 sequences were used for phylogenetic analyses, which consisted of 226 reference sequences of 81 related taxa downloaded from GenBank, as used in previous studies [2,4], as well as 12 new sequences (4 for LSU, 4 for atp6, and 4 for mtSSU) generated from voucher specimens collected in 2021 and 2022. Accession numbers for all newly generated sequences were obtained by submitting them to GenBank, and details of the sequences used for phylogenetic analyses are provided in Table 1. Additionally, four new ITS sequences were generated and submitted to GenBank. The atp6 dataset comprised 80 taxa and 722 characteristics, of with 240 were constant, 94 were variable and parsimony-uninformative, and 388 were variable and parsimony-informative. The mtSSU dataset comprised 77 taxa and 628 characteristics, of with 278 were constant, 135 were variable and parsimony-uninformative, and 215 were variable and parsimony-informative. The LSU dataset comprised 81 taxa and 714 characteristics, of with 337 were constant, 107 were variable and parsimony-uninformative, and 270 were variable and parsimony-informative. Furthermore, a combined atp6–mtSSU matrix with 1350 total characteristics and a combined LSU–atp6–mtSSU matrix with 2064 total characteristics were generated in this study.

Table 1.
Taxa used in phylogenetic analyses, along with their GenBank accession numbers for LSU, atp6, and mtSSU sequence data. “—” indicates that the sequence was unavailable in GenBank. Accession numbers for sequences generated in this study are denoted in boldface.
Phylogenetic analyses based on a single gene were analyzed at first (Supplementary Figures S1–S3). The results of the atp6 phylogenetic tree confirmed the paraphyletic status of genus Ramaria and the monophyletic status of 11 other genera except the genus Phaeoclavulina (Supplementary Figure S1). Differently, our result split Phaeoclavulina into only two clades instead of three clades, as Xu et al. described [4]. The results of the mtSSU phylogenetic tree agreed with Xu et al. [4]. Ten genera were confirmed as monophyletic, but the monophyletic status of Gloeocanantharellus and Clavariadelphus were not supported with the existence of more than one branch. Meanwhile, for Ramaria subg. Laeticolora; Ramaria subg. Ramaria; and Ramaria subg, Lentoramaria were not distinguished well, and only Ramaria subg. Echinoramaria formed a relatively independent clade (Supplementary Figure S2). In the LSU phylogenetic tree, only nine genera within Gomphales were confirmed as monophyletic. The genus Gautieria was spilt into two clades, which never occurred in the two previous phylogenetic trees (Supplementary Figure S3). Although the topological structures were inconsistent when analyzing these three individual genes, all phylogenetic trees indicated that our specimens formed a completely stable and independent clade, thus being of a monophyletic status.
When combining and analyzing multiple loci, the four subgenera of the paraphyletic genus Ramaria were clearly distinguished, and all other genera were confirmed as monophyletic, in line with previous studies [2,4]. The tree topologies of atp6–mtSSU and LSU–atp6–mtSSU phylogenetic trees were almost identical (Supplementary Figure S4, Figure 1). The MP analysis of the combined atp6–mtSSU dataset resulted in a highly parsimonious tree with a TL of 3678 steps, a CI of 0.368, an RI of 0.659, a RC of 0.243, and an HI of 0.632. The MP analysis of the combined LSU–atp6–mtSSU dataset resulted in a highly parsimonious tree with a TL of 5596 steps, a CI of 0.355, an RI of 0.611, a RC of 0.217, and an HI of 0.645. The ML, MP, and BI analyses of combined datasets yielded very similar tree topologies with minimal variation in statistical support values, so only the tree inferred from the ML analysis is presented.
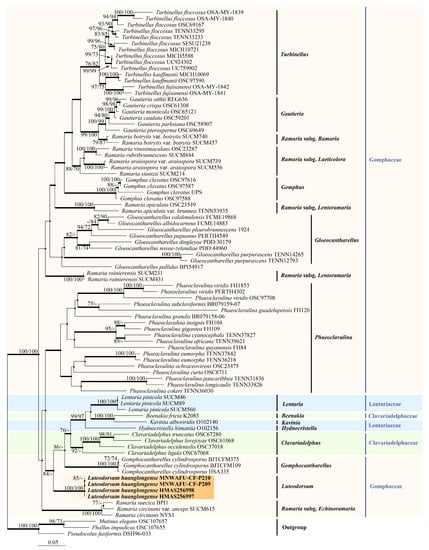
Figure 1.
Phylogenetic tree generated from a maximum likelihood analysis based on combined LSU, atp6, and mtSSU sequences depicting the phylogenetic relationships of Gomphales. Phallus impudicus, Mutinus elegans, and Pseudocolus fusiformis were used as outgroups. The nodes above the branches indicate the maximum likelihood bootstrap support (MLBS) values (≥70%) and maximum parsimony bootstrap support (MPBS) values (≥70%). The branches that presented a Bayesian posterior probability (BPP) ≥ 0.95 are thicker. The novel sequences are highlighted in bold and an orange shade.
The phylogenetic tree revealed that the Gomphales sequences formed a distinct clade presenting significant support values (MLBS/MPBS/BPP = 100%/98%/1 in the atp6–mtSSU tree, and MLBS/MPBS/BPP = 100%/100%/1 in the LSU–atp6–mtSSU tree), with the sequences from northwest China being well-clustered within Gomphales (Supplementary Figure S4, Figure 1). Within the Gomphales phylogenetic tree, the Gomphaceae family contained seven genera, with Ramaria being paraphyletic and the other six genera being monophyletic. The Lentariaceae and Clavariadelphaceae families were paraphyletic and clustered in a clade with a high BPP of 0.99 and a moderate MLBS/MPBS of 68%/69% in the atp6–mtSSU tree (Supplementary Figure S4), as well as high MLBS/BPP values of 76%/0.97 and a moderate MPBS of 64% in the LSU–atp6–mtSSU tree (Figure 1). These results concurred with those of previous studies by Giachini et al. and Xu et al. [2,4]. Four specimens collected from the Loess Plateau of northwest China (HMAS256997, HMAS256998, MNWAFU-CF-P209, and MNWAFU-CF-P210) formed an independent clade with significantly strong support (MLBS = 100%, MPBS = 100%, BPP = 1) in both the atp6–mtSSU and LSU–atp6–mtSSU trees, and were grouped together with the taxa of Gomphocantharellus, Lentariaceae, and Clavariadelphaceae with significant MLBS/MPBS/BPP values of 80%/71%/1 in the atp6–mtSSU tree (Supplementary Figure S4), as well as significant MLBS/BPP values of 84%/1 and a moderate MPBS value of 65% in the LSU–atp6–mtSSU tree (Figure 1), respectively. Based on the phylogenetic tree, we proposed a novel genus, Luteodorsum, which had a closer phylogenetic relationship to Gomphocantharellus and the Ramaria subg. Echinoramaria of Gomphaceae, Lentariaceae, and Clavariadelphaceae than with other genera of Gomphaceae.
3.2. SEM Observation and Qualitative X-ray Microanalysis
Several basidiospores and basal mycelia were scraped from dried specimens and photographed under SEM to observe their ultrastructure. Basidiospores were finely warted, and the basal mycelium was smooth. Interestingly, rosette-like crystals were observed above the hyphal surface of the basal mycelium (Figure 2B). To further understand the characteristics of this fungus, qualitative X-ray microanalysis was conducted using the same SEM to detect the crystal elements.
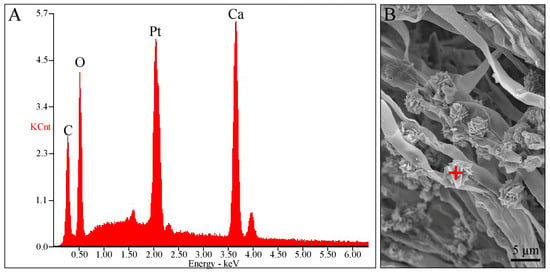
Figure 2.
Qualitative X-ray microanalysis performed via energy-dispersive X-ray spectrometry (EDS). (A) EDS spectrum of crystals on the surface of the basal mycelium, (B) Spot-tested using EDS (red plus).
The results of the EDS spectrum showed four large X-ray peaks of calcium (Ca), platinum (Pt), carbon (C), and oxygen (O) (Figure 2A). The weight percentage and atomic percentage of the rosette-like crystals are listed in Table 2. The Pt was entirely derived from the ion-sputter coater and should be ignored. The crystals were confirmed to be calcium salt crystals, with the presence of only one metallic element. After removing the Pt, the corrected crystal weight comprised 60.90% Ca, 32.41% O, and 6.69% C, and the corrected crystal atoms comprised 37.04% Ca, 49.38% O, and 13.58% C, respectively.
Table 2.
The content of different elements in crystals on the surface of the basal mycelium.
3.3. Taxonomy

Figure 3.
Habitat and basidiomes of Luteodorsum huanglongense (holotype HMAS256997). (A,B) Habitat in the field. (C–F) Young basidiomes. (G) Mature basidiomes.
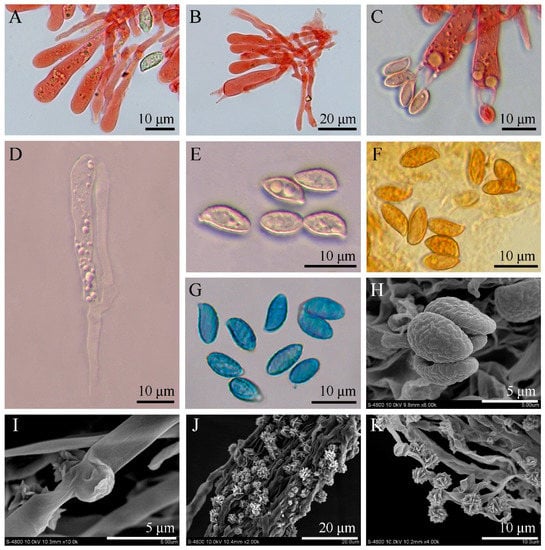
Figure 4.
Microscopic and ultramicroscopic features of Luteodorsum huanglongense. (A) Pleurocystidia, basidioles, and basidiospores in Congo red. (B) Pleurocystidia, basidioles, and basidia in Congo red. (C) Basidia and basidiospores in Congo red. (D) Pleurocystidia and basidioles in KOH. (E) Basidiospores in KOH. (F) Basidiospores in Melzer’s reagent. (G) Basidiospores in Cotton blue. (H) Basidiospores under SEM observation. (I) Clamp connections of basal mycelium under SEM observation. (J,K) Basal mycelium and calcium salt crystals under SEM observation.
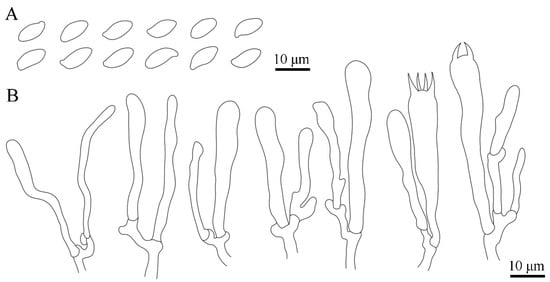
Figure 5.
Microscopic features of Luteodorsum huanglongense. (A) Basidiospores. (B) Pleurocystidia, basidioles, and basidia.
MycoBank 848312
Type species: Luteodorsum huanglongense Z.J. Peng, X.Y. Liu, and Z.D. Yu (described below).
Etymology: Lute- (Lat.), meaning yellow, in reference to the color of the dried hymenium; -dorsum (Lat.), derived from the wrinkled and ridge-like surface of the hymenium; Luteodorsum (Lat.), referring to the color and morphological similarity of the dried hymenium to the famous Loess Plateau of China, which is exactly the typical geomorphology of the city where the type species was collected.
Diagnosis: Luteodorsum differs from the five other cantharelloid–gomphoid genera of Gomphocantharellus, Gloeocantharellus, Gomphus, Phaeoclavulina, and Turbinellus due to its stipitate-pileate basidiomes presenting an almost glabrous to fibrillose pileus without obvious scales; its wrinkled, ridged, salmon to rosy-brown hymenophore; and its ellipsoid to obovoid warted basidiospores. Genetically, Luteodorsum forms a strongly autonomous well-clustered branch of Gomphales based on the LSU, atp6, and mtSSU sequences.
Description: Basidiomes stipitate-pileate, gomphoid, fleshy. Pileus clavate to horse-hoof-like at first, fan-shaped to funnel-shaped at maturity, surface coarse, almost glabrous to fibrillose, with sporadic warts, slightly hygrophanous, margin subundulate. Hymenophore decurrent; wrinkled and ridged; occasionally in irregular patches; light salmon, dark salmon, to rosy-brown; unchanging when exposed. Stipe central or slightly eccentric, cylindrical to slightly tapering downward, solid, with a white basal mycelial cord. Pleurocystidia scattered among and scarcely projecting beyond the basidia, cylindrical to clavate, flexuous, smooth. Hyphae with clamp connections. Basel mycelium smooth, with rosette-like druse crystals. Basidiospores ellipsoid to obovoid, ornamented with warts, light orange to light cinnamon, inamyloid, cyanophilic.
Luteodorsumhuanglongense Z.J. Peng, X.Y. Liu, and Z.D. Yu, sp. nov. (Figure 3, Figure 4 and Figure 5)
MycoBank 848313
Typification: China, Shaanxi Province, Yan’an City, Huanglong County, Huanglong Mountains, Caijiachuan Forest Farm, on litter in mixed broadleaf–conifer forest dominated by Pinus tabuliformis Carrières [33], Quercus mongolica Fisch. ex Turcz. [34], and Betula pendula subsp. mandshurica (Regel) Ashburner and McAll. [35]; elev. 1330 m; 35°49′27″ N, 109°54′49″ E; 1 October 2021; Z.J. Peng, Z.Y. Luo, and Z.D. Yu, HL152 (holotype HMAS256997). GenBank: ITS = OQ801492; LSU = OQ801490; mtSSU = OQ801494; atp6 = OQ790052.
Etymology: huanglongense (Lat.), referring to the type locality in the Huanglong Mountains in Shaanxi Province, China.
Diagnosis: L. huanglongense differs from other species of gomphoid fungi due to the following combination of characteristics: basdiomes unipileate; pileus surface coarse; almost glabrous to fibrillose; slightly hygrophanous; sandy-brown, orange-brown to coffee-brown when fresh; cream, light yellow to fawn when drying out; hymenium occasionally in irregular patches; light salmon, dark salmon to rosy brown when fresh; wheat to earth yellow when drying out; basidiospores (8.5–)8.7–10.7(–12.4) × (4.1–)4.2–5.5(–5.9) μm with Q = 1.6–2.5, Qm = 2.01 (±0.19); basidia 46–67 × 7–9 μm; pleurocystidia 34–49 × 2.7–5 μm, scattered among and scarcely projecting beyond the basidia; basal mycelium with rosette-like calcium salt crystals.
Description: Basidiomes erect; unipileate; fleshy when fresh; fragile when dry, solitary, scattered or in small groups. Pileus 4–12 mm wide; clavate to horse-hoof-like when young, then center depressed; fan-shaped to funnel-shaped (infundibuliform) at maturity; surface coarse; almost glabrous to fibrillose; slightly hygrophanous; sandy-brown (#d29459), orange-brown (#ab7257) to coffee-brown (#975c3c) when fresh; cream (#f9f2d8), light yellow (#f2e1b3) to fawn (#dfbc94) when drying out; margin subundulate; white to light orange (#d8ac93). Hymenium decurrent, with dichotomous, wrinkled, and ridged veins; occasionally in irregular patches; light salmon (#db9a86); dark salmon (#d08a71) to rosy-brown (#a07475) when fresh; wheat (#d8c6ae) to earth yellow (#ad7f44) when drying out. Stipe 27–43 × 5–14 mm, central or slightly eccentric, cylindrical to slightly tapering downward, solid, almost oncolorous with pileus, with white basal mycelial cord. Odor not distinctive, taste not recorded.
Basidiospores (8.5–)8.7–10.7(–12.4) × (4.1–)4.2–5.5(–5.9) μm, Lm × Wm = 9.7 (±0.62) × 4.8 (±0.39) μm, Q = 1.6–2.5, Qm = 2.01 (±0.19) [100/8/4], ellipsoid to obovoid, ornamented with warts, light yellowish to light cinnamon in mass, inamyloid, cyanophilic; apiculus rounded, eccentric. Basidia 46–67 × 7–9 μm, subcylindrical to clavate, sinuous, hyaline with four sterigmata, sterigmata 6.3–9.5 μm long, basal clamp connections present. Pleurocystidia 34–49 × 2.7–5 μm, scattered among and scarcely projecting beyond the basidia, cylindrical to clavate, flexuous, thin-walled, smooth, hyaline, clamped. Hymenophoral trama of hyaline, thin-walled, interwoven hyphae. Pileipellis composed of thin-walled, frequently branched, tightly interwoven hyphae; hyaline to light yellowish; 2–5 μm wide; inflated in the hyphal termini. Stipitipellis of thin-walled, parallel, and interwoven cylindrical hyphae; hyaline to light yellowish; 2–6 μm wide; terminations that are difficult to observe. Pileus and stipe context white to light yellowish white; composed of thin-walled, interwoven, hyaline hyphae; 3–7 μm wide; sometimes with embryo-like structure in the hyphal termini but difficult to observe. Caulocystidia not observed. Basal mycelium smooth, with clamp connections and rosette-like calcium salt crystals on the surface. Clamp connections present in all tissues.
Ecology and habitat: Solitary to scattered on the ground with moss, humus, and debris in mixed broadleaf–conifer forest dominated by Pinus L., Quercus L., and Betula L. [36], sometimes forming obvious fairy ring, elev. 1300–1394 m, currently only known to exist in northwestern China from September to October.
Other specimens examined: China, Shaanxi Province, Yan’an City, Huanglong County, Huanglong Mountains, Caijiachuan Forest Farm, on litter in mixed broadleaf–conifer forest dominated by P. tabuliformis, Q. mongolica, and B. pendula subsp. mandshurica; elev. 1330 m; 35°49′27″ N, 109°54′49″ E; 29 September 2022; B.Y. Ma, J.X. Wei, HL202 (HMAS256998); GenBank: ITS = OQ801493; LSU = OQ801491; mtSSU = OQ801495; atp6 = OQ790053; ibid., HL203 (MNWAFU-CF-P209); GenBank: ITS = OQ929929; LSU = OQ929933; mtSSU = OQ929931; atp6 = OQ924518; ibid., HL204 (MNWAFU-CF-P210); GenBank: ITS = OQ929930; LSU = OQ929934; mtSSU = OQ929932; atp6 = OQ924519.
4. Discussion
Previous molecular phylogenetic analyses for the cantharelloid, clavarioid, gomphoid, and phalloid fungi were conducted based on the multiple loci of LSU, nuclear small subunit rDNA (SSU), atp6, mtSSU, the second largest subunit of RNA polymerase (RPB2), and translation elongation factor subunit 1a (EF-1a) [1,2,4,37,38]. In our study, three loci of LSU, atp6, and mtSSU were chosen for analyzing the phylogenetic relationships of Gomphales, referring to Giachini et al. [2] and Xu et al. [4]. The results of phylogenetic analyses showed some inevitable topological incongruence and unreliable paraphyletic status of some genera when the three genes were analyzed individually (Supplementary Figure S1–S3). While two or three genes were concatenated, the divergences above were well settled. The phylogenetic analyses based on the combined atp6–mtSSU and LSU–atp6–mtSSU datasets indicated that Luteodorsum huanglongense generated an autonomous branch that fit well within Gomphales (Supplementary Figure S4, Figure 1). Thus, Luteodorsum was proposed as a novel genus. Instead of being closely related to morphologically similar species from Gomphus, Phaeoclavulina, and Turbinellus, L. huanglongense was clustered in a clade with Gomphocantharellus, Ramaria subg. Echinoramaria, Clavariadelphaceae, and Lentariaceae. Interestingly, this clade did not exhibit similar macromorphologies due to the phylogenetic affinity, but instead exhibited a great morphological diversity, including coral, gomphoid, cantharelloid, clavarioid, and resupinate hydnoid mushrooms [1,8,9,10,11,39].
Based on its macromorphology, L. huanglongense is easily recognizable as a cantharelloid–gomphoid mushroom in the field due to its typical features of sturdy flesh, fan to funnel shape, and wrinkled outer surfaces (Figure 3). Before our study, five genera of Gomphales were considered as cantharelloid–gomphoid mushrooms, namely Gomphocantharellus, Gloeocantharellus, Gomphus, Phaeoclavulina, and Turbinellus. Among them, Gomphocantharellus and Gloeocantharellus are more similar to chantarelle mushrooms due to their distinct gill-like hymenophore (false lamellae). In particular, Gomphocantharellus has smooth cylindrical basidiospores and a white spore print, while Gloeocantharellus has special gloeocystidia in the hymenium. These above characteristics help to distinguish Gomphocantharellus and Gloeocantharellus from other gomphoid genera due to their common echinulate or verrucose basidiospores, brownish spore print, and the absence of gleoplerous hyphae, including Luteodorsum (Figure 4) [4,19,40,41]. The other three genera, Gomphus, Phaeoclavulina, and Turbinellus, are traditional gomphoid mushrooms typified by wrinkled hymenophore; large and coarse scales on the cap surface; and stipes that are fused together, sharing two or more caps. Among them, Phaeoclavulina contains a minority of gomphoid taxa but a majority of representative coral fungi. In contrast to other gomphoid genera, L. huanglongense has unipileate basidiomes, an almost glabrous to fibrillose pileus without obvious scales, and prominent separate stipes (Figure 3).
Remarkably, the smooth basal mycelium of L. huanglongense was observed to be covered with abundant rosette-like druse crystals (Figure 2B and Figure 4J,K). While some other Gomphales species from Hydnocristella, Clavariadelphus, Lentaria, and Phaeoclavulina have also been reported to produce various crystals on the basal mycelium and rhizomorph hyphae, their composition remains unknown [8,9,10,20,42]. The EDS analyses of the druse crystals from L. huanglongense confirmed the presence of Ca, C, and O (Figure 2, Table 2). The druse crystals from L. huanglongense were found to be similar to spherical aggregates of calcium oxalate (CaOx) formed in mesophyll cells from Abutilon theophrasti Medik. and Acacia robeorum Maslin, instead of the needle-shaped, prismatic, or flaky CaOx found in multiple white-rotting Agaricomycotina fungi, mycorrhiza fungi, and plant pathogen fungi [8,43,44,45,46,47,48]. Some studies have indicated that the production of CaOx crystals by several oxalate-producing fungi is associated with the extraction of Ca2+ from calcium-containing minerals [47,49]. The soil type of the sampling site in a mixed broadleaf–conifer forest dominated by Pinus, Quercus, and Betula was alkaline cinnamon soil [50,51]. In 0–6-, 6–13-, and 13–32-cm cinnamon soil layers, the calcium carbonate (CaCO3) content reached 2.00%, 10.51%, and 17.01%, respectively, indicating a relative abundance of Ca2+ [52]. It has been suggested that an excess of Ca2+ could enhance the CaOx crystal production of fungi [47]; thus, the capability of L. huanglongense to produce CaOx suggested that it may play an important role in soil ecology.
Many gomphoid mushrooms have been reported to be edible and form mycorrhizal associations with trees [41,53,54]. Although L. huanglongense was found around Pinus, Quercus, and Betula and may have an ectomycorrhizal association with these genera, the evidence as to whether it is edible or has any ecological functions is currently insufficient. Overall, the distinctive features of L. huanglongense and its phylogenetic placement in Gomphales make it a unique and interesting addition to the cantharelloid–gomphoid mushroom category. Further research is needed to investigate the edibility and ecological role of L. huanglongense.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9060664/s1, Figure S1: Phylogenetic tree generated from a maximum likelihood analysis based on atp6 sequences, depicting the phylogenetic relationships of Gomphales. Phallus impudicus, and Mutinus elegan were used as outgroups. The nodes above the branches indicate the maximum likelihood bootstrap support (MLBS) values (≥70%) and maximum parsimony bootstrap support (MPBS) values (≥70%). The branches that presented a Bayesian posterior probability (BPP) ≥0.95 are thicker. The novel sequences are highlighted in bold and orange shade; Figure S2: Phylogenetic tree generated from a maximum likelihood analysis based on mtSSU sequences, depicting the phylogenetic relationships of Gomphales. Phallus impudicus, Mutinus elegans, and Pseudocolus fusiformis were used as outgroups. The nodes above the branches indicate the maximum likelihood bootstrap support (MLBS) values (≥70%) and maximum parsimony bootstrap support (MPBS) values (≥70%). The branches that presented a Bayesian posterior probability (BPP) ≥ 0.95 are thicker. The novel sequences are highlighted in bold and orange shade; Figure S3: Phylogenetic tree generated from a maximum likelihood analysis based on LSU sequences, depicting the phylogenetic relationships of Gomphales. Phallus impudicus, Mutinus elegans, and Pseudocolus fusiformis were used as outgroups. The nodes above the branches indicate the maximum likelihood bootstrap support (MLBS) values (≥70%) and maximum parsimony bootstrap support (MPBS) values (≥70%). The branches that presented a Bayesian posterior probability (BPP) ≥ 0.95 are thicker. The novel sequences are highlighted in bold and orange shade; Figure S4: Phylogenetic tree generated from a maximum likelihood analysis based on combined atp6 and mtSSU sequences, depicting the phylogenetic relationships of Gomphales. Phallus impudicus, Mutinus elegans, and Pseudocolus fusiformis were used as outgroups. The nodes above the branches indicate the maximum likelihood bootstrap support (MLBS) values (≥70%) and maximum parsimony bootstrap support (MPBS) values (≥70%). The branches that presented a Bayesian posterior probability (BPP) ≥ 0.95 are thicker. The novel sequences are highlighted in bold and orange shade.
Author Contributions
Conceptualization, Z.Y. and Z.P.; methodology, Z.P., Y.W., Z.L., C.X. and B.W.; software, Z.P., Y.W., B.W. and C.X.; investigation, Z.P., Z.L., B.M., J.W. and Z.Y.; writing—original draft preparation, Z.P. and Y.W.; writing—review and editing, X.L. and Z.Y.; supervision, Z.Y.; project administration, B.M., J.W. and Z.Y.; funding acquisition, Z.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (31670650).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We appreciate the kind support given by L.Y. Zhu, Key Laboratory of Edible Fungal Resources and Utilization (North), Ministry of Agriculture and Rural Affairs, Jilin Agricultural University, for her help in morphological identification. We thank G.Y. Zhang, State Key Laboratory of Crop Stress Biology in Arid Areas, Northwest A & F University, for his assistance in analyzing salt crystal elements. We also express our thanks to T.C. Wang and Y.P. Wang, State-owned Forest Administration Bureau of Huanglong Mountains, for their guidance in the Huanglong Mountains and some photos that were provided (Figure 3A,B).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Hosaka, K.; Bates, S.T.; Beever, R.E.; Castellano, M.A.; Colgan, W., 3rd; Domínguez, L.S.; Nouhra, E.R.; Geml, J.; Giachini, A.J.; Kenney, S.R.; et al. Molecular phylogenetics of the gomphoid-phalloid fungi with an establishment of the new subclass Phallomycetidae and two new orders. Mycologia 2006, 98, 949–959. [Google Scholar] [CrossRef]
- Giachini, A.J.; Hosaka, K.; Nouhra, E.; Spatafora, J.; Trappe, J.M. Phylogenetic relationships of the Gomphales based on nuc-25S-rDNA, mit-12S-rDNA, and mit-atp6-DNA combined sequences. Fungal Biol. 2010, 114, 224–234. [Google Scholar] [CrossRef]
- He, M.-Q.; Zhao, R.-L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef]
- Xu, Y.-Y.; Jian, S.-P.; Mao, N.; Yang, Z.-L.; Fan, L. Gomphocantharellus, a new genus of Gomphales. Mycologia 2022, 114, 748–756. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008; p. 289. ISBN 978-1-84593-933-5. [Google Scholar]
- Corner, E.J.H. Supplement to a monograph of Clavaria and allied genera. Beih. Nova Hedwig. 1970, 33, 1–299. [Google Scholar]
- Methven, A.S. The genus Clavariadelphus in North America. Bibl. Mycol. 1990, 138, 1–192. [Google Scholar]
- Huang, H.-Y.; Zhao, J.; Zhang, P.; Ge, Z.-W.; Li, X.; Tang, L.-P. The genus Clavariadelphus (Clavariadelphaceae, Gomphales) in China. Mycokeys 2020, 70, 89–121. [Google Scholar] [CrossRef]
- Chen, J.-J.; Shen, L.-L.; Cui, B. Morphological characters and molecular data reveal a new species of Hydnocristella (Gomphales, Basidiomycota) from southwestern China. Nova Hedwig. 2015, 101, 139–146. [Google Scholar] [CrossRef]
- Liu, L.-N.; Wu, L.; Chen, Z.-H.; Bau, T.; Zhang, P. The species of Lentaria (Gomphales, Basidiomycota) from China based on morphological and molecular evidence. Mycol. Prog. 2017, 16, 605–612. [Google Scholar] [CrossRef]
- Robledo, G.L.; Urcelay, C. Kavinia chacoserrana sp. nov. (Gomphales, Basidiomycota): A new species from South America based on morphological and molecular data. Mycosphere 2017, 8, 1028–1034. [Google Scholar] [CrossRef]
- González-Ávila, P.A.; Luna-Vega, I.; Ríos, M.V.; Saade, R.L.; Blanco, J.C. Current knowledge and importance of the order Gomphales (Fungi: Basidiomycota) in Mexico. Nova Hedwig. 2013, 97, 55–86. [Google Scholar] [CrossRef]
- Donk, M.A. Four New Families of Hymenomycetes. Persoonia 1961, 1, 405–407. [Google Scholar]
- Donk, M.A. A Conspectus of the Families of Aphyllophorales. Persoonia 1964, 3, 199–324. [Google Scholar]
- Petersen, R.H.; Pearman, W.R. Spore ornamentation in Ramaria as depicted by scanning electron micrographs. Persoonia 1973, 7, 289–292. [Google Scholar]
- Jülich, W.; Star, W. Ultrastructure of basidiospores I. Beenakia. Pers. 1983, 12, 67–74. [Google Scholar]
- Villegas, M.; Cifuentes, J.; Torres, A.E. Sporal characters in Gomphales and their significance for phylogenetics. Fungal Divers. 2005, 18, 157–175. [Google Scholar]
- Maneevun, A.; Dodgson, J.; Sanoamuang, N. Phaeoclavulina and Ramaria (Gomphaceae, Gomphales) from Nam Nao National Park, Thailand. Trop. Nat. Hist. 2012, 12, 147–164. [Google Scholar]
- Deng, C.Y.; Li, T.H. Gloeocantharellus persicinus, a new species from China. Mycotaxon 2008, 106, 449–453. [Google Scholar]
- Mifsud, S. Phaeoclavulina decurrens (Gomphales, Basidiomycetes)—The first record for a coral fungus for the Maltese Islands. Microb. Biosyst. 2019, 4, 55–59. [Google Scholar] [CrossRef]
- Dring, D.M. Techniques for microscopic preparation. In Methods in Microbiology; Booth, C., Ed.; Academic Press: New York, NY, USA, 1971; Volume 4, pp. 95–112. ISBN 978-0-12-521504-6. [Google Scholar] [CrossRef]
- Peng, Z.J.; Yu, A.; Luo, Z.Y.; Liu, X.Y.; Chen, W.F.; Yu, Z.D. Punctularia atropurpurascens (Punctulariaceae, Basidiomycota): A new record to China. Microbiol. China 2021, 48, 4232–4239. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Kretzer, A.M.; Bruns, T.D. Use of atp6 in Fungal Phylogenetics: An Example from the Boletales. Mol. Phylogenet. Evol. 1999, 13, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.-L.; Li, G.-J.; Sánchez-Ramírez, S.; Stata, M.; Yang, Z.-L.; Wu, G.; Dai, Y.-C.; He, S.-H.; Cui, B.-K.; Zhou, J.-L.; et al. A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 2017, 84, 43–74. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or Bootstrap? A Simulation Study Comparing the Performance of Bayesian Markov Chain Monte Carlo Sampling and Bootstrapping in Assessing Phylogenetic Confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef]
- Carrière, E.A. Traité Conifer des Conifers, 2nd ed.; Chez l’Auteur: Paris, France, 1867; p. 510. [Google Scholar]
- Moskovskoe, O.L.P. Bulletin de la Société impériale des naturalists de Moscou. Soc. Imp. Nat. Moscou 1838, 11, 101. [Google Scholar]
- Ashburner, K.; McAllister, H.A. The Genus Betula: A Taxonomic Revision of Birches; Kew Publishing: London, UK, 2013; p. 291. ISBN 9781842461419. [Google Scholar]
- Linné, C.V.; Salvius, L. Species Plantarum; Impensis Laurentii Salvii: Stockholm, Sweden, 1753; pp. 982+994+1000. [Google Scholar]
- Pine, E.M.; Hibbett, D.S.; Donoghue, M.J. Phylogenetic relationships of cantharelloid and clavarioid Homobasidiomycetes based on mitochondrial and nuclear rDNA sequences. Mycologia 1999, 91, 944–963. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Pine, E.M.; Langer, E.; Langer, G.; Donoghue, M.J. Evolution of gilled mushrooms and puffballs inferred from ribosomal DNA sequences. Proc. Natl. Acad. Sci. USA 1997, 94, 12002–12006. [Google Scholar] [CrossRef]
- Nunez, M.P.; Ryvarden, L. A note on the genus Beenakia. Sydowia 1994, 46, 321–328. [Google Scholar]
- Singer, R. New genera of fungi II. Lloydia 1945, 8, 139–144. [Google Scholar]
- Giachini, A.J.; Castellano, M.A. A new taxonomic classification for species in Gomphus sensu lato. Mycotaxon 2011, 115, 183–201. [Google Scholar] [CrossRef]
- González-Ávila, A.; Martínez-González, C.R.; Espinosa, D.; Estrada-Torres, A. Phaeoclavulina liliputiana sp. nov. (Gomphaceae, Gomphales) a new endemic species from Tlaxcala, Mexico. Phytotaxa 2020, 470, 155–164. [Google Scholar] [CrossRef]
- Whitney, K.D.; Arnott, H.J. Calcium Oxalate Crystal Morphology and Development in Agaricus Bisporus. Mycologia 1987, 79, 180–187. [Google Scholar] [CrossRef]
- Webb, M.A. Cell-Mediated Crystallization of Calcium Oxalate in Plants. Plant Cell 1999, 11, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Agerer, R. Fungal relationships and structural identity of their ectomycorrhizae. Mycol. Prog. 2006, 5, 67–107. [Google Scholar] [CrossRef]
- Stodůlková, E.; Sulc, M.; Cisarova, I.; Novak, P.; Kolarik, M.; Flieger, M. Production of (+)-globulol needle crystals on the surface mycelium of Quambalaria cyanescens. Folia Microbiol. 2008, 53, 15–22. [Google Scholar] [CrossRef]
- Guggiari, M.; Bloque, R.; Aragno, M.; Verrecchia, E.; Job, D.; Junier, P. Experimental calcium-oxalate crystal production and dissolution by selected wood-rot fungi. Int. Biodeterior. Biodegrad. 2011, 65, 803–809. [Google Scholar] [CrossRef]
- He, H.; Bleby, T.M.; Veneklaas, E.J.; Lambers, H.; Kuo, J. Morphologies and elemental compositions of calcium crystals in phyllodes and branchlets of Acacia robeorum (Leguminosae: Mimosoideae). Ann. Bot. 2012, 109, 887–896. [Google Scholar] [CrossRef]
- Schilling, J.S.; Jellison, J. Extraction and translocation of calcium from gypsum during wood biodegradation by oxalate-producing fungi. Int. Biodeter. Biodegr. 2007, 60, 8–15. [Google Scholar] [CrossRef]
- Wang, J.B.; Yang, P. Analysis of Physical and Chemical Properties of Forest Soil in Huanglongshan Crossptilon Nature Reserve in Hancheng, Shaanxi Province. For. Resour. Manag. 2014, 105–109. (In Chinese) [Google Scholar] [CrossRef]
- Wang, Y.; Dong, L.; Zhang, M.; Cui, Y.; Bai, X.; Song, B.; Zhang, J.; Yu, X. Dynamic microbial community composition, co-occurrence pattern and assembly in rhizosphere and bulk soils along a coniferous plantation chronosequence. Catena 2023, 223, 106914. [Google Scholar] [CrossRef]
- Jia, H.Y. The physical and chemical character of forest soil in loess regions. Sci. Silvae Sin. 1990, 26, 74–78. (In Chinese) [Google Scholar]
- Lamus, V.; Franco, S.; Montoya, L.; Endara, A.R.; Caballero, L.A.; Bandala, V.M. Mycorrhizal synthesis of the edible mushroom Turbinellus floccosus with Abies religiosa from central Mexico. Mycoscience 2015, 56, 622–626. [Google Scholar] [CrossRef]
- Liu, J.-W.; Luangharn, T.; Wan, S.-P.; Wang, R.; Yu, F.-Q. A new edible species of Gomphus (Gomphaceae) from southwestern China. Mycoscience 2022, 63, 293–297. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).