Role of Virulence Determinants in Candida albicans’ Resistance to Novel 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone


1
National Institute of Public Health-National Institute of Hygiene, Chocimska 24, Warsaw 00-791, Poland
2
Faculty of Chemistry, Warsaw University of Technology, Noakowskiego 3, Warsaw 00-664, Poland
*
Author to whom correspondence should be addressed.
J. Fungi 2017, 3(3), 32; https://doi.org/10.3390/jof3030032
Submission received: 6 April 2017
/
Revised: 17 June 2017
/
Accepted: 20 June 2017
/
Published: 24 June 2017
Abstract
:We investigated the role of KEX2, SAP4-6, EFG1, and CPH1 in the virulence of Candida under a novel compound 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone (Compound 4). We examined whether the exposure of C. albicans cells to Compound 4, non-cytotoxic to mammalian cells, reduces their adhesion to the human epithelium. We next assessed whether the exposure of C. albicans cells to Compound 4 modulates the anti-inflammatory response (IL-10) and induces human macrophages to respond to the Candida cells. There was a marked reduction in the growth of the sap4Δsap5Δsap6Δ mutant cells when incubated with Compound 4. Under Compound 4 (minimal fungicidal concentration MFC = 0.5–16 µg/mL): (1) wild type strain SC5314 showed a resistant phenotype with down-regulation of the KEX2 expression; (2) the following mutants of C. albicans: sap4Δ, sap5Δ, sap6Δ, and cph1Δ displayed decreased susceptibility with the paradoxical effect and up-regulation of the KEX2 expression compared to SC5314; (3) the immune recognition of C. albicans by macrophages and (4) the stimulation of IL-10 were not blocked ex vivo. The effect of deleting KEX2 in C. albicans had a minor impact on the direct activation of Compound 4’s antifungal activity. The adhesion of kex2Δ is lower than that of the wild parental strain SC5314, and tends to decrease if grown in the presence of a sub-endpoint concentration of Compound 4. Our results provide evidence that SAP4–6 play a role as regulators of the anti-Candida resistance to Compound 4. Compound 4 constitutes a suitable core to be further exploited for lead optimization to develop potent antimycotics.
1. Introduction
The emergence of antifungal resistance has driven the field to seek novel antifungal targets that could lead to the development of effective drugs with a new mode of action. Our recent findings [1,2] focused on the antifungal sulfones’ activity, dealing with multiple aspects of the virulence attributes and immune system evasion mechanisms employed by C. albicans. The studies [1,2] included diverse regulators of drug resistance in C. albicans such as: (1) lytic aspartic proteases (Saps), involved in the resistance of C. albicans to azoles (efflux pumps [3]); (2) subtilisin-type serine convertase Kex2 playing a role in the cell wall formation, interfering with Saps in the cell wall remodeling mechanisms, and responding to antifungal drugs [3,4,5]; (3) transcriptional regulators controlling hyphae formation, given that yeast-hyphae implicates morphological alterations (cell wall remodeling [5]); (4) development of biofilms overcoming environmental stress such as drug exposure and immune system attack (metabolic adaptation) [6]. These attributes include robust responses to local environmental stress exerted by sulfones upon C. albicans, the inactivation of which attenuates virulence.
We previously showed [1] that the primary sulfones’ action mode relies on the modulation of programmed cell death. Sulfones specifically affected the fungal cell membrane without injury in mammalian cells [1], indicating that their less evolutionary conserved target is presented in fungi (focused on the yeast-specific regulators of programmed cell death). It was described [7] that apoptosis requires active proteins’ synthesis. Our most recent studies [1,8] indicated that sulfones are responsible for the induction of KEX2, which can be associated with apoptosis and related to increased levels of pro-apoptotic proteins playing a role in the DNA repair check point. The proteinase Kex2 is a regulatory endoproteinase located in the trans-Golgi network, with multiple regulatory functions in Candida cells. Based on our results [1], it was proven that Kex2 is crucial for Candida to cause tissue damage. With respect to sulfone activity, it is conceivable that when overexpressed, KEX2 has an activity sufficient to sustain viability and hyphal growth of Candida. Moreover, we showed that the phenotype with attenuated KEX2 (abolished SAP processing) exhibited weak adhesion regardless of the influence of 4-chloro-3-nitrophenyldifluoroiodomethyl sulfone [1,8]. Therefore, more studies are required to identify KEX2’s biological functions in the sulfone resistance mechanisms displayed by C. albicans.
Transcriptional factors CPH1 and EFG1 participate in different biological processes (fungal cell wall organization, pathogenesis, and regulation of biofilm formation). Since these regulators are unique for C. albicans, they have been considered as potential antifungal drug targets [9]. As it was shown that EFG1 is also involved in antifungal resistance [10,11], and deletion of EFG1 increased susceptibility of C. albicans to halogenated methyl sulfones, here we investigated whether it may produce a similar effect against 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone (Compound 4). In light of the above, we applied these approaches to study the effect of mutation in the genes of serine protease KEX2 and aspartic proteases (SAPs) on the Candida albicans resistance mechanisms against Compound 4.
On the basis of our findings [2] and in continuation of our effort to develop an antifungal target of Compound 4, we report herein on the in vitro susceptibility of the C. albicans wild type reference strain of SC5314 and mutant strains (sap4Δsap5Δsap6Δ, efg1Δ, cph1Δ, and kex2Δ) to Compound 4. Additionally, screening against the C. glabrata wild type strain and mutants was included. We sought to determine whether the virulence factors (KEX2, SAP4-6, CPH1, and EFG1) could be associated with the paradoxical growth PG effect under sulfone influence. We tested the Candida albicans 90028 yeast cell growth inhibition under Compound 4 in time intervals. In addition, we also investigated the influence of Compound 4 on the KEX2 transcription in the C. albicans sessile cells growing on the human epithelium layer (Caco-2). Under Compound 4 the mutants were tested for the KEX2 role in the anti-sulfone response, including the adhesion to the epithelium and anti-inflammatory IL-10 stimulation. We examined whether the exposure of C. albicans cells to Compound 4 modulates the response of human macrophages against the Candida cells.
2. Materials and Methods
2.1. Synthesis of 2-Bromo-2-Chloro-2-[(4-Chlorophenyl)Sulfonyl]-1-Phenylethanone (Compound 4)
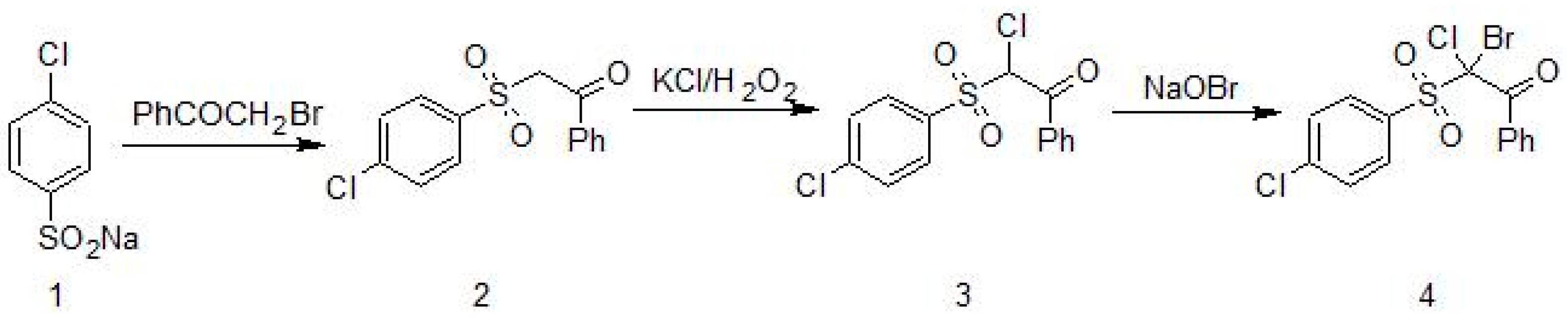
As is shown in Figure 1, 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone (Compound 4) was synthesized starting from sodium 4-chlorobenzene sulfinate (1) which reacted with 2-bromo-1-phenylethanone to obtain 2-(4-chlorophenylsulfonyl)-1-phenylethanone; (2) was transformed into 2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone; (3) by the reaction with potassium halide in the presence of hydrogen peroxide and acetic acid. The final product; (4) was obtained by treatment of β-chloro (3) with sodium hypobromite [2].
2.2. Strains, Media, and Growth Conditions
The Candida albicans wild type reference strains, the mutants of C. albicans and C. glabrata, and the Candida clinical isolates used in the study are listed in Table 1. The decreased azole susceptibility C. glabrata strain was included in the study to further assess the antifungal activity of Compound 4. Strains were stored on ceramic beads in a Microbank tube (Prolab Diagnostics, Richmond Hill, ON, Canada) at −70 °C.
Prior to the respective examinations, routine cultures were conducted at 30 °C for 18 h in the Yeast extract-peptone-dextrose medium (YEPD) [19]. RPMI 1640 medium with l-glutamine without sodium bicarbonate (Sigma, Saint Louis, MO, USA) was buffered with 0.165 M morpholinepropanesulfonic acid (Sigma, Saint Louis, MO, USA) to a pH of 7. Stock solutions of Compound 4 and AmB (Sigma, Saint Louis, MO, USA) were prepared in dimethyl sulfoxide (DMSO, Sigma, Saint Louis, MO, USA) and RPMI, respectively, and stored at −20 °C until use.
2.3. Compound 4 Susceptibility Testing
Compound 4 activity against the planktonic cells of Candida was tested by the broth microdilution method M27-A3 [20]. Compound 4 was tested at concentrations that ranged from 0.25 to 16 μg/mL. As described in [2]: (1) compound test wells (PTW) were prepared with stock solution of Compound 4 (1600 µg/mL) dissolved in DMSO (66% in water), then the yeast cell suspension and Compound 4 (final dilution 1:100) were dispensed at a final volume of 250 µL/well into 96-well microplates; (2) growth control wells (GCW) were prepared with planktonic cells at a final density of 2.5 × 102–2.5 × 103 cfu/mL suspended in RPMI 1640 medium containing 0.66% DMSO (v/v); (3) sterility control wells (SCW) were made with Compound 4, RPMI 1640 medium (Gibco®, Waltham, MA, USA), and sterile water replacing inoculum; (4) microtiter plates were incubated at 35 °C without agitation for 48 h. After incubation, the growth of the cells was measured by a microtiter plate reader Spark Control M10 (Tecan Group Ltd., Grödig, Austria). The endpoint was calculated as a 100% reduction in OD405 as compared to the growth in the control wells according to the formula: % of inhibition = 100—(OD405 CTW—OD405 SCW)/(OD405 GCW—OD405 SCW). Additionally, AmB (Sigma-Aldrich, St. Louis, MO, USA) at 0.5 μg/mL (MIC assessed visually, data not shown) against C. albicans SC5314 was tested. As described in [2], the minimal inhibitory concentrations (MICs) of the compound were read visually after 18 and 48 h. Paradoxical growth (PG), i.e., the reduced antifungal effect (<99.9% reduction of the starting inoculum) at concentrations of above clear endpoint (100% cell growth reduction) was evaluated. Tests were performed in three independent experiments.
2.4. Yeast Cell Growth Inhibition under Compound 4 in Time Intervals
We tested the C. albicans 90028 yeast cell growth inhibition under Compound 4 at the following concentrations: 32, 1, 0.0625, 0.0078, and 0.00195 µg/mL in time intervals [21]. AmB was used as a control against 90028. Furthermore, we used the untreated cells as a growth control. Briefly, the Candida cells were inoculated into the YEPD solid medium and incubated overnight at 35 °C. The final inoculum of 0.5 × 102 to 2.5 × 103 cfu/mL Sabouraud dextrose broth (Sigma-Aldrich, St. Louis, MO, USA) was prepared and placed into the microdilution plates that contained different Compound 4 concentrations, as described above. In addition, the growth control wells with DMSO were added. The visual results were further corroborated by using a spectrophotometer measuring the optical density at 405 nm during the period from 0 to 24 h of incubation. The latency period was defined as the time needed to reach the basal OD and start exponential growth [21]. All growth inhibition experiments were conducted in duplicates on different days and were presented as mean absorbance data.
2.5. Mononuclear Cell PBMCs. In Vitro Candida Stimulation
PBMCs were isolated from heparinized whole blood by gradient density centrifugation in Histopaque 1077 (Sigma-Aldrich, St. Louis, MO, USA) according to the manufacturer’s instructions. The quality of purification of PBMC was verified by staining the cell-containing slides with methylene blue and performing microscopic examination. The cells were counted to produce 5 × 105 cells/mL. To differentiate PBMCs into the monocyte-derived macrophages in vitro, fresh PBMCs were distributed in 24-well plates, and were suspended in RPMI 1640 (Dutch modification, Sigma) supplemented with 2 mM l-glutamine, 1 mM sodium pyruvate, and 100 µg/mL gentamicin for the experiment stimulation [22]. The cells were seeded for one and half hours and then the cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum, 20 mM l-glutamine, and 100 µg/mL gentamicin (all from Gibco) at 37 °C and 5% CO2 (medium was changed every day). After 7 days of incubation at 37 °C, the macrophages were collected and seeded into coverslips and cultured for 30 min (in the latter conditions). Then, the macrophages were stimulated with the Candida cells (untreated and pretreated with Compound 4 for 18 h) at the ratio of 1:5 for 3 h at 37 °C in CO2. Experiments were performed in duplicate and repeated at least three times. The study was approved by the Ethics Committee of the National Institute of Public Health-National Institute of Hygiene (Opinion 5/2012, 28.06.2012 and Appendix 18.04.2013) and all donors signed an informed consent form.
2.6. Fluorescence Microscopy
Cells from different co-cultures on coverslips were stained with Acridine orange AO (Roche Diagnostics GmbH, Mannheim, Germany) working solution (0.01%) at low pH, contrasted by crystal violet and were used for examination under fluorescence microscopy (excitation/emission: at 520–650 nm). Briefly, the cells were washed with PBS and stained with AO for 5 min at room temperature. Then, after washing with PBS, contrast staining with crystal violet was performed for 3 min in the dark, and was followed by three times washing with PBS. Acridine Orange (AO) was used to identify engulfed apoptotic cells and visualized under fluorescent microscopy (Axioskop 40 Zeiss, Oberkochen, Germany). Experiments were performed in triplicate.
2.7. Cytokine Analysis
Fresh PBMC (derived as described above) at a density of 5 × 106 cell/mL were pre-incubated with C. albicans (2 × 106/mL) in RPMI 1640 (Dutch modification, Sigma) supplemented with 2 mM L-glutamine, 1 mM sodium pyruvate, and 100 µg/mL gentamicin in 96-well plates for 24 h at 37 °C in 5% CO2 [23]. The monocytes were stimulated with the heat-killed (at 60 °C for 1 h) or live C. albicans cells, and then pre-incubated or not with Compound 4 for 18 h. The supernatants and cells were then separated by centrifugation and the supernatant was stored at −20 °C until cytokine quantification was performed. The levels of human IL-10 were measured by the ELISA sandwich technique (PeproTech EC Ltd., London, UK) and the results were expressed in ng/mL, according to the manufacturer’s instructions. Plate readings were conducted at 450 nm in the Infinite M200 PRO NANOQuant (Tecan Group Ltd., Grödig, Austria). The experiments were repeated at least three times.
2.8. Assay of Candida Adherence to Human Line Caco-2 (ATCC HTB27, LGC, Poland). Inhibition of Adhesion: Microwell-Based Assay
The adherence of C. albicans and C. glabrata to the Caco-2 cell line (ATCC HTB-37TM) under Compound 4 at a concentration of 0.25 µg/mL was performed as described previously [24]. The adherence was calculated according to the formula: % adhesion = 100% × X adherent cells/X control cells [24]. Where: X control cells stand for the number of cfu/mL of PBS; X adherent cells stands for cells showing adhesion after treatment with Compound 4. The adhesion assay was performed in triplicate and repeated at least twice.
2.9. Analysis of Gene Expression
Total RNA was extracted from cells after 18 h growth in YEPD at 30 °C as previously described [25]. Simultaneously, the overnight-grown cells in YEPD were washed with water and then 200 μL of the suspension was added to 1800 µL of RPMI (final density of 1−1.2 × 106 cells/mL) and inoculated onto the Caco-2 monolayer. Incubation was conducted for 18 h at 37 °C until the RNA extraction.
For the cells pre-treated with Compound 4, the blastoconidia (as described above) were suspended (1−1.2 × 106 cells/mL) in the YEPD medium containing 0.25 µg/mL (final concentration) of Compound 4. Then, after 2 h incubation with Compound 4, the cells were washed with water, suspended in 2000 µL YEPD, and incubated for 18 h at 30 °C until the RNA extraction. Simultaneously, pre-incubated cells with 0.25 µg/mL Compound 4 were re-suspended in 2000 µL of RPMI and inoculated onto the Caco-2 monolayer for 18 h at 37 °C until RNA extraction. Prior to further examinations, C. albicans total RNA was stored at −20 °C. First-strand cDNA synthesis was performed using the Enhanced Avian HS RT-PCR kit (Sigma-Aldrich, St. Louis, MO, USA) according to the manufacturer’s instructions. The sequence of the primer set of KEX2 was designed by using Primer3 (Primer-BLAST, NCBI, Table 2). The primer set of ACT1 (Table 2) was used as described previously by Naglik et al. [26].
cDNA was quantified using the FastStart Essential DNA Green Master (Roche Diagnostics GmbH, Mannheim, Germany), according to the manufacturer’s instructions. Each reaction mixture (15 µL) contained FastStartTaq DNA polymerase, reaction buffer, dNTP mix, SYBR Green I dye, 2 µL (250 nM) of each primer, water PCR grade, and 60 ng of template cDNA [26]. For reliable normalization of the KEX2 gene expression data, we used the housekeeping gene ACT1 as the reference gene [26]. Real-time PCR reactions were performed as described previously by Naglik et al. [26]: at 95 °C for 15 min, followed by 45 cycles of 15 s at 94 °C and 1 min at 60 °C with the LightCycler 96 Instrument (Roche Diagnostics GmbH, Mannheim, Germany). The specificity of each primer pair was determined by the presence of a single melting temperature peak. The CT values were provided from RT-PCR instrumentation and were imported into a Microsoft Excel 2010 spreadsheet. The relative quantification was calculated using an equation [27], where ΔCT = Avg. KEX2 CT—Avg. ACT1 CT and ΔΔCT = ΔCT—ΔCT parental strain.
2.10. Statistical Analysis
All of the above experiments were repeated at least in triplicate on different days, with experimental values expressed as mean ± SD. The statistical differences between the control and the test values were determined by means of the Wilcoxon signed-rank matched-pair test. A p value of 0.05 was assumed as the threshold for differences.
3. Results
3.1. In Vitro Anti-Candida Activity of Compound 4
For SC5314, the MIC value and clear endpoint (100% cell growth reduction) were not observed at the range of concentrations presented in Table 3. The screening of antifungal activity revealed phenotypes displaying the persistent growth at the supra-concentrations of the endpoint (Table 3). The incomplete growth inhibition observed at higher concentrations appeared to vary among the species and mutants and it depended on the deleted genes. Against most of the mutants tested, we observed the endpoint of Compound 4 at low concentrations of Compound 4 and its lower activity at higher concentrations (Table 3). The antifungal activity of Compound 4 (clear endpoint) was observed at 0.5 µg/mL against the null mutants cph1Δ and sap6Δ (PG phenotypes). We found that Compound 4 at 1 µg/mL inhibited the mutant cells completely (100% reduction for 5 out of 17 mutants tested). Thus a decrease in CFU count was observed mainly at 1 µg/mL followed by an increase in the number of CFU of the mutants at the higher concentration of 4 µg/mL (only 95–99% reduction, Table 3). The strain with EFG1 reinsertion exhibited the growth inhibition of approximately 100% at 2 µg/mL (PG phenotype). The antifungal activity screening revealed the phenotypes showing persistent growth at the supra-concentrations of the endpoint (6 out of 17 strains tested, Table 3). The exceptions were the mutants as follows: C. albicans sap4Δsap5Δsap6Δ, C. glabrata his3Δ trp1Δ kex2Δ::HIS3, and C. glabrata his3Δ trp1Δ::TRP1 kex2Δ::HIS3, which revealed 100% reduction at 16 µg/mL. Precisely, this agent showed a clear endpoint against C. glabrata his3Δ trp1Δ kex2Δ::HIS3 at 8 µg/mL and two-fold decreased activity (100% reduction at 16 µg/mL) against C. glabrata his3Δ trp1Δ::TRP1 kex2Δ::HIS3 (Table 3). Although the latter C. glabrata mutants showed cell reduction at 16 µg/mL, the corresponding genes (KEX2 and TRP1) reinsertion mutant was resistant at the whole range of concentrations tested. Thus the deletion of the KEX2 gene in the cells of C. glabrata exerted sensitivity to Compound 4 (Table 3). On the contrary, all of the mutants of C. albicans attenuated in genes KEX2 and/or URA3 displayed the resistant phenotype under Compound 4 at the concentrations ranging from 0.5 to 16 µg/mL. Based on the antifungal activity assay, we further tested the influence of Compound 4 on: (1) KEX2 expression; (2) adhesion; and (3) immune response.
3.2. Candida Cell Growth Inhibition under Compound 4
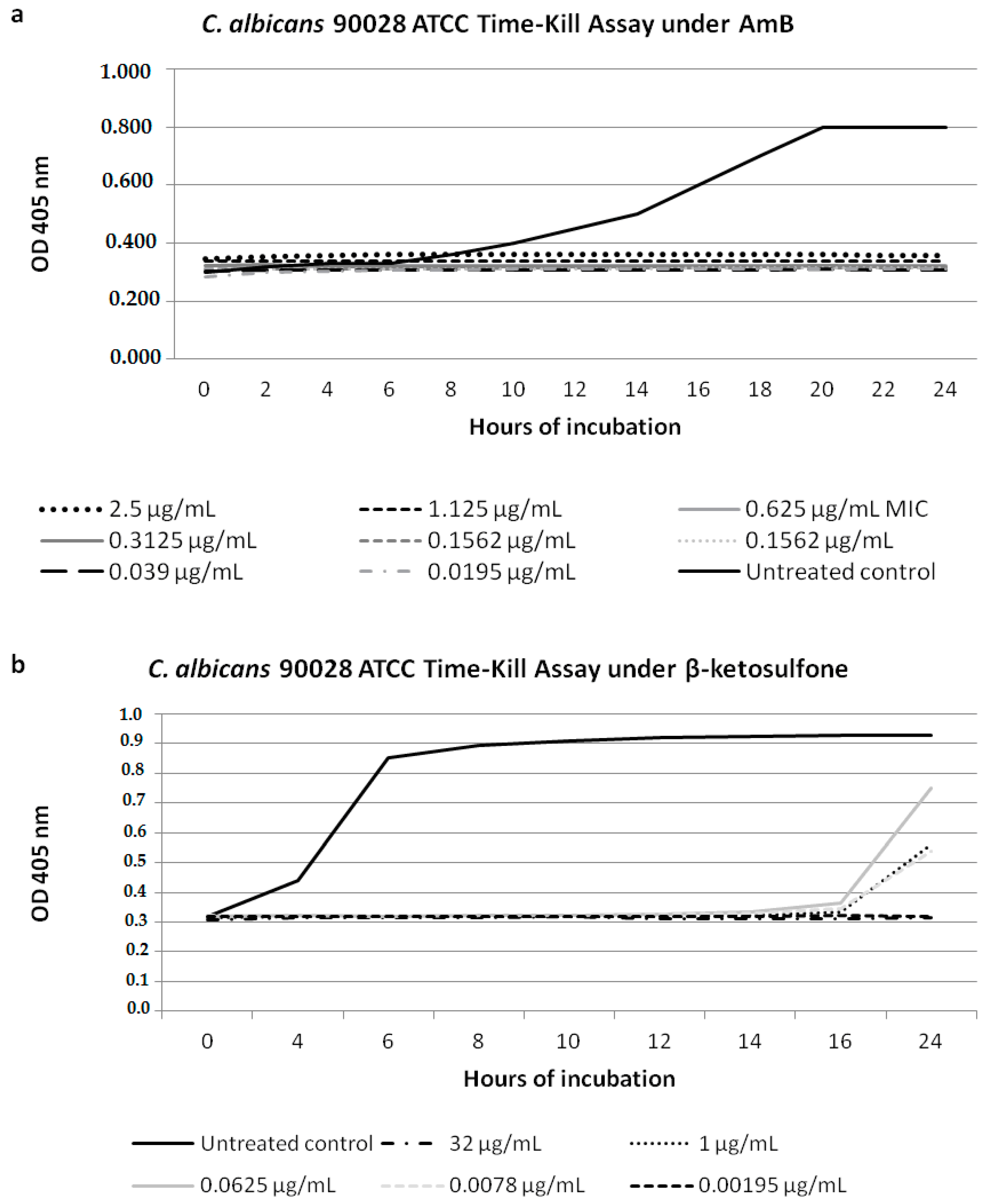
We performed growth inhibition of the strain C. albicans 90028 under Compound 4 and AmB (Figure 2). For Compound 4 at the three concentrations tested (0.0078; 0.0625; 1 µg/mL) against C. albicans 90028, the latency period was 16 h, which was four times longer than that of the growth control (Figure 2b). A dramatic decrease in growth of 90028 was evident for Compound 4 at 32 µg/mL (Figure 2b). The cell growth inhibition for AmB at the concentrations ranging from 2.5 to 0.0195 µg/mL was shown (Figure 2a). We suggested that the difference in the latent period of 90028 depended on the temperature fluctuation going beyond our control during the incubation period.
3.3. Compound 4 Treatment Results in Inhibition of Adhesive Properties of Candida Strains
In the case of SC5314, the adhesion decreased 1.97-fold under Compound 4 at 0.25 µg/mL. The geometric mean adhesion level was 4-fold lower (p ≤ 0.05) for the mutant carrying the homozygous KEX2 mutation (kex2Δ—CNA4) compared to the wild type strain SC5314. On the contrary, the heterozygous URA3 mutation (ura3Δ::URA3—CNA1) decreased adhesion (1.3-fold) of the yeast cells to Caco-2, compared to SC5314 (Table 4). The CNA1 mutant untreated with Compound 4 displayed lightly reduced adhesion vs. SC5314.
In contrast, the mutant reverted with one KEX2 copy (kex2Δ::KEX2—CNA2) but carrying a homozygous URA3 disruption (ura3Δ—CNA2) displayed significantly increased adhesion to Caco-2 (p ≤ 0.05). The mutants CNA1 and CNA3 (kex2Δ/ura3Δ::URA3) showed adhesion under these conditions, with the same level as the wild type strain SC5314 (p > 0.05, Table 4). The Compound 4-treated-cells at 0.25 µg/mL showed significant changes in their adhesion properties, when compared with their non-treated counterparts (p ≤ 0.05, Table 4). Exceptions worth noting were as follows: the C. albicans kex2Δ reintegrated with one URA3 copy (kex2Δ/ura3Δ::URA3—CNA3), and the cell attachment was not significantly inhibited (p > 0.05). The latter was similar for C. glabrata kex2Δ (strain 1006). The adhesion level of the mutants CNA1, CNA2, and CNA4 was inhibited as follows: 1.5-; 2.8-; 3-fold, respectively, at 0.25 µg/mL compared to their non-treated counterparts (Table 4). Additionally, the decreased adhesion was characteristic for C. glabrata 1008 (4.7-fold) and 1010 (2.9-fold). In the case of C. albicans sapΔ, the adhesion increased significantly (p ≤ 0.05) from 1.4- (sap5Δ) to 3.33-fold (sap4Δsap5Δsap6Δ) under 0.25 µg/mL. In contrast, in the sap6Δ mutant this feature was not significantly affected (p > 0.05). Pre-treating the cells with Compound 4 insignificantly affected the adhesion of cph1Δ/efg1Δ, compared with their non-treated counterparts (p > 0.05, Table 4). Interestingly, the remaining morphogenesis mutants displayed a statistically significant increase in the attachment to Caco-2 cells (p ≤ 0.05 compared to their untreated counterparts). In our adhesion assay all of the C. albicans cells displaying adherence to the epithelium were recovered on YEPD generating blastoconidial cells. Assessment of the number of viable cells (CFU) remaining after a 90-min pre-incubation with Compound 4 was carried out as described previously [24]. Ninety-minute incubation of the yeast suspension with Compound 4 did not decrease the viability of all of the tested strains (p > 0.05, data not shown).
3.4. Interaction of C. albicans Cells with Macrophages and Fungal Morphology under Compound 4
Acridine orange fluorescent staining was applied to both the Candida cells treated and untreated with Compound 4 and incubated with macrophages (Figure 3).
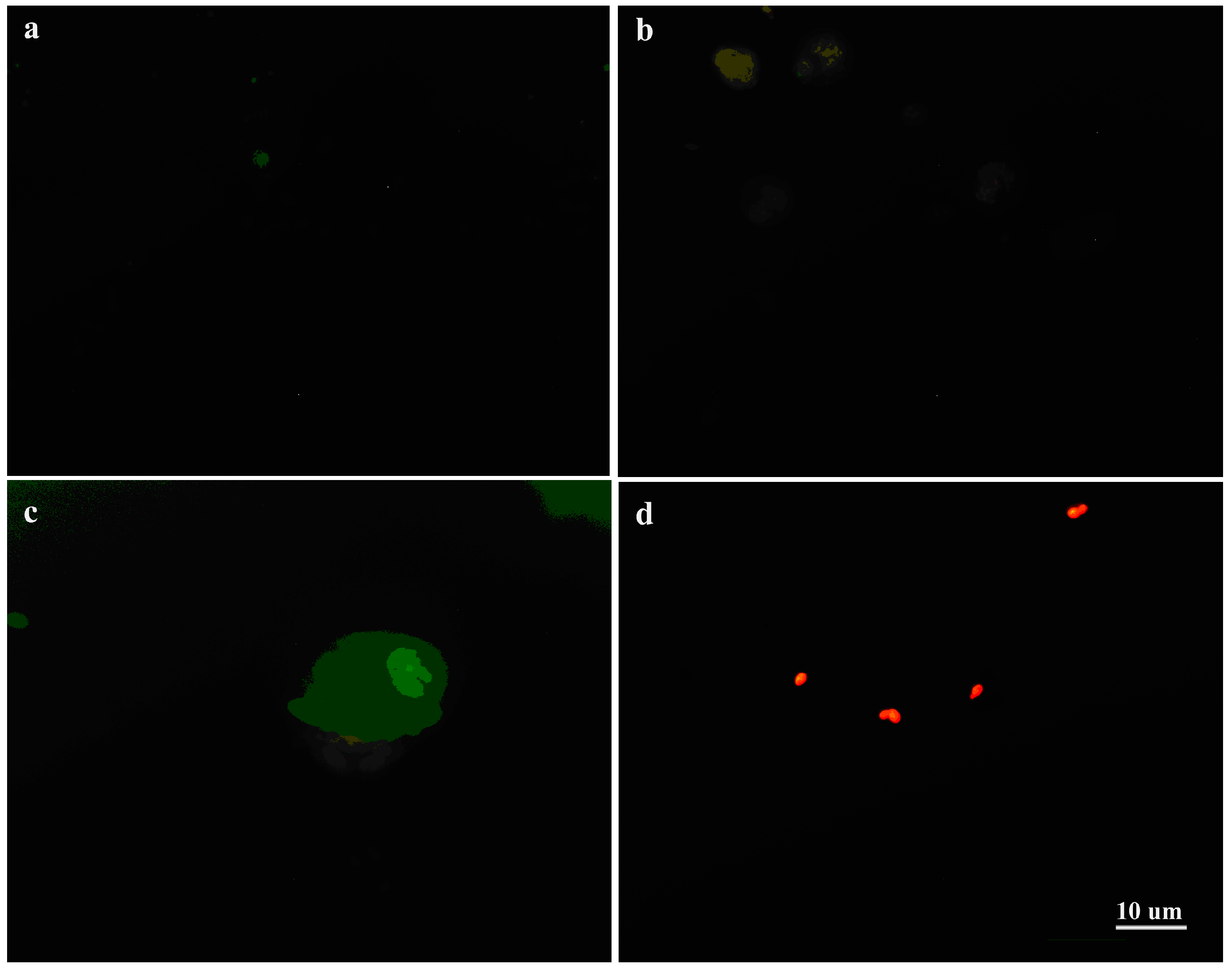
The Candida cells pretreated with Compound 4 were observed outside of the macrophages (Figure 3b,d). After 3 h of co-incubation of the Candida cells with the macrophages (Figure 3a,c), the cells were ingested or not by phagocytes. After pre-treatment with Compound 4, the cells of the wild type SC5314 were observed outside of the phagocytes (Figure 3a,c). Moreover, we depicted the untreated cells of SC5314 not ingested by macrophages (Figure 3a,c), emitting green fluorescence outside of the macrophages. Noticeable, SC5314 pre-treated with Compound 4 showed cells outside of the macrophages (Figure 3d). Based on our results, we suggested that there were no differences in the interaction between macrophages and the Candida cells untreated and pre-treated with Compound 4. Under microscopic examination, we found the C. albicans cells pre-treated with Compound 4 showing acidification, as detected by Acridine orange fluorescence (Figure 3d). Therefore, the cells exposed to Compound 4 prior to interactions with macrophages showed the plasma membrane and lysosomal membrane permeabilization causing the release of Acridine orange from the lysosomal lumen to the cytosol (Figure 3d). The argument is that, in the apoptotic cells, a proton pump increases the concentration of Acridine orange [28]. It appears that the Compound 4-treated cells stained with Acridine orange showed a red-orange cytoplasm, and their nuclei stained yellow. Then, cell death was associated with the decrease in the pH of the cytoplasm (acidification), conceivably by fusion with lysosomes. Finally, this staining procedure identified and selected those cells at the apoptotic stage after the treatment with Compound 4.
3.5. Compound 4 Modulates the KEX2 mRNA Expression
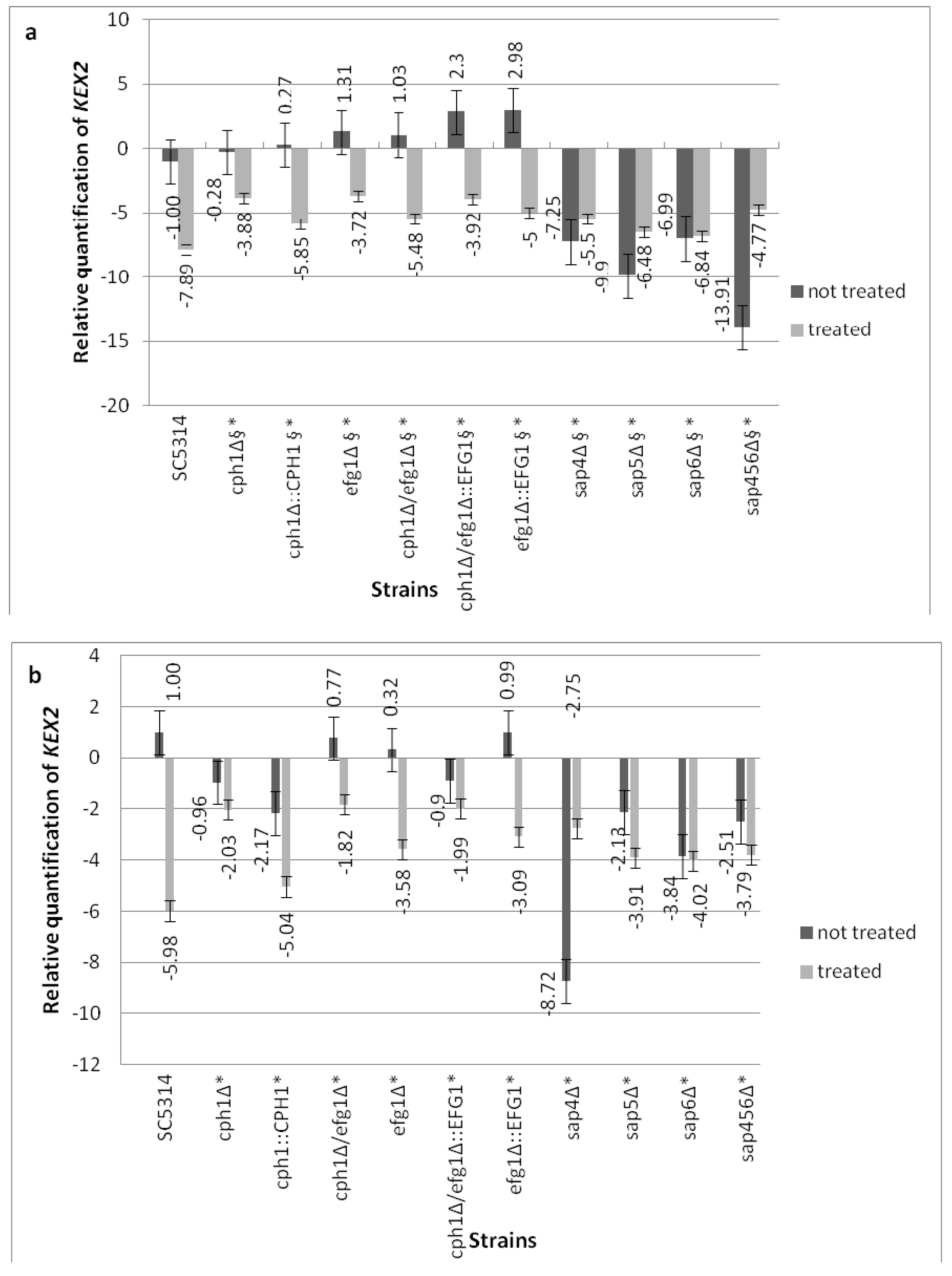
The ability of C. albicans strains to express KEX2 under Compound 4 was assessed with either the yeast cells pre-cultured in YEPD (Figure 4a) as well as in the cells growing on the Caco-2 monolayer (Figure 4b). To identify whether a certain class of genes (Figure 4a,b) was responsible for the KEX2 response under Compound 4 influence, we compared the wild-type strain with either the morphogenetic mutants or the sap4Δsap5Δsap6Δ triple null mutant. Under Compound 4 influence, the mRNA level for KEX2 was significantly affected in all of the mutants compared with the wild-type control cells. The decreased expression of KEX2 was observed for all of the morphogenetic mutants (treated with Compound 4) and significant differences (p < 0.05) appeared between the untreated and treated cells in both the tested media (Figure 4a,b). The KEX2 ΔΔCT raw data were monitored after 18 h growth of C. albicans (untreated and pre-treated with Compound 4) in YEPD (Figure 4a). Firstly, the elevated KEX2 expression by Compound 4 in sap4Δsap5Δsap6Δ during pre-culture in YEPD was noted; secondly, in the case of sap6Δ, almost comparable expression was observed between the treated and untreated cells. The KEX2 expression analysis related to the sap4Δsap5Δsap6Δ treated cells grown on Caco-2 showed decreased levels compared with the untreated counterparts (except for sap4Δ showing three-fold increased expression vs. untreated cells, Figure 4b). Generally for Compound 4, the transcript results of KEX2 in all of the mutants tested were superior to the parental strain SC5314. Eighty percent viability of mammal epithelial cells was assessed for the exposure to Compound 4 at 0.25 µg/mL, as described in [15].
3.6. Interaction of Compound 4 with Interleukine 10
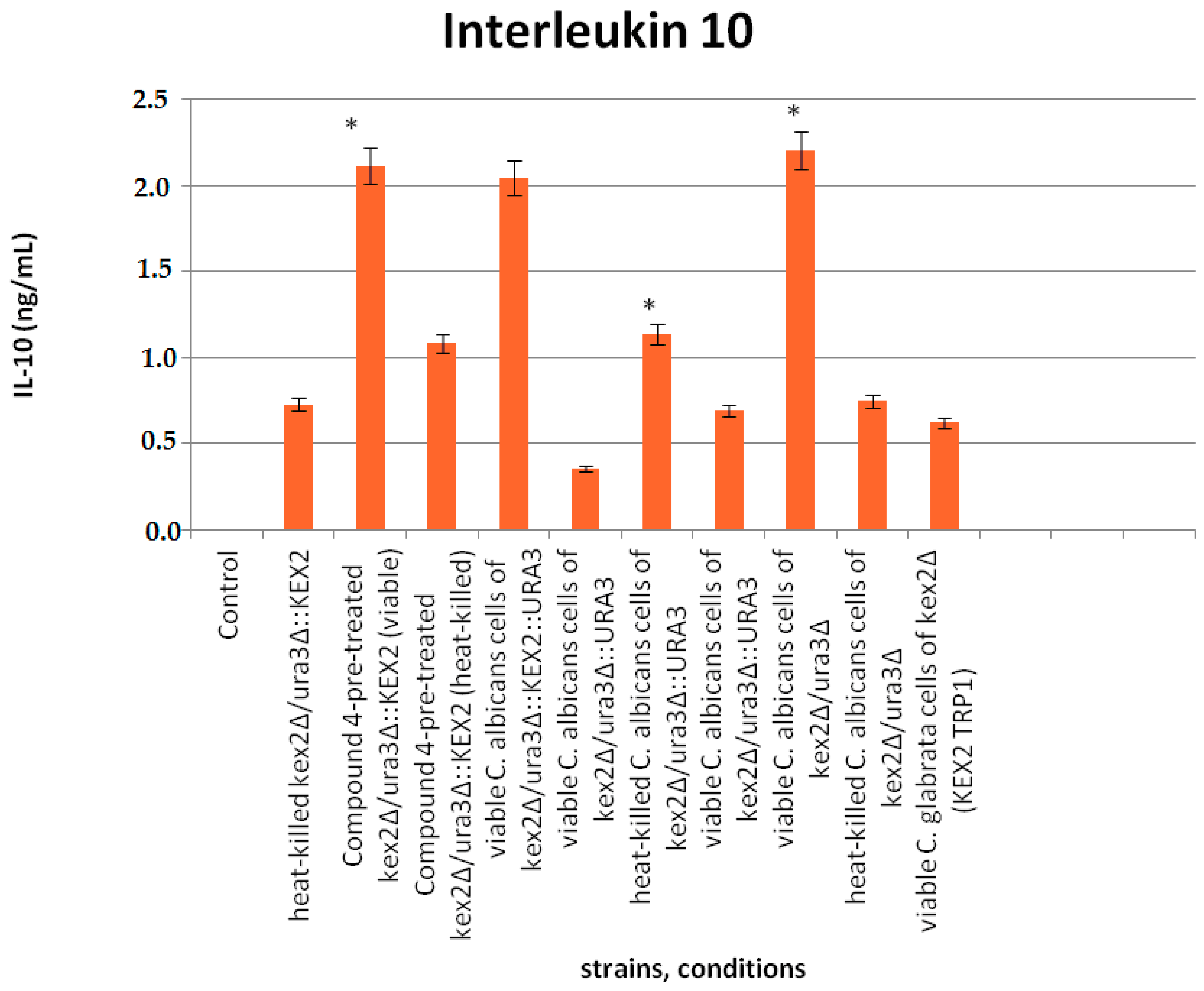
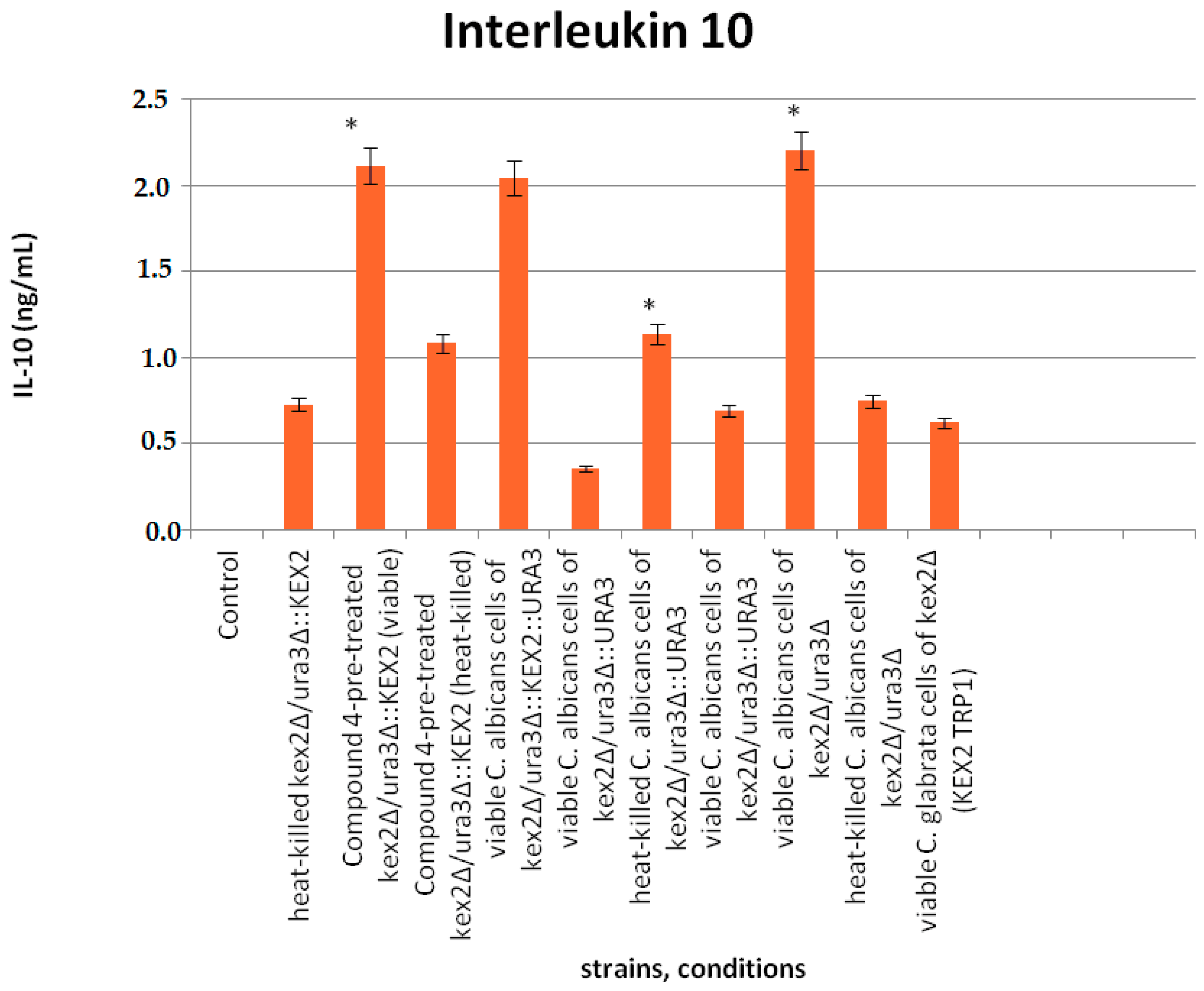
The experimental approach was conducted to evaluate whether Compound 4 affects the Candida mutant cells stimulating macrophages to produce IL-10 (Figure 5). The results indicated that IL-10 was significantly induced by the viable mutants (either untreated or pretreated with Compound 4) compared with their heat-killed counterparts (p ≤ 0.05). No significant differences between the homozygous mutant kex2Δ/ura3Δ (CNA4) and heterozygous mutant reverted with one copy of KEX2 (ura3Δ/kex2Δ::KEX2-strain CNA1) or another one complemented with one copy of both KEX2 and URA3 (kex2Δ::KEX2/ura3Δ::URA3—strain CNA1) either pretreated with Compound 4 and untreated were observed. Consistent with the results above, Compound 4 does not block IL-10 stimulation. The viable cells of the kex2Δ mutant complemented with one copy of URA3 (kex2Δ/ura3Δ::URA3—strain CNA3) down-regulated IL-10 almost 6-fold compared with CNA1 and CNA2, respectively. The C. glabrata kex2Δ cells (heat-killed strain 1006) led to an insignificant IL-10 induction.
4. Discussion
We found that the following genes: SAP4, SAP5, SAP6, and CPH1 are Compound 4 tolerance determinants, as planktonic cells of these mutants were characterized by a decreased Compound 4 sensitivity (PG effect noted at supra-endpoint concentrations) compared with SC5314. We showed that disruption of the morphogenic regulator EFG1 does not enhance the sensitivity of C. albicans cells to Compound 4. As reported previously [11], the sensitivity of efg1Δ to many classes of antimycotics remained unaffected. In our study, this mutant consistently showed a decreased sensitivity to Compound 4, notwithstanding a possible involvement of a decreased content of ergosterol or any passive diffusion. The findings of the first report of Prasad et al. [11] suggested no noticeable difference in the membrane fluidity and passive diffusion in efg1Δ under the tested antimycotics. Following the previous findings [10] that Efg1 is involved in the regulation of the ERG3 expression of the ergosterol biosynthesis pathway in C. albicans, we speculated that the action mode of Compound 4 might be different than targeting ergosterol or its metabolism. Moreover, in line with previous studies [11,22], we assumed that the unaltered (resistant) phenotype of the C. albicans cells under Compound 4 is not dependent on the drug efflux pump, which is affected by the mutation in EFG1. In view of this and of our above-presented findings, the phenomenon of being active in concert with the cell wall components remains a potential target for Compound 4 and its antifungal chemotherapy.
Considering the fact that the overall sensitivity of sap4Δsap5Δsap6Δ was restricted to Compound 4, we speculated that these genes target the cell wall-associated genes, affecting its composition. Since we found that the adhesion of cph1Δ and efg1Δ was superior to that of the parental strain SC5314, we further suggested that the Efg1/Cph1 pathways might not be entirely responsible for adhesin release either under or without the influence of Compound 4. The results obtained are in line with the findings of Carvalho et al. [29], which indicated another transcriptomic approach than EFG1/CPH1, acting under non-steroidal anti-inflammatory drugs. We suggested that Compound 4 interferes with the regulatory mechanisms in the Candida cells attenuated in the following genes: SAP4, SAP5, and SAP6, leading to a compensatory production of the virulence-related genes associated with the adherence, hyphal growth, and other proteases (ALS3, ECE1, SAP1, SAP9,10). Furthermore, Samaranayake et al. [30] reported that different ALS genes can complement each other.
We found that the overexpression of KEX2 influences the susceptibility to Compound 4 in the sap4Δsap5Δsap6Δ mutant or its single counterparts compared with the parental strain, depending on the growth medium and/or conditions. A possible explanation is that the upregulation of KEX2 is required for processing of the virulence-related genes associated with the cell wall remodeling, adherence, and hyphal growth [18]. During the adaptive response to Compound 4 it is possible that KEX2 provides modification to a maturation of the cell wall proteins (i.e., Sap5, Sap6, and Sap 9, 10), involved in the cell surface integrity. It is possible that the KEX2-dependent aspartic protease Sap2 can also be activated in sap4Δ under Compound 4. Previous studies [19,28,31,32] showed an increased activity of Saps in the Candida strains resistant to antifungals. In addition, the Sap2 secretion is induced in the media containing proteins as the sole nitrogen source [33,34]. We are inclined to think that this enzyme can contribute to the Candida growth on the epithelial monolayer where the pH is reduced under Compound 4 (cell acidification).
Our study revealed a marked reduction in the activity of Compound 4 against the C. albicans kex2Δ mutant compared with the C. glabrata kex2Δ mutant (Table 3). The deletion of both KEX2 and URA3 directly activated the antifungal activity of Compound 4 (decreased adhesion, Table 4). However, no significant changes in the cell wall composition or immunogenicity of C. albicans were dependent on URA3 [35]. URA3 and KEX2 are not an important aspect of the Candida biology stimulating PBMCs to produce anti-inflammatory IL-10 under the influence of Compound 4. Herein, we did not test the direct impact of Compound 4 on the activity of monocytes/macrophages. Surprisingly, we found no differences in phagocytosis between the Candida cells treated with Compound 4 (physiologically inactive or dead) and those that were untreated.
Of further note, our findings were consistent with earlier reports [36,37] suggesting that PG represents a compensatory response to a range of cell wall insults. Since the wild type strain displayed a resistant phenotype under Compound 4, the proper regulation of the cell wall integrity pathway is crucial to eliminate this effect. While the biological functions of Sap proteinases are still uncertain, we demonstrated that SAP4, SAP5, and SAP6 can negatively regulate the maintenance of the cell surface integrity under Compound 4. Our findings were consistent with earlier reports suggesting that this effect represents a generalized compensatory response to a range of cell wall insults, rather than specific responses to glucan synthesis.
5. Conclusions
We conclude that the molecular level changes of the signaling pathway can impact the paradoxical effect and overcome its negative effect on the treatment. We propose that the paradoxical effect may be due to a compensatory upregulation of the synthesis of the cell wall components in the Candida cells under the influence of Compound 4. Compound 4 acting as KEX2 inhibitor may prove useful for the treatment of Candida infections.
Acknowledgments
Monika Staniszewska and Małgorzata Bondaryk were supported by funding from the National Science Centre (No. DEC-2011/03/D/NZ7/06198; No. 2014/15/N/NZ6/03710). Zbigniew Ochal was supported by Warsaw University of Technology, Faculty of Chemistry. We thank Hsiu-Jung Lo, Oliver Bader, and Joachim Morschhäuser for providing mutants.
Author Contributions
Monika Staniszewska: substantial contribution to the conception and design of the whole manuscript (know-how), contribution in all the experiments included in the manuscript, acquisition of data, and analysis and interpretation of data, revising it critically for important intellectual content; Małgorzata Bondaryk: participated in the experimental part: antifungal activity of Compound 4, RT-PCR, and adhesion studies; Zbigniew Ochal: contribution in the acquisition of data on the synthesis of beta-ketosulfone, substantial contributions to the conception and design of the sulfone chemical synthesis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Staniszewska, M.; Bondaryk, M.; Kazek, M.; Gliniewicz, A.; Braunsdorf, C.; Schaller, M.; Mora-Montes, HM.; Ochal, Z. Effect of the serine protease KEX2 on Candida albicans virulence under halogenated methyl sulfones. Future Microbiol. 2017, 12, 285–306. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M.; Bondaryk, M.; Wieczorek, M.; Estrada-Mata, E.; Mora-Montes, H.M.; Ochal, Z. Antifungal effect of novel 2-bromo-2-chloro-2(4-chlorophenylsulfonyl)-1-phenylethanone against Candida strains. Front. Microbiol. 2016, 7, 1309. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yu, D.; Gao, S.; Lin, J.; Chen, Z.; Zhao, W. Role of Candida albicans-secreted aspartyl proteinases (Saps) in severe early childhood caries. Int. J. Mol. Sci. 2014, 15, 10766–10779. [Google Scholar] [CrossRef] [PubMed]
- Newport, G.; Agabian, N. KEX2 influences Candida albicans proteinase secretion and hyphal formation. J. Biol. Chem. 1997, 272, 28954–28961. [Google Scholar] [CrossRef] [PubMed]
- Bader, O.; Schaller, M.; Klein, S.; Kukula, J.; Haack, K.; Mühlschlegel, F.; Korting, H.C.; Schäfer, W.; Hube, B. The KEX2 gene Candida glabrata is required for cell surface integrity. Mol. Microbiol. 2001, 41, 1431–1444. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Budge, S.; Kaloriti, D.; Tillmann, A.; Jacobsen, M.D.; Yin, Z.; Ene, J.V.; Bohovych, I.; Sandai, D.; Kastora, S.; et al. Stress adaptation in a pathogenic fungus. J. Exp. Biol. 2014, 217, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Sudbery, I.; Ramsdale, M. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proc. Natl. Acad. Sci. USA 2003, 100, 14327–14332. [Google Scholar] [CrossRef] [PubMed]
- Bondaryk, M.; Grabowska-Jadach, I.; Ochal, Z.; Sygitowicz, G.; Staniszewska, M. Possible role of hydrolytic enzymes (Sap, Kex2) in Candida albicans response to aromatic compounds bearing a sulfone moiety. Chem. Papers 2016, 70, 1336–1350. [Google Scholar] [CrossRef]
- Tripathi, H.S.; Luqman, S.; Meena, A.; Khan, F. Genomic identification of potential targets unique to Candida albicans for the discovery of antifungal agents. Curr. Drug Targets 2014, 15, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.J.; Wang, J.S.; Lin, C.Y.; Chen, C.G.; Hsiao, T.Y.; Hsu, C.T.; Su, C.L.; Fann, M.J.; Ching, Y.T.; Yang, Y.L. Efg1 involved in drug resistance by regulating the expression of ERG3 in Candida albicans. Antimicrob. Agents Chemother. 2005, 49, 1213–1215. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.; Hameed, S.; Manoharlal, R.; Biswas, S.; Mukhopadhyay, C.K.; Goswami, S.K.; Prasad, R. Morphogenic regulator EFG1 affects the drug susceptibilities of pathogenic Candida albicans. FEMS Yeast Res. 2010, 10, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Bale, M.; Buschelman, B.; Lancaster, M.; Espinel-Ingroff, A.; Rex, J.H.; Rinaldi, M.G. Selection of candidate quality control isolates and tentative quality control ranges for in vitro susceptibility testing of yeast isolates by National Committee for Clinical Laboratory Standards proposed standard methods. J. Clin. Microbiol. 1994, 32, 1650–1653. [Google Scholar] [PubMed]
- Gillum, A.M.; Tsai, E.Y.; Kirsch, D.R. Isolation of the Candida albicans gene for orotidine-5′-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 1984, 198, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar] [PubMed]
- Liu, H.; Köhler, J.; Fink, G.R. Suppression of hyphal formation in Candida albicans by mutation of a STE12 homolog. Science 1994, 266, 1723–1726. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.J.; Köhler, J.R.; DiDomenico, B.; Loebenberg, D.; Cacciapuoti, A.; Fink, G.R. Nonfilamentous C. albicans mutants are avirulent. Cell 1997, 90, 939–949. [Google Scholar] [CrossRef]
- Lermann, U.; Morschhäuser, J. Secreted aspartic proteases are not required for invasion of reconstituted human epithelia by Candida albicans. Microbiolology 2008, 154, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Newport, G.; Kuo, A.; Flattery, A.; Gill, C.; Blake, J.J.; Kurtz, M.B.; Abruzzo, G.K.; Agabian, N. Inactivation of Kex2p diminishes the virulence of Candida albicans. J. Biol. Chem. 2003, 278, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M.; Bondaryk, M.; Ochal, Z. Susceptibility of Candida albicans to new synthetic sulfone derivatives. Arch. Pharm. 2015, 348, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. CLSI Document M27-A3, Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, Approved Standard, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Shields, R.K.; Nguyen, M.H.; Du, C.; Press, E.; Cheng, S.; Clancy, C.J. Paradoxical effect of caspofungin against Candida bloodstream isolates is mediated by multiple pathways but eliminated in human serum. Antimicrob. Agents Chemother. 2011, 55, 2641–2647. [Google Scholar] [CrossRef] [PubMed]
- Bink, A.; Govaert, G.; Vandenbosh, D.; Kuchariková, S.; Coenye, T.; Nelis, H.; Van Dijck, P.; Cammue, B.P.; Thevissen, K. Transcription factor Efg1 contributes to the tolerance of Candida albicans biofilms against antifungal agents in vitro and in vivo. J. Med. Microbiol. 2012, 61, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Temple, S.E.; Pham, K.; Glendenning, P.; Philips, M.; Waterer, G.W. Endotoxin induced TNF and IL-10 mRNA production is higher in male than female donors, correlation with elevated expression of TLR4. Cell. Immunol. 2008, 251, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M.; Bondaryk, M.; Ochal, Z. Polish Patent Application PL-P.408765. 4 July 2014. [Google Scholar]
- Amberg, D.; Burke, D.; Strathern, J. (Eds.) Yeast RNA isolations, techniques and protocols. In Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2007; pp. 127–131. [Google Scholar]
- Naglik, J.R.; Moyes, D.; Makwana, J.; Kanzaria, P.; Tsichlaki, E.; Weindl, G.; Tappuni, A.R.; Rogers, C.A.; Woodman, A.J.; Challacombe, S.J.; et al. Quantitative expression of the Candida albicans secreted aspartyl proteinase gene family in human oral and vaginal candidasis. Microbiology 2008, 154, 3266–3280. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time Quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Shukla, P.K. Amphotericin B resistance leads to enhanced proteinase and phospholipase activity and reduced germ tube formation in Candida albicans. Fungal Biol. 2010, 114, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.P.; Gursky, L.C.; Rosa, R.T.; Rymovicz, A.U.; Campelo, P.M.; Grégio, A.M.; Koga-Ito, C.Y.; Samaranayake, L.P.; Rosa, E.A. Non-steroidal anti-inflammatory drugs may modulate the protease activity of Candida albicans. Microb. Pathog. 2010, 49, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Samaranayake, Y.H.; Cheung, B.P.; Yau, J.Y.; Yeung, S.K.; Samaranayake, L.P. Human serum promotes Candida biofilm growth and virulence gene expression on silicone biomaterial. PLoS ONE 2013, 8, e6292. [Google Scholar] [CrossRef] [PubMed]
- Bondaryk, M.; Ochal, Z.; Staniszewska, M. Sulfone derivatives reduce growth, adhesion and aspartic protease SAP2 gene expresion. World J. Microbiol. Biotechnol. 2014, 30, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Ripeau, J.S.; Aumont, F.; Belhumeur, P.; Ostrosky-Zeichner, L.; Rex, J.H.; de Repentigny, L. Effect of the echinocandin caspofungin on expression of Candida albicans secretory aspartyl proteinases and phospholipase in vitro. Antimicrob. Agents Chemother. 2002, 46, 3096–3100. [Google Scholar] [CrossRef] [PubMed]
- Copping, V.M.; Berelle, C.J.; Hube, B.; Gow, N.A.; Brown, A.J.; Odds, F.C. Exposure of Candida albicans to antifungal agents affects expression of SAP2 and SAP9 secreted aspartyl proteinase genes. J. Antimicrob. Chemother. 2005, 55, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, E.; Schneider, A.E.; Sándor, N.; Lermann, U.; Staib, P.; Kremlitzka, M.; Bajtay, Z.; Barz, D.; Erdei, A.; Józsi, M. Secreted aspartic protease 2 of Candida albicans inactivates factor H and the macrophage factor H-receptors CR3 (CD11b/CD18) and CR4 (CD11c/CD18). Immunol. Lett. 2015, 168, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Zavrel, M.; Majer, O.; Kuchler, K.; Rupp, S. Transcription factor Efg1 shows a haploinsufficiency phenotype in modulating the cell wall architecture and immunogenicity of Candida albicans. Eukaryot. Cell 2012, 129, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Rueda, C.; Cuenca-Estrella, M.; Zaragoza, O. Paradoxical growth of Candida albicans in the presence of caspofungin is associated with multiple cell well rearrangements and decreased virulence. Antimicrob. Agents Chemother. 2014, 58, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Juvvadi, P.R.; Muñoz, A.; Lamoth, F.; Soderblom, E.J.; Moseley, M.A.; Read, N.D.; Steinbach, W.J. Calcium mediated induction of paradoxical growth following caspofungin treatment is associated with calcineurin activation and phosphorylation in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2015, 59, 4946–4955. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Synthesis of 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone (4) [2].
Figure 1.
Synthesis of 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone (4) [2].
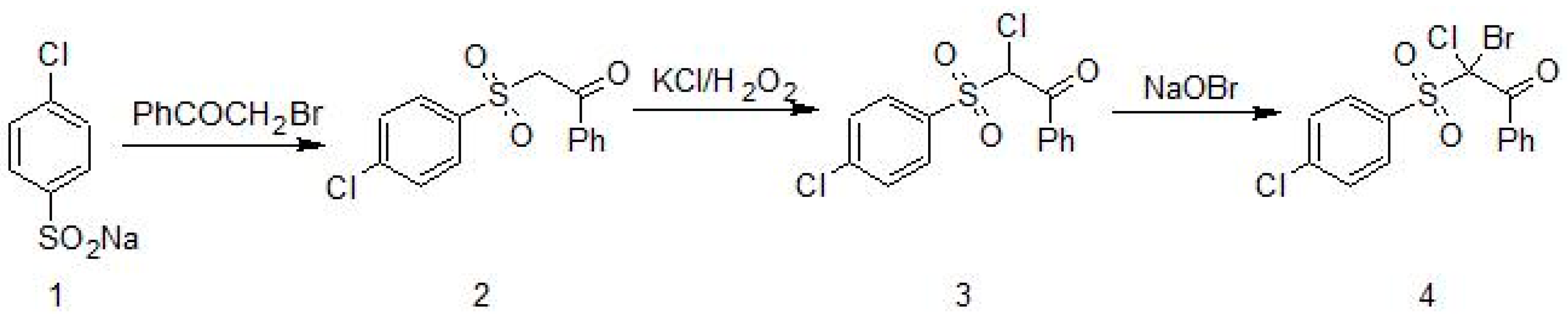
Figure 2.
Characterization of growth of the Candida strains in the presence of Compound 4. The suspensions were prepared with organisms in log phase growth and diluted in Sabouraud Dextrose Broth SDB medium to obtain a final inoculum of 2.5 × 102−103 cfu/mL (OD405 nm). The effect of the antifungal agent of AmB was used as a control growth inhibition assay during 24 h against the reference strain of 90028 (a); (b) Compound 4 showed a very good inhibitory effect against 90028. The antifungal nature of Compound 4 against 90028 was determined in the growth inhibition curve studies. Compound 4 was tested at the highest concentration of 32 µg/mL and at the lowest one of 0.00195 µg/mL.
Figure 2.
Characterization of growth of the Candida strains in the presence of Compound 4. The suspensions were prepared with organisms in log phase growth and diluted in Sabouraud Dextrose Broth SDB medium to obtain a final inoculum of 2.5 × 102−103 cfu/mL (OD405 nm). The effect of the antifungal agent of AmB was used as a control growth inhibition assay during 24 h against the reference strain of 90028 (a); (b) Compound 4 showed a very good inhibitory effect against 90028. The antifungal nature of Compound 4 against 90028 was determined in the growth inhibition curve studies. Compound 4 was tested at the highest concentration of 32 µg/mL and at the lowest one of 0.00195 µg/mL.
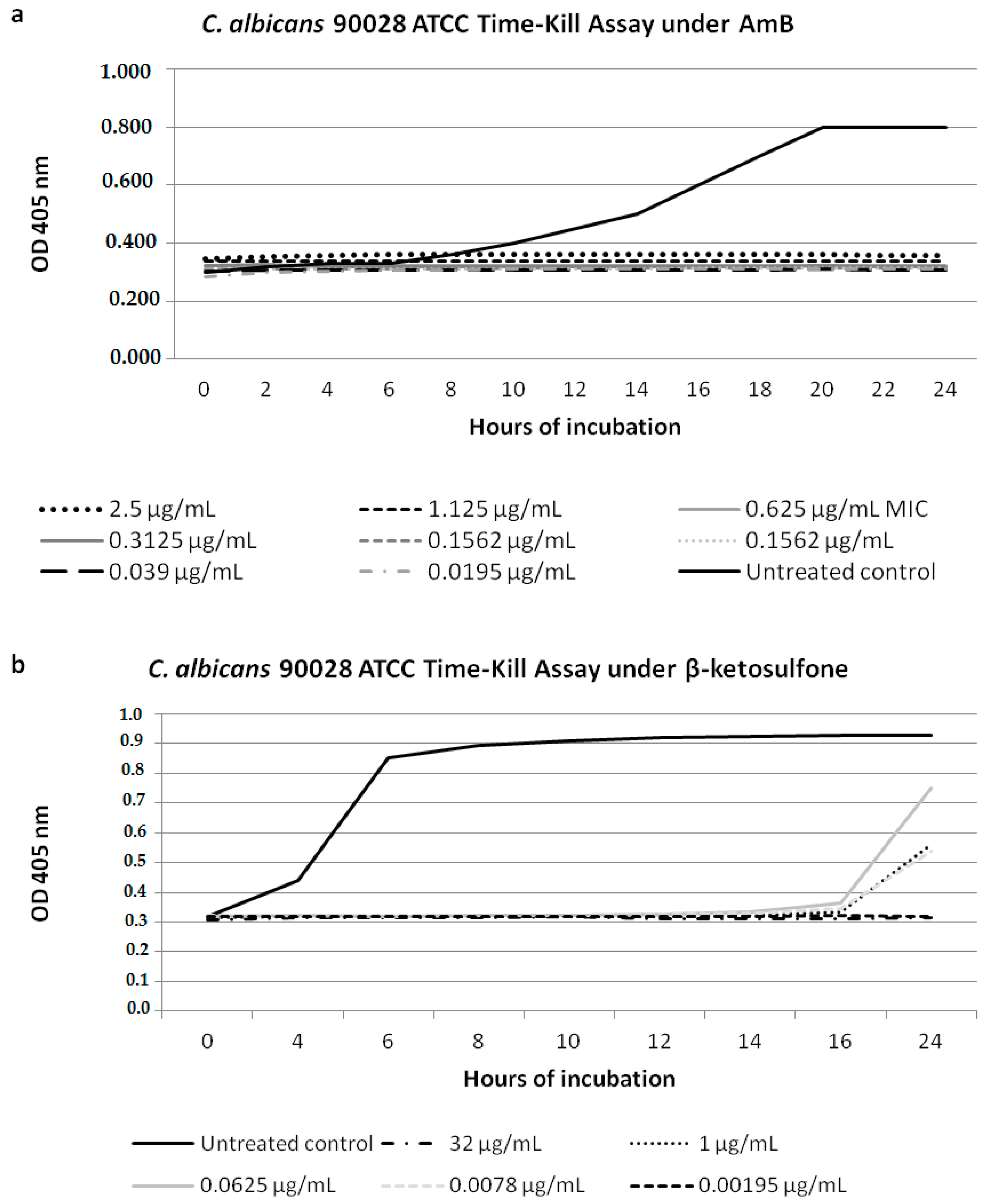
Figure 3.
Candida albicans—Macrophage Interactions. Candida albicans cells were untreated or pre-treated with Compound 4. (a) Control macrophage and C. albicans SC5314 cells (live) untreated with Compound 4 were seen in the macrophage’s vicinity; (b) Macrophages displaying signs of acidification (red fluorescence), suggesting that SC5314 cells treated with Compound 4 were ingested by them. Yeast cells showing acidification outside macrophages were observed (spotting red fluorescence); (c) Macrophages showed acidification (red fluorescence), suggesting that SC5314 cells untreated with Compound 4 were engulfed by it. Live yeast cells attached to macrophages were seen (green fluorescence); (d) SC5314 cells pre-treated with Compound 4 emitting red fluorescence.
Figure 3.
Candida albicans—Macrophage Interactions. Candida albicans cells were untreated or pre-treated with Compound 4. (a) Control macrophage and C. albicans SC5314 cells (live) untreated with Compound 4 were seen in the macrophage’s vicinity; (b) Macrophages displaying signs of acidification (red fluorescence), suggesting that SC5314 cells treated with Compound 4 were ingested by them. Yeast cells showing acidification outside macrophages were observed (spotting red fluorescence); (c) Macrophages showed acidification (red fluorescence), suggesting that SC5314 cells untreated with Compound 4 were engulfed by it. Live yeast cells attached to macrophages were seen (green fluorescence); (d) SC5314 cells pre-treated with Compound 4 emitting red fluorescence.
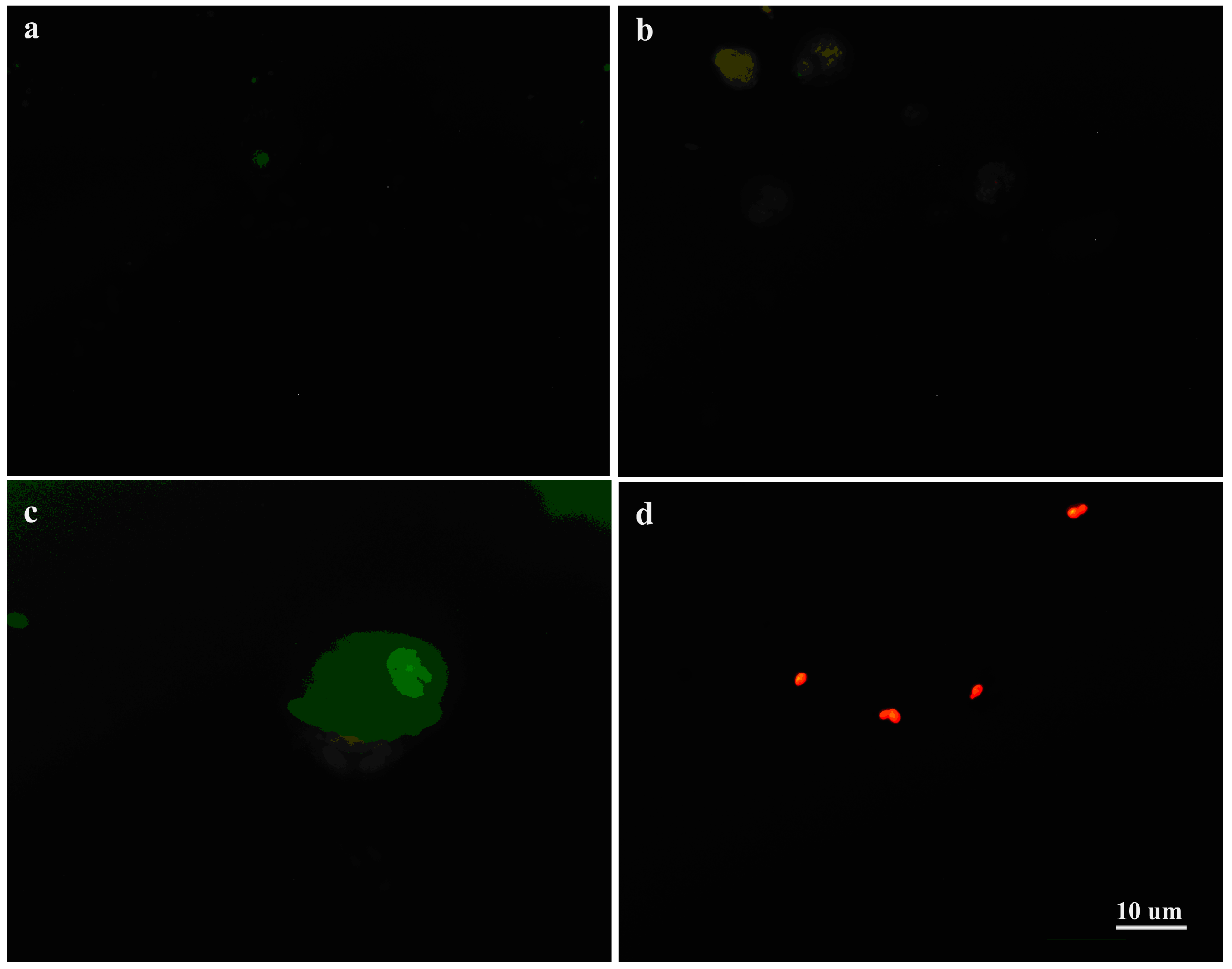
Figure 4.
Expression level of KEX2 normalized with the housekeeping gene ACT1 for both the Compound 4-treated (at 0.25 µg/mL) as well as untreated cells of C. albicans. (a) The transcript levels in the C. albicans cells growing in Yeast extract-peptone-dextrose growth YEPD medium; (b) The transcript levels in the C. albicans cells growing on Caco-2 cells; Legend: § means statistically significant differences between the untreated and treated cells (p < 0.05); * means statistically significant differences between SC5314 and mutants (both treated cells, p < 0.05).
Figure 4.
Expression level of KEX2 normalized with the housekeeping gene ACT1 for both the Compound 4-treated (at 0.25 µg/mL) as well as untreated cells of C. albicans. (a) The transcript levels in the C. albicans cells growing in Yeast extract-peptone-dextrose growth YEPD medium; (b) The transcript levels in the C. albicans cells growing on Caco-2 cells; Legend: § means statistically significant differences between the untreated and treated cells (p < 0.05); * means statistically significant differences between SC5314 and mutants (both treated cells, p < 0.05).
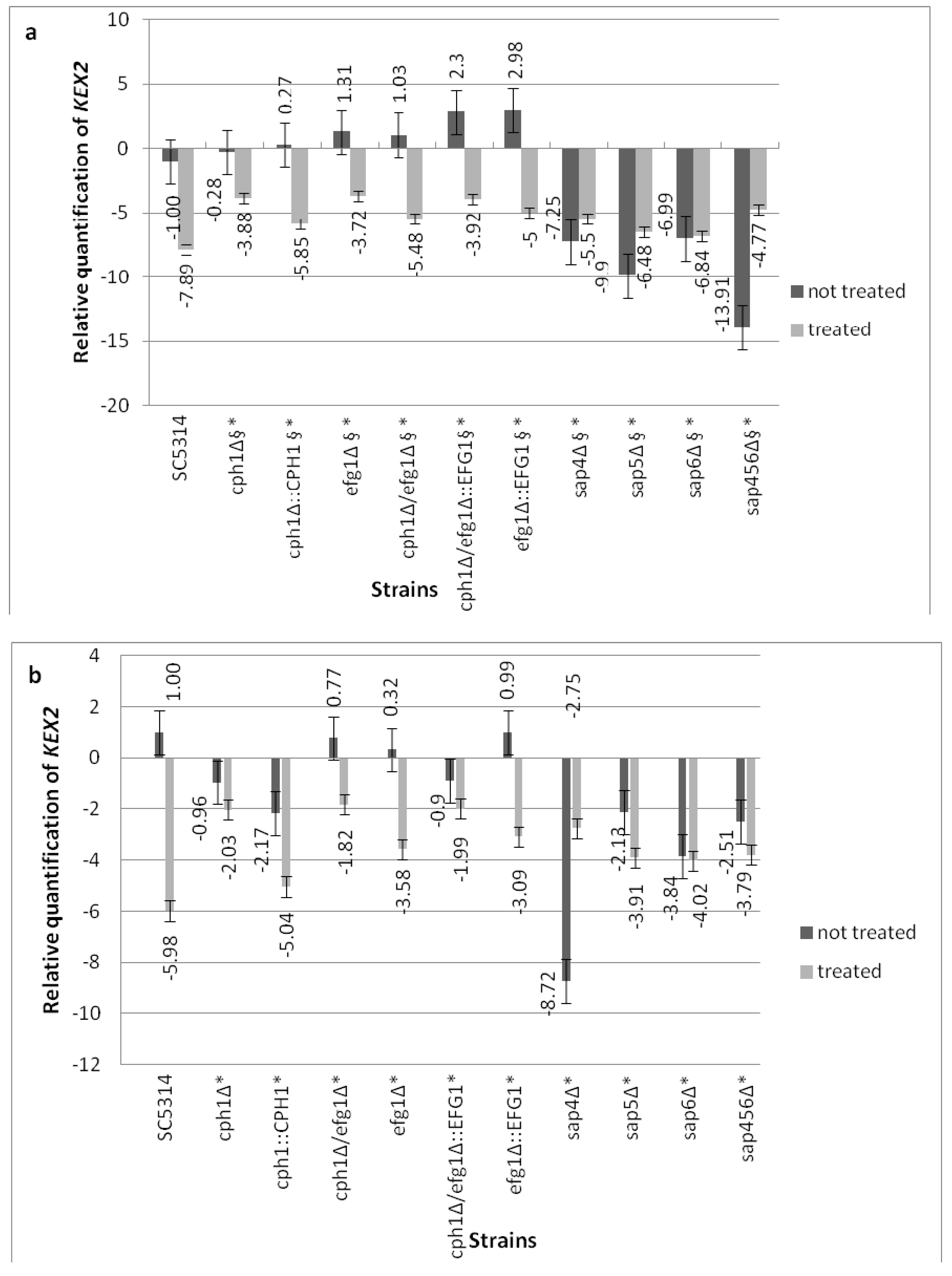
Figure 5.
Production of IL-10 upon interaction of peripheral blood mononuclear cells PBMCs with the Candida mutant cells in the presence or absence of Compound 4 at 0.25 µg/mL. Legend: * stands for statistically significant differences between the viable mutants and their heat-killed counterparts in IL-10 induction. All data are presented as mean values ± standard deviations (SDs).
Figure 5.
Production of IL-10 upon interaction of peripheral blood mononuclear cells PBMCs with the Candida mutant cells in the presence or absence of Compound 4 at 0.25 µg/mL. Legend: * stands for statistically significant differences between the viable mutants and their heat-killed counterparts in IL-10 induction. All data are presented as mean values ± standard deviations (SDs).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The strains used in this study.
| Species | Strains | Parental | Genotype/Characteristic | Reference |
|---|---|---|---|---|
| Candida albicans | 90028 | None | Prototrophic wild-type strain | [12] |
| SC5314 | None | Prototrophic wild-type strain | [13] | |
| CAI4 | SC5314 | ura3Δ::1imm434/ura3Δ::1imm434 | [14] | |
| cph1∆ | CAI4 | ura3Δ::1imm434/ura3Δ::1imm434 cph1Δ::hisGΔ/cph1::hisG-URA3-hisG | [15] | |
| cph1∆ (CPH1) | CAI4 | ura3Δ::1imm434/ura3Δ::1imm434 cph1Δ::hisG/cph1Δ::hisG (CPH1) | [16] | |
| efg1∆ | CAI4 | ura3Δ::1imm434/ura3Δ::1imm434 efg1Δ::hisG/efg1Δ::hisG-URA3-hisG | [16] | |
| cph1∆/efg1∆ | CAI4 | ura3Δ::1imm434/ura3Δ::1imm434Δ cph1::hisG/cph1Δ::hisG Δefg1::hisG/efg1Δ::hisG-URA3-hisG | [16] | |
| efg1∆ (EFG1) | CAI4 | ura3Δ::1 imm434/ura3Δ::1 imm434Δ efg1::hisG/efg1Δ::hisG (EFG1) | [16] | |
| cph1∆/efg1∆ (EFG1) | CAI4 | ura3Δ::1 imm434/ura3Δ::1 imm434 cph1Δ::hisG/cph1Δ::hisG efg1Δ::hisG/efg1Δ::hisG (EFG1) | [16] | |
| sap4∆ | SC5314 | sap4-1Δ::FRT/Δsap4-2::FRT | [17] | |
| sap5∆ | SC5314 | sap5-1Δ::FRT/Δsap5-2::FRT | [17] | |
| sap6∆ | SC5314 | sap6-1Δ::FRT/sap6-2 Δ::FRT | [17] | |
| sap456∆ | SC5314 | sap4-1Δ::FRT/sap4-2Δ::FRT; Δsap5-1::FRT/sap5Δ-2::FRT; sap6-1Δ::FRT/sap6-2Δ::FRT | [17] | |
| kex2∆CNA1 | CAI4 | ura3Δ::imm434/ura3Δ::imm434KEX2kex2Δ::hisGURA3hisG | [18] | |
| kex2∆CNA2 | CAI4 | ura3Δ::imm434/ura3Δ::imm434KEX2kex2Δ::hisG | [18] | |
| kex2∆CNA3 | CAI4 | ura3Δ::imm434/ura3Δ::imm434kex2Δ::hisGkex2Δ::hisGURA3hisG | [18] | |
| kex2∆CNA4 | CAI4 | ura3Δ::imm434/ura3Δ::imm434kex2Δ::hisGkex2Δ::hisG | [18] | |
| Candida glabrata | kex2∆1006 | Cg∆HT6 | his3Δ rp1Δt kex2Δ::HIS3 | [5] |
| kex2∆1008 | 1005 | his3 Δtrp1Δ kex2Δ::HIS3 pKRT1(KEX2 TRP1) | [5] | |
| kex2∆1010 | 1005 | his3Δ trp1Δ kex2Δ::HIS3 pACT-14(TRP1) | [5] |
Table 2.
List of primers used in the study.
| Primers | Sequence (5′-3′) |
|---|---|
| ACT1-1 | GACAATTTCTCTTTCAGCACTAGTAGTGA |
| ACT1-2 | GCTGGTAGAGACTTGACCAACCA |
| KEX2-1 | TTTATATTGGGATATTTATTATCA |
| KEX2-2 | TGGGATTTTAATAATAAAGGCAAA |
Table 3.
Antifungal activity (cell inhibition %; Means ± SD) of 2 bromo-2-chloro-2-(-4-chlorophenyl sulfonyl)-1-phenyloethanon (Compound 4) against Candida strains after 48 h tested by using method M27-A3 [20].
Table 3.
Antifungal activity (cell inhibition %; Means ± SD) of 2 bromo-2-chloro-2-(-4-chlorophenyl sulfonyl)-1-phenyloethanon (Compound 4) against Candida strains after 48 h tested by using method M27-A3 [20].
| Candida Spp. | µg/mL | |||||||
|---|---|---|---|---|---|---|---|---|
| 16 | 8 | 4 | 2 | 1 | 0.5 | 0.5 | 0.5 1 | |
| SC5314 | 99.69 ± 0.01 | 99.55 ± 0.03 | 95.01 ± 0.01 | 92.70 ± 0.21 | 93.14 ± 0.05 | 95.59 ± 0.01 | 93.50 ± 0.01 | 100 |
| Δsap4 3 | 97.97 ± 0.22 | 96.90 ± 0.02 | 95.50 ± 0.01 | 92.85 ± 0.02 | 100 ± 0.04 2 | 99.90 ± 0.03 | 98.89 ± 0.02 | 100 |
| Δsap5 3 | 99.50 ± 0.00 | 99.31 ± 0.01 | 95.10 ± 0.20 | 40.05 ± 0.04 | 100 ± 0.03 2 | 98.32 ± 0.10 | 97.00 ± 0.04 | 100 |
| Δsap6 3 | 99.80 ± 0.05 | 98.90 ± 0.01 | 99.42 ± 0.01 | 99.90 ± 0.01 | 96.95 ± 0.02 | 100 ± 0.01 2 | 97.67 ± 0.01 | 100 |
| Δsap456 3 | 100 ± 0.02 | 98.70 ± 0.02 | 99.13 ± 0.01 | 99.85 ± 0.02 | 100 ± 0.01 | 97.89 ± 0.02 | 98.00 ± 0.04 | 100 |
| Can16 3 | 99.40 ± 0.03 | 99.42 ± 0.04 | 99.94 ± 0.05 | 100 ± 0.216 | 100 ± 0.01 | 100 ± 0.04 2 | 99.00 ± 0.02 | 100 |
| YLO323 | 99.71 ± 0.02 | 99.48 ± 0.03 | 95.67 ± 0.02 | 86.22 ± 0.02 | 92.04 ± 0.02 | 95.53 ± 0.22 | 95.67 ± 004 | 100 |
| HLC52 | 99.63 ± 0.03 | 98.94 ± 0.04 | 99.48 ± 0.025 | 99.74 ± 0.06 | 99.77 ± 0.02 | 98.99 ± 0.04 | 98.90 ± 0.01 | 100 |
| HLC54 3 | 99.75 ± 0.01 | 99.16 ± 0.09 | 99.33 ± 0.08 | 99.83 ± 0.05 | 100 ± 0.06 2 | 99.43 ± 0.09 | 99.00 ± 0.01 | 100 |
| HLC74 3 | 99.76 ± 0.01 | 98.93 ± 0.04 | 98.96 ± 0.04 | 100 ± 0.02 | 99.38 ± 0.01 | 98.85 ± 0.03 | 98.50 ± 0.03 | 100 |
| HLC84 | 99.71 ± 0.02 | 99.31 ± 0.01 | 99.24 ± 0.02 | 98.86 ± 0.04 | 99.15 ± 0.02 | 99.16 ± 0.03 | 99.00 ± 0.01 | 100 |
| CNA1 | 99.06 ± 0.00 | 99.04 ± 0.00 | 99.00 ± 0.01 | 98.98 ± 0.01 | 98.97 ± 0.01 | 98.96 ± 0.01 | 98.00 ± 0.02 | 100 |
| CNA2 | 99.05 ± 0.00 | 99.00 ± 0.01 | 98.99 ± 0.01 | 98.98 ± 0.00 | 98.970 ± 0.00 | 98.95 ± 0.00 | 98.77 ± 0.03 | 100 |
| CNA3 | 99.44 ± 0.00 | 99.19 ± 0.00 | 99.17 ± 0.00 | 98.98 ± 0.00 | 98.77 ± 0.01 | 96.50 ± 0.00 | 96.00 ± 0.01 | 100 |
| CNA4 | 99.96 ± 0.01 | 99.48 ± 0.00 | 99.12 ± 0.01 | 99.11 ± 0.01 | 98.67 ± 0.01 | 95.64 ± 0.01 | 94.78 ± 0.01 | 100 |
| 1006 | 100 ± 0.00 | 100 ± 0.21 2 | 99.47 ± 0.00 | 99.35 ± 0.01 | 99.27 ± 0.00 | 99.01 ± 0.01 | 99.00 ± 0.03 | 100 |
| 1008 | 99.08 ± 0.00 | 99.04 ± 0.01 | 98.99 ± 0.00 | 98.99 ± 0.01 | 98.97 ± 0.01 | 98.97 ± 0.00 | 97.90 ± 0.02 | 100 |
| 1010 | 100 ± 0.00 2 | 99.27 ± 0.01 | 99.21 ± 0.00 | 99.16 ± 0.00 | 98.94 ± 0.01 | 98.94 ± 0.00 | 98.94 ± 0.03 | 100 |
1 Amphotericin B at a concentration of 0.5 µg/mL was used as a control (clear endpoint = cell growth inhibition at 100%); 2 stands for clear endpoint (cell growth inhibition % = 100); 3 stands for PG phenotype; Results were confirmed in three separate experiments on different days.
Table 4.
The percentage of adhesion of Candida cells to the Caco-2 cell line after pre-treatment with 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone (named Compound 4). Adhesion data was calculated for cells grown on Sabouraud agar of a 24-well-plate. Adherence was expressed as a percentage of the total number of cells added (control cells non-treated). Data are expressed as the mean ± SD of three independent experiments. Values in bold indicate significantly affected adhesive properties compared to the non-treated counterparts (p ≤ 0.05).
Table 4.
The percentage of adhesion of Candida cells to the Caco-2 cell line after pre-treatment with 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone (named Compound 4). Adhesion data was calculated for cells grown on Sabouraud agar of a 24-well-plate. Adherence was expressed as a percentage of the total number of cells added (control cells non-treated). Data are expressed as the mean ± SD of three independent experiments. Values in bold indicate significantly affected adhesive properties compared to the non-treated counterparts (p ≤ 0.05).
| Species | Strains | Genotypes | Cells Treated at 0.25 µg/mL | Non Treated Cells |
|---|---|---|---|---|
| Candida albicans | SC5314 (wt) | Prototrophic wild-type strain | 29.99 ± 0.34 | 59.34 ± 0.06 |
| sap4Δ | sap4-1Δ::FRT/Δsap4-2::FRT | 76.67 ± 0.15 | 46.32 ± 0.16 | |
| sap5Δ | sap5-1Δ::FRT/Δsap5-2::FRT | 48.88 ± 0.17 | 35.55 ± 0.21 | |
| sap6Δ | sap6-1Δ::FRT/sap6-2 Δ::FRT | 25.29 ± 0.16 | 22.99 ± 0.04 | |
| sap4Δ/sap5Δ/sap6 | sap4-1Δ::FRT/sap4-2Δ::FRT; Δsap5-1::FRT/sap5Δ-2::FRT; sap6-1Δ::FRT/sap6-2Δ::FRT | 52.91 ± 0.33 | 15.88 ± 0.03 | |
| cph1Δ | ura3Δ::1imm434/ura3Δ::1imm434 cph1Δ::hisG/cph1Δ::hisG | 63.89 ± 0.25 | 35.13 ± 0.24 | |
| cph1Δ::CPH1 | ura3Δ::1imm434/ura3Δ::1imm434 cph1Δ::hisG/cph1Δ::hisG (CPH1) | 30.56 ± 0.03 | 12.82 ± 0.09 | |
| efg1Δ | ura3Δ::1imm434/ura3Δ::1imm434 efg1Δ::hisG/efg1Δ::hisG-URA3-hisG | 46.40 ± 0.25 | 29.26 ± 0.16 | |
| cph1Δ/efg1Δ | ura3Δ::1imm434/ura3Δ::1imm434Δ cph1::hisG/cph1Δ::hisG Δefg1::hisG/efg1Δ::hisG-URA3-hisG | 68.00 ± 0.16 | 72.21 ± 0.22 | |
| efg1Δ::EFG1 | ura3Δ::1 imm434/ura3Δ::1 imm434Δ efg1::hisG/efg1Δ::hisG (EFG1) | 48.33 ± 0.12 | 8.34 ± 0.09 | |
| cph1Δ/efg1Δ::EFG1 | ura3Δ::1 imm434/ura3Δ::1 imm434 cph1Δ::hisG/cph1Δ::hisG efg1Δ::hisG/efg1Δ::hisG (EFG1) | 61.87 ± 0.38 | 48.55 ± 0.078 | |
| kex2Δ CNA1 | ura3Δ::imm434/ura3Δ::imm434KEX2kex2Δ::hisGURA3hisG | 37.85 ± 0.04 | 55.8 ± 0.18 | |
| kex2Δ CNA2 | ura3Δ::imm434/ura3Δ::imm434KEX2kex2Δ::hisG | 22.49 ± 0.12 | 62.45 ± 0.27 | |
| kex2Δ CNA3 | ura3Δ::imm434/ura3Δ::imm434kex2Δ::hisGkex2Δ::hisGURA3hisG | 48.17 ± 0.41 | 52.33 ± 0.16 | |
| kex2Δ CNA4 | ura3Δ::imm434/ura3Δ::imm434kex2Δ::hisGkex2Δ::hisG | 7.33 ± 0.06 | 22.20 ± 0.06 | |
| Candida glabrata | Cg kex2Δ 1006 | his3Δ rp1Δt kex2Δ::HIS3 | 45.13 ± 0.22 | 56.99 ± 0.24 |
| Cg kex2Δ 1008 | his3Δtrp1Δ kex2Δ::HIS3 pKRT1 (KEX2 TRP1) | 17.43 ± 0.08 | 82.15 ± 0.19 | |
| Cg kex2Δ 1010 | his3Δ trp1Δ kex2Δ::HIS3 pACT-14 (TRP1) | 18.34 ± 0.07 | 53.85 ± 0.28 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Staniszewska, M.; Bondaryk, M.; Ochal, Z. Role of Virulence Determinants in Candida albicans’ Resistance to Novel 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone. J. Fungi 2017, 3, 32. https://doi.org/10.3390/jof3030032
AMA Style
Staniszewska M, Bondaryk M, Ochal Z. Role of Virulence Determinants in Candida albicans’ Resistance to Novel 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone. Journal of Fungi. 2017; 3(3):32. https://doi.org/10.3390/jof3030032
Chicago/Turabian StyleStaniszewska, Monika, Małgorzata Bondaryk, and Zbigniew Ochal. 2017. "Role of Virulence Determinants in Candida albicans’ Resistance to Novel 2-bromo-2-chloro-2-(4-chlorophenylsulfonyl)-1-phenylethanone" Journal of Fungi 3, no. 3: 32. https://doi.org/10.3390/jof3030032
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.