
Correlation Between Microbial Communities and Volatile Organic Compounds in Camel Milk at Different Lactation Stages in Xinjiang, China

Abstract
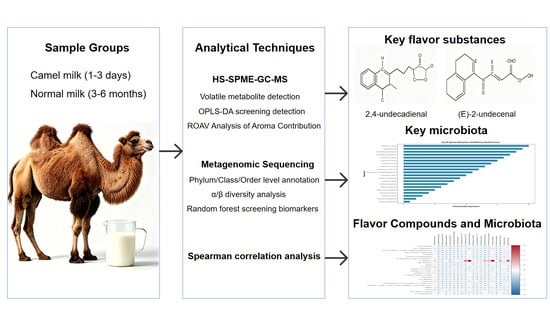
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. Microbial Metagenomic Analysis
2.3. Solid-Phase Microextraction Extraction
2.4. GC-MS
2.5. Qualitative and Quantitative Analysis of GC-MS
2.6. Relative Odor Activity Value
2.7. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Characteristic Flavor of Camel Milk at Different Stages of Lactation by GC-MS
3.2. OPLS-DA Analysis of Volatile Flavor Compounds
3.3. ROAV Analysis of Key Volatile Compounds in Camel Milk at Different Lactation Stages
3.4. Bacterial Composition and Taxonomic Annotation of Camel Milk from Different Lactations
3.5. Alpha Diversity and Beta Diversity Analysis of Microbiota in Camel Milk from Different Lactations
3.6. Correlation Analysis Between Microorganisms and Flavor in Camel Milk at Different Lactation Stages
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ho, T.M.; Zou, Z.; Bansal, N. Camel milk: A review of its nutritional value, heat stability, and potential food products. Food Res. Int. 2022, 153, 110870. [Google Scholar] [CrossRef]
- Arain, M.A.; Rasheed, S.; Jaweria, A.; Khaskheli, G.B.; Barham, G.S.; Ahmed, S. A Review on Processing Opportunities for the Development of Camel Dairy Products. Food Sci. Anim. Resour. 2023, 43, 383–401. [Google Scholar] [CrossRef]
- Alhaj, O.A.; Ibrahim, M.O.; Elsahoryi, N.A.; Al-Maseimi, O.D. Processing-Induced Modifications of Camel Milk Immunoglobulins and Lactoferrin: Implications for Immunocompromised Pediatric Populations and Therapeutic Applications. Foods 2026, 15, 1028. [Google Scholar] [CrossRef]
- Almasri, R.S.; Bedir, A.S.; Ranneh, Y.K.; El-Tarabily, K.A.; Al Raish, S.M. Benefits of camel milk over cow and goat milk for infant and adult health in fighting chronic diseases: A review. Nutrients 2024, 16, 3848. [Google Scholar] [CrossRef] [PubMed]
- Bhumireddy, S.R.; Rocchetti, G.; Pallerla, P.; Lucini, L.; Sripadi, P. A combined targeted/untargeted screening based on GC/MS to detect low-molecular-weight compounds in different milk samples of different species and as affected by processing. Int. Dairy J. 2021, 118, 105045. [Google Scholar] [CrossRef]
- He, J.; Wu, X.; Yu, Z. Microwave pretreatment of camellia (Camellia oleifera Abel.) seeds: Effect on oil flavor. Food Chem. 2021, 364, 130388. [Google Scholar] [CrossRef]
- Son, J.H.; Islam, A.; Hong, J.H.; Jeong, J.Y.; Song, O.Y.; Kim, H.E.; Khan, N.; Jamila, N.; Kim, K.S. Extraction of volatile organic compounds from leaves of Ambrosia artemisiifolia L. and Artemisia annua L. by headspace-solid phase micro extraction and simultaneous distillation extraction and analysis by gas chromatography/mass spectrometry. Food Sci. Biotechnol. 2021, 30, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Guo, Y.; Zhang, Y.; Pang, X.; Wang, Y.; Lv, J.; Zhang, S. Effects of different heat treatments on maillard reaction products and volatile substances of camel milk. Front. Nutr. 2023, 10, 1072261. [Google Scholar] [CrossRef]
- Nalepa, B.; Olszewska, M.A.; Markiewicz, L.H. Seasonal variances in bacterial microbiota and volatile organic compounds in raw milk. Int. J. Food Microbiol. 2018, 267, 70–76. [Google Scholar] [CrossRef]
- Xiong, Z.-Q.; Li, Y.-Y.; Xiang, Y.-W.; Xia, Y.-J.; Zhang, H.; Wang, S.-J.; Ai, L.-Z. Short communication: Dynamic changes in bacterial diversity during the production of powdered infant formula by PCR-DGGE and high-throughput sequencing. J. Dairy Sci. 2020, 103, 5972–5977. [Google Scholar] [CrossRef]
- Zhang, C.; Bijl, E.; Muis, K.E.; Hettinga, K. Stability of fat globules in UHT milk during proteolysis by the AprX protease from Pseudomonas fluorescens and by plasmin. J. Dairy Sci. 2020, 103, 179–190. [Google Scholar] [CrossRef]
- Soto Del Rio, M.L.D.; Dalmasso, A.; Civera, T.; Bottero, M.T. Characterization of bacterial communities of donkey milk by high-throughput sequencing. Int. J. Food Microbiol. 2017, 251, 67–72. [Google Scholar] [CrossRef]
- Jeong, J.; Yun, K.; Mun, S.; Chung, W.-H.; Choi, S.-Y.; Nam, Y.-D.; Lim, M.Y.; Hong, C.P.; Park, C.; Ahn, Y.J.; et al. The effect of taxonomic classification by full-length 16S rRNA sequencing with a synthetic long-read technology. Sci. Rep. 2021, 11, 1727. [Google Scholar] [CrossRef] [PubMed]
- Lyu, C.; Chen, C.; Ge, F.; Liu, D.; Zhao, S.; Chen, D. A preliminary metagenomic study of puer tea during pile fermentation. J. Sci. Food Agric. 2013, 93, 3165–3174. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.Y.; Mei, R.; Wu, Z.; Lee, P.K.; Liu, W.-T.; Lee, P.-H. Superior resolution characterisation of microbial diversity in anaerobic digesters using full-length 16S rRNA gene amplicon sequencing. Water Res. 2020, 178, 115815. [Google Scholar] [CrossRef] [PubMed]
- Niyazbekova, Z.; Yao, X.-T.; Liu, M.-J.; Bold, N.; Tong, J.-Z.; Chang, J.-J.; Wen, Y.; Li, L.; Wang, Y.; Chen, D.-K.; et al. Compositional and functional comparisons of the microbiota in the colostrum and mature milk of dairy goats. Animals 2020, 10, 1955. [Google Scholar] [CrossRef]
- Pranata, J.; Cadwallader, D.C.; Drake, M.; Barbano, D.M. Effects of storage time and temperature on the protein fraction of aseptic milk. J. Dairy Sci. 2025, 108, 8086–8097. [Google Scholar] [CrossRef]
- Quince, C.; Walker, A.W.; Simpson, J.T.; Loman, N.J.; Segata, N. Shotgun metagenomics, from sampling to analysis. Nat. Biotechnol. 2017, 35, 833–844. [Google Scholar] [CrossRef]
- Chi, X.; Shao, Y.; Pan, M.; Yang, Q.; Yang, Y.; Zhang, X.; Ai, N.; Sun, B. Distinction of volatile flavor profiles in various skim milk products via HS-SPME–GC–MS and E-nose. Eur. Food Res. Technol. 2021, 247, 1539–1551. [Google Scholar] [CrossRef]
- Wang, D.; Zhou, Y.; Zhao, J.; Guo, Y.; Yan, W. Analysis of Flavor Differences in Yak Milk Powder at Different Milk Production Stages by Headspace Solid-Phase Microextraction and Gas Chromatography–Mass Spectrometry. Foods 2025, 14, 91. [Google Scholar] [CrossRef]
- Dong, W.; Hu, R.; Long, Y.; Li, H.; Zhang, Y.; Zhu, K.; Chu, Z. Comparative evaluation of the volatile profiles and taste properties of roasted coffee beans as affected by drying method and detected by electronic nose, electronic tongue, and HS-SPME-GC-MS. Food Chem. 2019, 272, 723–731. [Google Scholar] [CrossRef]
- Zhao, M.; Li, T.; Yang, F.; Cui, X.; Zou, T.; Song, H.; Liu, Y. Characterization of key aroma-active compounds in Hanyuan Zanthoxylum bungeanum by GC-O-MS and switchable GC× GC-O-MS. Food Chem. 2022, 385, 132659. [Google Scholar] [CrossRef]
- Liu, H.; Hui, T.; Fang, F.; Ma, Q.; Li, S.; Zhang, D.; Wang, Z. Characterization and discrimination of key aroma compounds in pre-and postrigor roasted mutton by GC-O-MS, GC E-Nose and aroma recombination experiments. Foods 2021, 10, 2387. [Google Scholar] [CrossRef]
- Mais, E.; Alolga, R.N.; Wang, S.L.; Linus, L.O.; Yin, X.; Qi, L.W. A comparative UPLC-Q/TOF-MS-based metabolomics approach for distinguishing Zingiber officinale Roscoe of two geographical origins. Food Chem. 2018, 240, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Cui, C.; Zhang, S.; Zhu, J.; Peng, C.; Cai, H.; Yang, X.; Hou, R. Use of headspace GC/MS combined with chemometric analysis to identify the geographic origins of black tea. Food Chem. 2021, 360, 130033. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wen, D.; Zhou, Z.; Suolang, Q.; Siwang, W.; Kangji, L.; Tang, T.; Liu, Z.; Wang, Y. Dietary fiber enhances milk yield in plateau dairy cows via activation of the rumen microbiota-mammary gland axis. Front. Vet. Sci. 2025, 12, 1654799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kollie, S.C.; Xia, T.; Yang, Z.; Ian, M.T.; Elolimy, A.A.; Wang, W.; Lu, D.; Li, Y.; Li, M.; et al. Rumen-Plasma-Milk Metabolomics Profiling Revealed Metabolic Alterations Associated with Milk Fat Synthesis in Chinese Holstein Cows. Animals 2026, 16, 1136. [Google Scholar] [CrossRef]
- Sakwinska, O.; Bosco, N. Host microbe interactions in the lactating mammary gland. Front. Microbiol. 2019, 10, 1863. [Google Scholar] [CrossRef]
- Timlin, M.; Tobin, J.T.; Brodkorb, A.; Murphy, E.G.; Dillon, P.; Hennessy, D.; O’donovan, M.; Pierce, K.M.; O’callaghan, T.F. The impact of seasonality in pasture-based production systems on milk composition and functionality. Foods 2021, 10, 607. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, X.; Hu, R.; Zhang, J.; Dong, W. Effects of different roasting temperatures on the flavor characteristics of shelled macadamia kernels: Analysis based on HS-GC-IMS and HS-SPME-GC–MS. Food Res. Int. 2026, 232, 118878. [Google Scholar] [CrossRef]
- Fu, L.; Tian, J.; Liu, L.; Ma, Y.; Zhang, X.; Ma, C.; Kang, W.; Sun, Y. Analysis of Volatile Components in Tremella fuciformis by Electronic Nose Combined with GC-MS. J. Food Qual. 2022, 1, 9904213. [Google Scholar] [CrossRef]
- Forte, L.; Parabita, N.; Santoro, M.; Longobardi, F.; Natrella, G.; Quiñones, J.; Ponnampalam, E.N.; Tomasevic, I.; De Palo, P.; Maggiolino, A. From rumen to milk: Dietary polyphenols in dairy cows-A critical review. Vet. Anim. Sci. 2026, 31, 100569. [Google Scholar] [CrossRef]
- Bilal, Z.; Kondybayev, A.; Ospanova, A.; Tormo, H.; Akhmetsadykova, S.; Amutova, F.; Faye, B.; Konuspayeva, G. Volatile organic compounds of camel milk and shubat across Kazakhstan’s regions, seasons, and breeds. Heliyon 2024, 10, e35365. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, H.; Li, L.; Tan, J.; Wang, Y.; Liu, M.; Jiang, L. Multi-omics analysis reveals that the metabolite profile of raw milk is associated with dairy cows’ health status. Food Chem. 2023, 428, 136813. [Google Scholar] [CrossRef]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Rosa, L.; Ianiro, G.; Conte, A.L.; Conte, M.P.; Ottolenghi, L.; Valenti, P.; Cutone, A. Antibacterial, anti-invasive, and anti-inflammatory activity of bovine lactoferrin extracted from milk or colostrum versus whole colostrum. Biochem. Cell Biol. 2024, 102, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Králová, S. Role of fatty acids in cold adaptation of Antarctic psychrophilic Flavobacterium spp. Syst. Appl. Microbiol. 2017, 40, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Lérias, J.R.; Hernández-Castellano, L.E.; Suárez-Trujillo, A.; Castro, N.; Pourlis, A.; Almeida, A.M. The mammary gland in small ruminants: Major morphological and functional events underlying milk production—A review. J. Dairy Res. 2014, 81, 304–318. [Google Scholar] [CrossRef]
- Sauvageot, N.; Mokhtari, A.; Joyet, P.; Budin-Verneuil, A.; Blancato, V.S.; Repizo, G.D.; Henry, C.; Pikis, A.; Thompson, J.; Magni, C.; et al. Enterococcus faecalis uses a phosphotransferase system permease and a host colonization-related ABC transporter for maltodextrin uptake. J. Bacteriol. 2017, 199, e00878-16. [Google Scholar] [CrossRef]
- Zeng, L.; Burne, R.A. Subpopulation behaviors in lactose metabolism by Streptococcus mutans. Mol. Microbiol. 2021, 115, 58–69. [Google Scholar] [CrossRef]
- Vertès, A.A. Protein Secretion Systems of Corynebacterium glutamicum; Springer: Berlin/Heidelberg, Germany, 2012; pp. 351–389. [Google Scholar] [CrossRef]
- Shi, H.; Zhu, J.; Luo, J.; Cao, W.; Shi, H.; Yao, D.; Li, J.; Sun, Y.; Xu, H.; Yu, K.; et al. Genes regulating lipid and protein metabolism are highly expressed in mammary gland of lactating dairy goats. Funct. Integr. Genom. 2015, 15, 309–321. [Google Scholar] [CrossRef]
- Hinsu, A.T.; Tulsani, N.J.; Panchal, K.J.; Pandit, R.J.; Jyotsana, B.; Dafale, N.A.; Patil, N.V.; Purohit, H.J.; Joshi, C.G.; Jakhesara, S.J. Characterizing rumen microbiota and CAZyme profile of Indian dromedary camel (Camelus dromedarius) in response to different roughages. Sci. Rep. 2021, 11, 9400. [Google Scholar] [CrossRef]
- Lin, L.; Shao, X.; Yang, Y.; Murero, A.K.; Wang, L.; Xu, G.; Zhao, Y.; Han, S.; Su, Z.; Xu, K.; et al. Lysobacter enzymogenes: A fully armed biocontrol warrior. J. Integr. Agric. 2025, 24, 23–25. [Google Scholar] [CrossRef]
- Elizondo-Reyna, E.; Martínez-Montoya, H.; Tamayo-Ordoñez, Y.; Cruz-Hernández, M.A.; Carrillo-Tripp, M.; Tamayo-Ordoñez, M.C.; Sosa-Santillán, G.d.J.; la Garza, J.A.R.-D.; Hernández-Guzmán, M.; Bocanegra-García, V.; et al. Insights from a Genome-Wide Study of Pantoea agglomerans UADEC20: A Promising Strain for Phosphate Solubilization and Exopolysaccharides Production. Curr. Issues Mol. Biol. 2025, 47, 56. [Google Scholar] [CrossRef]
- Zhou, M.; Huang, F.; Du, X.; Wang, C.; Liu, G. Microbial Quality of Donkey Milk during Lactation Stages. Foods 2023, 12, 4272. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, J.; Liu, H.; Feng, C.; Shuang, Q. Novel insight into physicochemical and flavor formation in koumiss based on microbial metabolic network. Food Res. Int. 2021, 149, 110659. [Google Scholar] [CrossRef]
- Ming, L.; Yi, L.; Ai, Y.; Ji, R. A new insight into the exosome protein and lipid composition in camel colostrum and mature milk using comparative proteome and lipidomics analyses. Food Chem. X 2025, 29, 102729. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, F.J.; Zamora, R. Amino acid degradations produced by lipid oxidation products. Crit. Rev. Food Sci. Nutr. 2016, 56, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- da Silva Rodrigues, R.; Machado, S.G.; Nero, L.A. Spoilage microbial groups in dairy industry. Adv. Food Nutr. Res. 2025, 113, 519–561. [Google Scholar] [CrossRef]
- Sun, M.; Shao, W.; Liu, Z.; Ma, X.; Chen, H.; Zheng, N.; Zhao, Y. Microbial diversity in camel milk from Xinjiang, China as revealed by metataxonomic analysis. Front. Microbiol. 2024, 15, 1367116. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | Material Name | Camel Colostrum (%) | Regular Camel Milk (%) | p Value | VIP Value |
|---|---|---|---|---|---|
| 1 | .delta.-Nonalactone | 5.27 ± 1.91 | 6.72 ± 2.94 | 0.004 | 1.210 |
| 2 | Betazole | 4.24 ± 1.87 | 4.82 ± 2.27 | 0.008 | 1.204 |
| 3 | 1-Tridecyne | 0.51 ± 0.18 | 0.65 ± 0.23 | 0.004 | 1.239 |
| 4 | Dodecane, 2,6,11-trimethyl- | 1.84 ± 0.69 | 2.07 ± 0.77 | 0.004 | 1.239 |
| 5 | 2,4-Diamino-6-methyl-1,3,5-triazine | 1.51 ± 0.66 | 1.59 ± 0.75 | 0.007 | 1.204 |
| 6 | 3,5-Heptanedione, 4-ethyl-2,2,6,6-tetramethyl- | 15.20 ± 6.54 | 14.83 ± 7.48 | 0.006 | 1.216 |
| 7 | 2-pentyl-Piperidine | 28.83 ± 11.07 | 2.15 ± 14.34 | 0.004 | 1.225 |
| 8 | 3-Dodecanone | 1.05 ± 0.42 | 1.25 ± 0.40 | 0.006 | 1.283 |
| 9 | 3-Ethyl-2,6,10-trimethylundecane | 17.41 ± 6.80 | 19.42 ± 4.71 | 0.005 | 1.279 |
| 10 | 6-Tridecene | 19.77 ± 7.62 | 21.85 ± 9.80 | 0.004 | 1.223 |
| 11 | Cyclohexane, 1-ethenyl-1-methyl-2,4-bis(1-methylethenyl)-, (1.alpha.,2.beta.,4.beta.)- | 0.88 ± 0.38 | 0.90 ± 0.41 | 0.006 | 1.239 |
| 12 | 2,4-Undecadienal | 1.03 ± 0.32 | 1.28 ± 0.41 | 0.002 | 1.299 |
| 13 | 2,6,10-Trimethyltridecane | 28.11 ± 13.87 | 19.37 ± 10.42 | 0.009 | 1.204 |
| 14 | 2-Undecenal, E- | 6.41 ± 2.96 | 5.76 ± 3.79 | 0.002 | 1.236 |
| 15 | threo-2,5-Dimethyl-2- (2-methyl-2-tetrahydrofuryl) tetrahydrofuran | 25.40 ± 8.94 | 30.82 ± 12.62 | 0.003 | 1.228 |
| 16 | Naphthalene, 2-methyl- | 1.67 ± 0.65 | 1.86 ± 0.86 | 0.004 | 1.227 |
| 17 | 2(4H)-Benzofuranone, 5,6,7,7a-tetrahydro-4,4,7a-trimethyl-, (R)- | 0.96 ± 0.50 | 0.88 ± 0.35 | 0.013 | 1.223 |
| 18 | Fumaric acid, ethyl 2-methylallyl ester | 20.22 ± 7.07 | 22.90 ± 11.33 | 0.003 | 1.240 |
| 19 | 10-Undecenal | 1.94 ± 0.68 | 2.35 ± 0.86 | 0.003 | 1.219 |
| 20 | 2-Undecanone | 0.92 ± 0.31 | 1.18 ± 0.50 | 0.002 | 1.224 |
| 21 | Biphenyl | 0.37 ± 0.14 | 0.25 ± 0.21 | 0.002 | 1.214 |
| 22 | cis-4,5-Epoxy-(E)-2-decenal | 0.27 ± 0.06 | 0.20 ± 0.20 | 0.000 | 1.318 |
| 23 | 1,3-Cyclohexanediamine | 0.12 ± 0.02 | 0.07 ± 0.00 | 0.000 | 1.417 |
| 24 | Dodecane, 2,6,10-trimethyl- | 1.84 ± 0.69 | 2.07 ± 0.77 | 0.004 | 1.239 |
| Serial Number | Material Name | Flavor Description | Threshold (μg/kg) | Camel Colostrum | Regular Camel Milk |
|---|---|---|---|---|---|
| 1 | 2,4-Undecadienal | green, buttery, spicy, baked, fruity, fatty, aldehydic, chicken | 0.00001 | 100 | 27.72 |
| 2 | 2-Undecenal, E- | fresh, fruity, citrus, orange, peel | 0.00078 | 7.95 | 1.62 |
| 3 | 10-Undecenal | waxy, aldehydic, rose, mandarin, citrus, soapy, fatty | 0.0035 | 0.54 | 0.15 |
| 4 | 2(4H)-Benzofuranone,5,6,7,7a-tetrahydro-4,4,7a-trimethyl-, (R)- | musky, coumarin | 0.0021 | 0.44 | 0.09 |
| 5 | Naphthalene, 2-methyl- | sweet, floral, woody | 0.004 | 0.42 | 0.10 |
| 6 | 2-Undecanone | waxy, fruity, creamy, fatty, orris, floral | 0.0062 | 0.16 | 0.04 |
| 7 | 3-Dodecanone | - | 0.0083 | 0.12 | 0.03 |
| 8 | Biphenyl | pungent, rose, green, geranium | 0.0033 | 0.11 | 0.02 |
| 9 | .delta.-Nonalactone | coconut, creamy, sweet, milky, coumarin | 2.6 | 0.002 | 0.0006 |
| Group | Chao 1 | Observed Species | Shannon | Simpson | Pielou’s Evenness | Good’s Coverage |
|---|---|---|---|---|---|---|
| C | 372.28 ± 68.66 | 285.67 ± 56.56 | 1.07 ± 0.92 | 0.30 ± 0.29 | 0.19 ± 0.17 | 1 ± 0 |
| N | 1129.23 ± 223.27 | 689.0 ± 121.69 | 4 ± 0.22 | 0.96 ± 0.01 | 0.61 ± 0.02 | 0.99 ± 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Duo, Q.; Zhao, Y.; Osman, H.; Shao, W.; Zhao, Y. Correlation Between Microbial Communities and Volatile Organic Compounds in Camel Milk at Different Lactation Stages in Xinjiang, China. Foods 2026, 15, 1804. https://doi.org/10.3390/foods15101804
Duo Q, Zhao Y, Osman H, Shao W, Zhao Y. Correlation Between Microbial Communities and Volatile Organic Compounds in Camel Milk at Different Lactation Stages in Xinjiang, China. Foods. 2026; 15(10):1804. https://doi.org/10.3390/foods15101804
Chicago/Turabian StyleDuo, Qianqian, Yan Zhao, Henigul Osman, Wei Shao, and Yankun Zhao. 2026. "Correlation Between Microbial Communities and Volatile Organic Compounds in Camel Milk at Different Lactation Stages in Xinjiang, China" Foods 15, no. 10: 1804. https://doi.org/10.3390/foods15101804
APA StyleDuo, Q., Zhao, Y., Osman, H., Shao, W., & Zhao, Y. (2026). Correlation Between Microbial Communities and Volatile Organic Compounds in Camel Milk at Different Lactation Stages in Xinjiang, China. Foods, 15(10), 1804. https://doi.org/10.3390/foods15101804