1. Introduction
The technology for generating plants with a gametic number of chromosomes and using them to produce completely homozygous inbred lines is generally referred to as doubled haploid (DH) technology. DH technology can fix gametes from segregating populations into homozygous inbred lines in a single generation, thereby dramatically accelerating the pace at which improved cultivars can be developed. DH technology is recognized as one of the important tools in maize breeding to accelerate genetic gains. The use of DH lines in maize breeding increases the genetic variance among the lines tested, phenotype-to-genotype correlation and selection accuracy [
1,
2]. Together, DH greatly improves the efficiency of breeding programs and potentially reduces the costs in the development of improved cultivars.
DH technology in maize encompasses (1) the production of haploids by crossing the germplasm from which the DH lines are anticipated with pollen from maternal haploid inducers, (2) the identification of haploids from diploids at the seed or seedling stage using genetic markers or natural differences among haploids and diploids, (3) subjecting the haploid seedlings to chromosomal-doubling treatments using mitosis-inhibiting chemicals, and (4) caring for the treated haploid plants in the field (referred to as D
0 nursery) and self-pollinating fertile DH plants to produce seed for DH lines (D
1 lines). The process of maize DH-line production is well documented in several publications [
2,
3,
4]. Great advances have been made in improving the efficiencies of haploid induction and identification [
2]. However, in the step of chromosomal doubling and the production of seed for DH lines, there is considerable scope for improving the efficiency and reducing the costs. The chromosomal-doubling step encompasses growing haploid seedlings in a growth chamber or a greenhouse, treating them with mitosis-inhibiting chemicals, post-treatment recovery in the greenhouse, and transplanting the seedlings to the field with some protocol-specific modifications [
2,
4]. The chemicals most commonly used for chromosomal doubling are colchicine, antimitotic herbicides, and N
2O gas [
5,
6,
7]. The purpose of chromosomal-doubling treatments is to produce pollen and form seeds from reproductively sterile haploid plants [
8]. Upon chromosomal-doubling treatments, some proportion of surviving haploids may double the chromosomes in the reproductive tissues and produce pollen and seed. The overall success rate (OSR) is a criterion that measures success for artificial chromosomal-doubling methods [
7], and the OSR varies from 5 to 35% depending on the protocol and the chemical used [
6,
7,
9]. Even though artificial chromosome-doubling methods are extensively practiced in the DH-production pipelines, this process demands considerable manual labor and laboratory and greenhouse facilities, besides employing toxic chemicals [
2,
10]. Hence, breeding programs are seeking alternative methods that can be operationally simpler, less expensive and not involve toxic chemicals.
A high proportion of haploids from very few maize genotypes show fertility without the application of anti-mitotic chemicals through spontaneously doubling their chromosomes, even though the reasons are not understood well [
11]. The genotype dependency of spontaneous doubling has limited its widespread use in DH pipelines to date. If this limitation can be addressed, spontaneous chromosomal doubling could dramatically simplify the chromosomal-doubling step and reduce the costs significantly [
10]; hence, it is pursued as an alternative to artificial chromosomal-doubling methods. Most haploid plants are infertile in their male reproductive organs, namely, tassels, but female reproductive organs, namely, ears, produce seed when pollinated with pollen from diploid plants even without being subjected to artificial chromosomal-doubling treatments [
12,
13]. Hence, the lack of haploid male fertility (HMF) is a constraint for producing seed for DH lines when relying on spontaneous doubling. The analysis of HMF in large numbers of inbreds showed greater genetic variance in temperate [
14] and tropical germplasm [
11]. QTL-mapping studies using bi-parental populations [
15,
16,
17,
18] and genome-wide association studies (GWAS) [
11,
14] indicated that HMF is controlled by a few QTLs with major effects. Another major aspect of spontaneous doubling that is also important for DH-production pipelines is overall haploid fertility (HF) [
11] or overall fertile haploid plants [
19], which considers both male and female fertility and is based on seed production from DH plants. A greater genetic variance was also detected for HF in tropical inbred lines, and GWAS identified 11 significantly associated single-nucleotide polymorphisms (SNPs) with HF [
11]. However, HF is less studied than HMF, and there are no previous studies revealing the genetic components and inheritance of this important trait.
In maize-breeding programs, combining-ability analyses are widely used to generate genetic information, such as general combining ability (GCA) and specific combining ability (SCA), for use in genetic-diversity evaluation, parental selection and hybrid development [
20,
21,
22]. Combining-ability analyses are always performed through diallel mating methods developed by Griffing [
23]. A diallel analysis of HMF involving six temperate inbreds revealed significant GCA and SCA effects [
24]. Additive effects were noted to be important for the selection of HMF, while epistatic effects could also play a role [
25]. The HMF can be increased by single-plant selection and recurrent selection [
25]. However, such information is not available for tropical germplasm. Moreover, there are no studies reported to date on the GCA and SCA effects for HF. Therefore, this study was designed to (i) evaluate the GCA and SCA effects for HMF and HF, and the maternal effects of selected maize inbred lines, and (ii) predict the performance of the hybrid for HMF and HF based on mid-parent value and GCA effects.
3. Discussion
Improving the chromosomal-doubling efficiency in maize haploids can unfold the full potential of DH technology, which could facilitate new selection approaches that are currently not feasible due to the lower efficiency of artificial chromosomal-doubling methods. High chromosomal-doubling efficiency can facilitate phenotypic or marker-assisted selection for simple traits and genomic selection for complex traits at the haploid stage [
10]. This could reduce breeding-cycle time and increase selection intensity and efficiency. This study confirmed previous observations [
11,
14] that the spontaneous chromosomal-doubling efficiency can be very high in a few maize genotypes compared to the doubling efficiency achieved by artificial chromosomal-doubling protocols. The use of spontaneous doubling can substantially reduce the labor costs, and accordingly, overall costs in DH-line production may be reduced by 20–50% [
10,
25]. However, as a high level of spontaneous doubling is limited to a few genotypes, breeding strategies need to be developed for using spontaneous doubling in DH-production pipelines. For this, estimating variance components and combining ability is important, and this is the first study reporting such estimates in the tropical inbred lines.
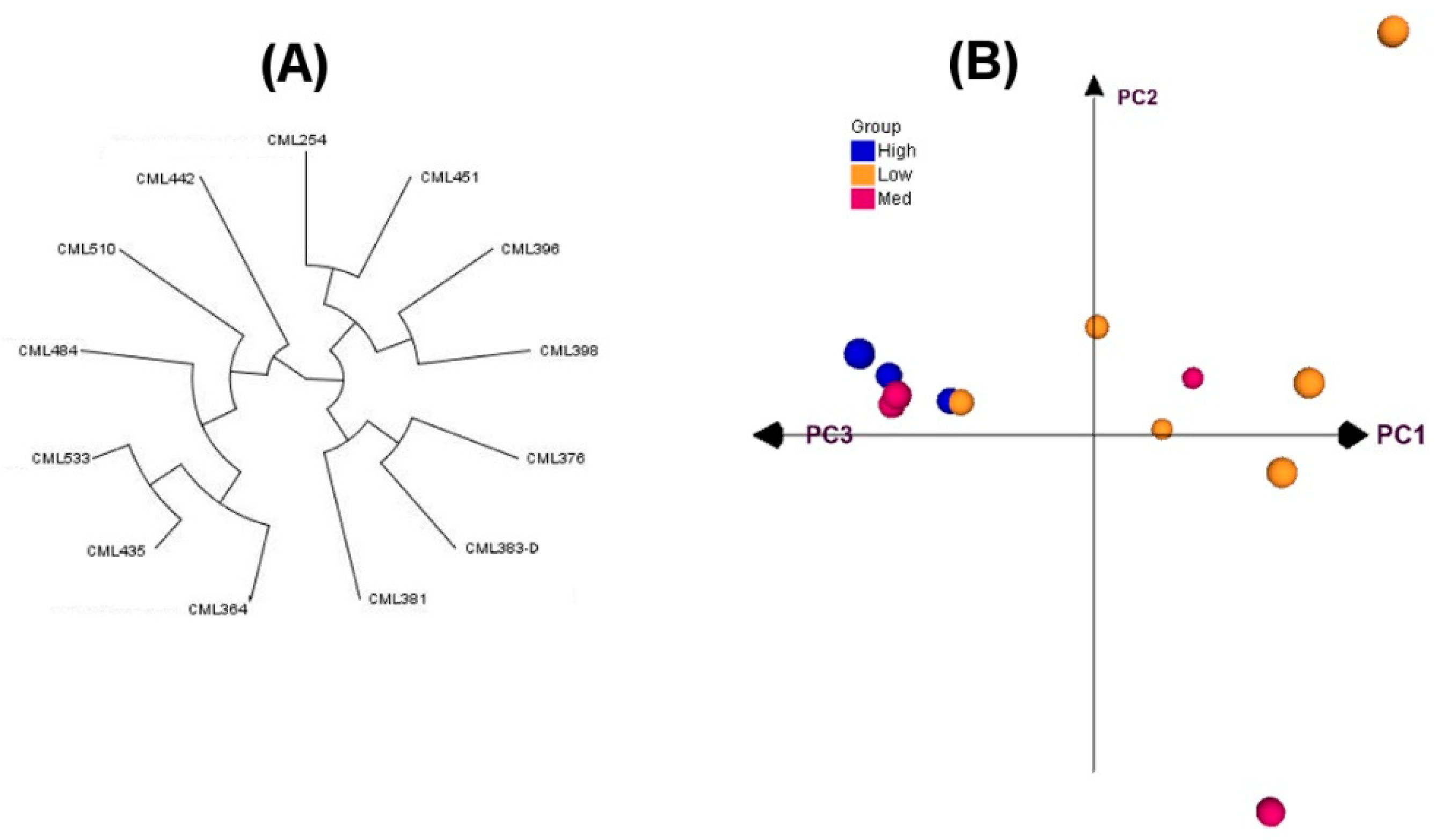
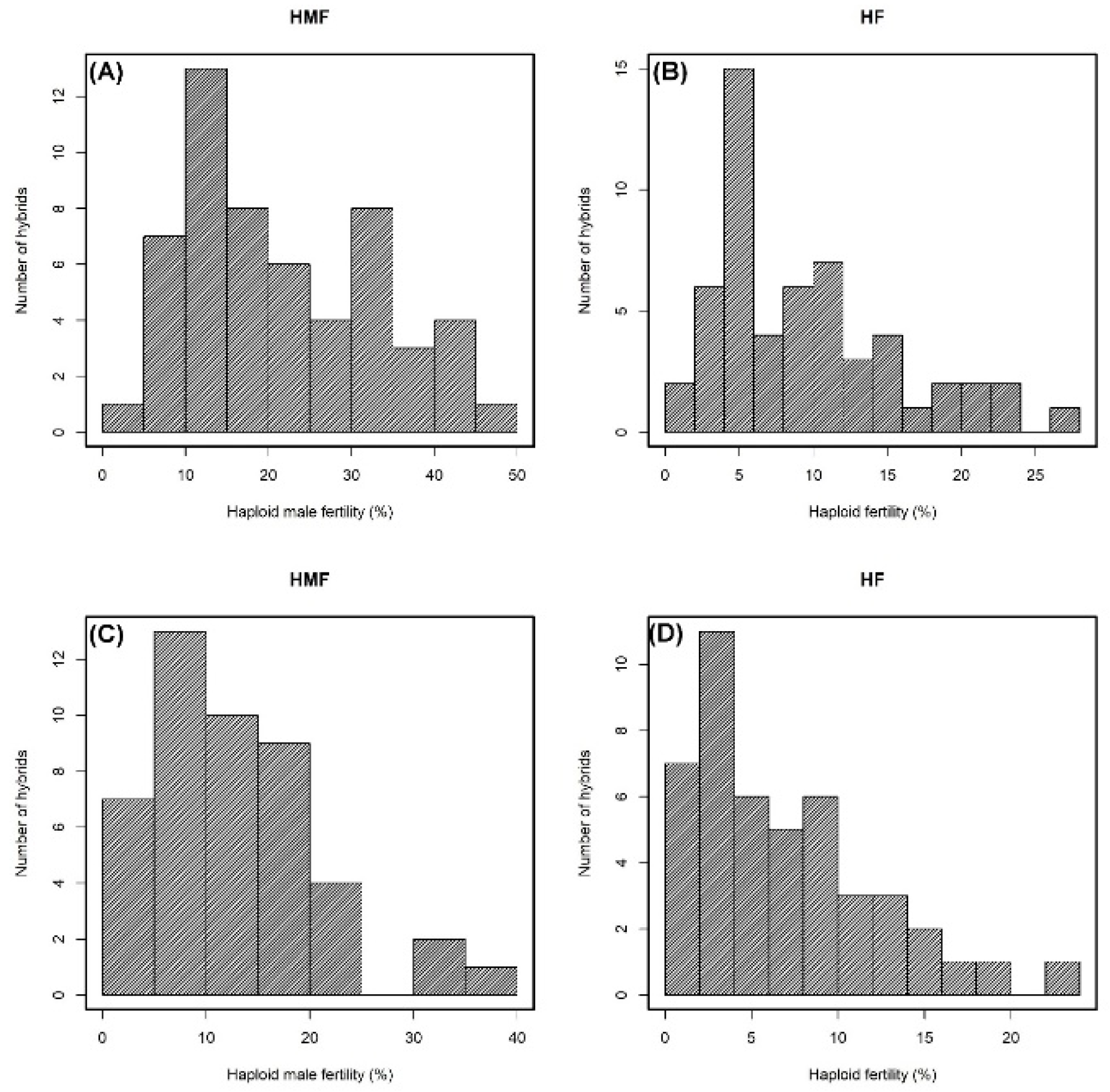
Two traits associated with spontaneous chromosomal doubling, namely, HMF and HF, showed greater variation in the inbred lines used in this study, and the data agree with a previous report on these lines [
11]. The significant genotype (hybrid) effect observed in ANOVA indicated considerable variability for HMF and HF among the F1 hybrids evaluated in this study. Observing significant GCA and SCA variances suggests that the performance of hybrids can be attributed simultaneously to both additive and non-additive genetic components. The environment had no significant effect on HF in both diallel experiments, while it strongly affected the HMF in the first diallel experiment. Fuente et al. [
24] also observed a significant environment effect for HMF. In both diallel experiments, non-significant GCA x environment-interaction variances were observed for both HMF and HF, indicating that the parental inbred lines do not perform differently in different environments. The significant SCA × environment-interaction variance for HMF observed in the second diallel experiment indicates the importance of non-additive genetic effects in this set of F1 crosses. The non-significant reciprocal variance and maternal variance for HF indicates that extra-nuclear factors are not important in the inheritance of HF. However, they seemed to play a role for HMF in the second diallel experiment.
The larger proportion of the mean squares of the GCA compared to SCA for HMF and HF suggests that the additive genetic effect was the main contributor to their inheritance in the F1 hybrids in the two evaluated environments. In a diallel experiment with a fixed-effects model, even though the estimates of additive and dominance variances are less important [
26], the GCA–SCA ratio (GSR) proposed by Baker [
27] (Baker’s ratio) is frequently used in several studies [
28,
29,
30] to determine the relative importance of GCA and SCA. If the Baker’s ratio is close to 1, then GCA is predominant for a given trait. The Baker’s ratios >0.7 for both the traits in both diallel experiments also indicated that GCA was relatively more important than SCA for both the traits. In addition, the higher additive genetic variance (σ
2A) compared to dominance variance (σ
2D) in both the experiments and higher estimates for the narrow-sense heritability over those for the broad-sense heritability also indicate the predominance of the additive effects for both HMF and HF in tropical maize.
All the F1 hybrids (excluding reciprocals) assessed in the two diallel experiments revealed a varied magnitude of SCA effects in different groups (
Table 6). In a group of hybrids derived from parents with a high rate of HMF, significant SCA effects were observed in a negative direction. This is probably one of the reasons for the low performance of hybrids derived from high x high HMF combinations, compared to their parents’ per se performance. Other possible reasons may be that HMF and HF are known to be controlled by a few genes with large effects, and these genes might have been fixed in the selected lines, which had a high performance of >65% for HMF and >26% for HF. Fuente et al. [
24] also observed that the F1 diallelic crosses show significantly less HMF compared to inbreds with high HMF in temperate maize germplasm. Contrary to the hybrids developed from lines with high and medium levels of HMF and HF, the hybrids developed with poor HMF- and HF-performing lines exhibited significant but positive SCA effects. Thus, non-additive effects play an important role in lines with poor HMF and HF performance. Non-additive effects generally include both dominance and epistatic effects. For the HMF and HF measured in haploid plants, the SCA effect might be partially determined by additive x additive epistatic effects only, as dominant effects are not possible due to a single copy of chromosomes in haploids [
24]. Even if the genome is doubled spontaneously at a very early stage, the duplicated chromosomes will be exact replicas of existing chromosomes, eliminating the possibility of dominant effects [
24]. Improving spontaneous doubling should be based on breeding strategies that exploit additive effects. The inbred lines CML364, CML435 and CML442 had the highest significant positive GCA effects for both HMF and HF, suggesting that these lines may harbor loci conferring a large positive effect and transmit such favorable alleles to their progenies through hybridization. These lines should also be useful as sources of QTLs for marker-assisted selection that would facilitate the transfer of alleles favorable for increased spontaneous doubling into commercial elite lines and hybrids.
The correlation between the mean parental values and F1-hybrid performance for HMF and HF in both the diallel experiments clearly indicates that merely knowing the per se performance is enough for predicting the performance of hybrids for HMF and HF with limited field experiments. Furthermore, by knowing the GCA effects of lines, the accuracy for finding the best possible combinations with high HMF and HF is also high (
Figure 3). These high correlations indicate the presence of a few genes controlling most of the variation of these two traits, as indicated by several genetic studies [
11,
14,
15,
16,
17,
18], and fixing these genes in lines could lead to high rates of spontaneous doubling. Focusing on a few genes for increasing spontaneous doubling also encourages the use of marker-assisted backcrossing to introgress these genes into other elite lines. The possibility of eliminating or reducing chemical-based chromosomal doubling in the process of DH-line development could ultimately save resources for maize-breeding programs.
5. Data Analysis
The statistical analysis was carried out using the AGD-R (Analysis of Genetic Designs with R for Windows) Version 5.0 statistical software [
37]. Analysis of variance (ANOVA) was performed using the following statistical model:
where
yijkm represents the phenotypic performance of the
ijth genotype (parental line
i =
j, or hybrid
i≠
j) in the
mth replication in the
kth environment,
µ is an intercept term,
gij is the genetic effect of the
ijth genotype (parental line
i =
j, or hybrid
i≠
j),
lk is the effect of the kth environment,
(gl)ijk is the interaction of the
ijth genotype (parental line
i =
j, or hybrid
i≠
j) with the
kth environment,
rmk is the effect of the
mth replication in the
kth environment, and
eijkm is the residual error term.
The diallel analysis was carried out by applying Griffing’s Model I (fixed model) Method 1 (parents and hybrids with reciprocal crosses) [
23]. The mean squares for GCA and SCA were tested against their respective error variances inferred from ANOVA using the SAS software program version 9.4 [
38]. The variance was partitioned into GCA, SCA, reciprocal, maternal and nonmaternal components and their interactions with years. To test the mean squares for the GCA, SCA, reciprocal, maternal and nonmaternal components for significance in the two-location analysis, the interaction between locations and the corresponding component was used as the error term for
F tests. Estimates of the GCA and SCA effects were calculated, and their significance was determined by
t tests. Further variance-components estimates were computed for GCA, SCA and reciprocal (REC). All the calculations were performed using the AGD-R software. Pairwise Pearson’s correlation coefficients (r) were calculated between the F1 hybrids (F1P) and mid-parent values (MPV), r(MPV: F1P). Furthermore, we also tested the Pearson’s correlation of F1P with the sum of the GCA effects of both the parents, r(GCA: F1P), for each diallel experiment.

 ,
, 




{kind=link}
{kind=link}
{kind=link}