Retinoic Acid, under Cerebrospinal Fluid Control, Induces Neurogenesis during Early Brain Development

Abstract
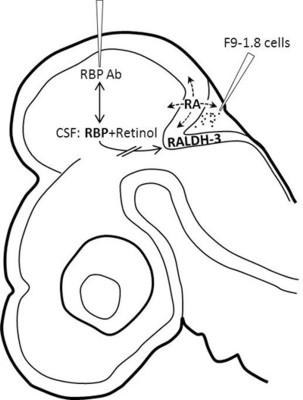
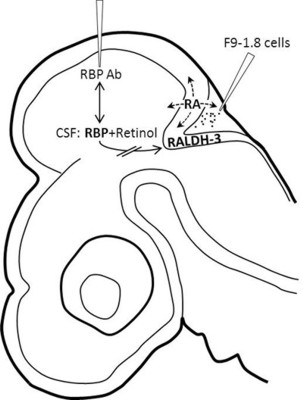
:
{kind=link}
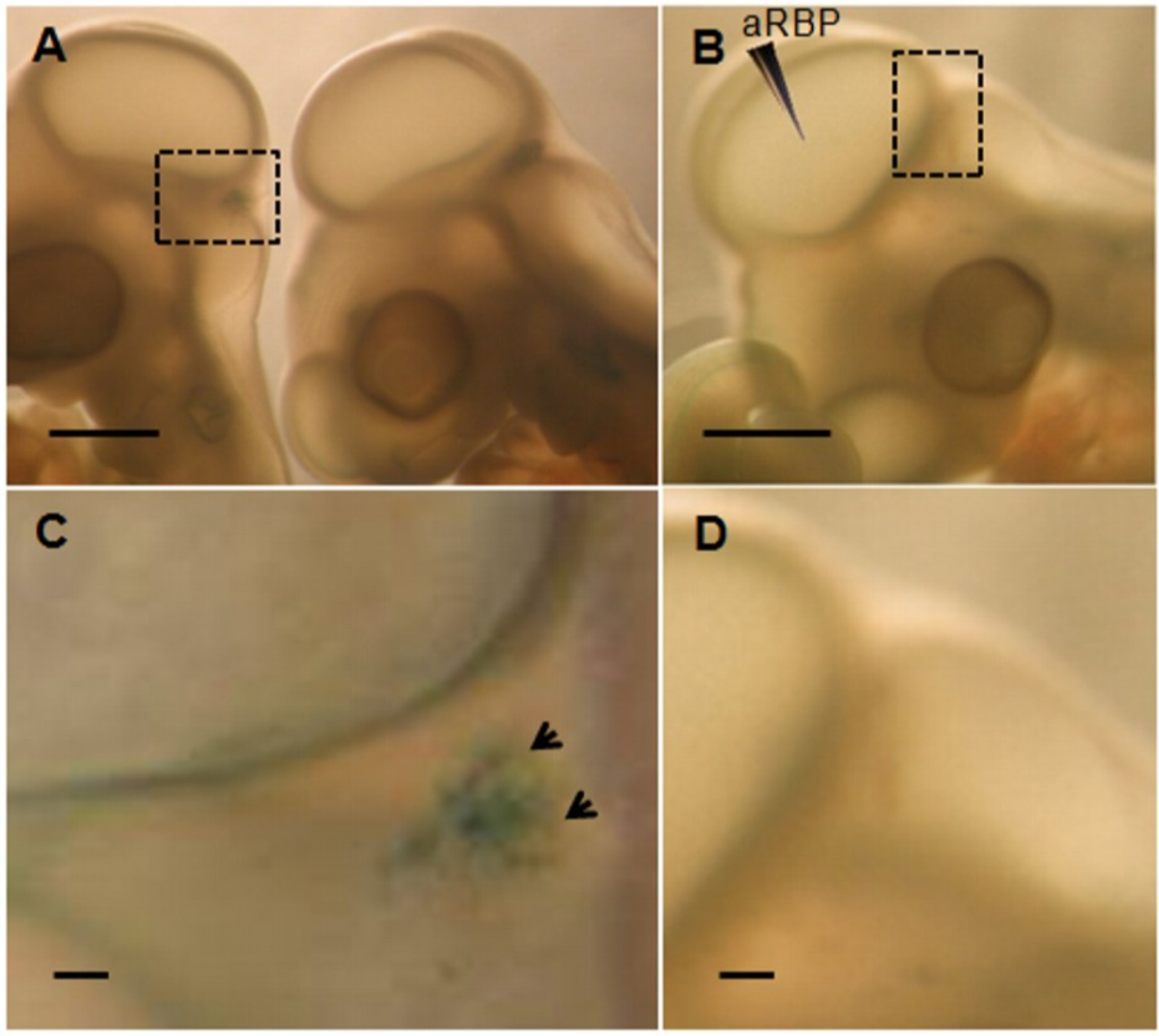
{kind=link}
{kind=link}
{kind=link}
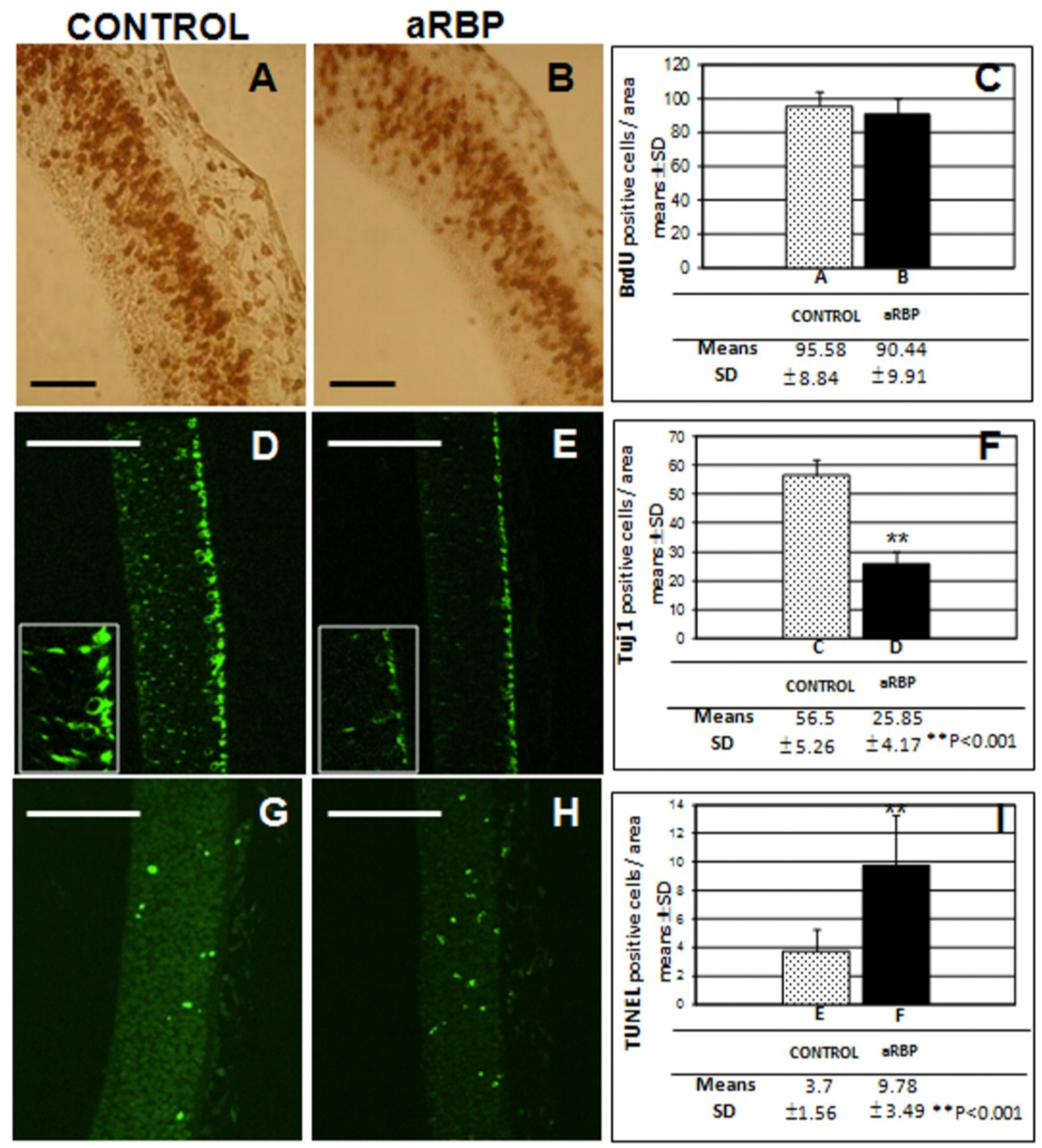{kind=link}
1. Introduction
2. Experimental Section
2.1. Obtaining Embryos and Cerebrospinal Fluid
2.2. Culture of F9-1.8 Cell Line
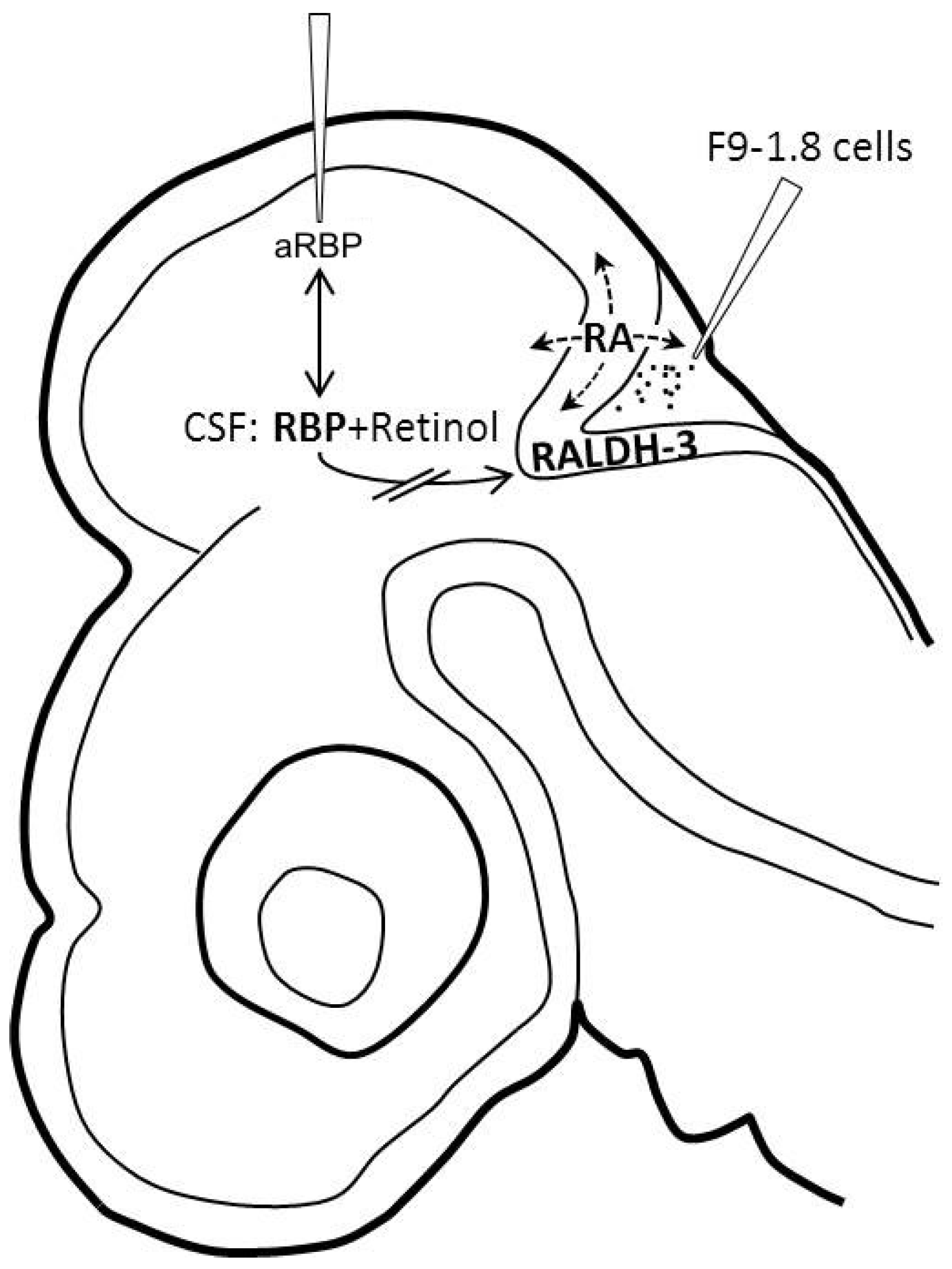
2.3. Micro-Injection of F.9-1.8 Cells “in Vivo”
2.4. Neuroepithelial Cell Behavior Test
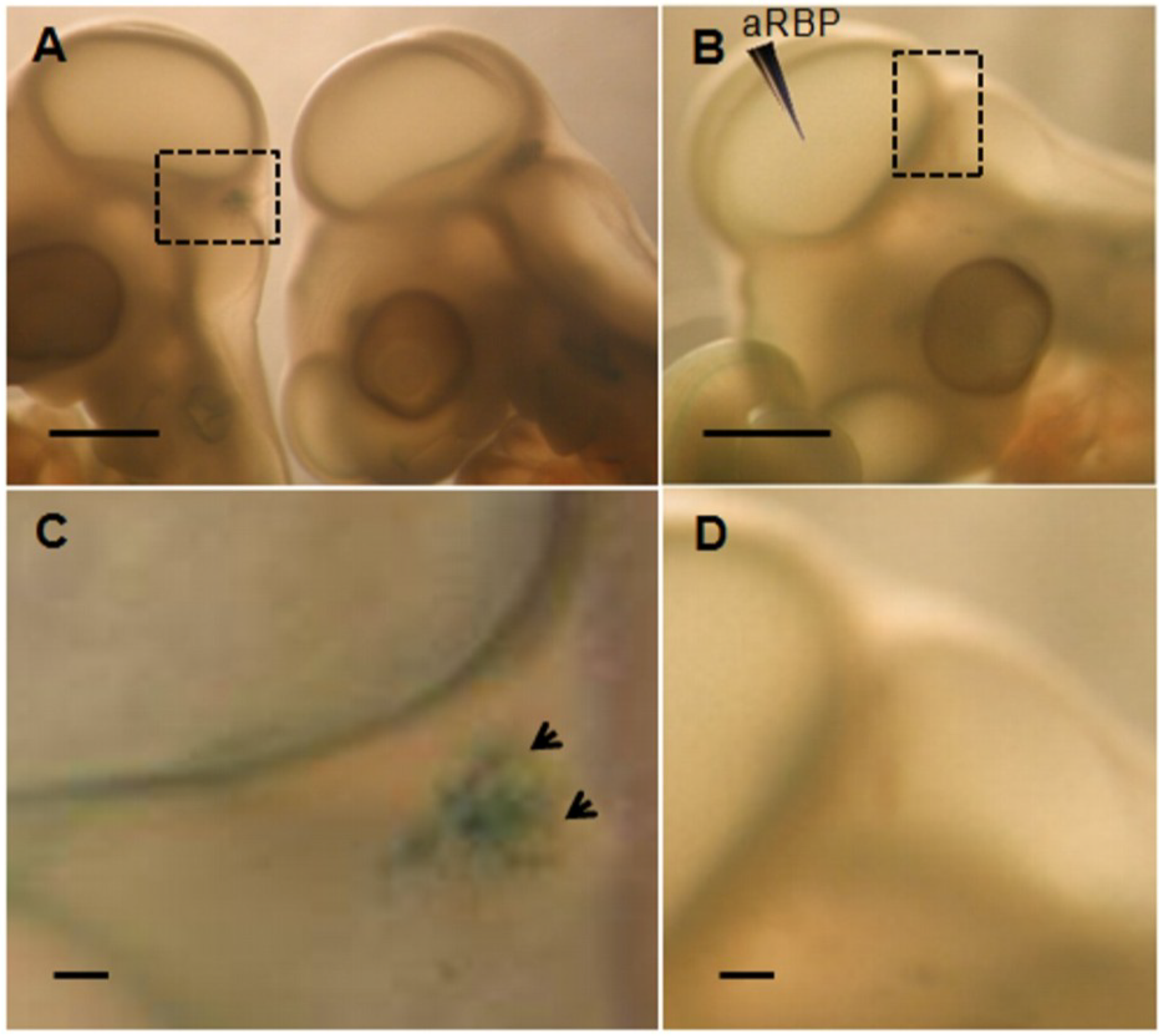
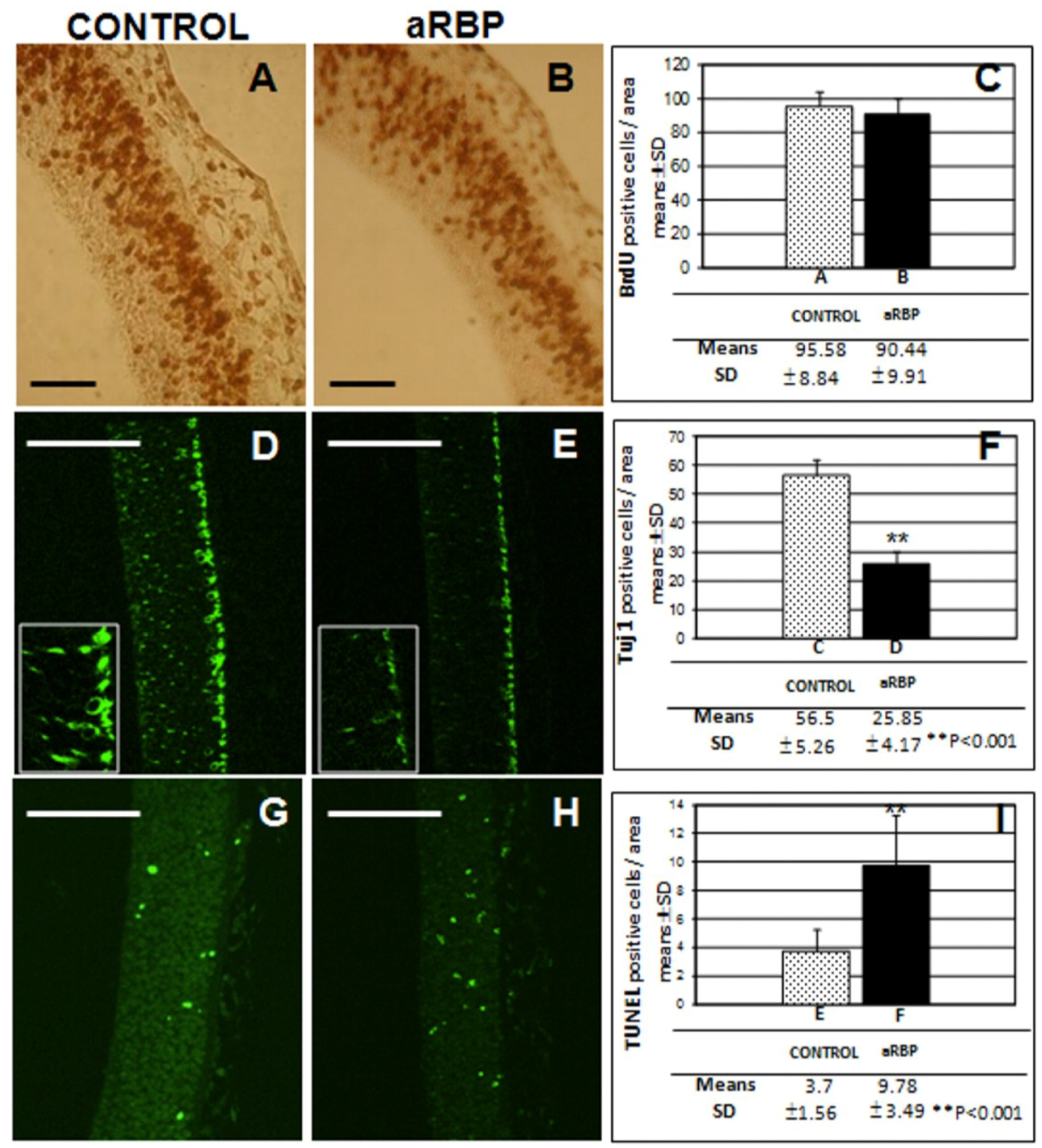
2.4.1. Determination of BrdU
2.4.2. Neurogenesis
2.4.3. TUNEL Assay
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Checiu, I.; Prelipceanu, O.; Popescu, O. The role of the cerebrospinal fluid during embryonic development. A biochemical study. Morphol. Embryol. (Bucur.) 1984, 30, 243–250. [Google Scholar]
- Desmond, M.E.; Jacobson, A.G. Embryonic brain enlargement requires cerebrospinal fluid pressure. Dev. Biol. 1977, 57, 188–198. [Google Scholar] [CrossRef]
- Dziegielewska, K.M.; Evans, C.A.; Lai, P.C.; Lorscheider, F.L.; Malinowska, D.H.; Møllgård, K.; Saunders, N.R. Proteins in cerebrospinal fluid and plasma of fetal rats during development. Dev. Biol. 1981, 83, 193–200. [Google Scholar] [CrossRef]
- Desmond, M.E.; Levitan, M.L. Brain expansion in the chick embryo initiated by experimentally produced occlusion of the spinal neurocoel. Anat. Rec. 2002, 268, 147–159. [Google Scholar] [CrossRef]
- Gato, A.; Moro, J.A.; Alonso, M.I.; Bueno, D.; De la Mano, A.; Martin, C. Embryonic cerebrospinal fluid regulates neuroepithelial survival, proliferation, and neurogenesis in chick embryos. Anat. Rec. A 2005, 284A, 475–484. [Google Scholar] [CrossRef]
- Miyan, J.A.; Nabiyouni, M.; Zendah, M. Development of the brain: A vital role for cerebrospinal fluid. Can. J. Physiol. Pharmacol. 2003, 81, 317–328. [Google Scholar] [CrossRef]
- Gato, A.; Desmond, M.E. Why the embryo still matters: CSF and the neuroepithelium as interdependent regulators of embryonic brain growth, morphogenesis and histiogenesis. Dev. Biol. 2009, 327, 263–272. [Google Scholar] [CrossRef]
- Zappaterra, M.W.; Lehtinen, M.K. The cerebrospinal fluid: Regulator of neurogenesis, behavior, and beyond. Cell. Mol. Life Sci. 2012, 69, 2863–2878. [Google Scholar] [CrossRef]
- Mashayekhi, F.; Draper, C.E.; Bannister, C.M.; Pourghasem, M.; Owen-Lynch, P.J.; Miyan, J.A. Deficient cortical development in the hydrocephalic Texas (H-Tx) rat: A role for CSF. Brain 2002, 125, 1859–1874. [Google Scholar] [CrossRef]
- Owen-Lynch, P.J.; Draper, C.E.; Mashayekhi, F.; Bannister, C.M.; Miyan, J.A. Defective cell cycle control underlies abnormal cortical development in the hydrocephalic Texas rat. Brain 2003, 126, 623–631. [Google Scholar] [CrossRef]
- Parada, C.; Martín, C.; Alonso, M.I.; Moro, J.A.; Bueno, D.; Gato, A. Embryonic cerebrospinal fluid collaborates with the isthmic organizer to regulate mesencephalic gene expression. J. Neurosci. Res. 2005, 82, 333–345. [Google Scholar] [CrossRef]
- Gato, A.; Martin, P.; Alonso, M.I.; Martin, C.; Pulgar, M.A.; Moro, J.A. Analysis of cerebro-spinal fluid protein composition in early developmental stages in chick embryos. J. Exp. Zool. A 2004, 301A, 280–289. [Google Scholar] [CrossRef]
- Parada, C.; Gato, A.; Bueno, D. Mammalian embryonic cerebrospinal fluid proteome has greater apolipoprotein and enzyme pattern complexity than the avian proteome. J. Proteome Res. 2005, 4, 2420–2428. [Google Scholar] [CrossRef]
- Parada, C.; Gato, A.; Aparicio, M.; Bueno, D. Proteome analysis of chick embryonic cerebrospinal fluid. Proteomics 2006, 6, 312–320. [Google Scholar] [CrossRef]
- Zappaterra, M.D.; Lisgo, S.N.; Lindsay, S.; Gygi, S.P.; Walsh, C.A.; Ballif, B.A. A comparative proteomic analysis of human and rat embryonic cerebrospinal fluid. J. Proteome Res. 2007, 6, 3537–3548. [Google Scholar] [CrossRef]
- Martin, C.; Bueno, D.; Alonso, M.I.; Moro, J.A.; Callejo, S.; Parada, C.; Martin, P.; Carnicero, E.; Gato, A. FGF2 plays a key role in embryonic cerebrospinal fluid trophic properties over chick embryo neuroepithelial stem cells. Dev. Biol. 2006, 297, 402–416. [Google Scholar] [CrossRef]
- Goncalves, M.B.; Boyle, J.; Webber, D.J.; Hall, S.; Minger, S.L.; Corcoran, J.P. Timing of the retinoid-signalling pathway determines the expression of neuronal markers in neural progenitor cells. Dev. Biol. 2005, 278, 60–70. [Google Scholar] [CrossRef]
- Jacobs, S.; Lie, D.C.; DeCicco, K.L.; Shi, Y.; DeLuca, L.M.; Gage, F.H.; Evans, R.M. Retinoic acid is required early during adult neurogenesis in the dentate gyrus. Proc. Nat. Acad. Sci. USA 2006, 103, 3902–3907. [Google Scholar]
- Wang, H.F.; Liu, F.C. Regulation of multiple dopamine signal transduction molecules by retinoids in the developing striatum. Neuroscience 2005, 134, 97–105. [Google Scholar] [CrossRef]
- Reijntjes, S.; Blentic, A.; Gale, E.; Maden, M. The control of morphogen signalling: regulation of the synthesis and catabolism of retinoic acid in the developing embryo. Dev. Biol. 2005, 285, 224–237. [Google Scholar] [CrossRef]
- Maden, M. Retinoic acid and limb regeneration—A personal view. Int. J. Dev. Biol. 2002, 46, 883–886. [Google Scholar]
- McCaffery, P.; Drager, U.C. Regulation of retinoic acid signaling in the embryonic nervous system: a master differentiation factor. Cytokine Growth Factor Rev. 2000, 11, 233–249. [Google Scholar] [CrossRef]
- Blentic, A.; Gale, E.; Maden, M. Retinoic acid signalling centres in the avian embryo identified by sites of expression of synthesising and catabolising enzymes. Dev. Dyn. 2003, 227, 114–127. [Google Scholar] [CrossRef]
- Parada, C.; Gato, A.; Bueno, D. All-trans retinol and retinol-binding protein from embryonic cerebrospinal fluid exhibit dynamic behaviour during early central nervous system development. Neuroreport 2008, 19, 945–950. [Google Scholar] [CrossRef]
- Martin, C.; Alonso, M.I.; Santiago, C.; Moro, J.A.; De la Mano, A.; Carretero, R.; Gato, A. Early embryonic brain development in rats requires the trophic influence of cerebrospinal fluid. Int. J. Dev. Neurosci. 2009, 27, 733–740. [Google Scholar] [CrossRef]
- Duester, G. Families of retinoid dehydrogenases regulating vitamin A function: Production of visual pigment and retinoic acid. Eur. J. Biochem. 2000, 267, 4315–4324. [Google Scholar] [CrossRef]
- Begemann, G.; Meyer, A. Hindbrain patterning revisited: Timing and effects of retinoic acid signalling. Bioessays 2001, 23, 981–986. [Google Scholar] [CrossRef]
- Clotman, F.; VanMaeleFabry, G.; Picard, J.J. Retinoic acid induces a tissue-specific deletion in the expression domain of Otx2. Neurotoxicol. Teratol. 1997, 19, 163–169. [Google Scholar] [CrossRef]
- Diez del Corral, R.; Storey, K.G. Markers in vertebrate neurogenesis. Nat. Rev. Neurosci. 2001, 2, 835–839. [Google Scholar]
- Diez del Corral, R.; Storey, K.G. Opposing FGF and retinoid pathways: A signalling switch that controls differentiation and patterning onset in the extending vertebrate body axis. Bioessays 2004, 26, 857–869. [Google Scholar] [CrossRef]
- Kolm, P.J.; Apekin, V.; Sive, H. Xenopus hindbrain patterning requires retinoid signaling. Dev. Biol. 1997, 192, 1–16. [Google Scholar] [CrossRef]
- Kudoh, T.; Wilson, S.W.; Dawid, I.B. Distinct roles for Fgf, Wnt and retinoic acid in posteriorizing the neural ectoderm. Development 2002, 129, 4335–4346. [Google Scholar]
- McCaffery, P.J.; Adams, J.; Maden, M.; Rosa-Molinar, E. Too much of a good thing: Retinoic acid as an endogenous regulator of neural differentiation and exogenous teratogen. Eur. J. Neurosci. 2003, 18, 457–472. [Google Scholar] [CrossRef]
- Alonso, M.I.; Martin, C.; Carnicero, E.; Bueno, D.; Gato, A. Cerebrospinal fluid control of neurogenesis induced by retinoic acid during early brain development. Dev. Dyn. 2011, 240, 1650–1659. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. 1951. Dev. Dyn. 1992, 195, 231–272. [Google Scholar] [CrossRef]
- Schulz, W.A.; Gais, G. Constitutive c-myc expression enhances proliferation of differentiating F9 teratocarcinoma cells. Biochim. Biophys. Acta 1989, 1013, 125–132. [Google Scholar]
- Sonneveld, E.; van den Brink, C.E.; van der Leede, B.J.; Maden, M.; van der Saag, P.T. Embryonal carcinoma cell lines stably transfected with mRARbeta2-lacZ: Sensitive system for measuring levels of active retinoids. Exp. Cell. Res. 1999, 250, 284–297. [Google Scholar] [CrossRef]
- Nakamura, H.; Watanabe, Y. Isthmus organizer and regionalization of the mesencephalon and metencephalon. Int. J. Dev. Biol. 2005, 49, 231–235. [Google Scholar] [CrossRef]
- Wilson, L.J.; Myat, A.; Sharma, A.; Maden, M.; Wingate, R.J. Retinoic acid is a potential dorsalising signal in the late embryonic chick hindbrain. BMC Dev. Biol. 2007, 7. [Google Scholar] [CrossRef]
- Chatzi, C.; Cunningham, T.J.; Duester, G. Investigation of retinoic acid function during embryonic brain development using retinaldehyde-rescued Rdh10 knockout mice. Dev. Dyn. 2013, 242, 1056–1065. [Google Scholar] [CrossRef]
- Maden, M. Retinoic acid in the development, regeneration and maintenance of the nervous system. Nat. Rev. Neurosci. 2007, 8, 755–765. [Google Scholar] [CrossRef]
- Haskell, G.T.; LaMantia, A.S. Retinoic acid signaling identifies a distinct precursor population in the developing and adult forebrain. J. Neurosci. 2005, 25, 7636–7647. [Google Scholar] [CrossRef]
- Merkle, F.T.; Alvarez-Buylla, A. Neural stem cells in mammalian development. Curr. Opin. Cell. Biol. 2006, 18, 704–709. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Alonso, M.I.; Carnicero, E.; Carretero, R.; De la Mano, A.; Moro, J.A.; Lamus, F.; Martín, C.; Gato, A. Retinoic Acid, under Cerebrospinal Fluid Control, Induces Neurogenesis during Early Brain Development. J. Dev. Biol. 2014, 2, 72-83. https://doi.org/10.3390/jdb2020072
Alonso MI, Carnicero E, Carretero R, De la Mano A, Moro JA, Lamus F, Martín C, Gato A. Retinoic Acid, under Cerebrospinal Fluid Control, Induces Neurogenesis during Early Brain Development. Journal of Developmental Biology. 2014; 2(2):72-83. https://doi.org/10.3390/jdb2020072
Chicago/Turabian StyleAlonso, M. Isabel, Estela Carnicero, Raquel Carretero, Aníbal De la Mano, Jose Antonio Moro, Francisco Lamus, Cristina Martín, and Angel Gato. 2014. "Retinoic Acid, under Cerebrospinal Fluid Control, Induces Neurogenesis during Early Brain Development" Journal of Developmental Biology 2, no. 2: 72-83. https://doi.org/10.3390/jdb2020072