Genetic Alterations in the INK4a/ARF Locus: Effects on Melanoma Development and Progression


1
Faculty of Medicine, Health and Human Sciences, Macquarie University, Sydney, NSW 2109, Australia
2
Melanoma Institute Australia, Sydney, NSW 2065, Australia
*
Author to whom correspondence should be addressed.
Biomolecules 2020, 10(10), 1447; https://doi.org/10.3390/biom10101447
Submission received: 1 September 2020
/
Revised: 11 October 2020
/
Accepted: 12 October 2020
/
Published: 15 October 2020
(This article belongs to the Special Issue Deciphering alternative functions of the INK4a/ARF locus)
Abstract
:Genetic alterations in the INK4a/ARF (or CDKN2A) locus have been reported in many cancer types, including melanoma; head and neck squamous cell carcinomas; lung, breast, and pancreatic cancers. In melanoma, loss of function CDKN2A alterations have been identified in approximately 50% of primary melanomas, in over 75% of metastatic melanomas, and in the germline of 40% of families with a predisposition to cutaneous melanoma. The CDKN2A locus encodes two critical tumor suppressor proteins, the cyclin-dependent kinase inhibitor p16INK4a and the p53 regulator p14ARF. The majority of CDKN2A alterations in melanoma selectively target p16INK4a or affect the coding sequence of both p16INK4a and p14ARF. There is also a subset of less common somatic and germline INK4a/ARF alterations that affect p14ARF, while not altering the syntenic p16INK4a coding regions. In this review, we describe the frequency and types of somatic alterations affecting the CDKN2A locus in melanoma and germline CDKN2A alterations in familial melanoma, and their functional consequences in melanoma development. We discuss the clinical implications of CDKN2A inactivating alterations and their influence on treatment response and resistance.
1. Introduction
Melanoma is an aggressive and highly metastatic form of skin cancer that causes nearly 60,000 deaths globally each year [1]. Melanoma originates from neural-crest-derived pigment-producing cells known as melanocytes. These specialized, dendritic cells are predominantly located in the basal layer of the epidermis, and are also found in the vascular uvea of the eye and in mucosal membranes [2]. The transformation of melanocytes into melanoma, termed melanomagenesis, involves the sequential selection of genetic and epigenetic alterations that promote proliferation, invasion, and immune escape [3,4].
Cutaneous melanoma has the highest median coding mutation rate of any cancer type (14.4 coding mutations per megabase) and this reflects the high proportion of ultraviolet (UV)-induced C>T substitutions that occur at pyrimidines [5,6]. The high mutation burden in melanoma is also associated with a high rate of silent passenger mutations and this is consistent with the 2:1 ratio of non-synonymous to synonymous mutations in melanoma [7]. The systematic evaluation of over 500 melanoma genomes in the last decade has identified a series of frequently and significantly altered oncogenes and tumor suppressor genes, including BRAF, NRAS, RAC1, NF1, CDKN2A, PTEN, and ARID2 [7,8,9].
The CDKN2A locus is located on chromosome band 9p21 and is also referred to as the INK4a/ARF locus. This is one of the most commonly altered sequences in cancer and is mutated, deleted, or methylated in 40–70% of sporadic melanomas [8]. Germline alterations in the CDKN2A locus have also been identified in approximately 40% of high-risk melanoma families with three or more melanoma cases [10]. The frequent alteration of CDKN2A reflects its capacity to encode two distinct tumor suppressor proteins, p16INK4a and p14ARF, which are translated in alternate reading frames, from alternatively spliced transcripts with independent start sites and unique first exons (exon 1α for p16INK4a and exon 1ß for p14ARF) [11]. p16INK4a forms binary complexes with the cyclin-dependent kinases 4 and 6 (CDK4/6) to inhibit cyclin D-CDK4/6-mediated phosphorylation of the retinoblastoma protein and, thus, prevents G1 to S phase cell cycle transition [12,13,14]. The functions of p14ARF are more complex, but it plays a central role in the stabilization and activation of p53 via the inhibition of the major p53 negative regulator MDM2 [15,16,17,18]. Despite sharing largely overlapping DNA sequences, the functional impact of CDKN2A alterations is complex and can be difficult to predict.
In this review, we examine genetic alterations affecting the CDKN2A locus in melanoma, the functional impact of altered CDKN2A, and the contribution of this locus to melanoma development and progression. We also discuss the impact of CDKN2A on the response of melanoma patients to current therapies.
2. Somatic CDKN2A Alterations in Melanoma
Inactivation of the CDKN2A locus has been detected in approximately 50% of primary melanomas and over 75% of metastatic melanomas [7,8,9]. The majority of CDKN2A alterations in melanoma selectively target p16INK4a or affect the coding sequences of both p16INK4a and p14ARF. Somatic CDKN2A alterations that solely affect p14ARF without compromising the syntenic p16INK4a coding region are rare. For example, analysis of the mutation spectrum of primary and metastatic cutaneous melanoma in The Cancer Genome Atlas (TCGA) (n = 363, data derived through the Memorial Sloan Kettering Cancer Center cBioPortal for Cancer Genomics [19,20] identified CDKN2A genetic alterations in approximately 45% (162/363) of melanoma cases; these included missense mutations (21/363, 5.8%), truncating mutations (34/363, 9.4%), in-frame deletions (2/363, 0.6%), amplifications (1/363, 0.3%), and homozygous deletions (112/363, 30.8%). Almost 43% of the missense, truncating, and in-frame mutations specifically affected p16INK4a, 57% affected both p14ARF and p16INK4a, and mutations that altered p14ARF alone were not observed. It is worth noting that alterations that appear to specifically alter one of the CDKN2A-encoded proteins may impact the co-ordinated regulation of p14ARF and/or p16INK4a [21]. These mutation frequencies are consistent with previous reports [22,23] and confirm that homozygous CDKN2A deletions are the most common alterations affecting this locus in melanoma (reviewed in reference [24]).
Promoter hypermethylation of CDKN2A leading to transcription silencing and loss of protein expression increases during melanoma progression and has been identified in approximately 19–60% of vertical-growth-phase melanomas, 40% of radial-growth-phase melanomas, and 25–33% of melanoma metastases [25,26,27,28]. Approximately 20–30% of melanomas show loss of p16INK4a due to promoter hypermethylation [23,27], and although p14ARF gene methylation is less studied, one study found that the p14ARF promoter was hypermethylated in 34/60 (57%) melanoma metastases. In this study, only 16/60 (27%) metastases displayed p16INK4a promoter methylation and seven of these tumors showed concurrent methylation on both the p16INK4a and p14ARF promoters [23]. Intriguingly, the epigenetic modifications affecting p14ARF and p16INK4a may vary in melanoma, with 5’ CpG promoter hypermethylation reported to be predominant in p16INK4a gene inactivation, whereas histone hypoacetylation is more commonly associated with p14ARF gene silencing [29]. In the TCGA cohort, methylation of three CpG islands in the p16INK4a exon 1α [cg12840719, cg13601799, cg04026675] and five CpG islands in p14ARF exon 1β [cg03079681, cg07562918, cg00718440, cg10848754, cg14430974] were analyzed. Although methylation levels (ß-value) were highly variable across the CDKN2A locus, the three p16INK4a CpG islands appeared to be the most highly methylated in the sequence (Figure 1).
Histone methylation also regulates CDKN2A transcription. The trimethylation of histone H3 at lysine 27 (H3K27me3) along the CDKN2A locus is induced by EZH2, the catalytic subunit of the polycomb repressive complex (PRC) 2. The co-operative binding of PRC1 (including the BMI1 subunit) and the long noncoding RNA ANRIL to H3K27me3 compacts the CDKN2A genomic locus and represses transcription [25,30,31,32], in particular the transcription of p16INK4a [33]. In line with these data, the levels of H3K27me3 increase from primary to metastatic melanoma, and high H3K27me3 is an independent poor prognostic marker in melanoma [34,35].
Mutations in the BRAF, NRAS, and NF1 genes are the predominant drivers in cutaneous melanoma, and CDKN2A genetic alterations are distributed evenly amongst these genetic melanoma subtypes, with CDKN2A genetic alterations detected in 48.9% (92/188) of BRAF-mutant, 44.7% (51/114) of NRAS-mutant, and 50% (33/66) of NF1-mutant cutaneous melanomas (Table 1) [8]. In contrast, CDKN2A promoter hypermethylation occurs most frequently in NRAS-mutant melanoma (87.7%) and is less common in melanoma with BRAF mutations (67.0% with CDKN2A promoter methylation) and in melanomas with NF1 mutations (77.3% showing CDKN2A promoter methylation) (Table 1). Analysis of the TCGA cutaneous melanoma dataset also showed that methylation of the p16INK4a probe cg12840719 was higher in NRAS-mutant melanoma, compared to BRAF-mutant melanoma, although this was not statistically significant (Figure 1C, Table 1). Interestingly CDKN2A methylation was rarely detected in sporadic primary melanoma and has not been identified in melanocytic nevi [36,37]. Altogether, CDKN2A alterations (including mutation, deletion, and promoter hypermethylation) appear to be differentially distributed across the cutaneous melanoma genomic subtypes, and have been identified in 58% of BRAF-mutant, 72% of NRAS-mutant, and 71% of NF1-mutant melanoma but only in 37% of triple wild-type melanomas [8].
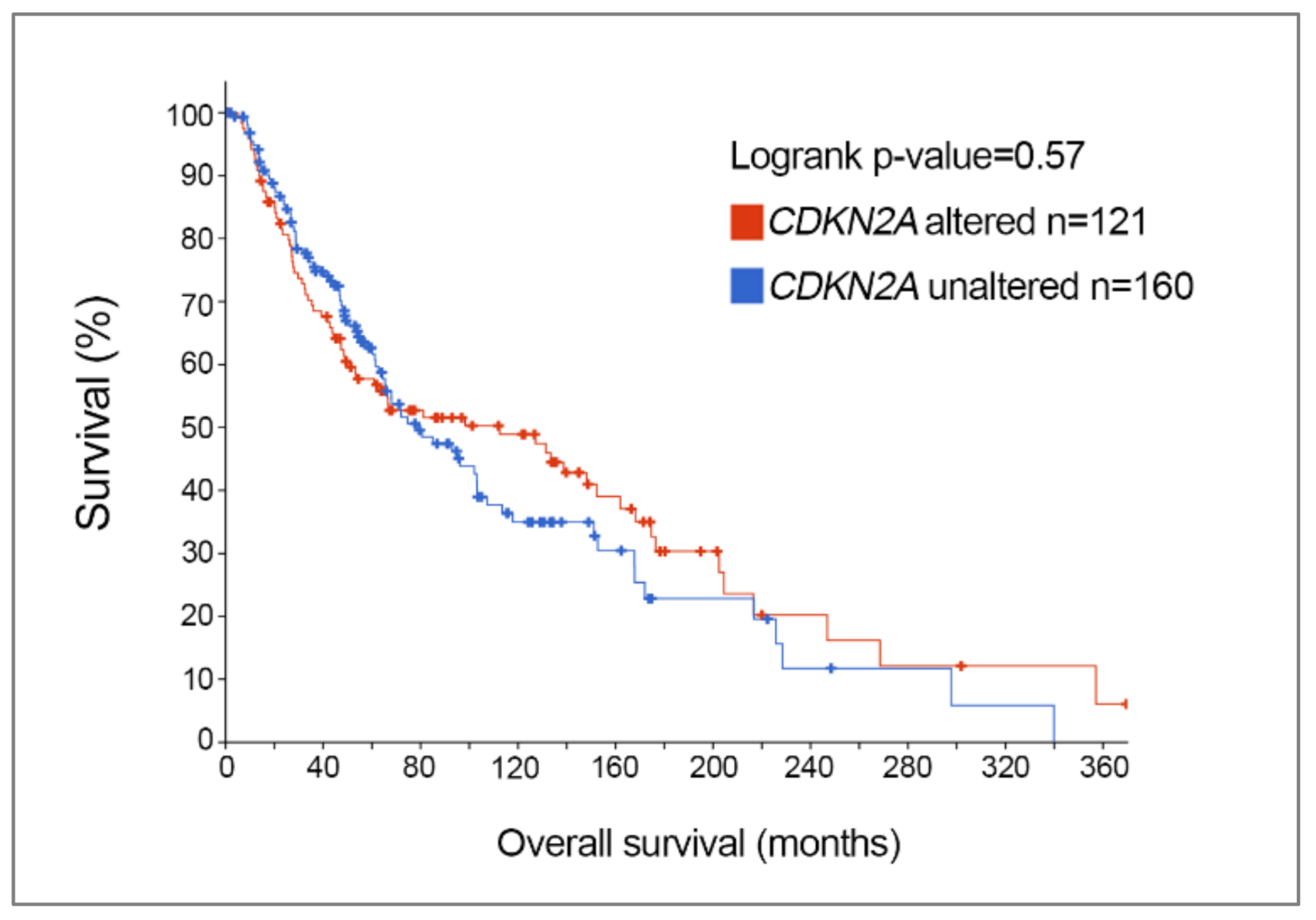
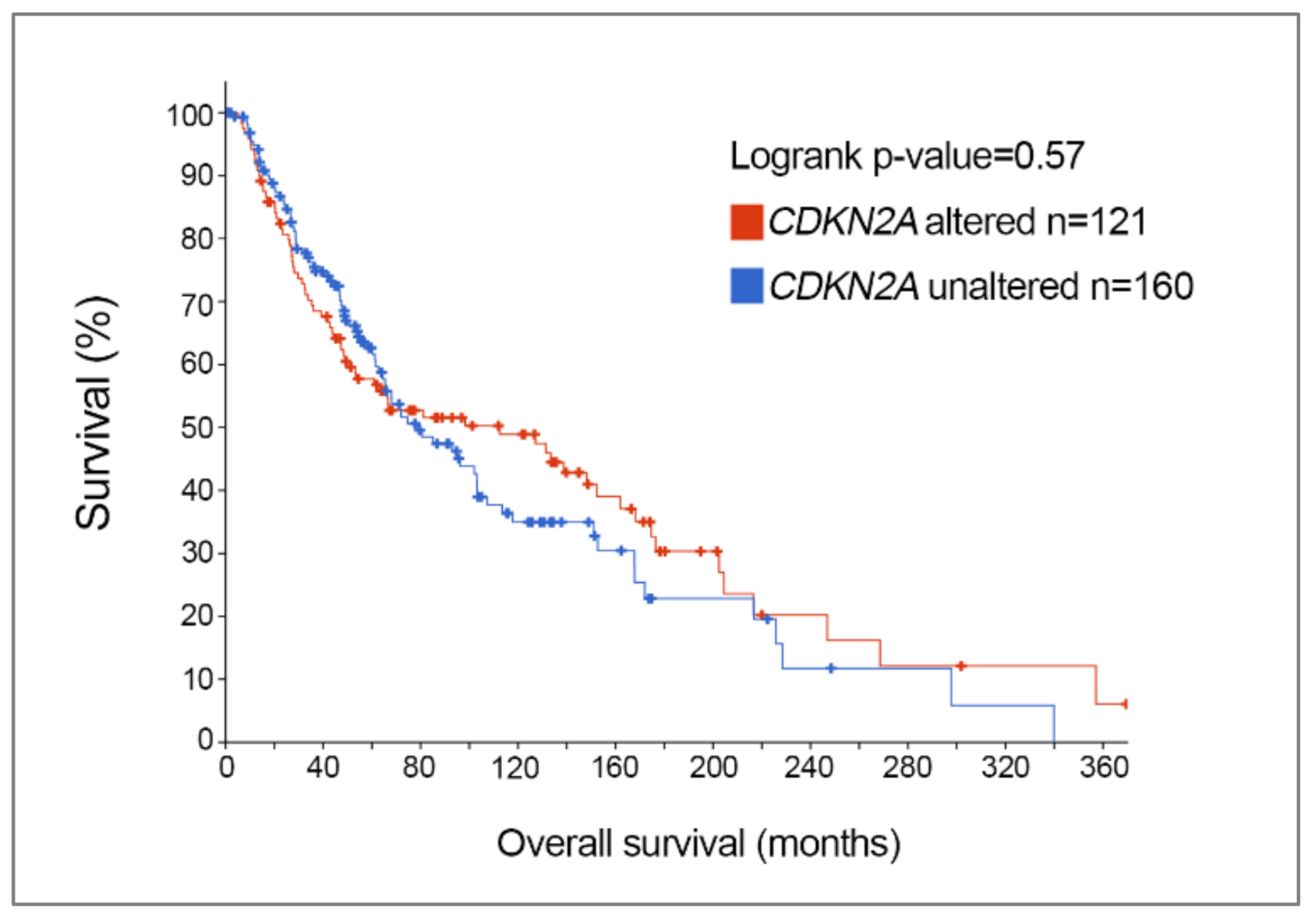
Further analysis of TCGA skin cutaneous melanoma dataset revealed that the frequency of CDKN2A genetic alterations was not influenced by gender; 63/135 (47%) females and 98/228 (43%) males showed melanoma-associated CDKN2A genetic changes, and this is in keeping with a previous report [38]. Melanoma patients diagnosed at a younger age tend to have a higher frequency of CDKN2A somatic alterations, however, with 72% of patients aged 30 years or younger displaying melanoma-associated CDKN2A alterations, and this diminishes with increasing age. In particular, 50% of melanoma patients aged 31–50, 41% of patients aged 51–70, and 36% of patients aged > 71 had somatic CDKN2A genetic changes [8]. Further, CDKN2A mutation status is not associated with overall survival in melanoma; melanoma patients with somatic CDKN2A alterations had a median overall survival of 112 months compared to a median overall survival of 79 months in patients with wild-type CDKN2A [8] (Figure 2).
CDKN2A is also highly mutated in rarer subtypes of cutaneous melanoma and in non-cutaneous melanoma. For instance, CDKN2A mutations are present in 20% (4 out of 20 cases; 3 truncating mutations and one missense mutation) of desmoplastic melanoma, and these alterations predominantly affected either p16INK4a alone or p16INK4a along with p14ARF [39]. CDKN2A mutations are less common in acral melanoma, with only 9–18% of acral melanomas showing somatic CDKN2A alterations, and these alterations were predominantly homozygous deletions [40,41]. CDKN2A mutation rates were similarly low in uveal melanoma, with only one missense mutation (V28G within exon 1α) detected in the 80 uveal melanomas included in TCGA uveal melanoma cohort. However, approximately 32% of primary uveal melanomas and 50% of uveal melanoma cell lines show hypermethylation of the CDKN2A promoter, and this promoter methylation predominantly affected p16INK4a expression [27,28,29].
3. Germline CDKN2A Alterations in Familial Melanoma
Loss of function alterations affecting the CDKN2A locus have been identified in the germline of multiple-case melanoma kindreds around the world [42,43]. These CDKN2A germline mutations are associated with a 65-fold increase in the risk of melanoma development [44]. Individuals in families with CDKN2A alterations commonly have higher numbers of atypical melanocytic nevi [45]. The systematic analysis of 80 melanoma-prone families identified 37 distinct mutations affecting the CDKN2A locus. Half of these mutations were in exon 1α (these will not affect the p14ARF coding sequence) and half were in exon 2. Whereas all exon 2 mutations altered the p16INK4a protein, only 14/20 of these exon 2 mutations also impacted the p14ARF protein sequence [45].
As is the case with CDKN2A somatic alterations, exon 1β-specific genetic alterations affecting p14ARF alone are less frequent than mutations targeting the p16INK4a coding regions (Table 2). Germline inactivation of p14ARF is mostly due to whole gene deletions, insertions, or splice mutations [46,47,48]. For instance, a germline deletion of p14ARF exon 1β coding sequence and a germline mutation (Gly16Asp) in exon 1β were identified in families with melanoma-neural system tumor syndrome [47,49]; a frameshift mutation (16 base pair insertion) was detected in a Spanish female who developed multiple primary melanomas at a young age [50]; a splice mutation resulting in haploinsufficiency was associated with melanoma in a single family [51]; and a mutation coding for the missense Arg54His mutation in p14ARF was found in an Italian melanoma family [52]. Large germline CDKN2A deletions encompassing exon 1α, 2, and/or 3 which affect both p14ARF and p16INK4a have also been identified in melanoma-prone kindreds [53,54], although these are uncommon and account for only 2% of germline CDKN2A alterations [55]. A germline splice site mutation removing exon 2 of CDKN2A was identified in a family with melanoma and multiple dysplastic nevi [56].
The penetrance of germline CDKN2A mutations for melanoma also varies according to geographical locations, with penetrance increasing with higher baseline melanoma incidence rates. For instance, lifetime penetrance of CDKN2A mutations was 0.58 in Europe, 0.76 in the United States, and 0.91 in Australia [38]. The median age of melanoma diagnosis was also younger in Australian melanoma-prone families compared to European families [10]. A GenoMEL study that screened for CDKN2A mutations in Australian, European, and North American families reported increased mutation frequency in families with melanoma and pancreatic cancer [10]. A prospective study examining cancer diagnoses in Swedish carriers of the Arg112dup alteration in CDKN2A concluded that mutation carriers had a significantly elevated risk of developing pancreatic, lung, head and neck, and gastro-oesophageal carcinomas [57]. Based on five clinical features (number of family members with melanoma, number of members with multiple primary melanomas, median age at diagnosis, presence of pancreatic cancer, and presence of upper airway cancer), Potjer et al. developed the CDKN2A Mutation (CM) score to predict CDKN2A germline mutation status among melanoma prone families; a CM score > 35 out of 49 possible points was associated with a > 90% probability of a melanoma-prone family sharing a CDKN2A mutation [58]. Importantly, CDKN2A mutation carriers have been reported to be at increased risk of developing other cancers, including breast, lung, and non-melanoma skin cancers [57,59,60,61,62]. These additional cancer risks are not consistently observed, however, and this may indicate that the risk of other cancer types reflects the specific germline CDKN2A mutation. For instance, whereas 25–36% of melanoma-prone families with Arg24Pro, c.34G>T, or Gly101Trp had pancreatic cancer, none of the 30 families with Met53Ile, c.IVS2-104A>G, or c.32_33ins9-32 developed pancreatic cancer [10]. Finally, a CDKN2A germline deletion of the p14ARF-specific exon 1ß was associated with excess risk for melanoma, astrocytoma, neurofibromas, and schwannomas [47].
Modifier genes for melanoma kindreds carrying CDKN2A mutations have also been reported. In an early study on Dutch melanoma families, the melanocortin-1 receptor (MC1R) variant Arg151Cys increased the risk of melanoma in carriers of a p16INK4a-inactivating deletion (known as p16-Leiden). This increased risk of melanoma was not wholly attributed to the fair skin type associated with this MC1R variant [63]. A later study confirmed that four frequent MC1R variants (Val60Leu, Val92Met, Arg151Cys, Arg160Tro) were associated with an increased melanoma risk in CDKN2A mutation carriers [64]. Polymorphisms in genes involved in DNA repair (POLN, PRKDC), immune regulation (IL9), and apoptosis (BCL7A, BCL2L1) have also been associated with increased melanoma risk, and in some instances, these polymorphisms (IL9 and BCL7A) have stronger risks in CDKN2A-positive families [65,66].
4. Functional Consequences of Melanoma-Associated CDKN2A Alterations
4.1. p16INK4a Mutations
Melanoma-associated CDKN2A missense mutations commonly diminish the capacity of p16INK4a to bind and inhibit CDK4/6 [67]. For example, Ranade et al. described germline CDKN2A substitutions that impaired the ability of p16INK4a to inhibit the catalytic activity of cyclin D1/CDK4 and cyclin D1/CDK6 complexes [14]. The Met53Ile and Arg24Pro germline mutants of p16INK4a have diminished capacity to bind CDK4 compared to wild-type p16INK4a [68]. Although, it is worth noting that the CDK4-binding affinity of the Arg24Pro mutation is controversial [69]. The somatic missense p16INK4a mutation (Pro48Leu) decreased the ability of the protein to bind and inhibit CDK6 kinase activity, thus, failing to arrest melanoma cell growth [70]. Melanoma cell lines with a Pro81Leu missense mutation in p16INK4a also showed defective binding ability to CDK4, and these cells had more aggressive cell growth compared to the wild-type cells [71].
There is some evidence that CDKN2A mutations do not only impact the binding affinity of p16INK4a for CDK4 or CDK6. Indeed, several melanoma-associated mutations (e.g., N-terminal 24bp p16INK4a duplication, Arg24Pro, Leu117Pro) retained CDK4 and/or CDK6 binding activity even though they displayed diminished cell cycle inhibitory activity, suggesting that other p16INK4a binding interactions may be important in melanoma susceptibility [69,72]. p16INK4a mutants may also dysregulate the oxidative stress response independent of cell cycle regulation. Elevated levels of intracellular reactive oxygen species and oxidative DNA damage were observed in p16INK4a-deficient melanocytes and cancer cells, likely due to altered p38 MAPK signaling [73]. Various p16INK4a mutants, including the melanoma-associated germline mutations Ala36Pro and Ala57Val, were also associated with impaired oxidative regulatory functions [74], and intracellular oxidative dysregulation in melanocytes can lead to genetic damage that contributes to increased melanoma susceptibility [73].
Inactivation of p16INK4a has been shown to contribute to the failure of senescence and progression from normal melanocytes to malignant melanoma via benign nevi, dysplastic nevi, radial growth phase, and vertical growth phase stages [75,76]. Loss of p16INK4a as a single event is not sufficient to induce melanomagenesis but does predispose one to melanoma development, especially in the presence of other driver mutations. For instance, loss of p16INK4a cooperates with BRAFV600E oncogenic mutation to promote melanoma progression in genetic mouse models [77]. More recently, Zeng et al. reported that bi-allelic deletion of p16INK4a induced hyper-motile and invasive melanocyte behavior via the transcriptional activation of the lineage-restricted transcription factor BRN2 [78].
4.2. p14ARF Mutations
The functional impact of exon 1β variants which alter only p14ARF has been less well studied. Specific germline deletions of exon 1β resulting in loss of p14ARF have been identified in melanoma cell lines and in a family with melanoma-neural system tumor syndrome [47,49]. A 16 base pair insertion (60ins16) caused by a duplication of a CG-rich region within exon 1β was detected in a Spanish female who had multiple primary melanomas. This mutant p14ARF retained the N-terminal nucleolar localization sequence but was restricted to the cytoplasm, failed to stabilize p53, and did not induce cell cycle arrest in p53-expressing melanoma cells [50]. In contrast, another p14ARF variant (Gly16Asp) in exon 1β retained its ability to bind HDM2 and stabilize p53 [49]. A splice site mutation in exon 1β was shown to cause p14ARF haploinsufficiency and was associated with melanoma susceptibility [79]. There have been several studies investigating the role of CDKN2A exon 2 mutations that impact the p14ARF coding sequence. In one study, 3/7 p14ARF mutations, encoded by CDKN2A exon 2 mutations, altered the subcellular distribution of p14ARF and diminished its ability to stabilize p53 [50].
5. Clinical Implications of CDKN2A: Impact on Response and Resistance to Current Treatments in Melanoma
The restoration of p14ARF and/or p16INK4a functions has not yet been possible, and most therapeutic strategies involve modulating downstream cell cycle regulators or pathways to overcome the loss of CDKN2A-encoded functions.
5.1. MDM2 Inhibitors
To overcome p14ARF loss, small molecule inhibitors targeting MDM2 activity or the MDM2–p53 interaction have been developed. These inhibitors have shown promising anti-tumor effects in the preclinical setting. The small molecules nutlins (nutlin-1, nutlin-2, and nutlin-3) sterically disrupt the interaction between MDM2 and p53, resulting in p53 accumulation and activation [80]. Nutlin-3 in particular has been shown to inhibit melanoma growth and induce apoptosis in patient-derived xenograft models [81]. Similarly, the MDM2 inhibitor KRT-232 inhibited tumor growth in xenografts derived from 15 melanoma patients, when used alone or in combination with BRAF and/or MEK inhibitors [82]. Importantly, although MDM2 inhibitors will only benefit melanoma patients with p53 wild-type tumors, melanoma often retains expression of wild-type p53 [81].
5.2. CDK Inhibitors
To circumvent p16INK4a loss, CDK inhibitors have been developed and tested with variable success. The first-generation CDK inhibitor, flavopiridol, has broad range activity against CDK1, CDK2, CDK4, and CDK7 and induced cell cycle arrest in preclinical melanoma models but failed to generate any significant clinical activity in a phase II trial of metastatic melanoma patients (NCT00005971 [83]). Second generation CDK-specific inhibitors such as ribociclib (LEE011) and abemaciclib selectively target CDK4 and CDK6, and these have shown more promising results, especially when combined with MAPK inhibitors. For instance, ribociclib in combination with binimetinib (MEK inhibitor) enhanced tumor regression in NRASQ61K-mutant melanoma xenograft models compared to treatment with either ribociclib or binimetinib alone [84]. Ribociclib also demonstrated synergistic effects in combination with encorafenib (BRAF inhibitor) in BRAFV600E-mutant melanoma models. Likewise, treatment with the selective CDK4/6 inhibitor abemaciclib inhibited tumor growth and delayed tumor recurrences in melanoma xenograft mouse models. Importantly, abemaciclib caused tumor regression in vemurafenib (BRAF inhibitor)-resistant tumors, suggesting that CDK4/6 inhibitors may be a viable therapeutic option for melanoma patients who progressed on BRAF/MEK inhibitors [85]. The combination of ribociclib and MDM2 inhibition also enhanced tumor regression and overcame resistance to CDK4/6 inhibitors in a melanoma xenograft model [86].
Selective CDK4/6 inhibitors have since been evaluated in clinical trials. The combination of CDK4 and MEK inhibitors (ribociclib and binimetinib) was tested in a phase Ib/II trial with advanced NRAS-mutant melanoma, and 60–70% of patients experienced clinical benefit (RECISR CR, PR, and SD) [87,88]. The combination of ribociclib and the BRAF inhibitor encorafenib was also evaluated in 18 patients with advanced BRAF-mutant melanoma, with more than half of patients showing clinical benefit (PR/SD) [89]. A triple combination of ribociclib, binimetinib, and encorafenib was evaluated in a phase Ib/II study of 21 patients with BRAF-mutant melanoma, and although increased toxicity was observed with this combination, clinical response was noted in over half of the patients [90].
Loss of CDKN2A has been shown to predict response to CDK4/6 inhibitors in melanoma, glioblastoma, ovarian, and rhabdoid tumor cells [91,92,93]. The activity of CDK4/6 inhibitors was also restricted to melanoma cells that retained expression of the retinoblastoma protein (the downstream effector of CDK4 and CDK6). The presence of an activating Arg24Cys CDK4 mutation, which abolishes the ability of CDK4 to bind to p16INK4 [94] was also associated with melanoma cell sensitivity to CDK4/6 inhibition [95]. These data suggest that activation of the CDK4/6 pathway via loss of p16INK4a or CDK4 activation and the retention of the retinoblastoma protein are key determinants of sensitivity to CDK4/6 inhibition.
5.3. Epigenetic Modulators
Considering that CDKN2A methylation can lead to p14ARF and p16INK4a loss, epigenetic reactivation of CDKN2A has also been attempted with inhibitors of DNA methyltransferase (DNMT), histone deacetylase (HDAC), histone methyltransferase, and histone acetyltransferase. These inhibitors have been shown to induce p14ARF and p16INK4a expression in cancer cell lines and preclinical models (reviewed in reference [96]). In melanoma, treatment of melanoma cell lines with 5-aza-2-deoxycytidine, a DNMT inhibitor, and vorinostat, an HDAC inhibitor, restored p14ARF and p16INK4a function, and this led to reduced cell proliferation, migration, and invasion [29]. However, given that these epigenetic modulators have promiscuous effects, it is difficult to attribute the consequent melanoma control on modulation of p14ARF and p16INK4a function alone.
5.4. BRAF/MEK Inhibitors
The BRAFV600 inhibitors dabrafenib and vemurafenib, in combination with MEK inhibitors trametinib or selumetinib, have now become standard of care in BRAFV600-mutant melanoma. The combination of dabrafenib and trametinib produced response rates of 64%, and progression-free survival and overall survival rates of 13% and 28% at 5 years, respectively, in patients with advanced BRAFV600-mutant melanoma [97], superior to single-agent treatment. However, despite improved response and progression-free survival rates, melanoma patients treated with these selective kinase inhibitors quickly develop resistance and progress within one year.
Despite the high frequency of CDKN2A alterations in melanomas, the impact of CDKN2A mutations on patient responses to BRAF/MEK inhibitors is not well established. For instance, in melanoma cell studies, the presence of p16INK4a-resistant CDK4 mutations (including the melanoma-associated germline CDK4 Arg24Cys mutation) did not alter cell sensitivity to BRAF inhibitors. Conversely, the overexpression of cyclin D1 was associated with BRAF inhibitor resistance and resistance was enhanced when cyclin D1 overexpression was combined with the CDK4 Arg24Cys mutation [98]. Recurrent CDKN2A loss has been implicated in BRAF inhibitor resistance [99,100], although CDKN2A alterations have been found to be pre-existing in responding patients, and CDKN2A alterations commonly co-occur with other mechanisms of BRAF inhibitor resistance (i.e., PTEN loss, N-RAS mutations) [101]. Finally, although reduced CDKN2A copy number at baseline has been associated with poor BRAF inhibitor responses in melanoma [102], the genetic loss of CDKN2A is also a poor prognostic marker in melanoma [103].
It is important to mention that 15–40% of mucosal and acral melanomas show activating mutations or amplification of the receptor tyrosine kinase KIT, and the kinase inhibitor imatinib has shown efficacy in KIT-mutant melanoma with an overall response rate of 54% [104]. Imatinib is also used commonly in the treatment of BCR–ABL chronic myelogenous leukemia but has not been as successful in BCR–ABL positive acute lymphoblastic leukemia showing deletion in the CDKN2A gene, suggesting that expression of p14ARF and/or p16INK4a may sensitize cancer cells to imatinib treatment [105]. Thus, it is tempting to speculate that CDKN2A inactivation in melanoma may analogously diminish sensitivity to imatinib in melanoma.
5.5. Immune Checkpoint Inhibitors
The immune checkpoint inhibitors targeting the inhibitory receptors cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) and programmed death-1 (PD-1) have significantly improved survival of patients with advanced and high-risk stage III melanoma. The CTLA-4 inhibitor ipilimumab generates a response rate of around 20% in melanoma patients, and a small proportion of patients remain disease-free past 10 years [106]. Response rates are higher with PD-1 inhibitors (up to 45%) and the combination of CTLA-4 and PD-1 inhibitors further enhances the response rate to 60% [107,108,109,110].
Immune checkpoint inhibitors show most activity in immunogenic cancers, and in tumors showing IFNγ transcriptome signatures and evidence of infiltrating T cells [111]. In this context, knockout of the CDKN2A gene in mice resulted in increased inflammatory cytokine expression in the skin following chronic UVB irradiation. Additionally, more myeloid cells were identified in the CDKN2A knockout mice [112]. Interestingly, chromosomal 9p losses encompassing CDKN2A can also affect the JAK2 gene (JAK2 is located on chromosome band 9p24.1 [113]). JAK2 is a critical transcription factor in IFNγ signaling, and the loss of JAK2 is associated with PD-1 inhibitor resistance [114]. Indeed, 75% of melanoma tumors carry concurrent loss of the JAK2 and CDKN2A alleles [115]. Hence, loss of CDKN2A may increase inflammatory responses, which may augment response to immune checkpoint blockade, but also confer susceptibility to immunotherapy resistance through IFNγ suppression. Given the complexity of the immune response and the heterogeneity of immune cell subsets, it is unclear if and how p14ARF and/or p16INK4a regulate melanoma response to immunotherapy.
CDKN2A mutations were not significantly associated with clinical outcomes such as median time to progression, overall survival, and disease control rate in a cohort of 102 cutaneous melanoma patients treated with immune checkpoint inhibitors. However, this study did report a trend towards improved time to progression and disease control rate in patients with CDKN2A mutations [116]. Similarly, melanoma patients with CDKN2A germline mutations also showed improved response to immune checkpoint blockade; approximately 58% of carriers responded to therapy, with 32% showing complete response [117], suggesting that CDKN2A mutation may be associated with better immunotherapy response rates. Although the mechanism for improved immunotherapy responsiveness in CDKN2A mutation carriers remains unclear, melanomas with somatic CDKN2A mutations have an increased mutational burden, and this may result in more neoantigens and stronger immune responses [117].
5.6. Chemotherapy
Chemotherapy is still used as salvage treatment for melanoma patients, especially those with BRAF wild-type disease, and in patients who have failed molecular targeted and/or immunotherapy [118]. Chemotherapy agents such as dacarbazine and temozolomide show low response rates of 12–13% and median overall survival of only 6–8 months [119], and although partial response rates for these agents can reach 15–28%, less than 2% of patients will have durable responses (reviewed in [120]).
Expression of p14ARF has been shown to enhance chemosensitivity. For example, p14ARF accumulation induced potent cell cycle arrest in a p53-dependent manner. On its own, p14ARF did not induce apoptosis, but rather sensitized cells to apoptosis in the presence of camptothecin and adriamycin, inhibitors of topoisomerase I and II, in osteosarcoma, colorectal, melanoma, and fibroblast cell lines [121]. This effect is also observed in osteosarcoma cell lines in response to cisplatin-induced apoptosis, however, effects were independent of p53 [122], suggesting a distinct regulatory mechanism that may be treatment dependent.
Similar to p14ARF, ectopic expression of p16INK4a in glioma cell lines also sensitized cells to the chemotherapy drug vincristine [123]. In melanoma cells, CDKN2A expression was associated with better response to chemotherapy in the form of melphalan or actinomycin-D, and enforced accumulation of p16INK4a induced cell death by augmenting response to these cytotoxic drugs [124].
6. Conclusions
The CDKN2A locus is the most common melanoma-dominant predisposition gene and somatic alterations encompassing this genetic sequence occur early in the development of melanoma. Many CDKN2A genetic and epigenetic changes impact both the p16INK4a and p14ARF protein products encoded by this locus, and although early studies confirmed the major contribution of p16INK4a in CDKN2A-associated melanoma, there is now significant evidence that p14ARF plays an important and additional role in melanomagenesis. CDKN2A loss is associated with histological features predictive of poor prognosis in melanoma and also correlates with diminished patient response to treatment, with loss of CDKN2A associated with poor response to BRAF/MEK inhibitors and chemotherapy but potentially improved responses to immune checkpoint inhibitors. The loss of the CDKN2A sequence also co-operates with the BRAF and NRAS oncogenes to promote melanoma development. Thus, there is renewed interest in restoring the functional loss of p16INK4a and p14ARF in melanoma, and the frequent loss of this locus in melanoma may provide unique therapeutic opportunities, as the downstream targets retinoblastoma protein and p53 are often retained. In recent work, the combination of CDK4 and MDM2 inhibitors demonstrated significant preclinical activity, and this combination was effective in melanoma models with genetic loss of CDKN2A. It remains to be determined whether combination therapies that functionally restore CDKN2A will be effective as salvage therapies. The CDKN2A locus may ultimately help stratify patients for optimal treatment and provide therapeutic options for patients who fail standard of care MAPK inhibitor-based and/or PD1-inhibitor-based therapies.
Author Contributions
All authors analyzed and interpreted the data and contributed to writing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was also supported in part by the National Health and Medical Research Council (APP1093017 and APP1128951). H.R. is supported by National Health and Medical Research Council Research Fellowship (APP1104503).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Karimkhani, C.; Green, A.C.; Nijsten, T.; Weinstock, M.A.; Dellavalle, R.P.; Naghavi, M.; Fitzmaurice, C. The global burden of melanoma: Results from the Global Burden of Disease Study 2015. Br. J. Dermatol. 2017, 177, 134–140. [Google Scholar] [CrossRef]
- Costin, G.E.; Hearing, V.J. Human skin pigmentation: Melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [CrossRef] [PubMed]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tyminska, A. Skin melanocytes: Biology and development. Postepy Dermatol. Alergol. 2013, 30, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Mihm, M.C., Jr. Melanoma. N. Engl. J. Med. 2006, 355, 51–65. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Borresen-Dale, A.L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galuppini, F.; Dal Pozzo, C.A.; Deckert, J.; Loupakis, F.; Fassan, M.; Baffa, R. Tumor mutation burden: From comprehensive mutational screening to the clinic. Cancer Cell Int. 2019, 19, 209. [Google Scholar] [CrossRef]
- Hodis, E.; Watson, I.R.; Kryukov, G.V.; Arold, S.T.; Imielinski, M.; Theurillat, J.P.; Nickerson, E.; Auclair, D.; Li, L.; Place, C.; et al. A landscape of driver mutations in melanoma. Cell 2012, 150, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas, N. Genomic Classification of Cutaneous Melanoma. Cell 2015, 161, 1681–1696. [Google Scholar]
- Hayward, N.K.; Wilmott, J.S.; Waddell, N.; Johansson, P.A.; Field, M.A.; Nones, K.; Patch, A.M.; Kakavand, H.; Alexandrov, L.B.; Burke, H.; et al. Whole-genome landscapes of major melanoma subtypes. Nature 2017, 545, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.M.; Chan, M.; Harland, M.; Hayward, N.K.; Demenais, F.; Bishop, D.T.; Azizi, E.; Bergman, W.; Bianchi-Scarra, G.; Bruno, W.; et al. Features associated with germline CDKN2A mutations: A GenoMEL study of melanoma-prone families from three continents. J. Med. Genet. 2007, 44, 99–106. [Google Scholar] [CrossRef]
- de Snoo, F.A.; Hayward, N.K. Cutaneous melanoma susceptibility and progression genes. Cancer Lett. 2005, 230, 153–186. [Google Scholar] [CrossRef]
- Serrano, M.; Hannon, G.J.; Beach, D. A new regulatory motif in cell-cycle control causing specific inhibition of cyclin D/CDK4. Nature 1993, 366, 704–707. [Google Scholar] [CrossRef]
- Serrano, M.; Lee, H.; Chin, L.; Cordon-Cardo, C.; Beach, D.; DePinho, R.A. Role of the INK4a locus in tumor suppression and cell mortality. Cell 1996, 85, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Ranade, K.; Hussussian, C.J.; Sikorski, R.S.; Varmus, H.E.; Goldstein, A.M.; Tucker, M.A.; Serrano, M.; Hannon, G.J.; Beach, D.; Dracopoli, N.C. Mutations associated with familial melanoma impair p16 INK4 function. Nature Genet. 1995, 10, 114–116. [Google Scholar] [CrossRef] [PubMed]
- Pomerantz, J.; Schreiber-Agus, N.; Liegeois, N.J.; Silverman, A.; Alland, L.; Chin, L.; Potes, J.; Chen, K.; Orlow, I.; Lee, H.W.; et al. The Ink4a tumor suppressor gene product, p19Arf, interacts with MDM2 and neutralizes MDM2’s inhibition of p53. Cell 1998, 92, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xiong, Y.; Yarbrough, W.G. ARF promotes MDM2 degradation and stabilizes p53: ARF-INK4a locus deletion impairs both the Rb and p53 tumor suppression pathways. Cell 1998, 92, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Kamijo, T.; Weber, J.D.; Zambetti, G.; Zindy, F.; Roussel, M.F.; Sherr, C.J. Functional and physical interactions of the ARF tumor suppressor with p53 and Mdm2. Proc. Natl. Acad. Sci. USA 1998, 95, 8292–8297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stott, F.J.; Bates, S.; James, M.C.; McConnell, B.B.; Starborg, M.; Brookes, S.; Palmero, I.; Ryan, K.; Hara, E.; Vousden, K.H. The alternative product from the human CDKN2A locus, p14ARF, participates in a regulatory feedback loop with p53 and MDM2. EMBO J. 1998, 17, 5001–5014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Rizos, H.; Becker, T.M.; Holland, E.A.; Kefford, R.F.; Mann, G.J. Differential expression of p16INK4a and p16beta transcripts in B-lymphoblastoid cells from members of hereditary melanoma families without CDKN2A exon mutations. Oncogene 1997, 15, 515–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fountain, J.W.; Karayiorgou, M.; Ernstoff, M.S.; Kirkwood, J.M.; Vlock, D.R.; Titus-Ernstoff, L.; Bouchard, B.; Vijayasaradhi, S.; Houghton, A.N.; Lahti, J. Homozygous deletions within human chromosome band 9p21 in melanoma. Proc. Natl. Acad. Sci. USA 1992, 89, 10557–10561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedberg, D.E.; Rigas, S.H.; Russak, J.; Gai, W.; Kaplow, M.; Osman, I.; Turner, F.; Randerson-Moor, J.A.; Houghton, A.; Busam, K.; et al. Frequent p16-independent inactivation of p14ARF in human melanoma. J. Natl. Cancer Inst. 2008, 100, 784–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; McArthur, G.A. CDK4 inhibitors an emerging strategy for the treatment of melanoma. Melanoma Manag. 2015, 2, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Leung, E.Y.; Baguley, B.C.; Finlay, G.J.; Askarian-Amiri, M.E. Epigenetic regulation in human melanoma: Past and future. Epigenetics 2015, 10, 103–121. [Google Scholar] [CrossRef] [Green Version]
- Kostaki, M.; Manona, A.D.; Stavraka, I.; Korkolopoulou, P.; Levidou, G.; Trigka, E.A.; Christofidou, E.; Champsas, G.; Stratigos, A.J.; Katsambas, A.J.E.d. High-frequency p16 INK 4A promoter methylation is associated with histone methyltransferase SETDB 1 expression in sporadic cutaneous melanoma. Exp. Dermatol. 2014, 23, 332–338. [Google Scholar] [CrossRef]
- Jonsson, A.; Tuominen, R.; Grafström, E.; Hansson, J.; Egyhazi, S. High frequency of p16INK4A promoter methylation in NRAS-mutated cutaneous melanoma. J. Investig. Dermatol. 2010, 130, 2809–2817. [Google Scholar] [CrossRef] [Green Version]
- Straume, O.; Smeds, J.; Kumar, R.; Hemminki, K.; Akslen, L.A. Significant impact of promoter hypermethylation and the 540 C>T polymorphism of CDKN2A in cutaneous melanoma of the vertical growth phase. Am. J. Pathol. 2002, 161, 229–237. [Google Scholar] [CrossRef]
- Venza, M.; Visalli, M.; Biondo, C.; Lentini, M.; Catalano, T.; Teti, D.; Venza, I. Epigenetic regulation of p14ARF and p16INK4A expression in cutaneous and uveal melanoma. Biochim. Biophys. Acta 2015, 1849, 247–256. [Google Scholar] [CrossRef]
- Pasini, D.; Bracken, A.P.; Helin, K. Polycomb group proteins in cell cycle progression and cancer. Cell Cycle 2004, 3, 396–400. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Feng, Y.; Wang, X. Regulation of Tumor Suppressor Gene CDKN2A and Encoded p16-INK4a Protein by Covalent Modifications. Biochemistry 2018, 83, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Li, S.; Munoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordero, F.J.; Huang, Z.; Grenier, C.; He, X.; Hu, G.; McLendon, R.E.; Murphy, S.K.; Hashizume, R.; Becher, O.J. Histone H3.3K27M Represses p16 to Accelerate Gliomagenesis in a Murine Model of DIPG. Mol. Cancer Res. 2017, 15, 1243–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, I.M.; Halvorsen, O.J.; Collett, K.; Stefansson, I.M.; Straume, O.; Haukaas, S.A.; Salvesen, H.B.; Otte, A.P.; Akslen, L.A. EZH2 expression is associated with high proliferation rate and aggressive tumor subgroups in cutaneous melanoma and cancers of the endometrium, prostate, and breast. J. Clin. Oncol. 2006, 24, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, F.; Niebel, D.; Aymans, P.; Ferring-Schmitt, S.; Dietrich, D.; Landsberg, J. H3K27me3 and EZH2 expression in melanoma: Relevance for melanoma progression and response to immune checkpoint blockade. Clin. Epigenetics 2020, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Gonzalgo, M.L.; Bender, C.M.; You, E.H.; Glendening, J.M.; Flores, J.F.; Walker, G.J.; Hayward, N.K.; Jones, P.A.; Fountain, J.W. Low frequency of p16/CDKN2A methylation in sporadic melanoma: Comparative approaches for methylation analysis of primary tumors. Cancer Res. 1997, 57, 5336–5347. [Google Scholar] [PubMed]
- Welch, J.; Millar, D.; Goldman, A.; Heenan, P.; Stark, M.; Eldon, M.; Clark, S.; Martin, N.G.; Hayward, N.K. Lack of genetic and epigenetic changes in CDKN2A in melanocytic nevi. J. Investig. Dermatol. 2001, 117, 383–384. [Google Scholar] [CrossRef] [Green Version]
- Bishop, D.T.; Demenais, F.; Goldstein, A.M.; Bergman, W.; Bishop, J.N.; Bressac-de Paillerets, B.; Chompret, A.; Ghiorzo, P.; Gruis, N.; Hansson, J.; et al. Geographical variation in the penetrance of CDKN2A mutations for melanoma. J. Natl. Cancer Inst. 2002, 94, 894–903. [Google Scholar] [CrossRef] [Green Version]
- Shain, A.H.; Yeh, I.; Kovalyshyn, I.; Sriharan, A.; Talevich, E.; Gagnon, A.; Dummer, R.; North, J.; Pincus, L.; Ruben, B.; et al. The Genetic Evolution of Melanoma from Precursor Lesions. N. Engl. J. Med. 2015, 373, 1926–1936. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.R.; Choi, Y.D.; Kim, J.M.; Jin, S.; Shin, M.-H.; Shim, H.-J.; Lee, J.-B.; Yun, S.J. Genetic alterations in primary acral melanoma and acral melanocytic nevus in Korea: Common mutated genes show distinct cytomorphological features. J. Investig. Dermatol. 2018, 138, 933–945. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.S.; Hendricks, W.; Kiefer, J.; Schmidt, J.; Sekar, S.; Carpten, J.; Craig, D.W.; Adkins, J.; Cuyugan, L.; Manojlovic, Z.; et al. Integrated genomic analyses reveal frequent TERT aberrations in acral melanoma. Genome Res. 2017, 27, 524–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussussian, C.J.; Struewing, J.P.; Goldstein, A.M.; Higgins, P.A.; Ally, D.S.; Sheahan, M.D.; Clark, W.H., Jr.; Tucker, M.A.; Dracopoli, N.C. Germline p16 mutations in familial melanoma. Nat. Genet. 1994, 8, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kamb, A.; Shattuck-Eidens, D.; Eeles, R.; Liu, Q.; Gruis, N.A.; Ding, W.; Hussey, C.; Tran, T.; Miki, Y.; Weaver-Feldhaus, J. Analysis of the p16 gene (CDKN2) as a candidate for the chromosome 9p melanoma susceptibility locus. Nat. Genet. 1994, 8, 22–26. [Google Scholar] [CrossRef]
- Helgadottir, H.; Hoiom, V.; Tuominen, R.; Nielsen, K.; Jonsson, G.; Olsson, H.; Hansson, J. Germline CDKN2A Mutation Status and Survival in Familial Melanoma Cases. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, J.A.; Wachsmuth, R.C.; Harland, M.; Bataille, V.; Pinney, E.; Mac, K.P.; Baglietto, L.; Cuzick, J.; Bishop, D.T. Genotype/phenotype and penetrance studies in melanoma families with germline CDKN2A mutations. J. Investig. Dermatol. 2000, 114, 28–33. [Google Scholar] [CrossRef]
- Bahuau, M.; Vidaud, D.; Jenkins, R.B.; Bieche, I.; Kimmel, D.W.; Assouline, B.; Smith, J.S.; Alderete, B.; Cayuela, J.M.; Harpey, J.P.; et al. Germ-line deletion involving the INK4 locus in familial proneness to melanoma and nervous system tumors. Cancer Res. 1998, 58, 2298–2303. [Google Scholar]
- Randerson-Moor, J.A.; Harland, M.; Williams, S.; Cuthbert-Heavens, D.; Sheridan, E.; Aveyard, J.; Sibley, K.; Whitaker, L.; Knowles, M.; Newton Bishop, J. A germline deletion of p14ARF but not CDKN2A in a melanoma–neural system tumour syndrome family. Human Mol. Genet. 2001, 10, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoude, L.G.; Wadt, K.A.; Pritchard, A.L.; Hayward, N.K. Genetics of familial melanoma: 20 years after CDKN2A. Pigment. Cell Melanoma Res. 2015, 28, 148–160. [Google Scholar] [CrossRef]
- Laud, K.; Marian, C.; Avril, M.-F.; Barrois, M.; Chompret, A.; Goldstein, A.M.; Tucker, M.A.; Clark, P.A.; Peters, G.; Chaudru, V. Comprehensive analysis of CDKN2A (p16INK4A/p14ARF) and CDKN2B genes in 53 melanoma index cases considered to be at heightened risk of melanoma. J. Med. Genet. 2006, 43, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Rizos, H.; Puig, S.; Badenas, C.; Malvehy, J.; Darmanian, A.P.; Jiménez, L.; Milà, M.; Kefford, R.F. A melanoma-associated germline mutation in exon 1 β inactivates p14ARF. Oncogene 2001, 20, 5543–5547. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, C.; Lee Wu, C.; Evans, G.; Howell, A.; Elles, R.G.; Jordan, R.; Sloan, P.; Read, A.P.; Thakker, N. Germline mutation of ARF in a melanoma kindred. Hum. Mol. Genet. 2002, 11, 1273–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binni, F.; Antigoni, I.; De Simone, P.; Majore, S.; Silipo, V.; Crisi, A.; Amantea, A.; Pacchiarini, D.; Castori, M.; De Bernardo, C. Novel and recurrent p14ARF mutations in Italian familial melanoma. Clin. Genet. 2010, 77, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Mistry, S.H.; Taylor, C.; Randerson-Moor, J.A.; Harland, M.; Turner, F.; Barrett, J.H.; Whitaker, L.; Jenkins, R.B.; Knowles, M.A.; Bishop, J.A.; et al. Prevalence of 9p21 deletions in UK melanoma families. Genes Chromosomes Cancer 2005, 44, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Knappskog, S.; Geisler, J.; Arnesen, T.; Lillehaug, J.R.; Lonning, P.E. A novel type of deletion in the CDKN2A gene identified in a melanoma-prone family. Genes Chromosomes Cancer 2006, 45, 1155–1163. [Google Scholar] [CrossRef]
- Lesueur, F.; de Lichy, M.; Barrois, M.; Durand, G.; Bombled, J.; Avril, M.F.; Chompret, A.; Boitier, F.; Lenoir, G.M.; Bressac-de Paillerets, B.; et al. The contribution of large genomic deletions at the CDKN2A locus to the burden of familial melanoma. Br. J. Cancer 2008, 99, 364–370. [Google Scholar] [CrossRef] [Green Version]
- Petronzelli, F.; Sollima, D.; Coppola, G.; Martini-Neri, M.E.; Neri, G.; Genuardi, M. CDKN2A germline splicing mutation affecting both P16INK4 and P14ARF RNA processing in a melanoma/neurofibroma kindred. Genes Chromosome Cancer 2001, 31, 398–401. [Google Scholar] [CrossRef]
- Helgadottir, H.; Hoiom, V.; Jonsson, G.; Tuominen, R.; Ingvar, C.; Borg, A.; Olsson, H.; Hansson, J. High risk of tobacco-related cancers in CDKN2A mutation-positive melanoma families. J. Med. Genet. 2014, 51, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Potjer, T.P.; Helgadottir, H.; Leenheer, M.; van der Stoep, N.; Gruis, N.A.; Hoiom, V.; Olsson, H.; van Doorn, R.; Vasen, H.F.A.; van Asperen, C.J.; et al. CM-Score: A validated scoring system to predict CDKN2A germline mutations in melanoma families from Northern Europe. J. Med. Genet. 2018, 55, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Potrony, M.; Puig-Butille, J.A.; Aguilera, P.; Badenas, C.; Carrera, C.; Malvehy, J.; Puig, S. Increased prevalence of lung, breast, and pancreatic cancers in addition to melanoma risk in families bearing the cyclin-dependent kinase inhibitor 2A mutation: Implications for genetic counseling. J. Am. Acad. Dermatol. 2014, 71, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, A.M.; Chan, M.; Harland, M.; Gillanders, E.M.; Hayward, N.K.; Avril, M.F.; Azizi, E.; Bianchi-Scarra, G.; Bishop, D.T.; Bressac-de Paillerets, B.; et al. High-risk melanoma susceptibility genes and pancreatic cancer, neural system tumors, and uveal melanoma across GenoMEL. Cancer Res. 2006, 66, 9818–9828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasen, H.F.; Gruis, N.A.; Frants, R.R.; van Der Velden, P.A.; Hille, E.T.; Bergman, W. Risk of developing pancreatic cancer in families with familial atypical multiple mole melanoma associated with a specific 19 deletion of p16 (p16-Leiden). Int. J. Cancer 2000, 87, 809–811. [Google Scholar] [CrossRef]
- Borg, A.; Sandberg, T.; Nilsson, K.; Johannsson, O.; Klinker, M.; Masback, A.; Westerdahl, J.; Olsson, H.; Ingvar, C. High frequency of multiple melanomas and breast and pancreas carcinomas in CDKN2A mutation-positive melanoma families. J. Natl. Cancer Inst. 2000, 92, 1260–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Velden, P.A.; Sandkuijl, L.A.; Bergman, W.; Pavel, S.; van Mourik, L.; Frants, R.R.; Gruis, N.A. Melanocortin-1 receptor variant R151C modifies melanoma risk in Dutch families with melanoma. Am. J. Hum. Genet. 2001, 69, 774–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demenais, F.; Mohamdi, H.; Chaudru, V.; Goldstein, A.M.; Newton Bishop, J.A.; Bishop, D.T.; Kanetsky, P.A.; Hayward, N.K.; Gillanders, E.; Elder, D.E.; et al. Association of MC1R variants and host phenotypes with melanoma risk in CDKN2A mutation carriers: A GenoMEL study. J. Natl. Cancer Inst. 2010, 102, 1568–1583. [Google Scholar] [CrossRef]
- Yang, X.R.; Pfeiffer, R.M.; Wheeler, W.; Yeager, M.; Chanock, S.; Tucker, M.A.; Goldstein, A.M. Identification of modifier genes for cutaneous malignant melanoma in melanoma-prone families with and without CDKN2A mutations. Int. J. Cancer 2009, 125, 2912–2917. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.S.; Pfeiffer, R.M.; Wheeler, W.; Maeder, D.; Burdette, L.; Yeager, M.; Chanock, S.; Tucker, M.A.; Goldstein, A.M.; Yang, X.R. Genetic variants in DNA repair genes and the risk of cutaneous malignant melanoma in melanoma-prone families with/without CDKN2A mutations. Int. J. Cancer 2012, 130, 2062–2066. [Google Scholar] [CrossRef]
- Byeon, I.J.; Li, J.; Ericson, K.; Selby, T.L.; Tevelev, A.; Kim, H.J.; O’Maille, P.; Tsai, M.D. Tumor suppressor p16INK4A: Determination of solution structure and analyses of its interaction with cyclin-dependent kinase 4. Mol. Cell 1998, 1, 421–431. [Google Scholar] [CrossRef]
- Harland, M.; Meloni, R.; Gruis, N.; Pinney, E.; Brookes, S.; Spurr, N.K.; Frischauf, A.M.; Bataille, V.; Peters, G.; Cuzick, J.; et al. Germline mutations of the CDKN2 gene in UK melanoma families. Hum. Mol. Genet. 1997, 6, 2061–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, T.M.; Rizos, H.; Kefford, R.F.; Mann, G.J. Functional impairment of melanoma-associated p16INK4a mutants in melanoma cells despite retention of cyclin-dependent kinase 4 binding. Clin. Cancer Res. 2001, 7, 3282–3288. [Google Scholar]
- Yarbrough, W.G.; Buckmire, R.A.; Bessho, M.; Liu, E.T. Biologic and biochemical analyses of p16INK4a mutations from primary tumors. J. Natl. Cancer Inst. 1999, 91, 1569–1574. [Google Scholar] [CrossRef] [Green Version]
- Walker, G.J.; Gabrielli, B.G.; Castellano, M.; Hayward, N.K. Functional reassessment of P16 variants using a transfection-based assay. Int. J. Cancer 1999, 82, 305–312. [Google Scholar] [CrossRef]
- Li, C.; Liu, T.; Liu, B.; Hernandez, R.; Facelli, J.C.; Grossman, D. A novel CDKN 2A variant (p16L117P) in a patient with familial and multiple primary melanomas. Pigment. Cell Melanoma Res. 2019, 32, 734–738. [Google Scholar] [PubMed]
- Jenkins, N.C.; Liu, T.; Cassidy, P.; Leachman, S.A.; Boucher, K.M.; Goodson, A.G.; Samadashwily, G.; Grossman, D. The p16(INK4A) tumor suppressor regulates cellular oxidative stress. Oncogene 2011, 30, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, N.C.; Jung, J.; Liu, T.; Wilde, M.; Holmen, S.L.; Grossman, D. Familial melanoma-associated mutations in p16 uncouple its tumor-suppressor functions. J. Investig. Dermatol. 2013, 133, 1043–1051. [Google Scholar] [CrossRef] [Green Version]
- Mooi, W.; Peeper, D. Oncogene-induced cell senescence—halting on the road to cancer. N. Engl. J. Med. 2006, 355, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Haferkamp, S.; Becker, T.M.; Scurr, L.L.; Kefford, R.F.; Rizos, H. p16INK4a-induced senescence is disabled by melanoma-associated mutations. Aging Cell 2008, 7, 733–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damsky, W.; Micevic, G.; Meeth, K.; Muthusamy, V.; Curley, D.P.; Santhanakrishnan, M.; Erdelyi, I.; Platt, J.T.; Huang, L.; Theodosakis, N. mTORC1 activation blocks BrafV600E-induced growth arrest but is insufficient for melanoma formation. Cancer Cell 2015, 27, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Jorapur, A.; Shain, A.H.; Lang, U.E.; Torres, R.; Zhang, Y.; McNeal, A.S.; Botton, T.; Lin, J.; Donne, M.; et al. Bi-allelic Loss of CDKN2A Initiates Melanoma Invasion via BRN2 Activation. Cancer Cell 2018, 34, 56–68.e59. [Google Scholar] [CrossRef]
- Garcia-Casado, Z.; Nagore, E.; Fernandez-Serra, A.; Botella-Estrada, R.; Lopez-Guerrero, J.A. A germline mutation of p14/ARF in a melanoma kindred. Melanoma Res. 2009, 19, 335–337. [Google Scholar] [CrossRef]
- Puszynski, K.; Gandolfi, A.; d’Onofrio, A. The pharmacodynamics of the p53-Mdm2 targeting drug Nutlin: The role of gene-switching noise. PLoS Comput. Biol. 2014, 10, e1003991. [Google Scholar] [CrossRef] [Green Version]
- Vilgelm, A.E.; Pawlikowski, J.S.; Liu, Y.; Hawkins, O.E.; Davis, T.A.; Smith, J.; Weller, K.P.; Horton, L.W.; McClain, C.M.; Ayers, G.D.; et al. Mdm2 and aurora kinase a inhibitors synergize to block melanoma growth by driving apoptosis and immune clearance of tumor cells. Cancer Res. 2015, 75, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Shattuck-Brandt, R.L.; Chen, S.-C.; Murray, E.; Johnson, C.A.; Crandall, H.; O’Neal, J.F.; Al-Rohil, R.N.; Nebhan, C.A.; Bharti, V.; Dahlman, K.B. Metastatic Melanoma Patient-derived Xenografts Respond to MDM2 Inhibition as a Single Agent or in Combination with BRAF/MEK Inhibition. Clin. Cancer Res. 2020, 15, 3803–3818. [Google Scholar] [CrossRef] [Green Version]
- Burdette-Radoux, S.; Tozer, R.G.; Lohmann, R.C.; Quirt, I.; Ernst, D.S.; Walsh, W.; Wainman, N.; Colevas, A.D.; Eisenhauer, E.A. Phase II trial of flavopiridol, a cyclin dependent kinase inhibitor, in untreated metastatic malignant melanoma. Investig. New Drugs 2004, 22, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Loo, A.; Chopra, R.; Caponigro, G.; Huang, A.; Vora, S.; Parasuraman, S.; Howard, S.; Keen, N.; Sellers, W.; et al. Abstract PR02: LEE011: An orally bioavailable, selective small molecule inhibitor of CDK4/6– Reactivating Rb in cancer. Mol. Cancer Ther. 2013, 12, PR02. [Google Scholar]
- Yadav, V.; Burke, T.F.; Huber, L.; Van Horn, R.D.; Zhang, Y.; Buchanan, S.G.; Chan, E.M.; Starling, J.J.; Beckmann, R.P.; Peng, S.B. The CDK4/6 inhibitor LY2835219 overcomes vemurafenib resistance resulting from MAPK reactivation and cyclin D1 upregulation. Mol. Cancer Ther. 2014, 13, 2253–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilgelm, A.E.; Saleh, N.; Shattuck-Brandt, R.; Riemenschneider, K.; Slesur, L.; Chen, S.C.; Johnson, C.A.; Yang, J.; Blevins, A.; Yan, C.; et al. MDM2 antagonists overcome intrinsic resistance to CDK4/6 inhibition by inducing p21. Sci. Transl. Med. 2019, 11, eaav7171. [Google Scholar] [CrossRef]
- Sosman, J.A.; Kittaneh, M.; Lolkema, M.P.J.K.; Postow, M.A.; Schwartz, G.; Franklin, C.; Matano, A.; Bhansali, S.; Parasuraman, S.; Kim, K. A phase 1b/2 study of LEE011 in combination with binimetinib (MEK162) in patients with NRAS-mutant melanoma: Early encouraging clinical activity. J. Clin. Oncol. 2014, 32, 9009. [Google Scholar] [CrossRef]
- Schuler, M.H.; Ascierto, P.A.; De Vos, F.Y.F.L.; Postow, M.A.; Van Herpen, C.M.L.; Carlino, M.S.; Sosman, J.A.; Berking, C.; Long, G.V.; Weise, A.; et al. Phase 1b/2 trial of ribociclib+binimetinib in metastatic NRAS-mutant melanoma: Safety, efficacy, and recommended phase 2 dose (RP2D). J. Clin. Oncol. 2017, 35, 9519. [Google Scholar] [CrossRef]
- Taylor, M.; Sosman, J.; Gonzalez, R.; Carlino, M.S.; Kittaneh, M.; Lolkema, M.P.; Miller, W.; Marino, A.; Zhang, V.; Bhansali, S.G.; et al. Phase Ib/Ii Study of Lee011 (Cdk4/6 Inhibitor) and Lgx818 (Braf Inhibitor) in Braf-Mutant Melanoma. Ann. Oncol. 2014, 25, iv374. [Google Scholar] [CrossRef]
- Ascierto, P.A.; Bechter, O.; Wolter, P.; Lebbe, C.; Elez, E.; Miller, W.H.; Long, G.V.; Omlin, A.G.; Siena, S.; Calvo, E.; et al. A phase Ib/II dose-escalation study evaluating triple combination therapy with a BRAF (encorafenib), MEK (binimetinib), and CDK 4/6 (ribociclib) inhibitor in patients (Pts) with BRAF V600-mutant solid tumors and melanoma. J. Clin. Oncol. 2017, 35, 9518. [Google Scholar] [CrossRef]
- Wiedemeyer, W.R.; Dunn, I.F.; Quayle, S.N.; Zhang, J.; Chheda, M.G.; Dunn, G.P.; Zhuang, L.; Rosenbluh, J.; Chen, S.; Xiao, Y.; et al. Pattern of retinoblastoma pathway inactivation dictates response to CDK4/6 inhibition in GBM. Proc. Natl. Acad. Sci. USA 2010, 107, 11501–11506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsumi, Y.; Iehara, T.; Miyachi, M.; Yagyu, S.; Tsubai-Shimizu, S.; Kikuchi, K.; Tamura, S.; Kuwahara, Y.; Tsuchiya, K.; Kuroda, H.; et al. Sensitivity of malignant rhabdoid tumor cell lines to PD 0332991 is inversely correlated with p16 expression. Biochem. Biophys. Res. Commun. 2011, 413, 62–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konecny, G.E.; Winterhoff, B.; Kolarova, T.; Qi, J.; Manivong, K.; Dering, J.; Yang, G.; Chalukya, M.; Wang, H.J.; Anderson, L.; et al. Expression of p16 and retinoblastoma determines response to CDK4/6 inhibition in ovarian cancer. Clin. Cancer Res. 2011, 17, 1591–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfel, T.; Hauer, M.; Schneider, J.; Serrano, M.; Wolfel, C.; Klehmann-Hieb, E.; De Plaen, E.; Hankeln, T.; Meyer zum Buschenfelde, K.H.; Beach, D. A p16INK4a-insensitive CDK4 mutant targeted by cytolytic T lymphocytes in a human melanoma. Science 1995, 269, 1281–1284. [Google Scholar] [CrossRef] [PubMed]
- Young, R.J.; Waldeck, K.; Martin, C.; Foo, J.H.; Cameron, D.P.; Kirby, L.; Do, H.; Mitchell, C.; Cullinane, C.; Liu, W.; et al. Loss of CDKN2A expression is a frequent event in primary invasive melanoma and correlates with sensitivity to the CDK4/6 inhibitor PD0332991 in melanoma cell lines. Pigment. Cell Melanoma Res. 2014, 27, 590–600. [Google Scholar] [CrossRef]
- Zhao, R.; Choi, B.Y.; Lee, M.H.; Bode, A.M.; Dong, Z. Implications of Genetic and Epigenetic Alterations of CDKN2A (p16(INK4a)) in Cancer. EBioMedicine 2016, 8, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Long, G.V.; Eroglu, Z.; Infante, J.; Patel, S.; Daud, A.; Johnson, D.B.; Gonzalez, R.; Kefford, R.; Hamid, O.; Schuchter, L.; et al. Long-Term Outcomes in Patients With BRAF V600-Mutant Metastatic Melanoma Who Received Dabrafenib Combined With Trametinib. J. Clin. Oncol. 2018, 36, 667–673. [Google Scholar] [CrossRef]
- Smalley, K.S.; Lioni, M.; Dalla Palma, M.; Xiao, M.; Desai, B.; Egyhazi, S.; Hansson, J.; Wu, H.; King, A.J.; Van Belle, P. Increased cyclin D1 expression can mediate BRAF inhibitor resistance in BRAF V600E–mutated melanomas. Mol. Cancer Ther. 2008, 7, 2876–2883. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Hugo, W.; Kong, X.; Hong, A.; Koya, R.C.; Moriceau, G.; Chodon, T.; Guo, R.; Johnson, D.B.; Dahlman, K.B.; et al. Acquired resistance and clonal evolution in melanoma during BRAF inhibitor therapy. Cancer Discov. 2014, 4, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Hugo, W.; Shi, H.; Sun, L.; Piva, M.; Song, C.; Kong, X.; Moriceau, G.; Hong, A.; Dahlman, K.B.; Johnson, D.B.; et al. Non-genomic and Immune Evolution of Melanoma Acquiring MAPKi Resistance. Cell 2015, 162, 1271–1285. [Google Scholar] [CrossRef] [Green Version]
- Moriceau, G.; Hugo, W.; Hong, A.; Shi, H.; Kong, X.; Yu, C.C.; Koya, R.C.; Samatar, A.A.; Khanlou, N.; Braun, J.; et al. Tunable-combinatorial mechanisms of acquired resistance limit the efficacy of BRAF/MEK cotargeting but result in melanoma drug addiction. Cancer Cell 2015, 27, 240–256. [Google Scholar] [CrossRef] [Green Version]
- Nathanson, K.L.; Martin, A.M.; Wubbenhorst, B.; Greshock, J.; Letrero, R.; D’Andrea, K.; O’Day, S.; Infante, J.R.; Falchook, G.S.; Arkenau, H.T.; et al. Tumor genetic analyses of patients with metastatic melanoma treated with the BRAF inhibitor dabrafenib (GSK2118436). Clin. Cancer Res. 2013, 19, 4868–4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conway, C.; Beswick, S.; Elliott, F.; Chang, Y.M.; Randerson-Moor, J.; Harland, M.; Affleck, P.; Marsden, J.; Sanders, D.S.; Boon, A.; et al. Deletion at chromosome arm 9p in relation to BRAF/NRAS mutations and prognostic significance for primary melanoma. Genes Chromosomes Cancer 2010, 49, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Hodi, F.S.; Corless, C.L.; Giobbie-Hurder, A.; Fletcher, J.A.; Zhu, M.; Marino-Enriquez, A.; Friedlander, P.; Gonzalez, R.; Weber, J.S.; Gajewski, T.F.; et al. Imatinib for melanomas harboring mutationally activated or amplified KIT arising on mucosal, acral, and chronically sun-damaged skin. J. Clin. Oncol. 2013, 31, 3182–3190. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.T.; Roussel, M.F.; Sherr, C.J. Arf gene loss enhances oncogenicity and limits imatinib response in mouse models of Bcr-Abl-induced acute lymphoblastic leukemia. Proc. Natl. Acad. Sci. USA 2006, 103, 6688–6693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schadendorf, D.; Hodi, F.S.; Robert, C.; Weber, J.S.; Margolin, K.; Hamid, O.; Patt, D.; Chen, T.T.; Berman, D.M.; Wolchok, J.D. Pooled Analysis of Long-Term Survival Data From Phase II and Phase III Trials of Ipilimumab in Unresectable or Metastatic Melanoma. J. Clin. Oncol. 2015, 33, 1889–1894. [Google Scholar] [CrossRef] [Green Version]
- Robert, C.; Schachter, J.; Long, G.V.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.; Lotem, M.; et al. Pembrolizumab versus Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2015, 372, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Ribas, A.; Schachter, J.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.M.; Lotem, M.; et al. Pembrolizumab versus ipilimumab in advanced melanoma (KEYNOTE-006): Post-hoc 5-year results from an open-label, multicentre, randomised, controlled, phase 3 study. Lancet Oncol. 2019, 20, 1239–1251. [Google Scholar] [CrossRef]
- Hodi, F.S.; Chesney, J.; Pavlick, A.C.; Robert, C.; Grossmann, K.F.; McDermott, D.F.; Linette, G.P.; Meyer, N.; Giguere, J.K.; Agarwala, S.S.; et al. Combined nivolumab and ipilimumab versus ipilimumab alone in patients with advanced melanoma: 2-year overall survival outcomes in a multicentre, randomised, controlled, phase 2 trial. Lancet Oncol. 2016, 17, 1558–1568. [Google Scholar] [CrossRef] [Green Version]
- Hodi, F.S.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Nivolumab plus ipilimumab or nivolumab alone versus ipilimumab alone in advanced melanoma (CheckMate 067): 4-year outcomes of a multicentre, randomised, phase 3 trial. Lancet Oncol. 2018, 19, 1480–1492. [Google Scholar] [CrossRef]
- Wargo, J.A.; Reddy, S.M.; Reuben, A.; Sharma, P. Monitoring immune responses in the tumor microenvironment. Curr. Opin. Immunol. 2016, 41, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Guroji, P.; DeBrot, A.H.; Manapragada, P.P.; Katiyar, S.K.; Elmets, C.A.; Yusuf, N. Loss of INK4a/Arf gene enhances ultraviolet radiation-induced cutaneous tumor development. Exp. Dermatol. 2017, 26, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Monti, S.; Rodig, S.J.; Juszczynski, P.; Currie, T.; O’Donnell, E.; Chapuy, B.; Takeyama, K.; Neuberg, D.; Golub, T.R.; et al. Integrative analysis reveals selective 9p24.1 amplification, increased PD-1 ligand expression, and further induction via JAK2 in nodular sclerosing Hodgkin lymphoma and primary mediastinal large B-cell lymphoma. Blood 2010, 116, 3268–3277. [Google Scholar] [CrossRef] [Green Version]
- Zaretsky, J.M.; Garcia-Diaz, A.; Shin, D.S.; Escuin-Ordinas, H.; Hugo, W.; Hu-Lieskovan, S.; Torrejon, D.Y.; Abril-Rodriguez, G.; Sandoval, S.; Barthly, L.; et al. Mutations Associated with Acquired Resistance to PD-1 Blockade in Melanoma. N. Engl. J. Med. 2016, 375, 819–829. [Google Scholar] [CrossRef]
- Horn, S.; Leonardelli, S.; Sucker, A.; Schadendorf, D.; Griewank, K.G.; Paschen, A. Tumor CDKN2A-Associated JAK2 Loss and Susceptibility to Immunotherapy Resistance. J. Natl. Cancer Inst. 2018, 110, 677–681. [Google Scholar] [CrossRef]
- DeLeon, T.T.; Almquist, D.R.; Kipp, B.R.; Langlais, B.T.; Mangold, A.; Winters, J.L.; Kosiorek, H.E.; Joseph, R.W.; Dronca, R.S.; Block, M.S.; et al. Assessment of clinical outcomes with immune checkpoint inhibitor therapy in melanoma patients with CDKN2A and TP53 pathogenic mutations. PLoS ONE 2020, 15, e0230306. [Google Scholar] [CrossRef] [Green Version]
- Helgadottir, H.; Ghiorzo, P.; van Doorn, R.; Puig, S.; Levin, M.; Kefford, R.; Lauss, M.; Queirolo, P.; Pastorino, L.; Kapiteijn, E.; et al. Efficacy of novel immunotherapy regimens in patients with metastatic melanoma with germline CDKN2A mutations. J. Med. Genet. 2020, 57, 316–321. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.A.; Schuchter, L.M. Chemotherapy for Melanoma. Cancer Treat. Res. 2016, 167, 209–229. [Google Scholar]
- Middleton, M.R.; Grob, J.J.; Aaronson, N.; Fierlbeck, G.; Tilgen, W.; Seiter, S.; Gore, M.; Aamdal, S.; Cebon, J.; Coates, A.; et al. Randomized phase III study of temozolomide versus dacarbazine in the treatment of patients with advanced metastatic malignant melanoma. J. Clin. Oncol. 2000, 18, 158–166. [Google Scholar] [CrossRef]
- Yang, A.S.; Chapman, P.B. The history and future of chemotherapy for melanoma. Hematol. Oncol. Clin. N. Am. 2009, 23, 583–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, S.; Kefford, R.F.; Rizos, H. Enforced expression of p14ARF induces p53-dependent cell cycle arrest but not apoptosis. Cell Cycle 2005, 4, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.W.; Zhu, X.F.; Huang, X.F.; Sheng, P.Y.; He, A.S.; Yang, Z.B.; Deng, R.; Feng, G.K.; Liao, W.M. P14ARF sensitizes human osteosarcoma cells to cisplatin-induced apoptosis in a p53-independent manner. Cancer Biol. Ther. 2007, 6, 1074–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, M.; Voss, D.; Park-Simon, T.W.; Mahlberg, R.; Koster, G. Role of p16 and p14ARF in radio- and chemosensitivity of malignant gliomas. Oncol. Rep. 2006, 16, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, S.J.; Thompson, J.F.; Indsto, J.; Scurr, L.L.; Lett, M.; Gao, B.F.; Dunleavey, R.; Mann, G.J.; Kefford, R.F.; Rizos, H. p16INK4a expression and absence of activated B-RAF are independent predictors of chemosensitivity in melanoma tumors. Neoplasia 2008, 10, 1231–1239. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
CDKN2A methylation (HumanMethylation450 arrays) ß-values (β-values range from 0, unmethylated, to 1, fully methylated). (A) Location of methylation probes and exons across the CDKN2A locus. (B) Methylation status of probes located across the CDKN2A locus. (C) Methylation of the cg12840719 probe according to the melanoma genomic subtypes. Data are derived from The Cancer Genome Atlas (TCGA) Skin Cutaneous Melanoma (SKCM) dataset, and only include 62 melanoma tumors without CDKN2A genomic deletions.
Figure 1.
CDKN2A methylation (HumanMethylation450 arrays) ß-values (β-values range from 0, unmethylated, to 1, fully methylated). (A) Location of methylation probes and exons across the CDKN2A locus. (B) Methylation status of probes located across the CDKN2A locus. (C) Methylation of the cg12840719 probe according to the melanoma genomic subtypes. Data are derived from The Cancer Genome Atlas (TCGA) Skin Cutaneous Melanoma (SKCM) dataset, and only include 62 melanoma tumors without CDKN2A genomic deletions.
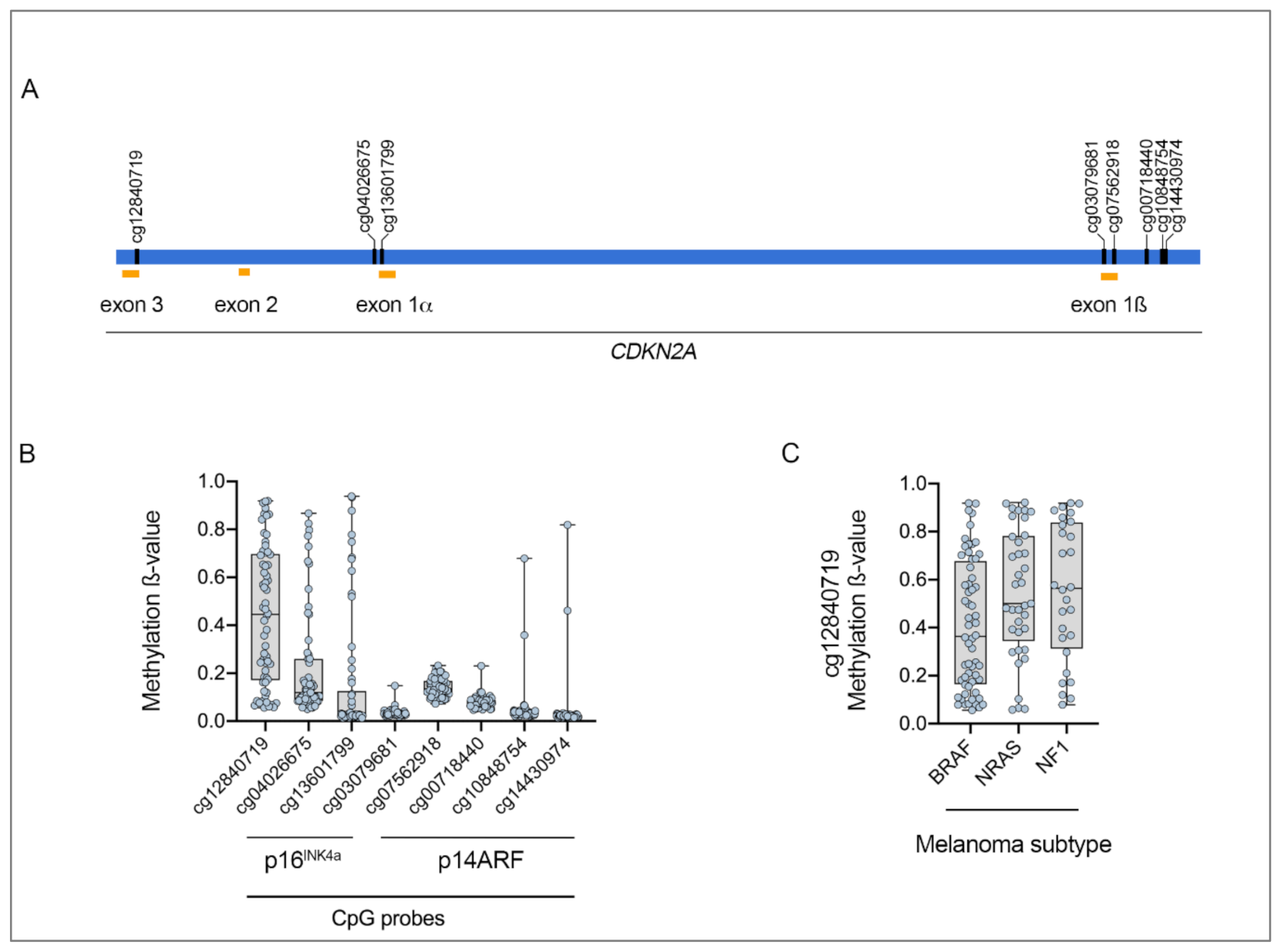
Figure 2.
Kaplan–Meier overall survival curve of patients from TCGA skin cutaneous melanoma cohort with altered vs. unaltered (wild-type) CDKN2A. Median overall survival for patients with CDKN2A alterations is 112 months, compared to a median overall survival of 79 months for melanoma patients with unaltered CDKN2A (p = 0.57, logrank test).
Figure 2.
Kaplan–Meier overall survival curve of patients from TCGA skin cutaneous melanoma cohort with altered vs. unaltered (wild-type) CDKN2A. Median overall survival for patients with CDKN2A alterations is 112 months, compared to a median overall survival of 79 months for melanoma patients with unaltered CDKN2A (p = 0.57, logrank test).

{kind=link}
{kind=link}
Table 1.
Frequency and type of CDKN2A alterations in the four cutaneous melanoma genomic subtypes, designated BRAF, RAS, NF1, and triple wild-type.
Table 1.
Frequency and type of CDKN2A alterations in the four cutaneous melanoma genomic subtypes, designated BRAF, RAS, NF1, and triple wild-type.
| Molecular Subtype | BRAF (n = 188) | NRAS (n = 114) | NF1 (n = 66) | Triple Wild-Type (n = 45) |
|---|---|---|---|---|
| Mutations in p16INK4a only | 7.4% (14/188) | 6.1% (7/114) | 9.1% (6/66) | 0% (0/45) |
| Mutations Affecting p16INK4a and p14ARF | 6.9% (13/188) | 7.9% (9/114) | 12.1% (8/66) | 4.4% (2/45) |
| CDKN2A Deletions | 34.6% (65/188) | 30.7% (35/114) | 28.8% (19/66) | 17.8% (8/45) |
| CDKN2A Hyper-Methylation | 67.0% (126/188) | 87.7% (100/114) | 77.3% (51/66) | 66.7% (30/45) |
Table 2.
Frequency and type of germline CDKN2A alterations identified in 676 individuals with various cancers, including melanoma and pancreatic cancers.
Table 2.
Frequency and type of germline CDKN2A alterations identified in 676 individuals with various cancers, including melanoma and pancreatic cancers.
| Germline Mutations | CDKN2 ANM_000077.4 (p16INK4a) | CDKN2A NM_058195.3 (p14ARF) |
|---|---|---|
| Total Number of Coding Variants | 435 | 179 |
| Number of Coding Variants | ||
| Exon 1a | 140/435 (32%) | 0/477 (0%) |
| Exon 1b | 0/435 (0% | 2/477 (0%) |
| Exon 2 | 295/435 (68%) | 177/477 (37%) |
| Exon 3 | 0/435 (0%) | 0/477 (0%) |
Data derived from LOVD3, Leiden Open Variant Database.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ming, Z.; Lim, S.Y.; Rizos, H. Genetic Alterations in the INK4a/ARF Locus: Effects on Melanoma Development and Progression. Biomolecules 2020, 10, 1447. https://doi.org/10.3390/biom10101447
AMA Style
Ming Z, Lim SY, Rizos H. Genetic Alterations in the INK4a/ARF Locus: Effects on Melanoma Development and Progression. Biomolecules. 2020; 10(10):1447. https://doi.org/10.3390/biom10101447
Chicago/Turabian StyleMing, Zizhen, Su Yin Lim, and Helen Rizos. 2020. "Genetic Alterations in the INK4a/ARF Locus: Effects on Melanoma Development and Progression" Biomolecules 10, no. 10: 1447. https://doi.org/10.3390/biom10101447
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.