Cytotoxicity and Pro-Apoptotic, Antioxidant and Anti-Inflammatory Activities of Geopropolis Produced by the Stingless Bee Melipona fasciculata Smith

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Geopropolis Samples
2.2. Extraction of Samples
2.3. Determination of Antioxidant Activity
2.3.1. DPPH• Radical Scavenging Activity
2.3.2. Ferric Reducing Antioxidant Power Assay (FRAP)
2.3.3. ABTS•+ Assay
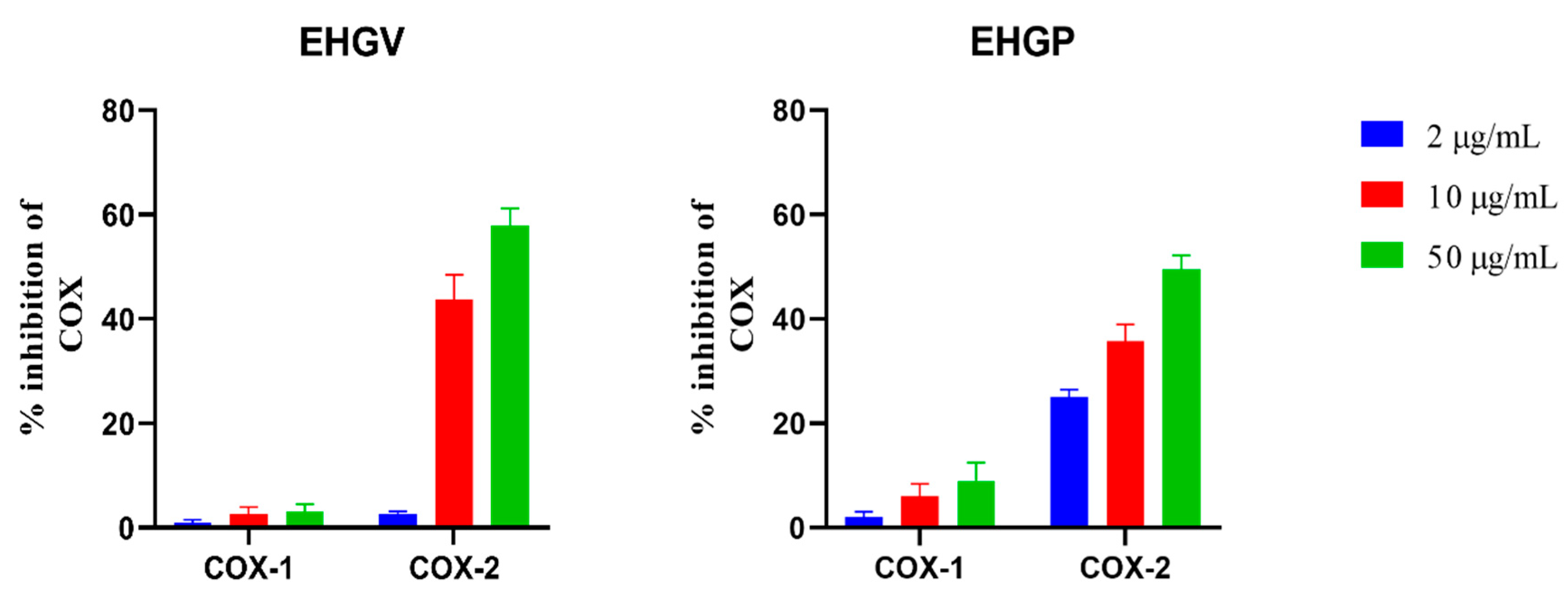
2.4. In Vitro COX Inhibition
2.5. Cell Culture and Morphological Analysis
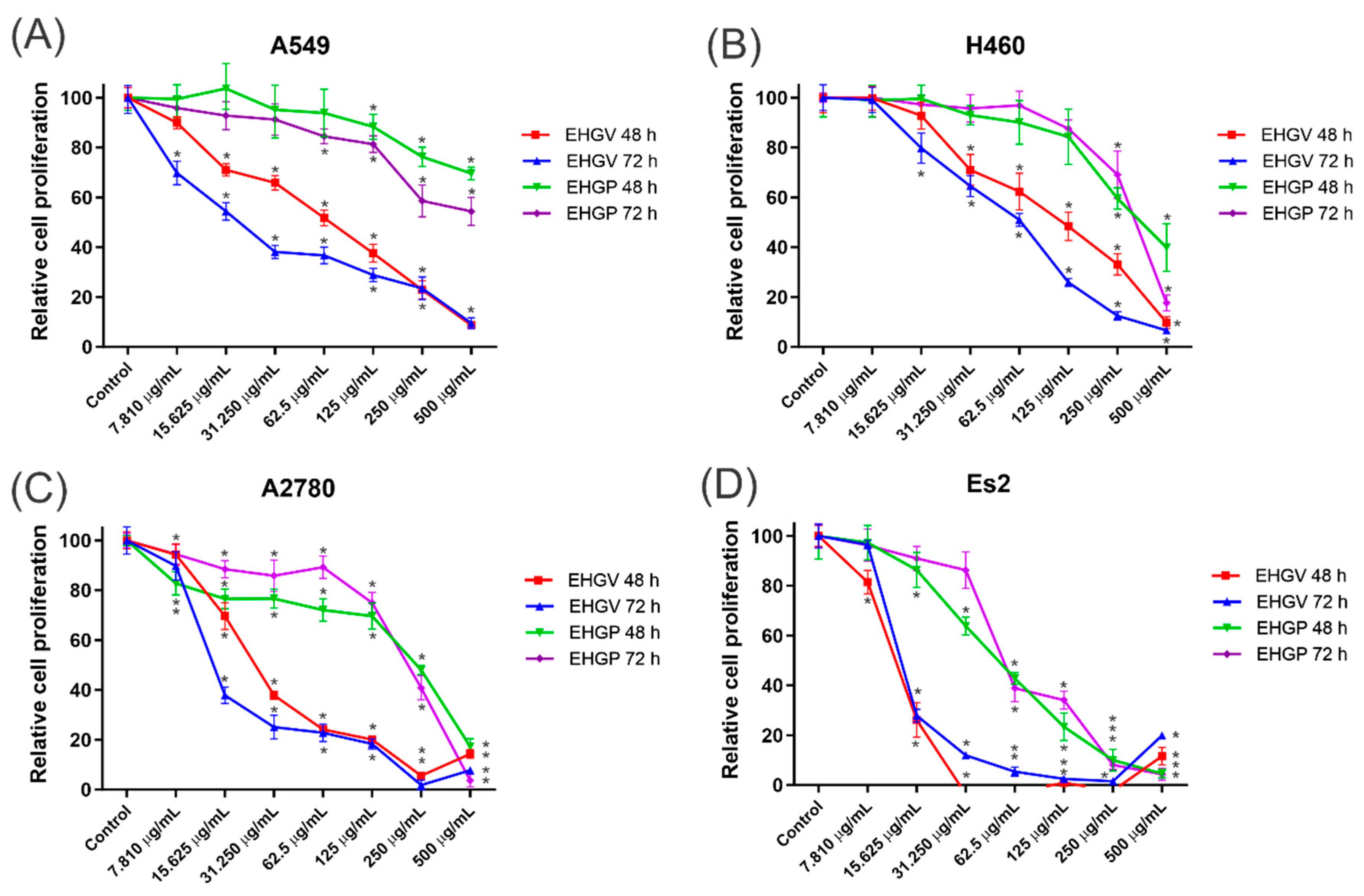
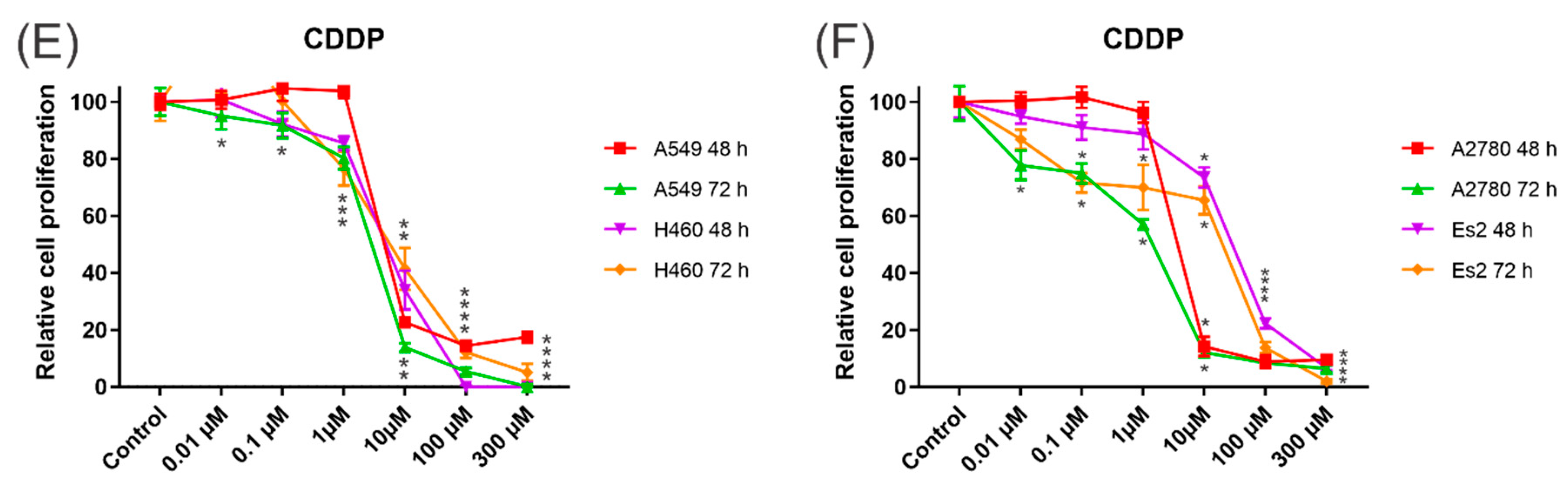
2.6. Cytotoxicity Activity
2.7. Cell Cycle and DNA Content Analysis by Flow Cytometry
2.8. Western Blotting
2.9. LC–ESI/IT–MS/MS Analysis
2.10. In Silico Assay
2.10.1. Predictive Models and Theoretical Calculations
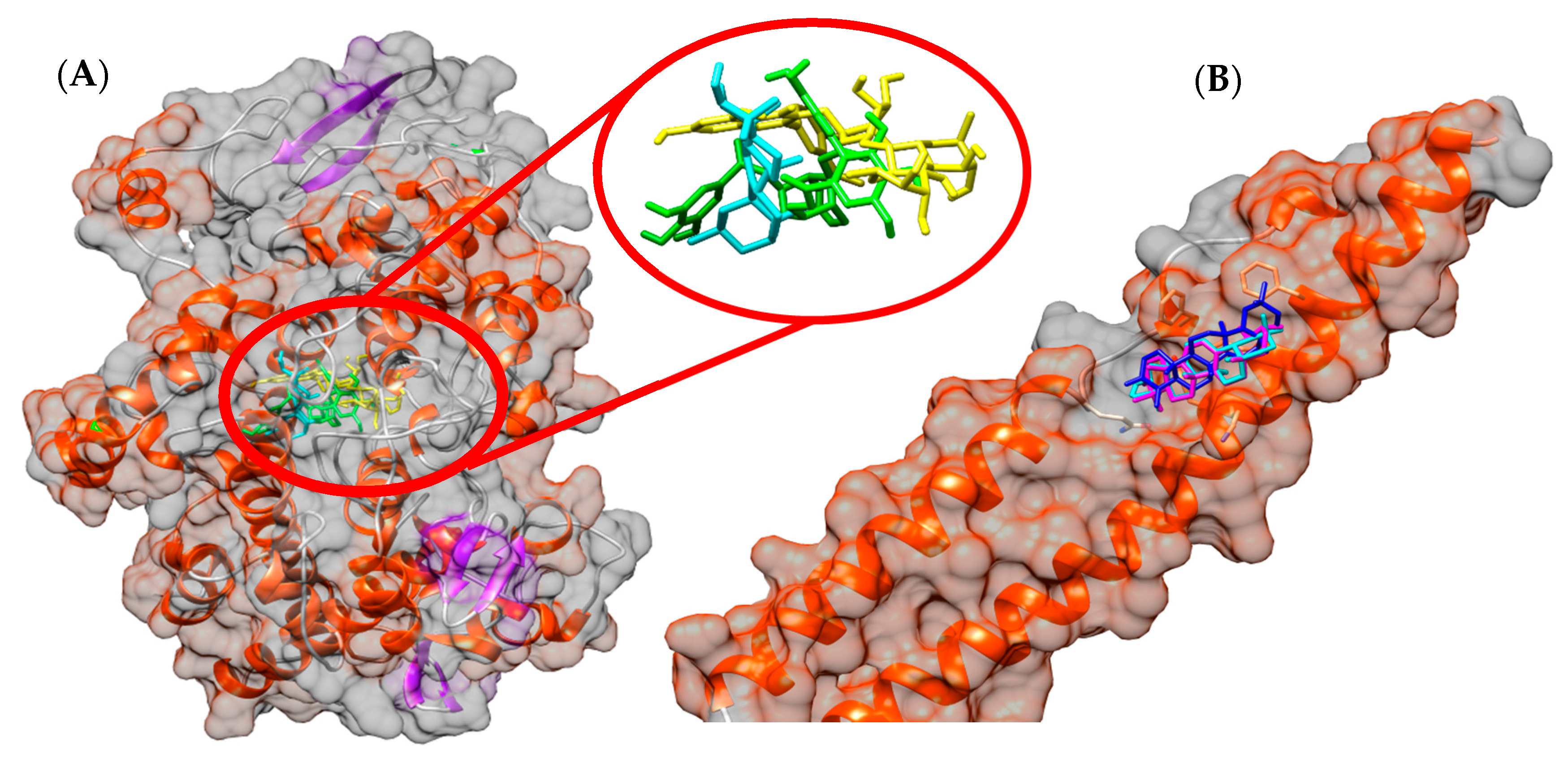
2.10.2. Molecular Docking (MD)
2.11. Statistical Analysis
3. Results
3.1. Antioxidant Activity
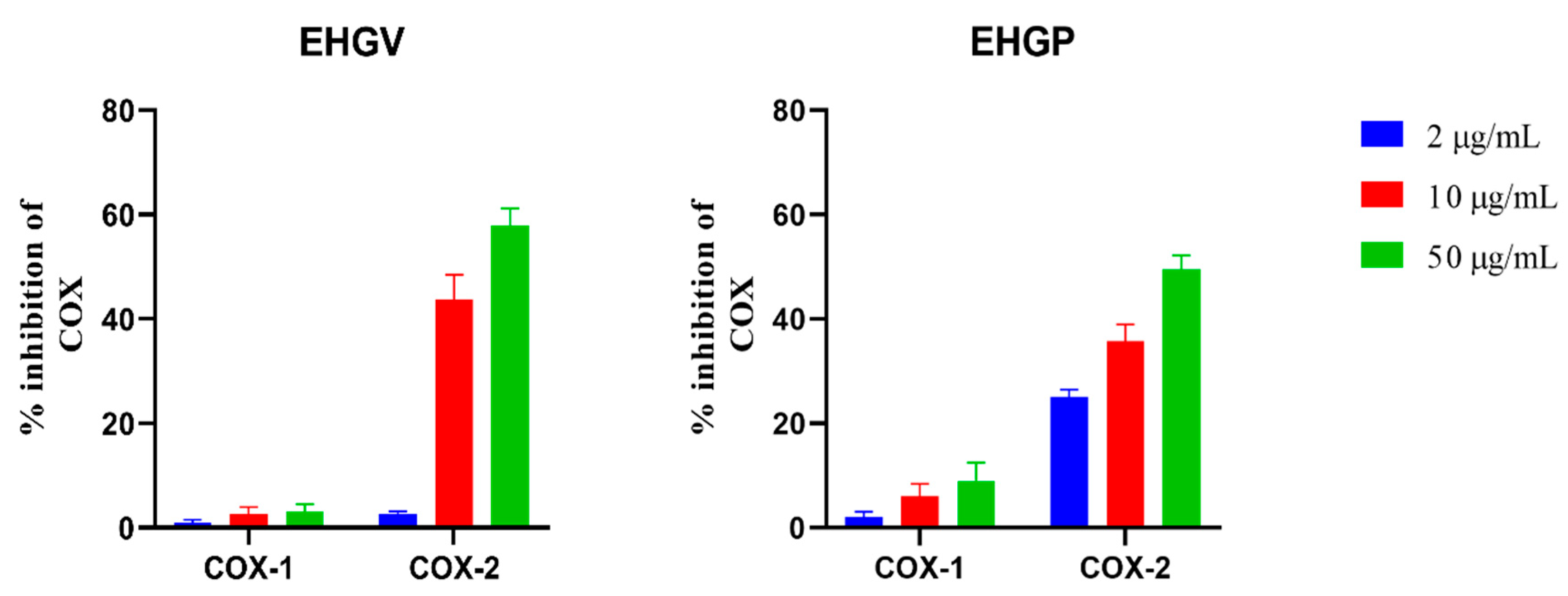
3.2. COX Inhibition Assay
3.3. Cell Culture and Morphological Analysis
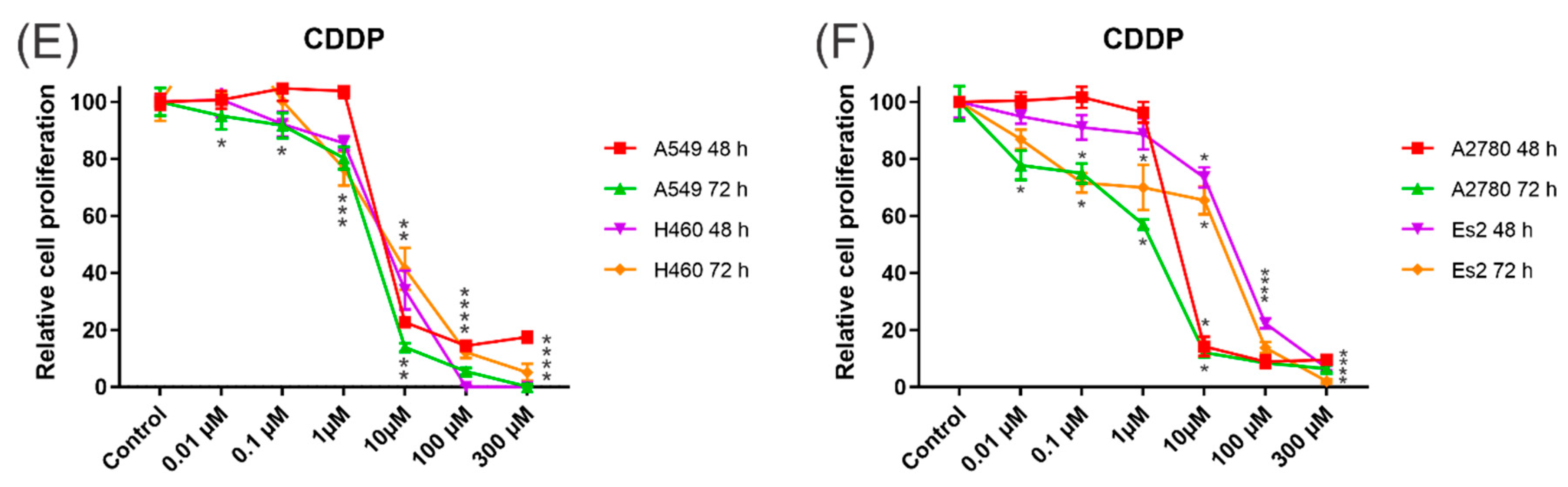
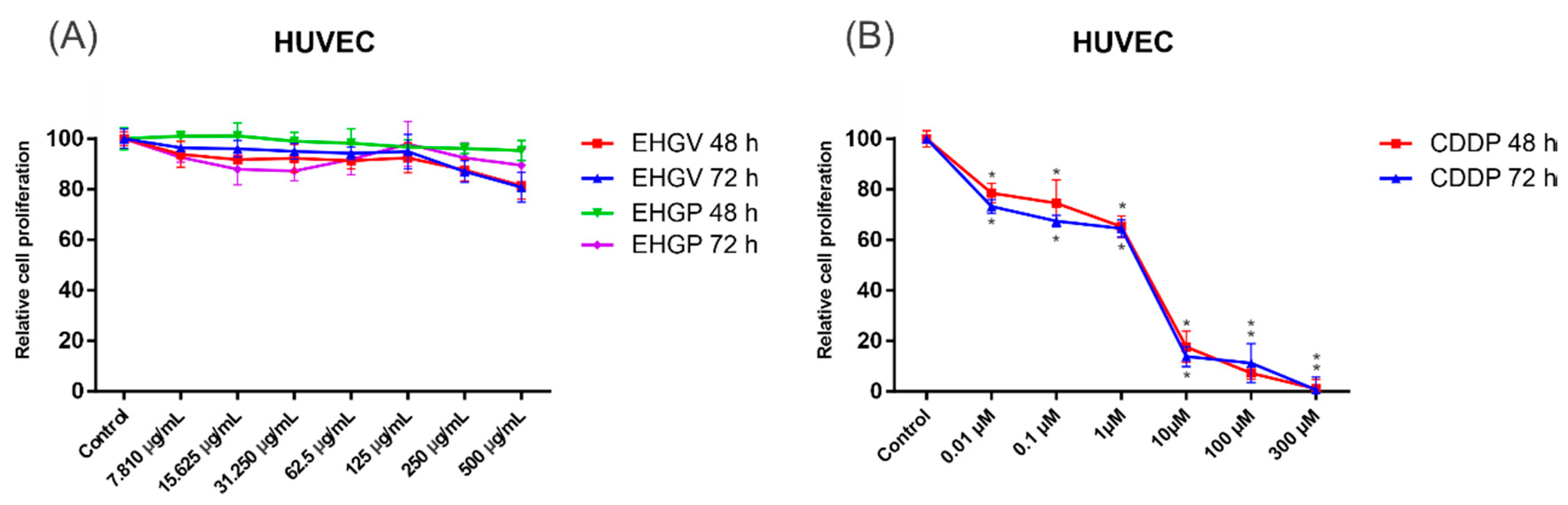
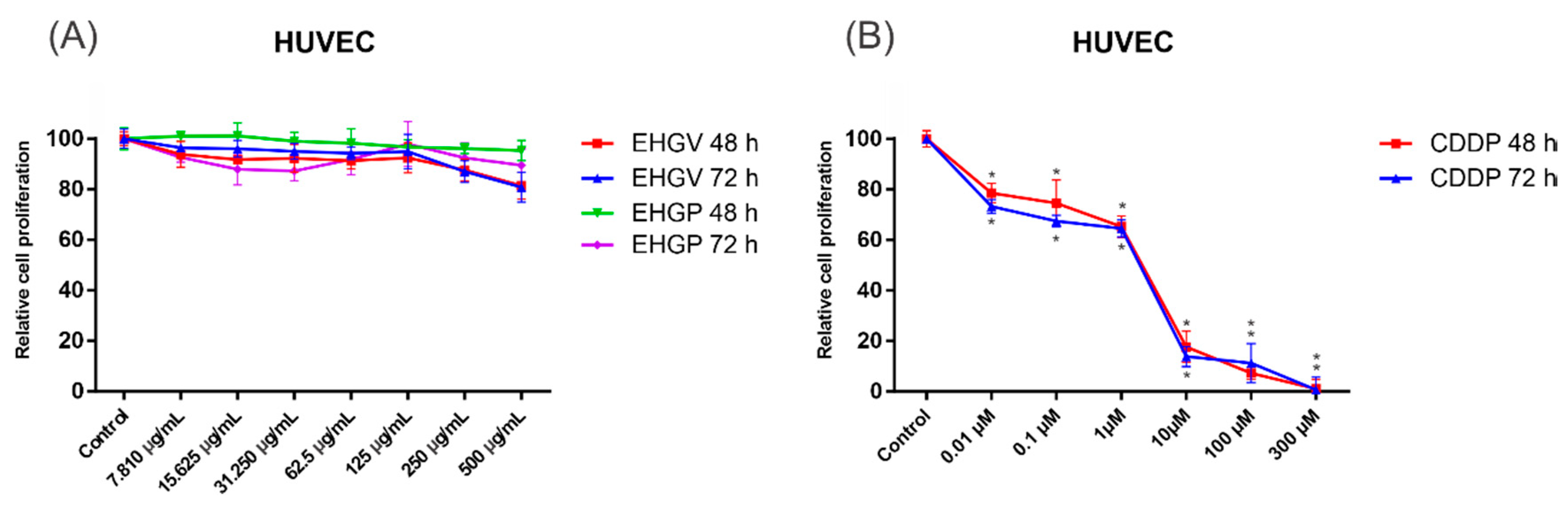
3.4. Cytotoxic Activity
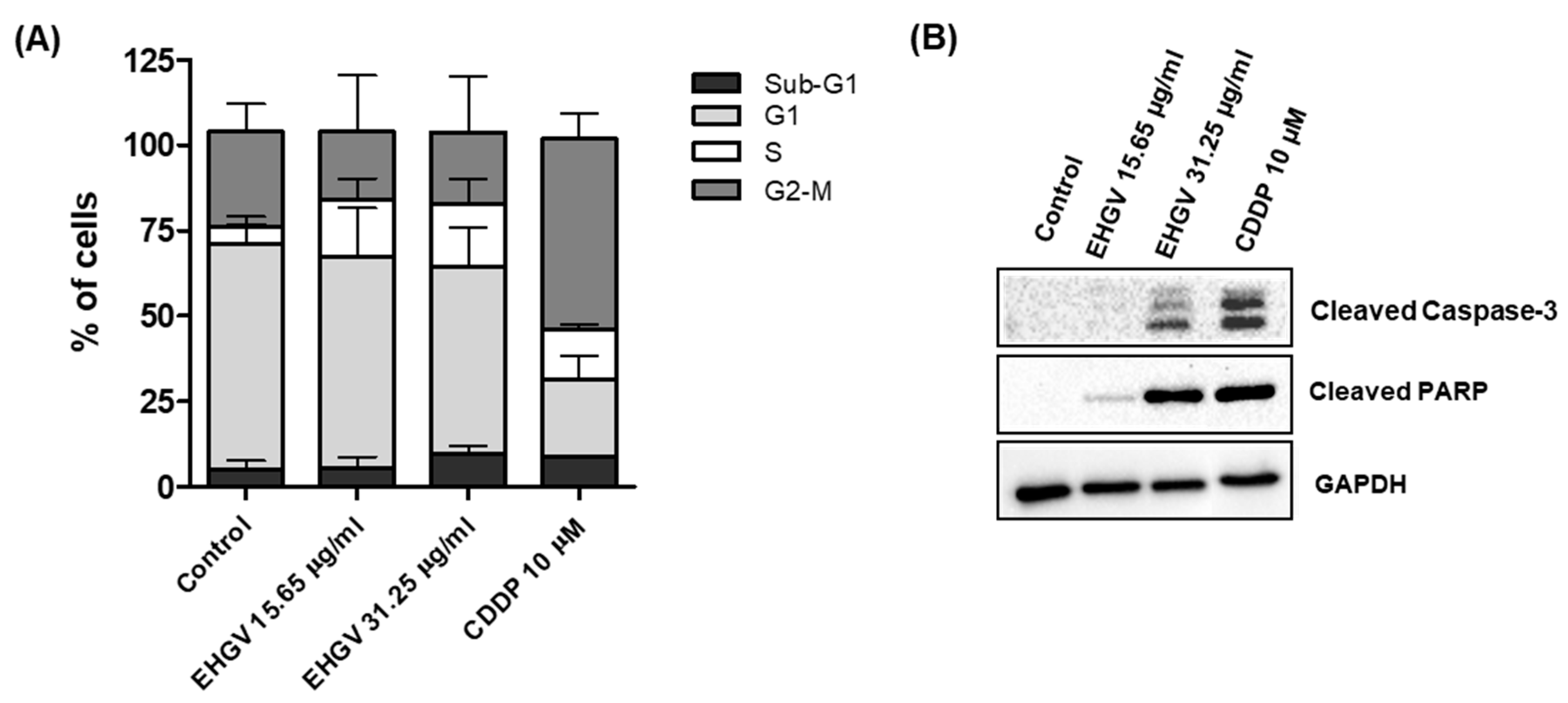
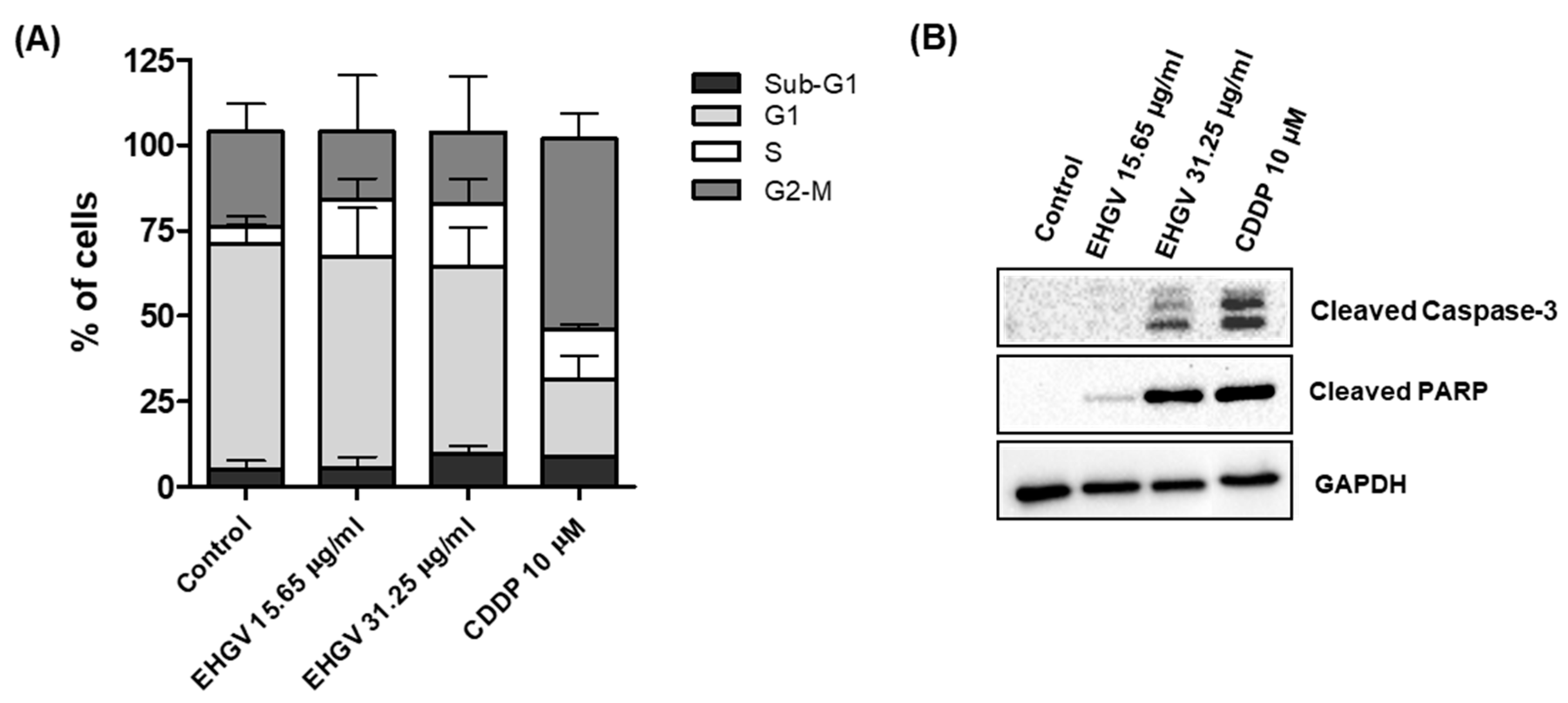
3.5. Cell Cycle Analysis by Flow Cytometry and Analysis of Apoptosis by Western Blot
3.6. LC–ESI/IT–MS/MS Analysis
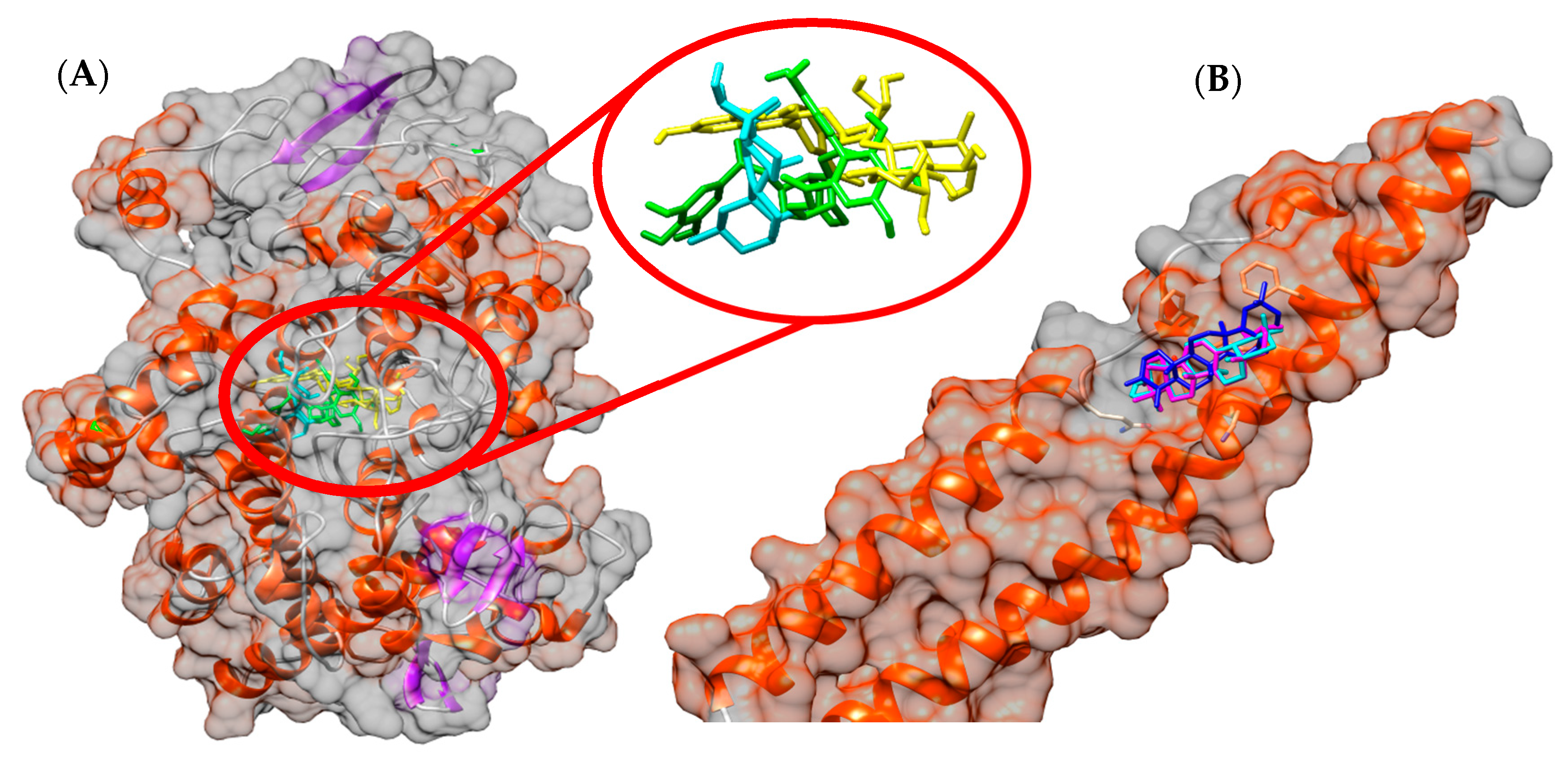
3.7. In Silico Study
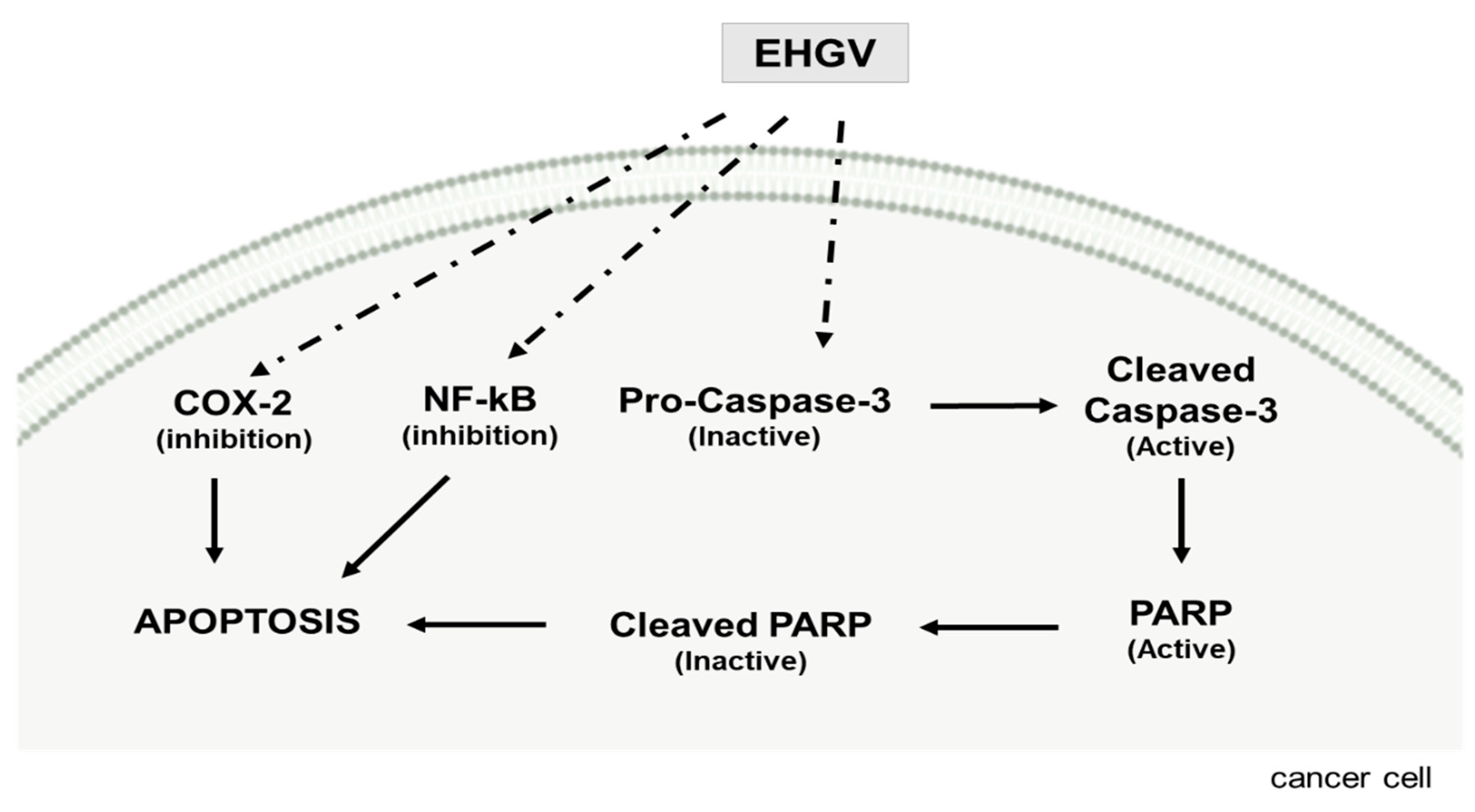
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sadeghi, S.; Davoodvandi, A.; Pourhanifeh, M.H.; Sharifi, N.; Nezhad, R.A.; Sahebnasagh, R.; Moghadam, S.A.; Sahebkar, A.; Mirzaei, H. Anti-cancer effects of cinnamon: Insights into its apoptosis effects. Eur. J. Med. Chem. 2019, 178, 131–140. [Google Scholar] [CrossRef]
- Bray, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Zanetti, R.; Ferlay, J. Cancer Incidence in Five Continents; International Agency for Research on Cancer: Lyon, France, 2019; Volume XI. [Google Scholar]
- Pokhriyal, R.; Hariprasad, R.; Kumar, L.; Hariprasad, G. Chemotherapy Resistance in Advanced Ovarian Cancer Patients. Biomark. Cancer 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Abbaszadeh, H.; Keikhaei, B.; Mottaghi, S. A review of molecular mechanisms involved in anticancer and antiangiogenic effects of natural polyphenolic compounds. Phytother. Res. 2019, 33, 2002–2014. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. Anticancer properties and mechanism of action of the quassinoid ailanthone. Phytother. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Goradel, N.H.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase-2 in cancer: A review. J. Cell. Physiol. 2018, 234, 5683–5699. [Google Scholar] [CrossRef]
- Man, S.; Wei, C.; Liu, C.; Gao, W. Anticancer Drugs from Traditional Toxic Chinese Medicines. Phytother. Res. 2012, 26. [Google Scholar] [CrossRef]
- Ghante, M.H.; Jamkhande, P.G. Role of Pentacyclic Triterpenoids in Chemoprevention and Anticancer Treatment: An Overview on Targets and Underling Mechanisms. J. Pharmacopunct. 2019, 22, 55–67. [Google Scholar]
- Nogueira, P.N. Vida e Criação de Abelhas Indígenas sem Ferrão; Editora Parma: São Paulo, Brazil, 1997. [Google Scholar]
- Kerr, W.E. Abelhas indígenas brasileiras (meliponíneos) na poplinizaçao e na produçao de mel, pólen, geoprópolis e cera. Inf. Agropecuário 1987, 13, 15–22. [Google Scholar]
- Lopes, A.J.O.; Vasconcelos, C.C.; Pereira, F.A.N.; Silva, R.H.M.; Queiroz, P.F.D.S.; Fernandes, C.V.; Garcia, J.B.S.; Ramos, R.M.; Da Rocha, C.Q.; Lima, S.T.; et al. Anti-Inflammatory and Antinociceptive Activity of Pollen Extract Collected by Stingless Bee Melipona fasciculata. Int. J. Mol. Sci. 2019, 20, 4512. [Google Scholar] [CrossRef] [Green Version]
- Franchin, M.; Da Cunha, M.G.; Denny, C.; Napimoga, M.H.; Cunha, F.Q.; Koo, H.; De Alencar, S.M.; Ikegaki, M.; Rosalen, P.L. Geopropolis from Melipona scutellaris decreases the mechanical inflammatory hypernociception by inhibiting the production of IL-1β and TNF-α. J. Ethnopharmacol. 2012, 143, 709–715. [Google Scholar] [CrossRef]
- Franchin, M.; Da Cunha, M.G.; Denny, C.; Napimoga, M.H.; Cunha, F.Q.; Bueno-Silva, B.; De Alencar, S.M.; Ikegaki, M.; Rosalen, P.L. Bioactive Fraction of Geopropolis from Melipona scutellaris Decreases Neutrophils Migration in the Inflammatory Process: Involvement of Nitric Oxide Pathway. Evid. Based Complement. Altern. Med. 2013, 2013, 907041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, C.M.; Campos, J.F.; Dos Santos, H.F.; Balestieri, J.B.P.; Silva, D.B.; Souza, K.D.P.; Carollo, C.A.; Estevinho, L.M.; Dos Santos, E.L. Chemical Composition and Pharmacological Effects of Geopropolis Produced byMelipona quadrifasciata anthidioides. Oxidative Med. Cell. Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liberio, S.A.; Pereira, A.L.A.; Dutra, R.P.; Reis, A.S.; Araújo, M.J.A.; Mattar, N.S.; Silva, L.A.; Ribeiro, M.N.S.; Nascimento, F.F.; Guerra, R.N.M.; et al. Antimicrobial activity against oral pathogens and immunomodulatory effects and toxicity of geopropolis produced by the stingless bee Melipona fasciculata Smith. BMC Complement. Altern. Med. 2011, 11, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.P.G.; Conte, F.L.; Cardoso, E.D.O.; Conti, B.J.; Santiago, K.B.; Golim, M.D.A.; Feltran, G.; Zambuzzi, W.F.; Sforcin, J.M. A new chemotherapeutic approach using doxorubicin simultaneously with geopropolis favoring monocyte functions. Life Sci. 2019, 217, 81–90. [Google Scholar] [CrossRef]
- De Sousa, D.M.N.; Olinda, R.G.; Martins, C.G.; Abrantes, M.R.; Coelho, W.A.C.; da Silva, J.B.A.; de Morais, S.M.; Batista, J.S. Phytochemical screening, in vitro toxicity and evaluation of antioxidant and antibacterial activities of jandaíra bee’s geopropolis. Acta Veter. Bras. 2015, 9, 134–140. [Google Scholar] [CrossRef]
- Sousa, J.P.L.D.M.; Pires, L.D.O.; Dos Santos, R.F.; Prudêncio, E.R.; Sant’Ana, L.D.; Ferreira, D.A.D.S.; Castro, R.N. Chemical and Antimicrobial Potential Study of Brazilian Propolis Produced by Different Species of Bees. Rev. Virtual Química 2019, 11, 1480–1497. [Google Scholar] [CrossRef]
- Dutra, R.P.; Bezerra, J.L.; Da Silva, M.C.P.; Batista, M.C.A.; Patrício, F.J.B.; Nascimento, F.F.; Ribeiro, M.N.S.; Guerra, R.N.M. Antileishmanial activity and chemical composition from Brazilian geopropolis produced by stingless bee Melipona fasciculata. Rev. Bras. Farm. 2019, 29, 287–293. [Google Scholar] [CrossRef]
- Da Silva, E.C.C.; Muniz, M.P.; Nunomura, R.D.C.S.; Nunomura, S.M.; Zilse, G.A.C. Constituintes fenólicos e atividade antioxidante da geoprópolis de duas espécies de abelhas sem ferrão amazônicas. Química Nova 2013, 36, 628–633. [Google Scholar] [CrossRef] [Green Version]
- Dutra, R.P.; Abreu, B.V.D.B.; Cunha, M.S.; Batista, M.C.A.; Torres, L.M.B.; Nascimento, F.F.; Ribeiro, M.N.S.; Guerra, R.N.M. Phenolic Acids, Hydrolyzable Tannins, and Antioxidant Activity of Geopropolis from the Stingless BeeMelipona fasciculataSmith. J. Agric. Food Chem. 2014, 62, 2549–2557. [Google Scholar] [CrossRef]
- Ferreira, J.M.; Fernandes-Silva, C.C.; Salatino, A.; Message, D.; Negri, G. Antioxidant Activity of a Geopropolis from Northeast Brazil: Chemical Characterization and Likely Botanical Origin. Evid. Based Complement. Altern. Med. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Torres, A.; Sandjo, L.; Friedemann, M.; Tomazzoli, M.; Maraschin, M.; Mello, C.F.; Santos, A.R. Chemical characterization, antioxidant and antimicrobial activity of propolis obtained from Melipona quadrifasciata quadrifasciata and Tetragonisca angustula stingless bees. Braz. J. Med. Boil. Res. 2018, 51. [Google Scholar] [CrossRef] [PubMed]
- Cinegaglia, N.C.; Bersano, P.R.O.; Araújo, M.J.A.M.; Búfalo, M.C.; Sforcin, J.M. Anticancer Effects of Geopropolis Produced by Stingless Bees on Canine Osteosarcoma Cells In Vitro. Evid. Based Complement. Altern. Med. 2013, 2013, 737386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.P.G.; Conte, F.L.; Cardoso, E.D.O.; Conti, B.J.; Santiago, K.B.; Golim, M.D.A.; Cruz, M.T.; Sforcin, J.M. Immunomodulatory/inflammatory effects of geopropolis produced byMelipona fasciculataSmith in combination with doxorubicin on THP-1 cells. J. Pharm. Pharmacol. 2016, 68, 1551–1558. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Boil. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Nicoletti, I.; Migliorati, G.; Pagliacci, M.; Grignani, F.; Riccardi, C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J. Immunol. Methods 1991, 139, 271–279. [Google Scholar] [CrossRef]
- De Moraes, G.N.; Carvalho, E.; Maia, R.C.; Sternberg, C. Immunodetection of caspase-3 by Western blot using glutaraldehyde. Anal. Biochem. 2011, 415, 203–205. [Google Scholar] [CrossRef]
- Dennington, R.; Keith, T.A.; Millam, J.M. GaussView5; Semichem. Inc.: Shawnee Mission, UK, 2016. [Google Scholar]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 09, Revision d. 01; Gaussian Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.J.O.; Vasconcelos, C.C.; Garcia, J.B.S.; Pinheiro, M.S.D.; Pereira, F.A.N.; Camelo, D.S.; De Morais, S.V.; Freitas, J.R.B.; Da Rocha, C.Q.; Ribeiro, M.N.D.S.; et al. Anti-Inflammatory and Antioxidant Activity of Pollen Extract Collected by Scaptotrigona affinis postica: In silico, in vitro, and in vivo Studies. Antioxidants 2020, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasconcelos, C.C.; Lopes, A.J.O.; Sousa, E.L.F.; Camelo, D.S.; Lima, F.C.V.M.; Da Rocha, C.Q.; Silva, G.E.B.; Garcia, J.B.S.; Cartágenes, M.D.S.D.S. Effects of Extract of Arrabidaea chica Verlot on an Experimental Model of Osteoarthritis. Int. J. Mol. Sci. 2019, 20, 4717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liston, D.R.; Davis, M. Clinically Relevant Concentrations of Anticancer Drugs: A Guide for Nonclinical Studies. Clin. Cancer Res. 2017, 23, 3489–3498. [Google Scholar] [CrossRef] [Green Version]
- De Souza, S.A.; Camara, C.A.; Da Silva, E.M.S.; Silva, T.M.S. Composition and Antioxidant Activity of Geopropolis Collected by Melipona subnitida (Jandaíra) Bees. Evid. Based Complement. Altern. Med. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Batista, M.C.A.; Abreu, B.V.d.B.; Dutra, R.P.; Cunha, M.S.; do Amaral, F.M.M.; Torres, L.M.B.; Ribeiro, M.N.d.S. Chemical composition and antioxidant activity of geopropolis produced by Melipona fasciculata (Meliponinae) in flooded fields and cerrado areas of MaranhÃ\poundso State, northeastern Brazil. Acta Amaz. 2016, 46, 315–322. [Google Scholar] [CrossRef]
- Al, M.L.; Daniel, D.; Moise, A.R.; Bobis, O.; Laslo, L.; Bogdanov, S. Physico-chemical and bioactive properties of different floral origin honeys from Romania. Food Chem. 2009, 112, 863–867. [Google Scholar] [CrossRef]
- Pauliuc, D.; Dranca, F.; Oroian, M. Antioxidant Activity, Total Phenolic Content, Individual Phenolics and Physicochemical Parameters Suitability for Romanian Honey Authentication. Foods 2020, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Esa, N.M.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxidative Med. Cell. Longev. 2016, 2016, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ninfali, P.; Antonini, E.; Frati, A.; Scarpa, E.S. C-Glycosyl Flavonoids from Beta vulgaris Cicla and Betalains from Beta vulgaris rubra: Antioxidant, Anticancer and Antiinflammatory Activities—A Review. Phytother. Res. 2017, 31, 871–884. [Google Scholar] [CrossRef]
- Liu, B.; Qu, L.; Yan, S. Cyclooxygenase-2 promotes tumor growth and suppresses tumor immunity. Cancer Cell Int. 2015, 15, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoellen, F.; Kelling, K.; Dittmer, C.; Diedrich, K.; Friedrich, M.; Thill, M. Impact of cyclooxygenase-2 in breast cancer. Anticancer Res. 2011, 31, 4359–4367. [Google Scholar] [PubMed]
- Divvela, A.K.C.; Challa, S.R.; Tagaram, I.K. Pathogenic Role of Cyclooxygenase-2 in Cancer. J. Health Sci. 2010, 56, 502–516. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Abrams, J.M.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; Dawson, T.M.; Dawson, V.L.; El-Deiry, W.S.; Fulda, S.; et al. Molecular definitions of cell death subroutines: Recommendations of the Nomenclature Committee on Cell Death 2012. Cell Death Differ. 2011, 19, 107–120. [Google Scholar] [CrossRef]
- Barboza, J.R.; Pereira, F.A.N.; Leite, J.A.C.; Coutinho, D.F.; Ribeiro, M.N.D.S. Abordagem química e toxicidade em modelo zebrafish de geoprópolis de melipona fasciculata Smith. Braz. J. Health Rev. 2019, 2, 5582–5594. [Google Scholar] [CrossRef]
- Suffness, M.; Pezzuto, J. Assays Related to Cancer Drug Discovery. In Methods in Plant Biochemistry: Assays for Bioactivity; Hostettmann, K., Ed.; Academic Press: London, UK, 1990; pp. 71–133. [Google Scholar]
- Trendowski, M. Recent Advances in the Development of Antineoplastic Agents Derived from Natural Products. Drugs 2015, 75, 1993–2016. [Google Scholar] [CrossRef] [Green Version]
- Ghorab, M.M.; Al-Said, M.S.; Samir, N.; Abdel-Latif, G.A.; Soliman, A.M.; Ragab, F.A.; El Ella, D.A.A. Aromatase inhibitors and apoptotic inducers: Design, synthesis, anticancer activity and molecular modeling studies of novel phenothiazine derivatives carrying sulfonamide moiety as hybrid molecules. Eur. J. Med. Chem. 2017, 134, 304–315. [Google Scholar] [CrossRef]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.C.; Arthurton, L.; Baena-Lopez, L.A. Learning on the Fly: The Interplay between Caspases and Cancer. BioMed Res. Int. 2018, 2018, 1–18. [Google Scholar] [CrossRef]
- Satoh, M.S.; Lindahl, T. Role of poly(ADP-ribose) formation in DNA repair. Nature 1992, 356, 356–358. [Google Scholar] [CrossRef]
- Oliver, F.J.; De La Rubia, G.; Rolli, V.; Ruiz-Ruiz, M.C.; De Murcia, G.; Murcia, J.M.-D. Importance of Poly(ADP-ribose) Polymerase and Its Cleavage in Apoptosis. J. Boil. Chem. 1998, 273, 33533–33539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoensinphon, N.; Qiu, P.; Dong, P.; Zheng, J.; Ngauv, P.; Cao, Y.; Li, S.; Ho, C.-T.; Xiao, H. 5-Demethyltangeretin inhibits human nonsmall cell lung cancer cell growth by inducing G2/M cell cycle arrest and apoptosis. Mol. Nutr. Food Res. 2013, 57, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.; Bufalo, M.; Conti, B.; Fernandes, A.R.Y., Jr.; Trusheva, B.; Bankova, V.; Sforcin, J. The chemical composition and pharmacological activities of geopropolis produced by Melipona fasciculata Smith in northeast Brazil. J. Mol. Pathophysiol. 2015, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Yam-Puc, A.; Santana-Hernández, A.A.; Nahuat, P.N.Y.; Ramón-Sierra, J.M.; Farfán, M.R.C.; Borges-Argáez, R.; Ortiz-Vázquez, E. Pentacyclic triterpenes and other constituents in propolis extract from Melipona beecheii collected in Yucatan, México. Rev. Bras. Farm. 2019, 29, 358–363. [Google Scholar] [CrossRef]
- Li, X.; Deng, Y.; Zheng, Z.; Huang, W.; Chen, L.; Tong, Q.; Ming, Y. Corilagin, a promising medicinal herbal agent. Biomed. Pharmacother. 2018, 99, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, S.-L.; Tao, J.-Y.; Pang, R.; Jin, F.; Guo, Y.-J.; Dong, J.-H.; Ye, P.; Zhao, H.-Y.; Zheng, G.-H. Preliminary exploration on anti-inflammatory mechanism of Corilagin (beta-1-O-galloyl-3,6-(R)-hexahydroxydiphenoyl-d-glucose) in vitro. Int. Immunopharmacol. 2008, 8, 1059–1064. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, A.K.; Kumar, R.; Ganguly, R.; Rana, H.K.; Pandey, P.K.; Sethi, G.; Bishayee, A.; Pandey, A. Corilagin in Cancer: A Critical Evaluation of Anticancer Activities and Molecular Mechanisms. Mol. 2019, 24, 3399. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, K.; Zhang, H.; Dong, W.; Peng, W.; Zhao, Y. Effect of typhaneoside on ventricular remodeling and regulation of PI3K/Akt/mTOR pathway. Herz 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.-D.; Cao, Y.; Bao, B.; Zhang, L.; Ding, A. Antioxidant capacity of Typha angustifolia extracts and two active flavonoids. Pharm. Boil. 2017, 55, 1283–1288. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Gu, D.; Zeng, H. Antitumor effects of beta-amyrin in Hep-G2 liver carcinoma cells are mediated via apoptosis induction, cell cycle disruption and activation of JNK and P38 signalling pathways. J. BUON 2018, 23, 965–970. [Google Scholar]
- Askari, V.R.; Fereydouni, N.; Rahimi, V.B.; Askari, N.; Sahebkar, A.H.; Rahmanian-Devin, P.; Kermani, A.S. β-Amyrin, the cannabinoid receptors agonist, abrogates mice brain microglial cells inflammation induced by lipopolysaccharide/interferon-γ and regulates Mφ1/Mφ2 balances. Biomed. Pharmacother. 2018, 101, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.; Mathew, L.E.; Vijayalakshmi, N.R.; Helen, A. Anti-inflammatory potential of β-amyrin, a triterpenoid isolated from Costus igneus. Inflammopharmacology 2014, 22, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Grösch, S.; Maier, T.J.; Schiffmann, S.; Geisslinger, G. Cyclooxygenase-2 (COX-2)–Independent Anticarcinogenic Effects of Selective COX-2 Inhibitors. J. Natl. Cancer Inst. 2006, 98, 736–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortazavi, H.; Nikfar, B.; Esmaeili, S.-A.; Rafieenia, F.; Saburi, E.; Chaichian, S.; Gorji, A.M.H.; Momtazi-Borojeni, A.A. Potential cytotoxic and anti-metastatic effects of berberine on gynaecological cancers with drug-associated resistance. Eur. J. Med. Chem. 2020, 187, 111951. [Google Scholar] [CrossRef]
- Rushe, M.; Silvian, L.F.; Bixler, S.; Chen, L.L.; Cheung, A.; Bowes, S.; Cuervo, H.; Berkowitz, S.; Zheng, T.; Guckian, K.; et al. Structure of a NEMO/IKK-Associating Domain Reveals Architecture of the Interaction Site. Structure 2008, 16, 798–808. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.-C.; Dong, S.; Zhang, S.-Y.; Jia, N.; Ou, S.-L. Taraxerone triterpene inhibits cancer cell growth by inducing apoptosis in non-small cell lung cancer cells. Bangladesh J. Pharmacol. 2016, 11, 342. [Google Scholar] [CrossRef] [Green Version]
- Mo, E.K.; Han, B.H.; Kim, S.M.; Yang, S.A.; Kang, S.K.; Oh, C.J.; Kim, R.; Kim, C.G.; Kang, H.J.; Sung, C.K. Identification of d-friedoolean-13-en-3-one (Taraxerone) as an antioxidant compound from sedom (Sedum sarmentosum). Food Sci. Biotechnol. 2012, 21, 485–489. [Google Scholar] [CrossRef]
- Patil, K.R.; Mohapatra, P.; Patel, H.M.; Goyal, S.N.; Ojha, S.; Kundu, C.N.; Patil, C.R. Pentacyclic Triterpenoids Inhibit IKKβ Mediated Activation of NF-κB Pathway: In Silico and In Vitro Evidences. PLoS ONE 2015, 10, e0125709. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Lehtonen, M.; Suuronen, T.; Kaarniranta, K.; Huuskonen, J. Terpenoids: Natural inhibitors of NF-κB signaling with anti-inflammatory and anticancer potential. Cell. Mol. Life Sci. 2008, 65, 2979–2999. [Google Scholar] [CrossRef]
- Hodon, J.; Borkova, L.; Pokorny, J.; Kazakova, A.; Urban, M. Design and synthesis of pentacyclic triterpene conjugates and their use in medicinal research. Eur. J. Med. Chem. 2019, 182, 111653. [Google Scholar] [CrossRef]
- Laszczyk, M.N. Pentacyclic Triterpenes of the Lupane, Oleanane and Ursane Group as Tools in Cancer Therapy. Planta Med. 2009, 75, 1549–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirupathi, A.; Silveira, P.; Nesi, R.; Pinho, R.A. Amyrin, a pentacyclic triterpene, exhibits anti-fibrotic, anti-inflammatory, and anti-apoptotic effects on dimethyl nitrosamine–induced hepatic fibrosis in male rats. Hum. Exp. Toxicol. 2016, 36, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-signaling pathway in cancer. Onco Targets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Shen, S.; Verma, I.M. NF-B, An Active Player in Human Cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [Green Version]
- Prescott, J.A.; Cook, S.J. Targeting IKKβ in Cancer: Challenges and Opportunities for the Therapeutic Utilisation of IKKβ Inhibitors. Cells 2018, 7, 115. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.; Edwards, J.; Pepper, C.; Mackay, S.P. Inhibitory-κB Kinase (IKK) α and Nuclear Factor-κB (NFκB)-Inducing Kinase (NIK) as Anti-Cancer Drug Targets. Cells 2018, 7, 176. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | DPPH• IC50 (μg/mL) | FRAP (mmol Fe2+/g) | ABTS+ IC50 (μg/mL) |
|---|---|---|---|
| EHGV | 76.16 ± 1.05 a | 2.91 ± 0.12 a | 13.28 ± 0.11 a |
| EHGP | 265.91 ± 0.29 b | 1.10 ± 0.25 b | 58.94 ± 0.09 b |
| Trolox | 3.01 ± 0.47 | 8.41 ± 0.28 | 3.69 ± 0.63 |
| Time | Sample | Cell Line | ||||
|---|---|---|---|---|---|---|
| A2780 | ES2 | H460 | A549 | HUVEC | ||
| 48 h | EHGV | 313.6 μg/mL | 133.1 μg/mL | 105.4 μg/mL | 105.4 μg/mL | 113,300 μg/mL |
| EHGP | 177.4 μg/mL | 51.4 μg/mL | 360.6 μg/mL | 1122 μg/mL | 40,650 μg/mL | |
| 72 h | EHGV | 16.92 μg/mL | 137.7 μg/mL | 56.51 μg/mL | 22.64 μg/mL | 5537 μg/mL |
| EHGP | 196 μg/mL | 64.83 μg/mL | 311.9 μg/mL | 551.1 μg/mL | indeterminate | |
| Compound | RT (min) | Identification | MW | [M − H]− (m/z) | MS/MS Fragments (m/z) |
|---|---|---|---|---|---|
| 1 | 2.8 | gluconic acid | 196 | 195 | 128; 177 |
| 2 | 3.1 | corilagin | 634 | 633 | 615; 484 |
| 3 | 15.9 | taraxerone | 424 | 423 | 304; 334; 406 |
| 4 | 18.2 | myricetin-3-O-α-arabinopyranoside | 450 | 449 | 430; 359; 329 |
| 5 | 19.1 | prunin | 434 | 433 | 313 |
| 6 | 20.5 | dipterocarpol | 443 | 442 | 209; 165 |
| 7 | 24 | taxifolin 7-O-rhamnoside | 450 | 449 | 405 |
| 8 | 24.7 | isoschaftoside | 564 | 563 | 548; 298 |
| 9 | 25.3 | marsformosanone | 422 | 421 | 377; 333; 297; 214; 179; 157 |
| 10 | 32 | β-amyrin | 427 | 426 | 232 |
| 11 | 40 | typhaneoside | 770 | 769 | 375; 331 |
| 12 | 44.5 | 3-[xyl]-28-glc-phytolaccagenin | 826 | 825 | 403; 360 |
| Compound | RT (min) | Identification | MW | [M − H]− (m/z) | MS/MS Fragments (m/z) |
|---|---|---|---|---|---|
| 1 | 2.8 | gluconic acid | 196 | 195 | 128; 177 |
| 2 | 16 | taraxerone | 424 | 423 | 304; 334; 364; 406 |
| 3 | 17.8 | dihydroquercetin-C-glycoside | 450 | 449 | 431; 359; 329; 287; 303 |
| 4 | 18.2 | dihydroquercetin-C-glycoside isomer | 450 | 449 | 430; 359; 329 |
| 5 | 19.1 | narigenin-C-glycoside | 434 | 433 | 313 |
| 6 | 20.2 | narigenin-C-glycoside isomer | 434 | 433 | 415; 313 |
| 7 | 21 | vitexin-O-galate | 584 | 583 | 169; 313; 932; 537 |
| 8 | 22.5 | pinobanksin glycosilated | 436 | 435 | 270; 151; 341; 391 |
| 9 | 22,9 | dihydroquercetin 3-O-ramnoside | 450 | 449 | 303; 405 |
| 10 | 33.8 | xantholaccaic acid A | 521 | 520 | 262; 357; 419; 458; 502; 542 |
| 11 | 42.5 | gallocatequin-xylose | 438 | 437 | 305; 357; 393; 437 |
| COX-2 | NF-κB | ||
|---|---|---|---|
| Ligand | ΔGbind (kcal/mol) | Ligand | ΔGbind (kcal/mol) |
| corilagin | −9.3 | taraxerone | −8.4 |
| typhaneoside | −8.8 | marsformosanone | −7.7 |
| β-amyrin | −8.7 | β-amyrin | −7.4 |
| isoschaftoside | −8.6 | dipterocarpol | −6.9 |
| 3-[xyl]-28-glc-phytolaccagenin | −8.5 | 3-[xyl]-28-glc-phytolaccagenin | −6.9 |
| marsformosanone | −8.5 | prunin | −6.8 |
| taraxerone | −8.3 | corilagin | −6.6 |
| prunin | −8.0 | typhaneoside | −6.5 |
| myricetin-3-O-α-arabinopyranoside | −7.9 | isoschaftoside | −6.4 |
| dipterocarpol | −7.7 | myricetin-3-O-α-arabinopyranoside | −6.3 |
| taxifolin 7-O-rhamnoside | −7.6 | taxifolin 7-O-rhamnoside | −6.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barboza, J.R.; Pereira, F.A.N.; Fernandes, R.A.; Vasconcelos, C.C.; Cartágenes, M.d.S.d.S.; Oliveira Lopes, A.J.; Melo, A.C.d.; Guimarães, I.d.S.; Rocha, C.Q.d.; Ribeiro, M.N.d.S. Cytotoxicity and Pro-Apoptotic, Antioxidant and Anti-Inflammatory Activities of Geopropolis Produced by the Stingless Bee Melipona fasciculata Smith. Biology 2020, 9, 292. https://doi.org/10.3390/biology9090292
Barboza JR, Pereira FAN, Fernandes RA, Vasconcelos CC, Cartágenes MdSdS, Oliveira Lopes AJ, Melo ACd, Guimarães IdS, Rocha CQd, Ribeiro MNdS. Cytotoxicity and Pro-Apoptotic, Antioxidant and Anti-Inflammatory Activities of Geopropolis Produced by the Stingless Bee Melipona fasciculata Smith. Biology. 2020; 9(9):292. https://doi.org/10.3390/biology9090292
Chicago/Turabian StyleBarboza, Josianne Rocha, Francisco Assis Nascimento Pereira, Renan Amphilophio Fernandes, Cleydlenne Costa Vasconcelos, Maria do Socorro de Sousa Cartágenes, Alberto Jorge Oliveira Lopes, Andreia Cristina de Melo, Isabella dos Santos Guimarães, Cláudia Quintino da Rocha, and Maria Nilce de Sousa Ribeiro. 2020. "Cytotoxicity and Pro-Apoptotic, Antioxidant and Anti-Inflammatory Activities of Geopropolis Produced by the Stingless Bee Melipona fasciculata Smith" Biology 9, no. 9: 292. https://doi.org/10.3390/biology9090292