
Augmentation of Performance, Carcass Trait, Biochemical Profile and Lipid Metabolism Concerning the Use of Organic Acidifier in Broiler Chickens

 ,
, 

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Additive and Experimental Design
2.2. Bird Management and Data Recording
2.3. Carcass Traits, Meat Quality and Sample Collection
2.4. Quantitative Real-Time PCR Analysis
2.5. Detection Assays Utilizing Commercial Kits
2.6. Statistical Analysis
3. Results
3.1. Performance, Carcass Traits and Meat Quality
3.2. Serum Indexes Related to Lipid Metabolism, Liver Health, Antioxidant Capacity, and Immunity
3.3. Jejunum Immunity
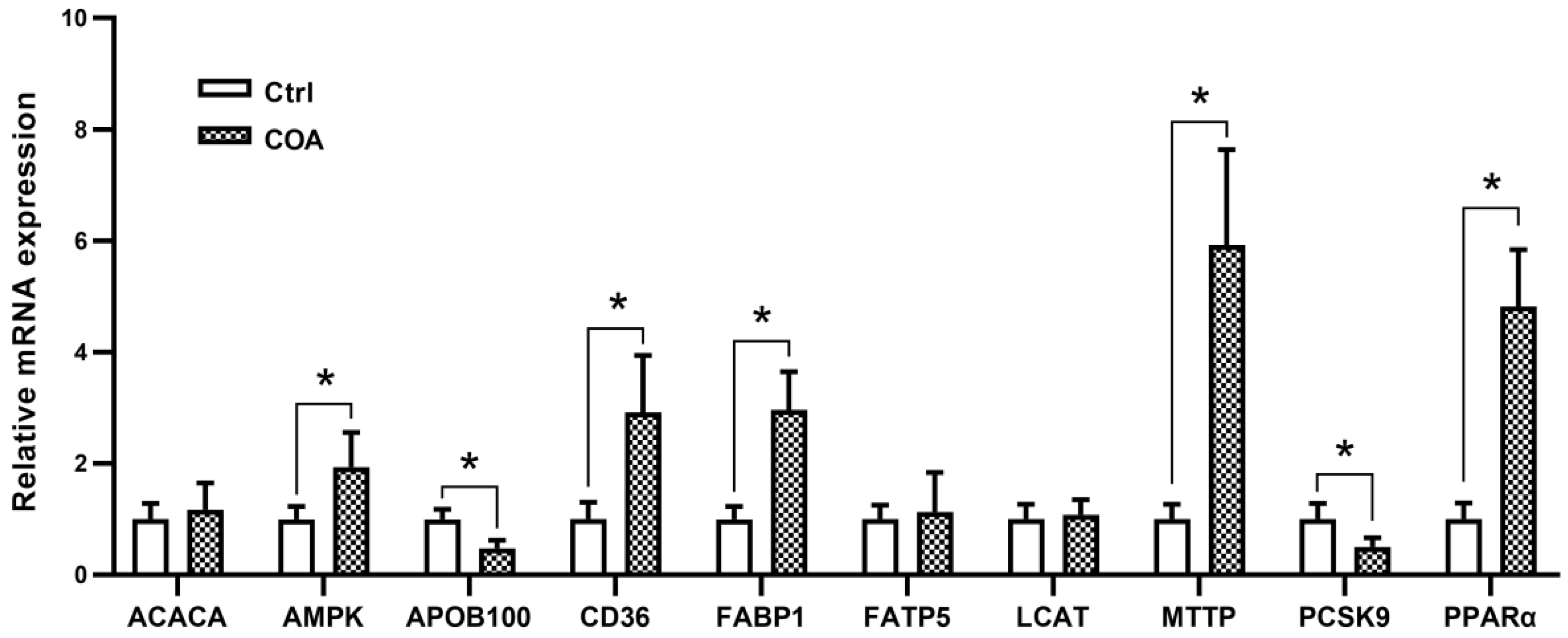
3.4. Liver Lipid Metabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Neveling, D.P.; Dicks, L.M.T. Probiotics: An antibiotic replacement strategy for healthy broilers and productive rearing. Probiot. Antimicrob. Proteins 2021, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Andrew Selaledi, L.; Mohammed Hassan, Z.; Manyelo, T.G.; Mabelebele, M. The current status of the alternative use to antibiotics in poultry production: An african perspective. Antibiotics 2020, 9, 594. [Google Scholar] [CrossRef]
- Zhu, Q.; Sun, P.; Zhang, B.; Kong, L.; Xiao, C.; Song, Z. Progress on gut health maintenance and antibiotic alternatives in broiler chicken production. Front. Nutr. 2021, 8, 692839. [Google Scholar] [CrossRef]
- Dai, D.; Qiu, K.; Zhang, H.J.; Wu, S.G.; Han, Y.M.; Wu, Y.Y.; Qi, G.H.; Wang, J. Organic acids as alternatives for antibiotic growth promoters alter the intestinal structure and microbiota and improve the growth performance in broilers. Front. Microbiol. 2020, 11, 618144. [Google Scholar] [CrossRef]
- Sun, H.Y.; Zhou, H.B.; Liu, Y.; Wang, Y.; Zhao, C.; Xu, L.M. Comparison of organic acids supplementation on the growth performance, intestinal characteristics and morphology, and cecal microflora in broilers fed corn-soybean meal diet. Anim. Biosci. 2022, 35, 1689–1697. [Google Scholar] [CrossRef]
- Sun, Y.; Ni, A.; Jiang, Y.; Li, Y.; Huang, Z.; Shi, L.; Xu, H.; Chen, C.; Li, D.; Han, Y.; et al. Effects of replacing in-feed antibiotics with synergistic organic acids on growth performance, health, carcass, and immune and oxidative statuses of broiler chickens under clostridium perfringens type a challenge. Avian Dis. 2020, 64, 393–400. [Google Scholar] [CrossRef]
- Ateya, A.I.; Arafat, N.; Saleh, R.M.; Ghanem, H.M.; Naguib, D.; Radwan, H.A.; Elseady, Y.Y. Intestinal gene expressions in broiler chickens infected with Escherichia coli and dietary supplemented with probiotic, acidifier and synbiotic. Vet. Res. Commun. 2019, 43, 131–142. [Google Scholar] [CrossRef]
- Li, D.; Zhang, K.; Pan, Z.; Yu, M.; Lu, Y.; Wang, G.; Wu, J.; Zhang, J.; Zhang, K.; Du, W. Antibiotics promote abdominal fat accumulation in broilers. Anim. Sci. J. 2020, 91, e13326. [Google Scholar] [CrossRef]
- Gao, C.Q.; Shi, H.Q.; Xie, W.Y.; Zhao, L.H.; Zhang, J.Y.; Ji, C.; Ma, Q.G. Dietary supplementation with acidifiers improves the growth performance, meat quality and intestinal health of broiler chickens. Anim. Nutr. 2021, 7, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Sharİf, M.; Suleman, M.N.; Saeed, M.; Ahamd, F.; Kamboh, A.A.; Ayaşan, T.; Arslan, M. Effect of dietary supplementation of a nonantibiotic growth promoter on growth performance and intestinal histomorphology in broilers. Pak. J. Zool. 2022, 54, 1629–1636. [Google Scholar] [CrossRef]
- Hamid, H.; Shi, H.Q.; Ma, G.Y.; Fan, Y.; Li, W.X.; Zhao, L.H.; Zhang, J.Y.; Ji, C.; Ma, Q.G. Influence of acidified drinking water on growth performance and gastrointestinal function of broilers. Poult. Sci. 2018, 97, 3601–3609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guo, Y.; Wang, Z.; Wang, Y.; Chen, B.; Du, P.; Zhang, X.; Huang, Y.; Li, P.; Michiels, J.; et al. Acidification of drinking water improved tibia mass of broilers through the alterations of intestinal barrier and microbiota. Anim. Biosci. 2022, 35, 902–915. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Lin, X.; Gou, Z.; Fan, Q.; Ye, J.; Jiang, S. Potential effects of acidifier and amylase as substitutes for antibiotic on the growth performance, nutrient digestion and gut microbiota in yellow-feathered broilers. Animals 2020, 10, 1858. [Google Scholar] [CrossRef]
- Arif, M.; Alagawany, M.; Abd El-Hack, M.E.; Saeed, M.; Arain, M.A.; Elnesr, S.S. Humic acid as a feed additive in poultry diets: A review. Iran J. Vet. Res. 2019, 20, 167–172. [Google Scholar]
- Adhikari, P.; Yadav, S.; Cosby, D.E.; Cox, N.A.; Jendza, J.A.; Kim, W.K. Research Note: Effect of organic acid mixture on growth performance and Salmonella Typhimurium colonization in broiler chickens. Poult. Sci. 2020, 99, 2645–2649. [Google Scholar] [CrossRef]
- Lu, T.; Marmion, M.; Ferone, M.; Wall, P.; Scannell, A.G.M. On farm interventions to minimise Campylobacter spp. contamination in chicken. Br. Poult. Sci. 2021, 62, 53–67. [Google Scholar] [CrossRef]
- Peh, E.; Kittler, S.; Reich, F.; Kehrenberg, C. Antimicrobial activity of organic acids against Campylobacter spp. and development of combinations-A synergistic effect? PLoS ONE 2020, 15, e0239312. [Google Scholar] [CrossRef]
- Pham, V.H.; Abbas, W.; Huang, J.; He, Q.; Zhen, W.; Guo, Y.; Wang, Z. Effect of blending encapsulated essential oils and organic acids as an antibiotic growth promoter alternative on growth performance and intestinal health in broilers with necrotic enteritis. Poult. Sci. 2022, 101, 101563. [Google Scholar] [CrossRef]
- Sevim, B.; Gümüş, E.; Harman, H.; Ayasan, T.; Başer, E.; Altay, Y.; Akbulut, K. Effects of dietary rosemary essential oil on growth performance, carcass traits and some hematological values of chukar partridge. Turk. J. Agric. Food Sci. Technol. 2020, 8, 430–435. [Google Scholar] [CrossRef]
- Stamilla, A.; Russo, N.; Messina, A.; Spadaro, C.; Natalello, A.; Caggia, C.; Randazzo, C.L.; Lanza, M. Effects of microencapsulated blend of organic acids and essential oils as a feed additive on quality of chicken breast meat. Animals 2020, 10, 640. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Wu, Y.; Lei, Z.; Zhang, M.; Xie, Z.; Tang, S. Lactic acid, a driver of tumor-stroma interactions. Int. Immunopharmacol. 2022, 106, 108597. [Google Scholar] [CrossRef] [PubMed]
- Choe, U.; Sun, J.; Wang, T.T.Y.; Rao, J.; Yu, L.L.; Gao, B.; Bailoni, E.; Chen, P.; Li, Y. Chemical composition of tomato seed flours, and their radical scavenging, anti-inflammatory and gut microbiota modulating properties. Molecules 2021, 26, 1478. [Google Scholar] [CrossRef] [PubMed]
- Qiu, K.; He, W.; Zhang, H.; Wang, J.; Qi, G.; Guo, N.; Zhang, X.; Wu, S. Bio-fermented malic acid facilitates the production of high-quality chicken via enhancing muscle antioxidant capacity of broilers. Antioxidants 2022, 11, 2309. [Google Scholar] [CrossRef]
- Kim, Y.J.; Youk, S.; Song, C.S. Effectiveness of administering a mixture of lactic acid bacteria to control Salmonella ser. Enteritidis infections in broilers. Animals 2022, 12, 374. [Google Scholar] [CrossRef]
- Qiu, K.; Zhao, Q.; Wang, J.; Qi, G.-H.; Wu, S.-G.; Zhang, H.-J. Effects of pyrroloquinoline quinone on lipid metabolism and anti-oxidative capacity in a high-fat-diet metabolic dysfunction-associated fatty liver disease chick model. Int. J. Mol. Sci. 2021, 22, 1458. [Google Scholar] [CrossRef]
- Qiu, K.; Wang, X.; Zhang, H.; Wang, J.; Qi, G.; Wu, S. Dietary supplementation of a new probiotic compound improves the growth performance and health of broilers by altering the composition of cecal microflora. Biology 2022, 11, 633. [Google Scholar] [CrossRef]
- Khan, R.U.; Naz, S.; Raziq, F.; Qudratullah, Q.; Khan, N.A.; Laudadio, V.; Tufarelli, V.; Ragni, M. Prospects of organic acids as safe alternative to antibiotics in broiler chickens diet. Environ. Sci. Pollut. Res. Int. 2022, 29, 32594–32604. [Google Scholar] [CrossRef]
- Sugiharto, S.; Ranjitkar, S. Recent advances in fermented feeds towards improved broiler chicken performance, gastrointestinal tract microecology and immune responses: A review. Anim. Nutr. 2019, 5, 1–10. [Google Scholar] [CrossRef]
- Al-Ghamdi, E.S. Growth performance, carcass characteristics, and blood biochemical indices of broilers affected by dietary organic acids blend’s supplementation. Anim. Biotechnol. 2022, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Singh, A.K.; Chen, X.; Lv, J.; Kim, W.K. Application of organic acids and essential oils as alternatives to antibiotic growth promoters in broiler chickens. Animals 2022, 12, 2178. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Toghyani, M.; Kheravii, S.K.; Pineda, L.; Han, Y.; Swick, R.A.; Wu, S.B. Potential of blended organic acids to improve performance and health of broilers infected with necrotic enteritis. Anim. Nutr. 2021, 7, 440–449. [Google Scholar] [CrossRef]
- Negm, M.H.; Aboul Ela, S.S.; Abd El-Hack, M.E. Does the use of lactic acid as an antibiotic substitute in broiler diets affect growth, carcass traits, blood indices and intestinal microbiota? Anim. Biotechnol. 2023, 34, 1163–1169. [Google Scholar] [CrossRef]
- Manvatkar, P.N.; Kulkarni, R.C.; Awandkar, S.P.; Chavhan, S.G.; Durge, S.M.; Avhad, S.R.; Channa, G.R.; Kulkarni, M.B. Performance of broiler chicken on dietary supplementation of protected organic acids blend. Br. Poult. Sci. 2022, 63, 633–640. [Google Scholar] [CrossRef]
- Liu, N.; Ding, K.; Wang, J.; Deng, Q.; Gu, K.; Wang, J. Effects of lactic acid bacteria and smectite after aflatoxin B(1) challenge on the growth performance, nutrient digestibility and blood parameters of broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 953–961. [Google Scholar] [CrossRef]
- Chen, L.; Duan, Y.; Wei, H.; Ning, H.; Bi, C.; Zhao, Y.; Qin, Y.; Li, Y. Acetyl-CoA carboxylase (ACC) as a therapeutic target for metabolic syndrome and recent developments in ACC1/2 inhibitors. Expert Opin. Investig. Drugs 2019, 28, 917–930. [Google Scholar] [CrossRef]
- Fang, C.; Pan, J.; Qu, N.; Lei, Y.; Han, J.; Zhang, J.; Han, D. The AMPK pathway in fatty liver disease. Front. Physiol. 2022, 13, 970292. [Google Scholar] [CrossRef]
- Garside, B.; Ho, J.H.; Kwok, S.; Liu, Y.; Dhage, S.; Donn, R.; Iqbal, Z.; Jones, S.A.; Soran, H. Changes in PCSK 9 and apolipoprotein B100 in Niemann-Pick disease after enzyme replacement therapy with olipudase alfa. Orphanet J. Rare Dis. 2021, 16, 107. [Google Scholar] [CrossRef]
- Li, Y.; Huang, X.; Yang, G.; Xu, K.; Yin, Y.; Brecchia, G.; Yin, J. CD36 favours fat sensing and transport to govern lipid metabolism. Prog. Lipid Res. 2022, 88, 101193. [Google Scholar] [CrossRef]
- Rodriguez Sawicki, L.; Bottasso Arias, N.M.; Scaglia, N.; Falomir Lockhart, L.J.; Franchini, G.R.; Storch, J.; Córsico, B. FABP1 knockdown in human enterocytes impairs proliferation and alters lipid metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zhang, G.; Zheng, Y.; Yang, Y.; Chen, C.; Xia, J.; Liang, L.; Lei, C.; Hu, Y.; Cai, X.; et al. SLC27A5 deficiency activates NRF2/TXNRD1 pathway by increased lipid peroxidation in HCC. Cell Death Differ. 2020, 27, 1086–1104. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, J.; Xiang, H.; Ding, P.; Wu, T.; Ji, G. LCAT- targeted therapies: Progress, failures and future. Biomed. Pharmacother. 2022, 147, 112677. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Chiu, T.Y.; Liang, Y.J.; Lee, C.J.; Liu, C.S.; Suen, C.S.; Yen, J.J.; Chen, H.T.; Hwang, M.J.; Hussain, M.M.; et al. PRAP1 is a novel lipid-binding protein that promotes lipid absorption by facilitating MTTP-mediated lipid transport. J. Biol. Chem. 2021, 296, 100052. [Google Scholar] [CrossRef]
- Pawlak, M.; Lefebvre, P.; Staels, B. Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 2015, 62, 720–733. [Google Scholar] [CrossRef]

{kind=link}
| Items | Day 1~14 | Day 15~28 | Day 29~42 |
|---|---|---|---|
| Ingredients, % | |||
| Corn | 58.20 | 58.70 | 61.00 |
| Soybean meal | 34.85 | 33.02 | 30.85 |
| Soybean oil | 2.54 | 4.48 | 4.72 |
| CaHPO4 | 2.27 | 2.00 | 1.82 |
| CaCO3 | 0.82 | 0.72 | 0.70 |
| NaCl | 0.35 | 0.35 | 0.35 |
| DL-Met | 0.34 | 0.26 | 0.20 |
| L-Lys·HCl | 0.31 | 0.15 | 0.04 |
| Premix 1 | 0.22 | 0.22 | 0.22 |
| Choline chloride (50%) | 0.10 | 0.10 | 0.10 |
| In total | 100.00 | 100.00 | 100.00 |
| Nutrient levels 2 | |||
| AME, MJ/kg | 12.35 | 12.97 | 13.18 |
| CP, % | 22.00 | 21.00 | 20.00 |
| Calcium, % | 1.00 | 1.00 | 0.90 |
| AP, % | 0.50 | 0.45 | 0.40 |
| Lys, % | 1.29 | 1.15 | 1.09 |
| Met, % | 0.55 | 0.52 | 0.48 |
| Met + Cys, % | 0.94 | 0.84 | 0.84 |
| Thr, % | 0.82 | 0.77 | 0.69 |
| Trp, % | 0.24 | 0.22 | 0.18 |
| Gene Name | Sequence (5′-3′) | Annealing Temperature (°C) | Product Size (bp) |
|---|---|---|---|
| ACACA | F: CCAGGCACAGTCTTTAGGGA | 60 | 208 |
| R: AATCTCGTTTCCTCCTGGCA | |||
| AMPK | F: CAAGCCCGCCAGATTCTTTT | 59 | 184 |
| R: CGACTCTGACTCCGTATCCC | |||
| APOB100 | F: AAATGTCCAAGGTGCAGCAG | 59 | 185 |
| R: CAATCAGCTTCCCGTTACCG | |||
| CD36 | F: ACACTGGGTAGCTGCAGAAT | 59 | 162 |
| R: GCTGCAGTCTTCCAAGCTTT | |||
| FABP1 | F: AAAGTACGAGCTCCAGTCCC | 61 | 227 |
| R: ACTTGGCCTTCTCTCCTGTC | |||
| FATP5 | F: GGGACCTTCAAGCAATGCAA | 59 | 158 |
| R: TACAGGTTGAGGCTCCCATC | |||
| GAPDH | F: AGCCATTCCTCCACCTTTGA | 61 | 190 |
| R: CAACAAAGGGTCCTGCTTCC | |||
| LCAT | F: TGTGGGAGGACATGAAGGAC | 59 | 184 |
| R: TCTGAGCTGCGTTTGTTGAC | |||
| MTTP | F: CCCATTAGCGTCGTGAAAGG | 59 | 174 |
| R: CCGGTTCTTGACGTTGGTTT | |||
| PCSK9 | F: GGTTGCTCCCTACTGTCCTT | 61 | 232 |
| R: CCAGCAGGTCCTTAACTCCA | |||
| PPARα | F: AGTGACCGCTCTACTTGACC | 61 | 250 |
| R: GATGCTGGTGAAAGGGTGTC |
| Ctrl | COA | SEM | p-Value | |
|---|---|---|---|---|
| Body Weight, g | ||||
| d 0 | 43.91 | 43.89 | 0.06 | 0.992 |
| d 14 | 537.48 | 561.14 | 13.45 | <0.001 |
| d 28 | 1704.44 | 1785.76 | 0.04 | <0.001 |
| d 42 | 3189.91 | 3250.06 | 0.09 | 0.040 |
| Weeks 1~2 | ||||
| ADG, g | 38.28 | 39.94 | 0.98 | <0.001 |
| ADFI, g | 43.65 | 43.14 | 1.05 | 0.144 |
| FCR, g/g | 1.14 | 1.08 | 0.01 | <0.001 |
| DCR, % | 0.01 | 0.01 | 0.02 | 0.609 |
| Weeks 3~4 | ||||
| ADG, g | 83.12 | 87.36 | 2.38 | <0.001 |
| ADFI, g | 122.12 | 127.13 | 2.82 | <0.001 |
| FCR, g/g | 1.47 | 1.46 | 0.02 | 0.035 |
| DCR, % | 0.02 | 0.02 | 0.03 | 0.818 |
| Weeks 5–6 | ||||
| ADG, g | 105.84 | 104.19 | 4.56 | 0.273 |
| ADFI, g | 181.86 | 182.09 | 5.24 | 0.896 |
| FCR, g/g | 1.72 | 1.75 | 0.06 | 0.121 |
| DCR, % | 0.03 | 0.03 | 0.04 | 0.617 |
| Weeks 1~6 | ||||
| ADG, g | 75.75 | 77.16 | 1.98 | 0.034 |
| ADFI, g | 115.88 | 117.45 | 2.51 | 0.061 |
| FCR, g/g | 1.44 | 1.43 | 0.02 | 0.031 |
| DCR, % | 0.05 | 0.06 | 0.05 | 0.427 |
| Item, % | Ctrl | COA | SEM | p-Value |
|---|---|---|---|---|
| Dressing percentage | 91.73 | 91.75 | 0.61 | 0.969 |
| Half evisceration | 84.79 | 84.38 | 1.25 | 0.270 |
| Full evisceration | 77.95 | 77.60 | 1.34 | 0.373 |
| Abdominal fat | 1.49 | 1.48 | 0.59 | 0.875 |
| Breast muscle | 29.65 | 29.61 | 1.22 | 0.630 |
| Leg muscle | 29.49 | 29.64 | 1.41 | 0.980 |
| Dropping loss | 4.34 | 3.82 | 0.43 | 0.032 |
| Ctrl | COA | SEM | p-Value | |
|---|---|---|---|---|
| TC, mmol/L | 2.98 | 3.05 | 0.55 | 0.803 |
| TG, mmol/L | 19.95 | 17.18 | 1.80 | 0.042 |
| HDL, mmol/L | 0.84 | 1.07 | 0.14 | 0.003 |
| VLDL, mmol/L | 0.65 | 0.58 | 0.13 | 0.280 |
| AST, U/L | 224.53 | 216.80 | 18.24 | 0.380 |
| ALT, U/L | 2.40 | 2.35 | 0.75 | 0.878 |
| TBil, μmol/L | 4.43 | 4.01 | 2.19 | 0.693 |
| Ctrl | COA | SEM | p-Value | |
|---|---|---|---|---|
| CAT, U/mL | 40.34 | 43.36 | 8.12 | 0.457 |
| SOD, U/mL | 69.75 | 77.33 | 6.47 | 0.026 |
| GSH-PX, U/mL | 427.08 | 390.27 | 53.48 | 0.162 |
| T-AOC, U/mL | 5.36 | 6.46 | 1.03 | 0.043 |
| MDA, nmol/mL | 4.69 | 4.46 | 0.94 | 0.607 |
| Ctrl | COA | SEM | p-Value | |
|---|---|---|---|---|
| IgA, g/L | 1.45 | 1.88 | 0.42 | 0.042 |
| IgM, g/L | 1.50 | 1.41 | 0.39 | 0.600 |
| IgG, g/L | 3.75 | 4.13 | 0.43 | 0.078 |
| C3, g/L | 0.51 | 0.61 | 0.06 | 0.049 |
| C4, g/L | 0.07 | 0.07 | 0.02 | 0.398 |
| Lysozyme, mg/L | 3.54 | 4.12 | 0.27 | 0.026 |
| Ctrl | COA | SEM | p-Value | |
|---|---|---|---|---|
| IL-2, ng/mL | 5.28 | 3.31 | 0.54 | 0.034 |
| IL-6, ng/mL | 0.67 | 0.74 | 0.16 | 0.713 |
| TNF-α, ng/mL | 1.84 | 1.33 | 0.21 | 0.027 |
| sIgA, µg/mL | 31.92 | 33.47 | 5.28 | 0.419 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, K.; Chen, Z.; Zheng, A.; Chang, W.; Cai, H.; Zhang, X.; Liu, G. Augmentation of Performance, Carcass Trait, Biochemical Profile and Lipid Metabolism Concerning the Use of Organic Acidifier in Broiler Chickens. Agriculture 2023, 13, 1765. https://doi.org/10.3390/agriculture13091765
Qiu K, Chen Z, Zheng A, Chang W, Cai H, Zhang X, Liu G. Augmentation of Performance, Carcass Trait, Biochemical Profile and Lipid Metabolism Concerning the Use of Organic Acidifier in Broiler Chickens. Agriculture. 2023; 13(9):1765. https://doi.org/10.3390/agriculture13091765
Chicago/Turabian StyleQiu, Kai, Zhimin Chen, Aijuan Zheng, Wenhuan Chang, Huiyi Cai, Xin Zhang, and Guohua Liu. 2023. "Augmentation of Performance, Carcass Trait, Biochemical Profile and Lipid Metabolism Concerning the Use of Organic Acidifier in Broiler Chickens" Agriculture 13, no. 9: 1765. https://doi.org/10.3390/agriculture13091765
APA StyleQiu, K., Chen, Z., Zheng, A., Chang, W., Cai, H., Zhang, X., & Liu, G. (2023). Augmentation of Performance, Carcass Trait, Biochemical Profile and Lipid Metabolism Concerning the Use of Organic Acidifier in Broiler Chickens. Agriculture, 13(9), 1765. https://doi.org/10.3390/agriculture13091765