
Urolithin A-Enhanced Multi-Bioactive Formulation Mitigates Cyclophosphamide-Induced Premature Ovarian Failure Through Suppression of Oxidative-Inflammatory Stress and Preservation of Follicle Fate

 ,
,
Highlights
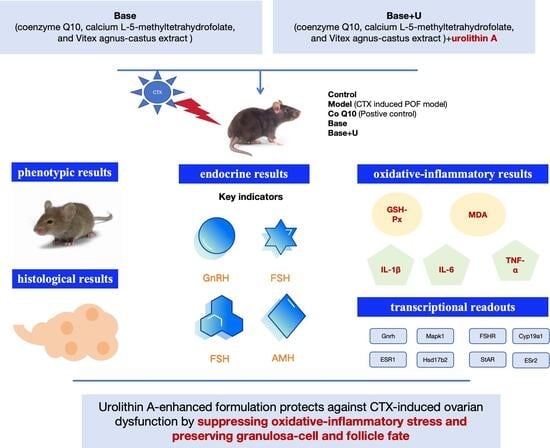
- CTX induced a POF-like phenotype with follicular atresia and endocrine disruption.
- CoQ10 and the designed formulations (Base and Base + U) improved cyclicity, ovarian histology, and hormone profiles.
- The treatments alleviated ovarian oxidative-inflammatory stress (GSH-Px↑, MDA↓, cytokines↓).
- Ovarian functional transcripts were restored (Fshr, Esr1, Hsd17b2) by the treatments.
- Urolithin A addition (Base + U) showed an incremental benefit on lipid peroxidation/inflammation.
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Study Formulations
2.3. Animals and Treatment
2.4. Evaluation of Ovarian Index
2.5. Vaginal Cytology and Estrous Cycle Staging
2.6. Serum Hormone Measurement
2.7. Histological Analysis
2.8. Terminal Deoxynucleotidyl Transferase dUTP Nick-End Labeling (TUNEL) Assay
2.9. Quantitative Real-Time PCR (qRT-PCR)
2.10. Oxidative Stress and Inflammatory Marker Assays
2.11. Statistical Analysis
3. Results
3.1. Effects of Treatments on the General Health of Mice with CTX-Induced POF
3.2. Effects of Treatments on Restoration of Estrous Cycle Regularity in CTX-Induced POF Model Mice
3.3. Effects of Treatments on Ovary Dysfunction in POF Mice
3.4. Effects of Treatments on Granulosa-Cell Apoptosis in POF Mice
3.5. Effects of Treatments on Serum Hormone Levels in POF Mice
3.6. Effects of the Treatments on Ovarian Oxidative Stress and Inflammatory Markers in POF Mice
3.7. Effects of Treatments on the Expression of Ovarian Function-Related Genes in POF Mice
3.8. Effects of Treatments on the Expression of Hypothalamic Reproductive Regulatory Genes in POF Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mikhael, S.; Punjala-Patel, A.; Gavrilova-Jordan, L. Hypothalamic-Pituitary-Ovarian Axis Disorders Impacting Female Fertility. Biomedicines 2019, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Eshre Guideline Group on POI; Webber, L.; Davies, M.; Anderson, R.; Bartlett, J.; Braat, D.; Cartwright, B.; Cifkova, R.; de Muinck Keizer-Schrama, S.; Hogervorst, E. ESHRE Guideline: Management of Women with Premature Ovarian Insufficiency. Hum. Reprod. 2016, 31, 926–937. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhao, Q.; Li, Y.; Zheng, Z.; Kong, X.; Shu, C.; Liu, Y.; Shi, Y. The Role of Oxidative Stress in Ovarian Aging: A Review. J. Ovarian Res. 2022, 15, 100. [Google Scholar] [CrossRef]
- Ataya, K.; Moghissi, K. Chemotherapy-Induced Premature Ovarian Failure: Mechanisms and Prevention. Steroids 1989, 54, 607–626. [Google Scholar] [CrossRef]
- Pouladvand, N.; Azarnia, M.; Zeinali, H.; Fathi, R.; Tavana, S. An Overview of Different Methods to Establish a Murine Premature Ovarian Failure Model. Anim. Model. Exp. Med. 2024, 7, 835–852. [Google Scholar] [CrossRef]
- Kawano, M.; McKey, J.; Batchvarov, I.S.; Capel, B. Granulosa Cell Death Is a Significant Contributor to DNA-Damaging Chemotherapy-Induced Ovarian Insufficiency. Biol. Reprod. 2025, 112, 906–915. [Google Scholar] [CrossRef]
- Drummond, A.E. The Role of Steroids in Follicular Growth. Reprod. Biol. Endocrinol. 2006, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, J.; Xin, X.; Jin, Y.; Zhu, Y.; Zhang, H.; Fan, R.; Ye, Y.; Li, D. Efficacy of Natural Products on Premature Ovarian Failure: A Systematic Review and Meta-Analysis of Preclinical Studies. J. Ovarian Res. 2024, 17, 46. [Google Scholar] [CrossRef]
- Ma, L.; Li, X.; Li, C.; Chen, P.; Lan, Y.; Huang, Y.; Xu, W.; Zhou, J. Association of Coenzyme Q10 with Premature Ovarian Insufficiency. Reprod. Sci. 2023, 30, 1548–1554. [Google Scholar] [CrossRef]
- Barbarska, O.; Zaryczny-Trojan, L.; Minkiewicz-Zochniak, A. Folate Supplementation Awareness Among Women of Reproductive Age in Poland: Focus on Active Forms and Updated National Recommendations. Nutrients 2025, 17, 3881. [Google Scholar] [CrossRef]
- Sirotkin, A. V Effects, Mechanisms of Action and Application of Vitex agnus-castus for Improvement of Health and Female Reproduction. Phyther. Res. 2025, 39, 1484–1493. [Google Scholar] [CrossRef]
- Sopjani, M.; Murati, V.; Mataj-Berisha, D.; Xuan, N.T.; Faggio, C.; Rifati-Nixha, A. Vitex agnus-castus in Menopause: Phytochemistry, Mechanistic Insights, Clinical Applications, and Safety Perspectives. Phytother. Res. 2026, 40, 1919–1934. [Google Scholar] [CrossRef]
- Csupor, D.; Lantos, T.; Hegyi, P.; Benkő, R.; Viola, R.; Gyöngyi, Z.; Csecsei, P.; Toth, B.; Vasas, A.; Marta, K. Vitex agnus-castus in Premenstrual Syndrome: A Meta-Analysis of Double-Blind Randomised Controlled Trials. Complement. Ther. Med. 2019, 47, 102190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cui, S.; Mao, B.; Zhang, Q.; Zhao, J.; Zhang, H.; Tang, X.; Chen, W. Ellagic Acid and Intestinal Microflora Metabolite Urolithin A: A Review on Its Sources, Metabolic Distribution, Health Benefits, and Biotransformation. Crit. Rev. Food Sci. Nutr. 2023, 63, 6900–6922. [Google Scholar] [CrossRef]
- Pidgeon, R.; Mitchell, S.; Shamash, M.; Suleiman, L.; Dridi, L.; Maurice, C.F.; Castagner, B. Diet-Derived Urolithin A Is Produced by a Dehydroxylase Encoded by Human Gut Enterocloster Species. Nat. Commun. 2025, 16, 999. [Google Scholar] [CrossRef]
- Andreux, P.A.; Blanco-Bose, W.; Ryu, D.; Burdet, F.; Ibberson, M.; Aebischer, P.; Auwerx, J.; Singh, A.; Rinsch, C. The Mitophagy Activator Urolithin A Is Safe and Induces a Molecular Signature of Improved Mitochondrial and Cellular Health in Humans. Nat. Metab. 2019, 1, 595–603. [Google Scholar] [CrossRef]
- Hirshfield, A.N.; Rees Midgley, A., Jr. Morphometric Analysis of Foilicular Development in the Rat. Biol. Reprod. 1978, 19, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Spears, N.; Lopes, F.; Stefansdottir, A.; Rossi, V.; De Felici, M.; Anderson, R.A.; Klinger, F.G. Ovarian Damage from Chemotherapy and Current Approaches to Its Protection. Hum. Reprod. Update 2019, 25, 673–693. [Google Scholar] [CrossRef]
- Kim, J.; You, S. Extended Adverse Effects of Cyclophosphamide on Mouse Ovarian Function. BMC Pharmacol. Toxicol. 2021, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Ataya, K.; Ramahi-Ataya, A. Reproductive Performance of Female Rats Treated with Cyclophosphamide and/or LHRH Agonist. Reprod. Toxicol. 1993, 7, 229–235. [Google Scholar] [CrossRef]
- McNatty, K.P.; Makris, A.; Degrazia, C.; Rapin, O.; Ryan, K.J. The Production of Progesterone, Androgens, and Estrogens by Granulosa Cells, Thecal Tissue, and Stromal Tissue from Human Ovaries in Vitro. J. Clin. Endocrinol. Metab. 1979, 49, 687–699. [Google Scholar] [CrossRef]
- Song, Y.; Guo, Z.; Song, L.; Ma, J.; Ma, Y.; Shang, L.; Meng, Y.; Fan, Z.; Hao, M.; Zhao, J. Role of DNA Damage Response in Cyclophosphamide-induced Premature Ovarian Failure in Mice. J. Obstet. Gynaecol. Res. 2024, 50, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, M.; Wu, C.; Zhu, Q.; Wu, T.; Zhu, X.; Wu, M.; Wang, S. Effect of Hysterectomy on Ovarian Function: A Systematic Review and Meta-Analysis. J. Ovarian Res. 2023, 16, 35. [Google Scholar] [CrossRef]
- Cedars, M.I. Evaluation of Female Fertility—AMH and Ovarian Reserve Testing. J. Clin. Endocrinol. Metab. 2022, 107, 1510–1519. [Google Scholar] [CrossRef]
- Awonuga, A.O.; Camp, O.G.; Abu-Soud, H.M. A Review of Nitric Oxide and Oxidative Stress in Typical Ovulatory Women and in the Pathogenesis of Ovulatory Dysfunction in PCOS. Reprod. Biol. Endocrinol. 2023, 21, 111. [Google Scholar] [CrossRef]
- Xue, L.; Li, X.; Zhu, X.; Zhang, J.; Zhou, S.; Tang, W.; Chen, D.; Chen, Y.; Dai, J.; Wu, M. Carbon Tetrachloride Exposure Induces Ovarian Damage through Oxidative Stress and Inflammatory Mediated Ovarian Fibrosis. Ecotoxicol. Environ. Saf. 2022, 242, 113859. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Pan, C.; Zhang, C. Unraveling the Complexity of Follicular Fluid: Insights into Its Composition, Function, and Clinical Implications. J. Ovarian Res. 2024, 17, 237. [Google Scholar] [CrossRef] [PubMed]
- Bu, Y.; Yan, J.; Zhang, Z.; Xue, S.; Chi, F.; Zheng, Y.; Xia, Y.; Chen, Y. Acupuncture and the HPO Axis: A Review of Neuroendocrine Mechanisms with Implications for Ovarian Function. J. Integr. Neurosci. 2025, 24, 39451. [Google Scholar] [CrossRef]
- Dai, F.; Wang, R.; Deng, Z.; Yang, D.; Wang, L.; Wu, M.; Hu, W.; Cheng, Y. Comparison of the Different Animal Modeling and Therapy Methods of Premature Ovarian Failure in Animal Model. Stem Cell Res. Ther. 2023, 14, 135. [Google Scholar] [CrossRef]
- Reynolds, A.C.; McKenzie, L.J. Cancer Treatment-Related Ovarian Dysfunction in Women of Childbearing Potential: Management and Fertility Preservation Options. J. Clin. Oncol. 2023, 41, 2281–2292. [Google Scholar] [CrossRef]
- Barberino, R.S.; Silva, R.L.S.; Palheta Junior, R.C.; Smitz, J.E.J.; Matos, M.H.T. Protective Effects of Antioxidants on Cyclophosphamide-Induced Ovarian Toxicity. Biopreserv. Biobank. 2023, 21, 121–141. [Google Scholar] [CrossRef]
- Olowe, T.G.; Oyovwi, M.O.; Nwangwa, K.E.; Ohwin, E.P.; Oghenetega, O.B. Cytotoxic Properties of Cyclophosphamide: A Focus on Its Mechanistic Impacts on Male Gonadal Functions. J. Explor. Res. Pharmacol. 2024, 9, 106–115. [Google Scholar] [CrossRef]
- Chen, H.; Nie, P.; Li, J.; Wu, Y.; Yao, B.; Yang, Y.; Lash, G.E.; Li, P. Cyclophosphamide Induces Ovarian Granulosa Cell Ferroptosis via a Mechanism Associated with HO-1 and ROS-Mediated Mitochondrial Dysfunction. J. Ovarian Res. 2024, 17, 107. [Google Scholar] [CrossRef]
- Guo, Y.; Xue, L.; Tang, W.; Xiong, J.; Chen, D.; Dai, Y.; Wu, C.; Wei, S.; Dai, J.; Wu, M. Ovarian Microenvironment: Challenges and Opportunities in Protecting against Chemotherapy-Associated Ovarian Damage. Hum. Reprod. Update 2024, 30, 614–647. [Google Scholar] [CrossRef]
- Bakırhan, E.G.; Kaplan, S. The Protective Role of Vitamin E against the Effects of Hyperthyroidism on the Rat Pituitary–Ovary Axis. Histochem. Cell Biol. 2025, 163, 72. [Google Scholar] [CrossRef] [PubMed]
- Oyola, M.G.; Handa, R.J. Hypothalamic–Pituitary–Adrenal and Hypothalamic–Pituitary–Gonadal Axes: Sex Differences in Regulation of Stress Responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef] [PubMed]
- Broekmans, F.J.M.; De Ziegler, D.; Howles, C.M.; Gougeon, A.; Trew, G.; Olivennes, F. The Antral Follicle Count: Practical Recommendations for Better Standardization. Fertil. Steril. 2010, 94, 1044–1051. [Google Scholar] [CrossRef]
- GOODMAN, R.L.; PICKOVER, S.M.; KARSCH, F.J. Ovarian Feedback Control of Follicle-Stimulating Hormone in the Ewe: Evidence for Selective Suppression. Endocrinology 1981, 108, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Homburg, R. Ovulation Induction and Controlled Ovarian Stimulation: A Practical Guide, 2nd ed.; Springer: Cham, Switzerland, 2014; ISBN 978-3-319-05612-8. [Google Scholar]
- Liu, T.; Huang, Y.; Lin, H. Estrogen Disorders: Interpreting the Abnormal Regulation of Aromatase in Granulosa Cells. Int. J. Mol. Med. 2021, 47, 73. [Google Scholar] [CrossRef]
- Chauvin, S.; Cohen-Tannoudji, J.; Guigon, C.J. Estradiol Signaling at the Heart of Folliculogenesis: Its Potential Deregulation in Human Ovarian Pathologies. Int. J. Mol. Sci. 2022, 23, 512. [Google Scholar] [CrossRef]
- Park, C.J.; Oh, J.-E.; Lin, P.; Zhou, S.; Bunnell, M.; Bikorimana, E.; Spinella, M.J.; Lim, H.J.; Ko, C.J. A Dynamic Shift in Estrogen Receptor Expression during Granulosa Cell Differentiation in the Ovary. Endocrinology 2025, 166, bqaf006. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Akey, C.W. Apoptosome Structure, Assembly, and Procaspase Activation. Structure 2013, 21, 501–515. [Google Scholar] [CrossRef]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular Growth and Atresia in Mammalian Ovaries: Regulation by Survival and Death of Granulosa Cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef]
- Yang, S.; Yuan, S.; Zhang, X.; Wang, S.; Wen, J.; Wu, M.; Shen, L. The Systemic Effects of Ovarian Aging. In Ovarian Aging; Wang, S., Ed.; Springer: Singapore, 2023; pp. 35–65. ISBN 978-981-19-8848-6. [Google Scholar]
- Berga, S.; Naftolin, F. Neuroendocrine Control of Ovulation. Gynecol. Endocrinol. 2012, 28, 9–13. [Google Scholar] [CrossRef]
- Xu, Y.; Nisenblat, V.; Lu, C.; Li, R.; Qiao, J.; Zhen, X.; Wang, S. Pretreatment with Coenzyme Q10 Improves Ovarian Response and Embryo Quality in Low-Prognosis Young Women with Decreased Ovarian Reserve: A Randomized Controlled Trial. Reprod. Biol. Endocrinol. 2018, 16, 29. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, R.; Ruan, Y.; Fan, S. Urolithin A Protects Ovarian Reserve Via Inhibiting PI3K/Akt Signaling and Preventing Chemotherapy-Induced Follicle Apoptosis. Biology 2025, 14, 829. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Dai, Y.; Zhang, S.; Yang, L.; Liu, P.; Zhang, T.; Li, H.; Pang, Y.; Ma, S.; Zhang, Y.; Zhao, T. Urolithin A-Enhanced Multi-Bioactive Formulation Mitigates Cyclophosphamide-Induced Premature Ovarian Failure Through Suppression of Oxidative-Inflammatory Stress and Preservation of Follicle Fate. Antioxidants 2026, 15, 662. https://doi.org/10.3390/antiox15060662
Dai Y, Zhang S, Yang L, Liu P, Zhang T, Li H, Pang Y, Ma S, Zhang Y, Zhao T. Urolithin A-Enhanced Multi-Bioactive Formulation Mitigates Cyclophosphamide-Induced Premature Ovarian Failure Through Suppression of Oxidative-Inflammatory Stress and Preservation of Follicle Fate. Antioxidants. 2026; 15(6):662. https://doi.org/10.3390/antiox15060662
Chicago/Turabian StyleDai, Yangyan, Silu Zhang, Lijia Yang, Penglong Liu, Tingfeng Zhang, Hailong Li, Yuchen Pang, Shijing Ma, Yehui Zhang, and Tiantian Zhao. 2026. "Urolithin A-Enhanced Multi-Bioactive Formulation Mitigates Cyclophosphamide-Induced Premature Ovarian Failure Through Suppression of Oxidative-Inflammatory Stress and Preservation of Follicle Fate" Antioxidants 15, no. 6: 662. https://doi.org/10.3390/antiox15060662
APA StyleDai, Y., Zhang, S., Yang, L., Liu, P., Zhang, T., Li, H., Pang, Y., Ma, S., Zhang, Y., & Zhao, T. (2026). Urolithin A-Enhanced Multi-Bioactive Formulation Mitigates Cyclophosphamide-Induced Premature Ovarian Failure Through Suppression of Oxidative-Inflammatory Stress and Preservation of Follicle Fate. Antioxidants, 15(6), 662. https://doi.org/10.3390/antiox15060662