
CyberGenomics: Application of Behavioral Genetics in Cybersecurity

 ,
,  , , , and
, , , and
Abstract
:
1. Introduction
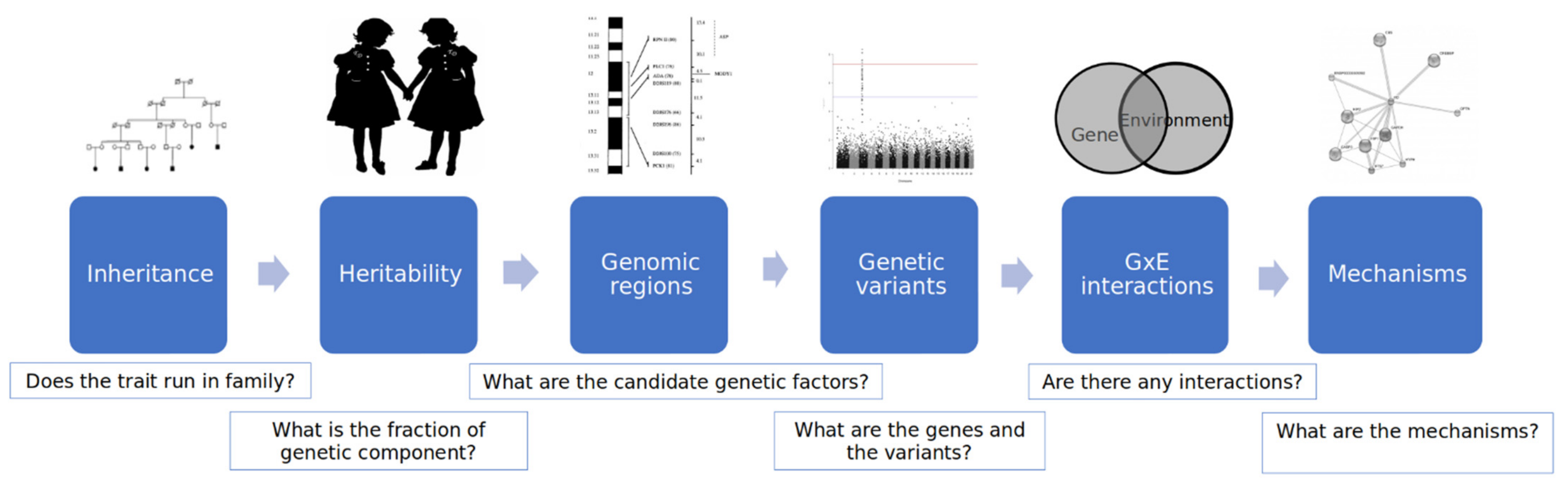
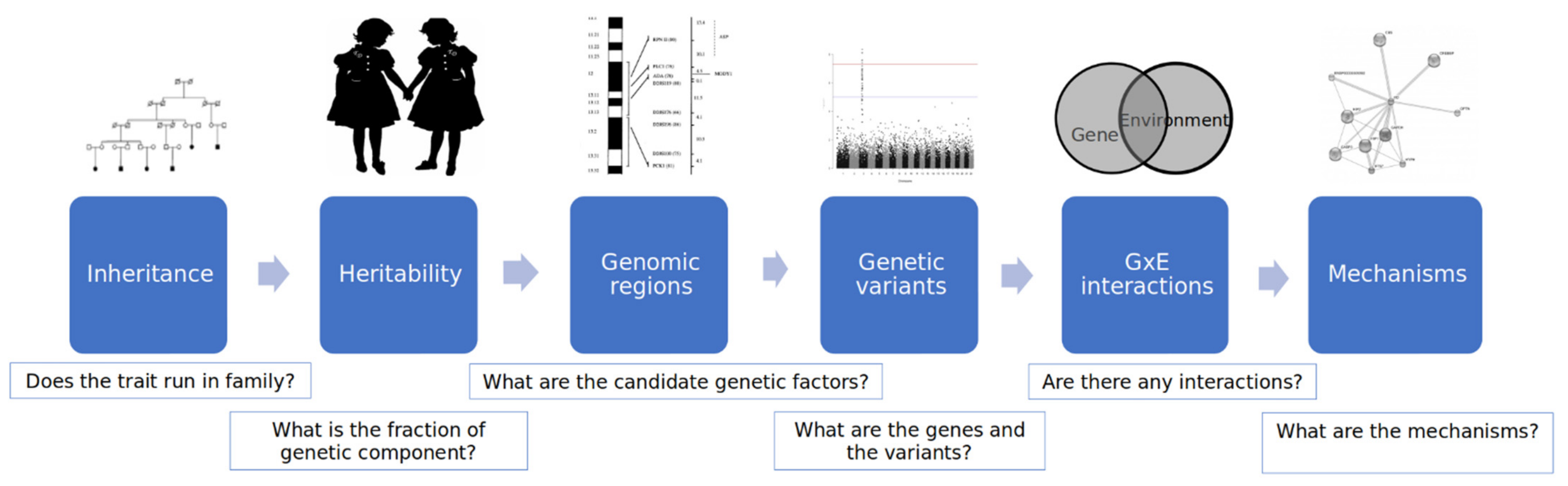
2. Genomic Factors
2.1. Determination of Behavior
2.2. Ethics
2.3. Human behind the Scene in Cybersecurity
3. Psychological Factors
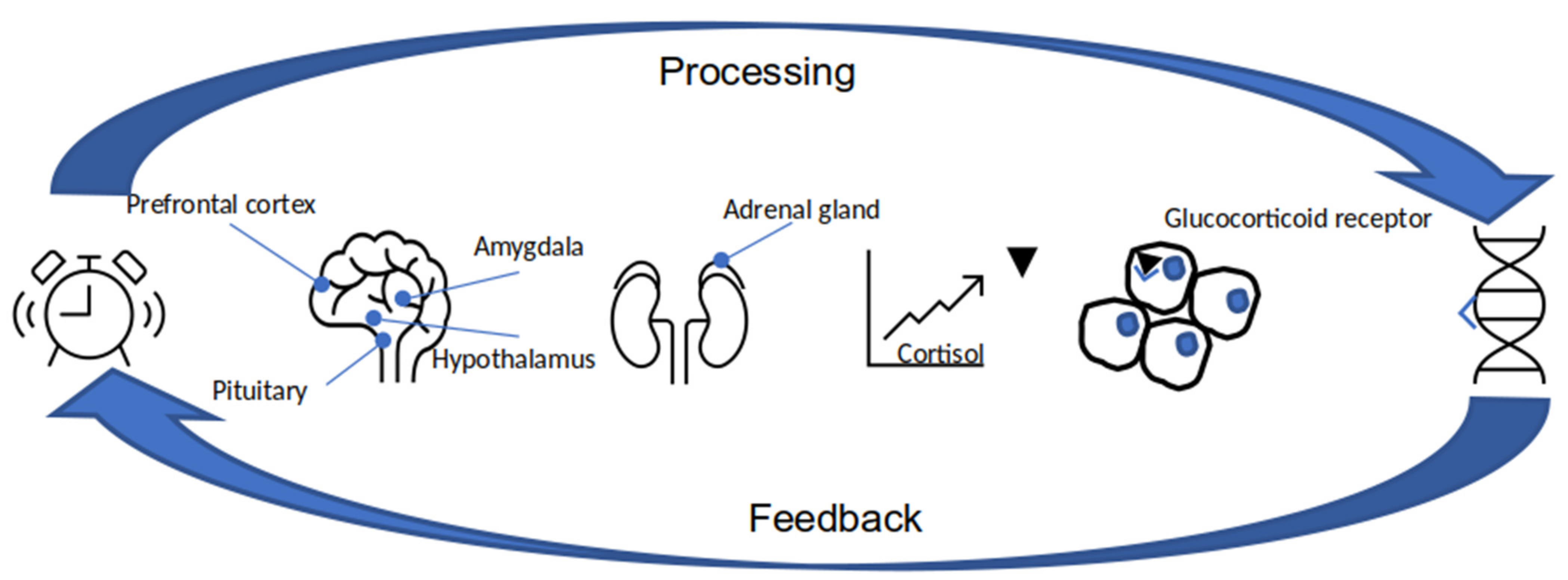
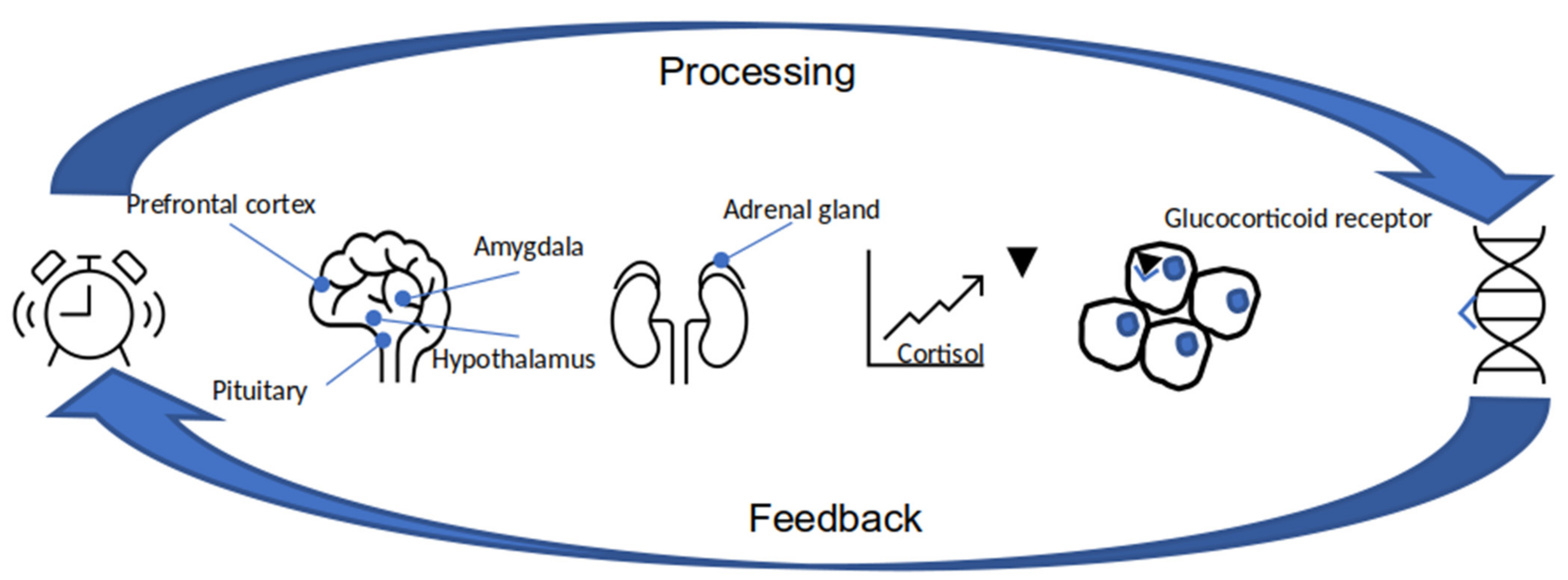
3.1. Stress
3.1.1. Stress Factor Defined by Genome
3.1.2. Stress in Cybersecurity Professional Career
- communication with collaborators that are busy with an incident response (their primary focus is not reporting);
- systemic thinking (IoCs must be correct and context-related);
- self-control (the report must be submitted in time).
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Human Genome Project Information Archive 1990–2003. Available online: http://web.ornl.gov/sci/techresources/Human_Genome (accessed on 28 July 2021).
- Assary, E.; Zavos, H.M.S.; Krapohl, E.; Keers, R.; Pluess, M. Genetic architecture of Environmental Sensitivity reflects multiple heritable components: A twin study with adolescents. Mol. Psychiatry 2020, 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R. Brain somatic mutations: The dark matter of psychiatric genetics? Mol. Psychiatry 2013, 19, 156–158. [Google Scholar] [CrossRef]
- Division of Behavioral and Social Sciences and Education. Social and Behavioral Sciences for National Security; National Academies Press: Washington, DC, USA, 2017. [Google Scholar]
- Balding, D.J.; Moltke, I.; Marioni, J. Handbook of Statistical Genomics; John Wiley & Sons: Hoboken, NJ, USA, 2019. [Google Scholar]
- Smoller, J.W. The Genetics of Stress-Related Disorders: PTSD, Depression, and Anxiety Disorders. Neuropsychopharmacology 2016, 41, 297–319. [Google Scholar] [CrossRef]
- Visscher, P.M.; Hill, W.G.; Wray, N.R. Heritability in the genomics era—Concepts and misconceptions. Nat. Rev. Genet. 2008, 9, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Felson, J. What can we learn from twin studies? A comprehensive evaluation of the equal environments assumption. Soc. Sci. Res. 2014, 43, 184–199. [Google Scholar] [CrossRef]
- Jelenkovic, A.; Sund, R.; Hur, Y.-M.; Yokoyama, Y.; Hjelmborg, J.; Möller, S.; Honda, C.; Magnusson, P.; Pedersen, N.; Ooki, S.; et al. Genetic and environmental influences on height from infancy to early adulthood: An individual-based pooled analysis of 45 twin cohorts. Sci. Rep. 2016, 6, 28496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lello, L.; Raben, T.G.; Hsu, S.D.H. Sibling validation of polygenic risk scores and complex trait prediction. Sci. Rep. 2020, 10, 13190. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.F. Spurious Genetic Associations. Biol. Psychiatry 2007, 61, 1121–1126. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, J.; Strandberg, E.; Arvelius, P.; Sánchez-Molano, E.; Pong-Wong, R.; Hickey, J.M.; Haskell, M.J.; Wiener, P. Genetic dissection of complex behaviour traits in German Shepherd dogs. Heredity 2019, 123, 746–758. [Google Scholar] [CrossRef]
- Alvarez, C.E. Naturally Occurring Cancers in Dogs: Insights for Translational Genetics and Medicine. ILAR J. 2014, 55, 16–45. [Google Scholar] [CrossRef] [Green Version]
- Zapata, I.; Serpell, J.A.; Alvarez, C.E. Genetic mapping of canine fear and aggression. BMC Genom. 2016, 17, 572. [Google Scholar] [CrossRef] [Green Version]
- Tang, R.; Noh, H.J.; Wang, D.; Sigurdsson, S.; Swofford, R.; Perloski, M.; Duxbury, M.; Patterson, E.E.; Albright, J.; Castelhano, M.; et al. Candidate genes and functional noncoding variants identified in a canine model of obsessive-compulsive disorder. Genome Biol. 2014, 15, R25. [Google Scholar] [CrossRef] [Green Version]
- Ha, J.H.; Alam, M.; Lee, D.H.; Kim, J.-J. Whole Genome Association Study to Detect Single Nucleotide Polymorphisms for Behavior in Sapsaree Dog (Canis familiaris). Asian-Aust. J. Anim. Sci. 2015, 28, 936–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erbe, M.; Hayes, B.; Matukumalli, L.; Goswami, S.; Bowman, P.; Reich, C.; Mason, B.; Goddard, M. Improving accuracy of genomic predictions within and between dairy cattle breeds with imputed high-density single nucleotide polymorphism panels. J. Dairy Sci. 2012, 95, 4114–4129. [Google Scholar] [CrossRef] [Green Version]
- Kemper, E.K.; Reich, C.M.; Bowman, P.J.; Jagt, C.J.V.; Chamberlain, A.J.; Mason, A.B.; Hayes, B.J.; Goddard, M. Improved precision of QTL mapping using a nonlinear Bayesian method in a multi-breed population leads to greater accuracy of across-breed genomic predictions. Genet. Sel. Evol. 2015, 47, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLeod, I.M.; Bowman, P.J.; Jagt, C.J.V.; Haile-Mariam, M.; Kemper, K.E.; Chamberlain, A.J.; Schrooten, C.; Hayes, B.J.; Goddard, M.E. Exploiting biological priors and sequence variants enhances QTL discovery and genomic prediction of complex traits. BMC Genom. 2016, 17, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, G.; Lee, S.H.; Hayes, B.; Goddard, M.; Wray, N.R.; Visscher, P. Simultaneous Discovery, Estimation and Prediction Analysis of Complex Traits Using a Bayesian Mixture Model. PLoS Genet. 2015, 11, e1004969. [Google Scholar] [CrossRef] [PubMed]
- Aliloo, H.; Pryce, J.E.; González-Recio, O.; Cocks, B.G.; Hayes, B.J. Accounting for dominance to improve genomic evaluations of dairy cows for fertility and milk production traits. Genet. Sel. Evol. 2016, 48, 8. [Google Scholar] [CrossRef] [Green Version]
- Clark, J. Medicalization of global health 1: Has the global health agenda become too medicalized? Glob. Health Action 2014, 7, 23998. [Google Scholar] [CrossRef] [Green Version]
- Hart, G. Sexual behaviour and its medicalisation: In sickness and in health. BMJ 2002, 324, 896–900. [Google Scholar] [CrossRef] [Green Version]
- Nuffield Council on Bioethics. Genetics and Human Behaviour: The Ethical Context–Summary and Recommendations. J. Int. Bioeth. Int. J. Bioeth. 2003, 14, 171–192. [Google Scholar] [CrossRef]
- Psychiatry, T.L. Medicalisation and its discontents. Lancet Psychiatry 2016, 3, 591. [Google Scholar] [CrossRef]
- Wyatt, W.J. Behavior Analysis in the Era of Medicalization: The State of the Science and Recommendations for Practitioners. Behav. Anal. Pr. 2009, 2, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Rothstein, A.M.; Anderlik, M.R. What is genetic discrimination, and when and how can it be prevented? Genet. Med. 2001, 3, 354–358. [Google Scholar] [CrossRef] [Green Version]
- Byrne, B.; Olson, R.K.; Samuelsson, S.; Wadsworth, S.; Corley, R.; DeFries, J.C.; Willcutt, E. Genetic and environmental influences on early literacy. J. Res. Read. 2006, 29, 33–49. [Google Scholar] [CrossRef]
- Petrill, S.A. Behavioural Genetic Studies of Reading and Mathematics Skills. Behav. Genet. Educ. 2016, 2016, 60–76. [Google Scholar] [CrossRef]
- Libertus, M.E.; Feigenson, L.; Halberda, J. Preschool acuity of the approximate number system correlates with school math ability. Dev. Sci. 2011, 14, 1292–1300. [Google Scholar] [CrossRef] [Green Version]
- Klitzman, R. Views of Discrimination among Individuals Confronting Genetic Disease. J. Genet. Couns. 2010, 19, 68–83. [Google Scholar] [CrossRef] [Green Version]
- Hatemi, P.; Alford, J.R.; Hibbing, J.R.; Martin, N.; Eaves, L.J. Is There a “Party” in Your Genes? Politi-Res. Q. 2009, 62, 584–600. [Google Scholar] [CrossRef] [Green Version]
- Berryessa, C.M.; Cho, M.K. Ethical, Legal, Social, and Policy Implications of Behavioral Genetics. Annu. Rev. Genom. Hum. Genet. 2013, 14, 515–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puscas, I.M. Military Enhancement: Technologies, Ethics and Operational Issues. In Ethics of Medical Innovation, Experimentation, and Enhancement in Military and Humanitarian Contexts; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2020; Volume 2020, pp. 127–146. [Google Scholar]
- Lemay, A.; Calvet, J.; Menet, F.; Fernandez, J.M. Survey of publicly available reports on advanced persistent threat actors. Comput. Secur. 2018, 72, 26–59. [Google Scholar] [CrossRef]
- Saalbach, K.-P. Attribution of Cyber Attacks. Inf. Technol. Peace Secur. 2019, 2019, 279–303. [Google Scholar] [CrossRef] [Green Version]
- MISP Project. MISP—Open Source Threat Intelligence Platform & Open Standards For Threat Information Sharing. 2019. Available online: https://www.misp-project.org (accessed on 31 October 2020).
- Hutchins, E.M.; Cloppert, M.J.; Amin, R.M. Intelligence-driven computer network defense informed by analysis of adversary campaigns and intrusion kill chains. Lead Issues Inf. Warf Secur. Res. 2011, 1, 80. [Google Scholar]
- Hoffmann, R. Stochastic Model of the Simple Cyber Kill Chain: Cyber Attack Process as a Regenerative Process. In Computer Information Systems and Industrial Management; Saeed, K., Dvorský, J., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 355–365. [Google Scholar]
- ISC22. How Views on Cybersecurity Professionals Are Changing and What Hiring Organizations Need to Know. The 2020 (ISC)2 Cybersecurity Perception Study; ISC: New Delhi, India, 2020. [Google Scholar]
- ACM. Joint Task Force on Cybersecurity Education. In Cybersecurity Curricula 2017: Curriculum Guidelines for Post-Secondary Degree Programs in Cybersecurity; ACM: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Newhouse, W.; Keith, S.; Scribner, B.; Witte, G. National Initiative for Cybersecurity Education (NICE) Cybersecurity Workforce Framework. NIST Spec. Publ. 2017, 800, 181. [Google Scholar] [CrossRef]
- Petersen, R.; Santos, D.; Smith, M.C.; Wetzel, K.A.; Witte, G. Workforce Framework for Cybersecurity (NICE Framework; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2020; Volume 181. [Google Scholar] [CrossRef]
- Parrish, A.; Impagliazzo, J.; Raj, R.K.; Santos, H.; Asghar, M.R.; Jøsang, A.; Pereira, T.; Stavrou, E. Global perspectives on cybersecurity education for 2030: A case for a meta-discipline. In Proceedings of the Companion of the 23rd Annual ACM Conference on Innovation and Technology in Computer Science Education, Larnaca, Cyprus, 2–4 July 2018; Volume 2018, pp. 36–54. [Google Scholar]
- Esparza, J.; Caporusso, N.; Walters, A. Addressing Human Factors in the Design of Cyber Hygiene Self-assessment Tools. In Advances in Human Factors in Cybersecurity; Corradini, I., Nardelli, E., Ahram, T., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 88–94. [Google Scholar]
- Alohali, M.; Clarke, N.; Li, F.; Furnell, S. Identifying and predicting the factors affecting end-users’ risk-taking behavior. Inf. Comput. Secur. 2018, 26, 306–326. [Google Scholar] [CrossRef]
- Corradini, I. Redefining the Approach to Cybersecurity; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2020; pp. 49–62. [Google Scholar]
- Zimmermann, V.; Renaud, K. Moving from a ‘human-as-problem” to a ‘human-as-solution” cybersecurity mindset. Int. J. Human-Comput. Stud. 2019, 131, 169–187. [Google Scholar] [CrossRef]
- Vukasović, T.; Bratko, D. Heritability of personality: A meta-analysis of behavior genetic studies. Psychol. Bull. 2015, 141, 769–785. [Google Scholar] [CrossRef] [PubMed]
- Briley, D.A.; Tucker-Drob, E.M. Comparing the Developmental Genetics of Cognition and Personality over the Life Span. J. Pers. 2017, 85, 51–64. [Google Scholar] [CrossRef]
- Rothbart, M.K.; Ahadi, S.A.; Evans, D.E. Temperament and personality: Origins and outcomes. J. Pers. Soc. Psychol. 2000, 78, 122–135. [Google Scholar] [CrossRef]
- Savitz, J.; Solms, M.; Ramesar, R. The molecular genetics of cognition: Dopamine, COMT and BDNF. Genes Brain Behav. 2006, 5, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Zmorzyński, S.; Styk, W.; Klinkosz, W.; Iskra, J.; Filip, A.A. Personality traits and polymorphisms of genes coding neurotransmitter receptors or transporters: Review of single gene and genome-wide association studies. Ann. Gen. Psychiatry 2021, 20, 7. [Google Scholar] [CrossRef] [PubMed]
- Freed, S.E. Examination of Personality Characteristics Among Cybersecurity and Information Technology Professionals; University of Tennessee: Chattanooga, TN, USA, 2014. [Google Scholar]
- Lugo, R.G.; Sütterlin, S. Cyber Officer Profiles and Performance Factors. In Lecture Notes in Computer Science; Springer: Dordrecht, The Netherlands, 2018; pp. 181–190. [Google Scholar]
- Ising, M.; Holsboer, F. Genetics of stress response and stress-related disorders. Dialog-Clin. Neurosci. 2006, 8, 433–444. [Google Scholar] [CrossRef]
- Pacák, K.; Palkovits, M. Stressor Specificity of Central Neuroendocrine Responses: Implications for Stress-Related Disorders. Endocr. Rev. 2001, 22, 502–548. [Google Scholar] [CrossRef] [PubMed]
- Albert, D.; Belsky, D.W.; Crowley, D.M.; Latendresse, S.J.; Aliev, F.; Riley, B.P.; Sun, C.; Dick, D.M.; Dodge, K.A. Conduct Problems Prevention Research Group Can Genetics Predict Response to Complex Behavioral Interventions? Evidence from a Genetic Analysis of the Fast Track Randomized Control Trial. J. Policy Anal. Manag. 2015, 34, 497–518. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Sharma, S. Physiology, Adrenocorticotropic Hormone (ACTH); StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Riese, H.; Rijsdijk, F.V.; Rosmalen, J.; Snieder, H.; Ormel, J. Neuroticism and Morning Cortisol Secretion: Both Heritable, But No Shared Genetic Influences. J. Pers. 2009, 77, 1561–1576. [Google Scholar] [CrossRef]
- CORtisolNETwork (CORNET) Consortium; Rietschel, L.; Streit, F.; Zhu, G.; McAloney, K.; Frank, J.; Couvy-Duchesne, B.; Witt, S.H.; Binz, T.; McGrath, J.; et al. Hair Cortisol in Twins: Heritability and Genetic Overlap with Psychological Variables and Stress-System Genes. Sci. Rep. 2017, 7, 15351. [Google Scholar] [CrossRef]
- Cornelis, M.C.; Nugent, N.; Amstadter, A.B.; Koenen, K.C. Genetics of Post-Traumatic Stress Disorder: Review and Recommendations for Genome-Wide Association Studies. Curr. Psychiatry Rep. 2010, 12, 313–326. [Google Scholar] [CrossRef] [Green Version]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. 2015, 11, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Goldman-Rakic, P.S.; Muly, E.C., III; Williams, G.V. D1 receptors in prefrontal cells and circuits. Brain Res. Rev. 2000, 31, 295–301. [Google Scholar] [CrossRef]
- Tartar, J.L.; Cabrera, D.; Knafo, S.; Thomas, J.D.; Antonio, J.; Peacock, C.A. The “Warrior” COMT Val/Met Genotype Occurs in Greater Frequencies in Mixed Martial Arts Fighters Relative to Controls. J Sports Sci Med 2020, 19, 38–42. [Google Scholar]
- Pflüger, L.S.; Gutleb, D.R.; Hofer, M.; Fieder, M.; Wallner, B.; Steinborn, R. Allelic variation of the COMT gene in a despotic primate society: A haplotype is related to cortisol excretion in Macaca fuscata. Horm. Behav. 2016, 78, 220–230. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Begni, V.; Pariante, C.M.; Riva, M.A. The human BDNF gene: Peripheral gene expression and protein levels as biomarkers for psychiatric disorders. Transl. Psychiatry 2016, 6, e958. [Google Scholar] [CrossRef] [PubMed]
- Lubin, F.D.; Roth, T.L.; Sweatt, J.D. Epigenetic Regulation of bdnf Gene Transcription in the Consolidation of Fear Memory. J. Neurosci. 2008, 28, 10576–10586. [Google Scholar] [CrossRef]
- Van Winkel, M.; Peeters, F.; van Winkel, R.; Kenis, G.; Collip, D.; Geschwind, N.; Jacobs, N.; Derom, C.; Thiery, E.; van Os, J.; et al. Impact of variation in the BDNF gene on social stress sensitivity and the buffering impact of positive emotions: Replication and extension of a gene–environment interaction. Eur. Neuropsychopharmacol. 2014, 24, 930–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, A.E.; Canli, T. Influence of life stress, 5-HTTLPR genotype, and SLC6A4 methylation on gene expression and stress response in healthy Caucasian males. Biol. Mood Anxiety Disord. 2015, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risch, N.; Herrell, R.; Lehner, T.; Liang, K.-Y.; Eaves, L.; Hoh, J.; Griem, A.; Kovacs, M.; Ott, J.; Merikangas, K.R. Interaction Between the Serotonin Transporter Gene (5-HTTLPR), Stressful Life Events, and Risk of Depression. JAMA 2009, 301, 2462–2471. [Google Scholar] [CrossRef] [Green Version]
- Qi, R.; Luo, Y.; Zhang, L.; Weng, Y.; Surento, W.; Li, L.; Cao, Z.; Lu, G.M. Effects of COMT rs4680 and BDNF rs6265 polymorphisms on brain degree centrality in Han Chinese adults who lost their only child. Transl. Psychiatry 2020, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Vogel, S.; Schwabe, L. Learning and memory under stress: Implications for the classroom. NPJ Sci. Learn. 2016, 1, 16011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Keulen, B.J.; Dolan, C.V.; Andrew, R.; Walker, B.R.; Pol, H.E.H.; Boomsma, D.I.; Rotteveel, J.; Finken, M.J.J. Heritability of Cortisol Production and Metabolism Throughout Adolescence. J. Clin. Endocrinol. Metab. 2019, 105, 443–452. [Google Scholar] [CrossRef]
- Van Keulen, B.J.; Dolan, C.V.; Andrew, R.; Walker, B.R.; Pol, H.E.H.; Boomsma, D.I.; Rotteveel, J.; Finken, M.J.J. Long-Term Stability of Cortisol Production and Metabolism Throughout Adolescence: Longitudinal Twin Study. Twin Res. Hum. Genet. 2020, 23, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Boonen, E.; Vervenne, H.; Meersseman, P.; Andrew, R.; Mortier, L.; Declercq, P.E.; Vanwijngaerden, Y.-M.; Spriet, I.; Wouters, P.J.; Perre, S.V.; et al. Reduced Cortisol Metabolism during Critical Illness. N. Engl. J. Med. 2013, 368, 1477–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allman, J.M.; Hakeem, A.; Erwin, J.M.; Nimchinsky, E.; Hof, P. The anterior cingulate cortex. The evolution of an interface between emotion and cognition. Ann. N. Y. Acad. Sci. 2001, 935, 107–117. [Google Scholar] [CrossRef]
- Fan, J.; Wu, Y.; Fossella, A.J.; Posner, I.M. Assessing the heritability of attentional networks. BMC Neurosci. 2001, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.P.; Bruce, J.; Gunnar, M.R. The anterior attention network: Associations with temperament and neuroendocrine activity in 6-year-old children. Dev. Psychobiol. 2001, 40, 43–56. [Google Scholar] [CrossRef]
- Liu, J.; Xu, X.; Luo, Q.; Luo, Y.; Chen, Y.; Lui, S.; Wu, M.; Zhu, H.; Kemp, G.; Gong, Q. Brain grey matter volume alterations associated with antidepressant response in major depressive disorder. Sci. Rep. 2017, 7, 10464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treadway, M.T.; Grant, M.M.; Ding, Z.; Hollon, S.D.; Gore, J.C.; Shelton, R.C. Early Adverse Events, HPA Activity and Rostral Anterior Cingulate Volume in MDD. PLoS ONE 2009, 4, e4887. [Google Scholar] [CrossRef]
- Thomason, M.E.; Hamilton, J.P.; Gotlib, I.H. Stress-induced activation of the HPA axis predicts connectivity between subgenual cingulate and salience network during rest in adolescents. J. Child. Psychol. Psychiatry 2011, 52, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Oltsik, J. The Life and Times of Cybersecurity Professionals. 2018. Available online: https://www.esg-global.com/hubfs/pdf/ESG-ISSA-Research-Report-Life-of-Cybersecurity-Professionals-Apr-2019.pdf (accessed on 31 October 2020).
- Exabeam. Exabeam 2019: Cybersecurity Professionals Salary, Skills, and Stress Survey. 2019. Available online: https://www.exabeam.com/library/2019-cybersecurity-professionals-salary-skills-and-stress-survey (accessed on 31 October 2020).
- Feher, A.; Vernon, P.A. Looking beyond the Big Five: A selective review of alternatives to the Big Five model of personality. Pers. Individ. Differ. 2021, 169, 110002. [Google Scholar] [CrossRef]
- Roberts, R.; Woodman, T. Personality and performance: Moving beyond the Big 5. Curr. Opin. Psychol. 2017, 16, 104–108. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Task | Description | Role |
|---|---|---|
| T0854 | Tip critical or time-sensitive information to appropriate customers | Multi-Disciplined Language Analyst |
| T0800 | Provide timely notice of imminent or hostile intentions or activities which may impact organization objectives, resources, or capabilities | Threat/Warning Analyst, All-Source Analyst |
| T0258 | Provide timely detection, identification, and alerting of possible attacks/intrusions, anomalous activities, and misuse activities and distinguish these incidents and events from benign activities | Cyber Defense Analyst |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domarkienė, I.; Ambrozaitytė, L.; Bukauskas, L.; Rančelis, T.; Sütterlin, S.; Knox, B.J.; Maennel, K.; Maennel, O.; Parish, K.; Lugo, R.G.; et al. CyberGenomics: Application of Behavioral Genetics in Cybersecurity. Behav. Sci. 2021, 11, 152. https://doi.org/10.3390/bs11110152
Domarkienė I, Ambrozaitytė L, Bukauskas L, Rančelis T, Sütterlin S, Knox BJ, Maennel K, Maennel O, Parish K, Lugo RG, et al. CyberGenomics: Application of Behavioral Genetics in Cybersecurity. Behavioral Sciences. 2021; 11(11):152. https://doi.org/10.3390/bs11110152
Chicago/Turabian StyleDomarkienė, Ingrida, Laima Ambrozaitytė, Linas Bukauskas, Tautvydas Rančelis, Stefan Sütterlin, Benjamin James Knox, Kaie Maennel, Olaf Maennel, Karen Parish, Ricardo Gregorio Lugo, and et al. 2021. "CyberGenomics: Application of Behavioral Genetics in Cybersecurity" Behavioral Sciences 11, no. 11: 152. https://doi.org/10.3390/bs11110152