
Effects of Sea-Buckthorn Flavonoids on Growth Performance, Serum Inflammation, Intestinal Barrier and Microbiota in LPS-Challenged Broilers

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection
2.3. Index Determination Method
2.3.1. Measurement of Growth Performance
2.3.2. Determination of Serum Indicators
2.3.3. Gut Histomorphological Analysis
2.3.4. Gene Expression Using Quantitative Real-Time PCR
2.3.5. 16S rRNA Sequencing
2.4. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Serum Inflammation
3.3. Intestinal Morphology
3.4. Intestinal Permeability and Expression of Jejunal Mucosa-Associated Gene mRNA
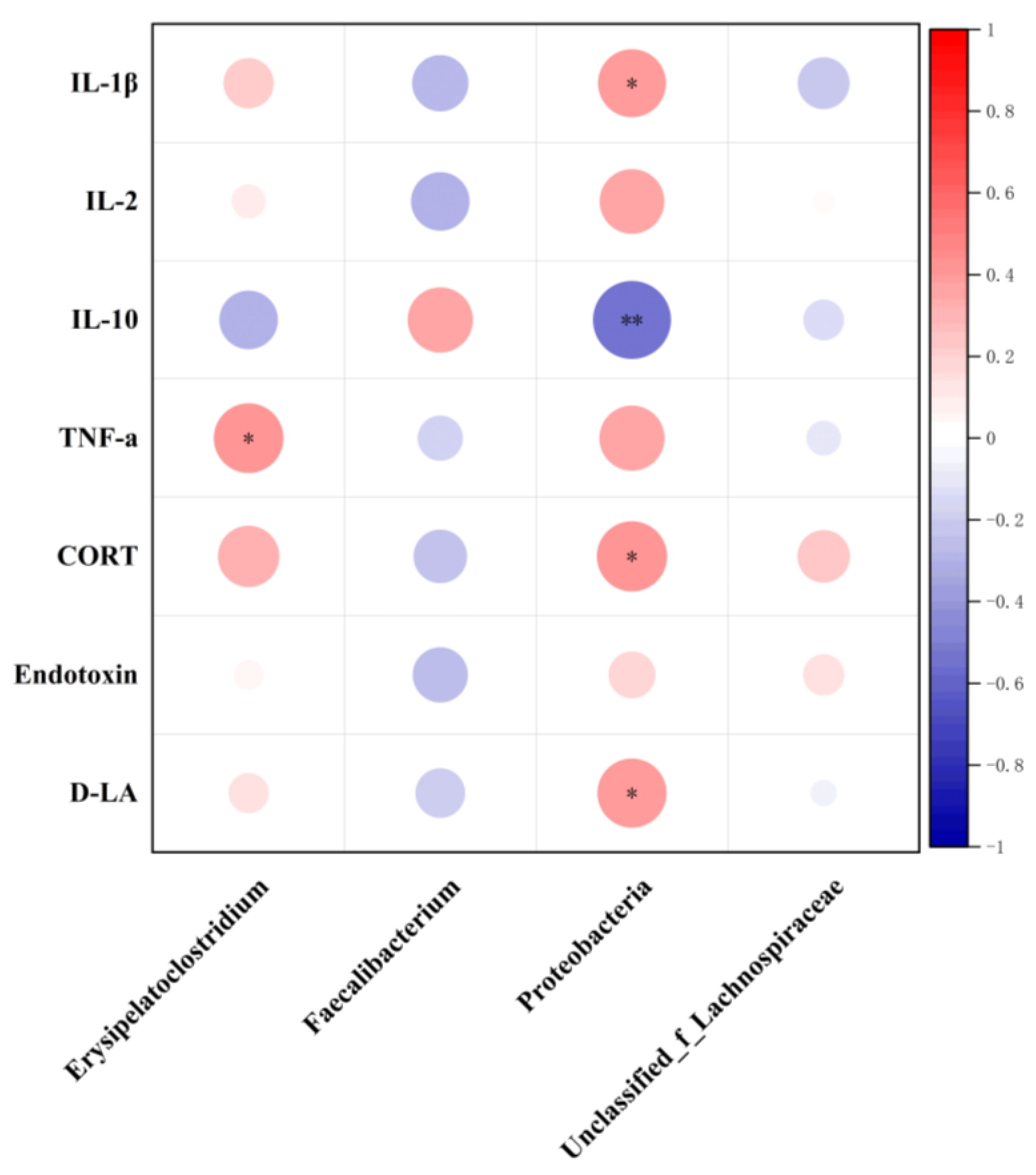
3.5. Gut Microbiota Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, L.A.; Er, M.W.; Li, L.H.; Wen, P.; Jia, Y.C.; Huo, L.M. Microclimate environment model construction and control strategy of enclosed laying brooder house. Poult Sci. 2022, 101, 101843. [Google Scholar] [CrossRef]
- Liu, Y.H.; Zhang, Y.; Bai, D.Y.; Li, Y.Q.; He, X.L.; Ito, K.; Liu, K.; Tan, H.; Zhen, W.; Zhang, B.; et al. Dietary Supplementation with Chlorogenic Acid Enhances Antioxidant Capacity, Which Promotes Growth, Jejunum Barrier Function, and Cecum Microbiota in Broilers under High Stocking Density Stress. Animals 2023, 13, 303. [Google Scholar] [CrossRef]
- Yao, M.; Gao, W.H.; Tao, H.X.; Yang, J.; Huang, T.H. The regulation effects of danofloxacin on pig immune stress induced by LPS. Res. Vet. Sci. 2017, 110, 65–71. [Google Scholar] [CrossRef]
- Wang, M.Y.; Zhang, Y.; Tong, Y.X.; Guo, P.T.; Zhang, J.; Wang, C.K.; Gao, Y.Y. Effects of lutein on jejunal mucosal barrier function and inflammatory responses in lipopolysaccharide-challenged yellow-feather broilers. Poult Sci. 2022, 101, 102191. [Google Scholar] [CrossRef]
- Dias, K.M.M.; Oliveira, C.H.; Calderano, A.A.; Rostagno, H.S.; Gomes, K.M.; O’Connor, K.E.; Davis, R.; Walsh, M.; Britton, J.; Altieri, E.A.; et al. Dietary Hydroxytyrosol Supplementation on Growth Performance, Gut Morphometry, and Oxidative and Inflammatory Status in LPS-Challenged Broilers. Animals 2024, 14, 871. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Yu, Y.; Zhu, Z.; Tian, H.; Guo, Y.; Zan, R.; Tian, Y.; Jiang, R.; Li, K.; Sun, G.; et al. Regulation of the MyD88 gene in chicken spleen inflammation induced by stress. J. Anim. Sci. 2023, 101, skad060. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, J.J.; Zhu, P.J.; Han, H.M.; Wan, X.L.; Yang, H.M.; Wang, Z.Y. Effects of betaine on growth performance, intestinal health, and immune response of goslings challenged with lipopolysaccharide. Poult Sci. 2022, 101, 102153. [Google Scholar] [CrossRef]
- Ciesarová, Z.; Murkovic, M.; Cejpek, K.; Kreps, F.; Tobolková, B.; Koplík, R.; Belajová, E.; Kukurová, K.; Daško, Ľ.; Panovská, Z.; et al. Why is sea buckthorn (Hippophae rhamnoides L.) so exceptional? A review. Food Res Int. 2020, 133, 109170. [Google Scholar] [CrossRef]
- Sireswar, S.; Biswas, S.; Dey, G. Adhesion and anti-inflammatory potential of Lactobacillus rhamnosus GG in a sea buckthorn based beverage matrix. Food Funct. 2020, 11, 2555–2572. [Google Scholar] [CrossRef]
- Sławińska, N.; Żuchowski, J.; Stochmal, A.; Olas, B. Extract from Sea Buckthorn Seeds-A Phytochemical, Antioxidant, and Hemostasis Study; Effect of Thermal Processing on Its Chemical Content and Biological Activity In Vitro. Nutrients 2023, 15, 686. [Google Scholar] [CrossRef]
- Jayashankar, B.; Singh, D.; Tanwar, H.; Mishra, K.P.; Murthy, S.; Chanda, S.; Mishra, J.; Tulswani, R.; Misra, K.; Singh, S.B.; et al. Augmentation of humoral and cellular immunity in response to Tetanus and Diphtheria toxoids by supercritical carbon dioxide extracts of Hippophae rhamnoides L. leaves. Int. Immunopharmacol. 2017, 44, 123–136. [Google Scholar] [CrossRef]
- Xiao, P.T.; Liu, S.Y.; Kuang, Y.J.; Jiang, Z.M.; Lin, Y.; Xie, Z.S.; Liu, E.H. Network pharmacology analysis and experimental validation to explore the mechanism of sea buckthorn flavonoids on hyperlipidemia. J Ethnopharmacol. 2021, 264, 113380. [Google Scholar] [CrossRef]
- Sharma, A.; Shukla, P.K.; Bhattacharyya, A.; Kumar, U.; Roy, D.; Yadav, B.; Prakash, A. Effect of dietary supplementation of sea buckthorn and giloe leaf meal on the body weight gain, feed conversion ratio, biochemical attributes, and meat composition of turkey poults. Vet. World 2018, 11, 93–98. [Google Scholar] [CrossRef]
- Hao, X.Y.; Zhang, X.Z.; Yang, D.Y.; Xie, Y.Z.; Mu, C.T.; Zhang, J.X. Effects of sea-buckthorn flavonoids on growth performance, nutrient digestibility, microbial protein synthesis, and plasma antioxidant capacity of finishing lambs. Anim. Feed. Sci. Technol. 2023, 305, 115783. [Google Scholar] [CrossRef]
- Ren, Q.C.; Li, X.H.; Li, Q.Y.; Yang, H.L.; Wang, H.L.; Zhang, H.; Zhao, L.; Jiang-Yong, S.L.; Meng, X.L.; Zhang, Y.; et al. Total flavonoids from sea buckthorn ameliorates lipopolysaccharide/cigarette smoke-induced airway inflammation. Phytother Res. 2019, 33, 2102–2117. [Google Scholar] [CrossRef]
- Xu, Y.; Li, J.; Lin, Z.; Liang, W.; Qin, L.; Ding, J.; Chen, S.; Zhou, L. Isorhamnetin Alleviates Airway Inflammation by Regulating the Nrf2/Keap1 Pathway in a Mouse Model of COPD. Front Pharmacol. 2022, 13, 860362. [Google Scholar] [CrossRef]
- Sul, O.J.; Ra, S.W. Quercetin Prevents LPS-Induced Oxidative Stress and Inflammation by Modulating NOX2/ROS/NF-kB in Lung Epithelial Cells. Molecules 2021, 26, 6949. [Google Scholar] [CrossRef]
- Bian, Y.; Lei, J.; Zhong, J.; Wang, B.; Wan, Y.; Li, J.; Liao, C.; He, Y.; Liu, Z.; Ito, K.; et al. Kaempferol reduces obesity, prevents intestinal inflammation, and modulates gut microbiota in high-fat diet mice. J. Nutr. Biochem. 2022, 99, 108840. [Google Scholar] [CrossRef]
- Ma, J.S.; Chang, W.H.; Liu, G.H.; Zhang, S.; Zheng, A.J.; Li, Y.; Xie, Q.; Liu, Z.Y.; Cai, H.Y. Effects of flavones of sea buckthorn fruits on growth performance, carcass quality, fat deposition and lipometabolism for broilers. Poult Sci. 2015, 94, 2641–2649. [Google Scholar] [CrossRef]
- An, J.S.; Shi, J.J.; Liu, K.B.; Li, A.K.; He, B.B.; Wang, Y.; He, J.H. Effects of Solid-State Fermented Wheat Bran on Growth Performance, Immune Function, Intestinal Morphology and Microflora in Lipopolysaccharide-Challenged Broiler Chickens. Animals 2022, 12, 1100. [Google Scholar] [CrossRef]
- Zhang, P.F.; Shi, B.L.; Su, J.L.; Yue, Y.X.; Cao, Z.X.; Chu, W.B.; Li, K.; Yan, S.M. Relieving effect of Artemisia argyi aqueous extract on immune stress in broilers. J. Anim. Physiol. Anim. Nutr. 2017, 101, 251–258. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, S.; Wang, F.; Wang, T.; Yin, L.; Chen, J.; Fu, C. Effects of Dietary Terminalia chebula Extract on Growth Performance, Immune Function, Antioxidant Capacity, and Intestinal Health of Broilers. Animals 2024, 14, 746. [Google Scholar] [CrossRef]
- Chang, W.Y.; Yu, Y.H. Effect of Bacillus species-fermented products and essential oils on growth performance, gut morphology, cecal short-chain fatty acid levels, and microbiota community in broilers. Poult. Sci. 2022, 101, 101970. [Google Scholar] [CrossRef]
- Xing, Y.Y.; Zheng, Y.K.; Yang, S.; Zhang, L.H.; Guo, S.W.; Shi, L.L.; Xu, Y.Q.; Jin, X.; Yan, S.M.; Shi, B.L. Artemisia ordosica Polysaccharide Alleviated Lipopolysaccharide-induced Oxidative Stress of Broilers via Nrf2/Keap1 and TLR4/NF-κB Pathway. Ecotoxicol. Environ. Saf. 2021, 223, 112566. [Google Scholar] [CrossRef]
- Li, X.; Bian, J.; Xing, T.; Zhao, L.; Li, J.L.; Zhang, L.; Gao, F. Effects of guanidinoacetic acid supplementation on growth performance, hypothalamus-pituitary-adrenal axis, and immunity of broilers challenged with chronic heat stress. Poult Sci. 2023, 102, 103114. [Google Scholar] [CrossRef]
- Kalia, S.; Bharti, V.K.; Giri, A.; Kumar, B.; Arora, A.; Balaje, S.S. Hippophae rhamnoides as novel phytogenic feed additive for broiler chickens at high altitude cold desert. Sci Rep. 2018, 8, 5954. [Google Scholar] [CrossRef]
- Abdel-Latif, M.A.; Elbestawy, A.R.; El-Far, A.H.; Noreldin, A.E.; Emam, M.; Baty, R.S.; Albadrani, G.M.; Abdel-Daim, M.M.; Abd El-Hamid, H.S. Quercetin Dietary Supplementation Advances Growth Performance, Gut Microbiota, and Intestinal mRNA Expression Genes in Broiler Chickens. Animals 2021, 11, 2302. [Google Scholar] [CrossRef]
- Liu, X.; Huang, X.Y.; Fu, Y.; Wang, Y.Z.; Lu, Z.Q. The Positive Effects of Exogenous Pancreatin on Growth Performance, Nutrient Digestion and Absorption, and Intestinal Microbiota in Piglets. Front. Physiol. 2022, 13, 906522. [Google Scholar] [CrossRef]
- Song, X.; Anas, M.A.; Kurniawati, A.; Hanim, C.; Muhlisin, A.M.A.; Madani, A.M.A.; Wang, Q.; Chen, H. Effects of reduced-protein diets with protease supplementation on growth, carcass yield, intestinal morphology, organ development, nutrient digestibility, and blood biochemical of broiler chickens. Transl. Anim. Sci. 2023, 7, txad098. [Google Scholar] [CrossRef]
- Zou, T.D.; Deng, C.X.; Wang, Z.R.; Ye, Y.L.; You, J.M. Dietary alanyl-glutamine improves growth performance of weaned piglets through maintaining intestinal morphology and digestion-absorption function. Animal 2019, 13, 1826–1833. [Google Scholar] [CrossRef]
- Xing, Y.; Zheng, Y.; Yang, S.; Zhang, L.; Guo, S.; Shi, L.; Xu, Y.; Jin, X.; Yan, S.; Shi, B.L. Artemisia ordosica polysaccharide ameliorated LPS-induced growth inhibition and intestinal injury in broilers through enhancing immune-regulation and antioxidant capacity. J. Nutr. Biochem. 2023, 115, 109284. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zheng, X.; Yang, B.; Yan, M.; Wang, H.; Yang, S.; Shi, D.; Guo, S.; Liu, C. Effect of Wu Zhi San supplementation in LPS-induced intestinal inflammation and barrier damage in broilers. Front. Vet. Sci. 2023, 10, 1234769. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Shang, X.; Lin, G.; Li, H.; Feng, X.; Zhang, H. Effects of Zinc Glycinate on Growth Performance, Serum Biochemical Indexes, and Intestinal Morphology of Yellow Feather Broilers. Biol. Trace Elem. Res. 2022, 200, 4089–4097. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.Y.; Lou, W.J.; Zhang, D.Y.; Sun, S.Y. ROS Plays a Role in the Neonatal Rat Intestinal Barrier Damages Induced by Hyperoxia. Biomed. Res. Int. 2020, 2020, 8819195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Ma, L.; Wan, X.Y.; Shan, J.J.; Qu, Y.G.; Hashimoto, K.J. (R)-Ketamine attenuates LPS-induced endotoxin-derived delirium through inhibition of neuroinflammation. Psychopharmacology 2021, 238, 2743–2753. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Y.; Li, T.; Li, Y.; Zhang, T.; Du, M.; Zhang, Q.; Wang, X.; Hu, G. Protective role of Cecropin AD against LPS-induced intestinal mucosal injury in chickens. Front Immunol. 2023, 14, 1290182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, L.; Zhan, X.; Zeng, X.; Zhou, L.; Cao, G.; Chen, A.; Yang, C. Effects of dietary supplementation of probiotic, Clostridium butyricum, on growth performance, immune response, intestinal barrier function, and digestive enzyme activity in broiler chickens challenged with Escherichia coli K88. J. Anim. Sci. Biotechnol. 2016, 7, 3. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, K.; Zhang, A.; Chang, W.; Zheng, A.; Chen, Z.; Cai, H.; Liu, G.H. Effects of Lactobacillus acidophilus on the growth performance, immune response, and intestinal barrier function of broiler chickens challenged with Escherichia coli O157. Poult. Sci. 2021, 100, 101323. [Google Scholar] [CrossRef]
- Guo, Y.; Yu, Y.; Li, H.; Ding, X.; Li, X.; Jing, X.; Chen, J.; Liu, G.; Lin, Y.; Jiang, C.; et al. Inulin supplementation ameliorates hyperuricemia and modulates gut microbiota in Uox-knockout mice. Eur. J. Nutr. 2021, 60, 2217–2230. [Google Scholar] [CrossRef]
- Zhang, L.H.; Wang, J.; Piao, X.S. Potential Effects of 25-Hydroxycholecalciferol on the Growth Performance, Blood Antioxidant Capacity, Intestinal Barrier Function and Microbiota in Broilers under Lipopolysaccharide Challenge. Antioxidants 2022, 11, 2094. [Google Scholar] [CrossRef]
- Zhang, B.L.; Zhong, Q.Z.; Liu, N.; Song, P.; Zhu, P.; Zhang, C.C.; Sun, Z.W. Dietary Glutamine Supplementation Alleviated Inflammation Responses and Improved Intestinal Mucosa Barrier of LPS-Challenged Broilers. Animals 2022, 12, 1729. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Wang, J.; Lv, Z. Supplementing Genistein for Breeder Hens Alters the Growth Performance and Intestinal Health of Offspring. Life 2023, 13, 1468. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Shan, C.; Geng, S.; Ning, D. Oleocanthal alleviated lipopolysaccharide-induced acute lung injury in chickens by inhibiting TLR4/NF-κB pathway activation. Poult. Sci. 2023, 102, 102458. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Li, X.; Xue, C.; Zhang, L.; Wang, C.; Xu, X.; Shan, A. Astragalus polysaccharides alleviates LPS-induced inflammation via the NF-κB/MAPK signaling pathway. J. Cell Physiol. 2020, 235, 5525–5540. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Qu, Y.Y.; Yin, K.; Zhang, X.T.; Lin, H.J. Selenomethionine ameliorates LPS-induced intestinal immune dysfunction in chicken jejunum. Metallomics 2021, 13, mfab003. [Google Scholar] [CrossRef] [PubMed]
- Kothari, P.; Dhaniya, G.; Sardar, A.; Sinha, S.; Girme, A.; Rai, D.; Chutani, K.; Hingorani, L.; Trivedi, R. A glucuronated flavone TMMG spatially targets chondrocytes to alleviate cartilage degeneration through negative regulation of IL-1β. Biomed. Pharmacother. 2023, 163, 114809. [Google Scholar] [CrossRef]
- Xia, C.X.; Gao, A.X.; Zhu, Y.; Dong, T.T.; Tsim, K.W. Flavonoids from Seabuckthorn (Hippophae rhamnoides L.) restore CUMS-induced depressive disorder and regulate the gut microbiota in mice. Food Funct. 2023, 14, 7426–7438. [Google Scholar] [CrossRef]
- Jiang, M.; Lv, Z.; Huang, Y.; Cheng, Z.; Meng, Z.; Yang, T.; Yan, Q.; Lin, M.; Zhan, K.; Zhao, G. Quercetin Alleviates Lipopolysaccharide-Induced Inflammatory Response in Bovine Mammary Epithelial Cells by Suppressing TLR4/NF-κB Signaling Pathway. Front. Vet. Sci. 2022, 9, 915726. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Li, X.; Xu, F.; Zhao, S.; Wu, X.; Wang, Y.; Xie, J. Kaempferol Alleviates Murine Experimental Colitis by Restoring Gut Microbiota and Inhibiting the LPS-TLR4-NF-κB Axis. Front. Immunol. 2021, 12, 679897. [Google Scholar] [CrossRef]
- He, C.; Yu, W.; Yang, M.; Li, Z.; Yu, J.; Zhong, D.; Deng, S.; Song, Z.; Cheng, S. Qi Fu Yin ameliorates neuroinflammation through inhibiting RAGE and TLR4/NF-κB pathway in AD model rats. Aging 2023, 15, 13239–13264. [Google Scholar] [CrossRef]
- Song, B.; Li, P.; Yan, S.; Liu, Y.; Gao, M.; Lv, H.; Lv, Z.; Guo, Y.M. Effects of Dietary Astragalus Polysaccharide Supplementation on the Th17/Treg Balance and the Gut Microbiota of Broiler Chickens Challenged with Necrotic Enteritis. Front. Immunol. 2022, 13, 781934. [Google Scholar] [CrossRef] [PubMed]
- Adewole, D.; Akinyemi, F. Gut Microbiota Dynamics, Growth Performance, and Gut Morphology in Broiler Chickens Fed Diets Varying in Energy Density with or without Bacitracin Methylene Disalicylate (BMD). Microorganisms 2021, 9, 787. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Hu, X.; Bao, L.; Wu, K.; Zhao, Y.; Xiang, K.; Li, S.; Wang, Y.; Qiu, M.; Feng, L.; et al. Gut dysbiosis induces the development of mastitis through a reduction in host anti-inflammatory enzyme activity by endotoxemia. Microbiome 2022, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Gureev, A.P.; Shaforostova, E.A.; Vitkalova, I.Y.; Sadovnikova, I.S.; Kalinina, Y.I.; Cherednichenko, V.R.; Reznikova, K.A.; Valuyskikh, V.V.; Popov, V.N. Long-term mildronate treatment increased Proteobacteria level in gut microbiome, and caused behavioral deviations and transcriptome change in liver, heart and brain of healthy mice. Toxicol. Appl. Pharmacol. 2020, 398, 115031. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Chi, X.; Wang, H.; Dai, A.; Dong, J.; Liu, S.; Zhang, D. Mechanism of action of buckwheat quercetin in regulating lipid metabolism and intestinal flora via Toll-like receptor 4 or nuclear factor κB pathway in rats on a high-fat diet. Nutrition 2023, 115, 112148. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.X.; Zhang, M.M.; Wang, Y.M.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Yang, Y.; Kong, Y.; Guo, Q.; Xu, Y.; Xing, P.; Sun, Y.; Qian, J.; Xu, R.; Xie, L.; et al. Gut Bacteria Erysipelatoclostridium and Its Related Metabolite Ptilosteroid a Could Predict Radiation-Induced Intestinal Injury. Front. Public Health 2022, 10, 862598. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Liu, Y.; Liu, S.; An, Y.; Xue, H.; Wang, J.; Xia, F.; Chen, X.; Cao, Y.G. Microbiome-metabolome responses of Fuzhuan brick tea crude polysaccharides with immune-protective benefit in cyclophosphamide-induced immunosuppressive mice. Food Res. Int. 2022, 157, 111370. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, % | Content | Nutrient Levels, % 2 | Content |
|---|---|---|---|
| Corn | 56.78 | Metabolizable energy, MJ/kg | 2879 |
| Soybean meal | 29.00 | Crude protein | 20.06 |
| Corn gluten meal | 5.00 | Calcium | 0.921 |
| Wheat bran | 3.60 | Phosphorus | 0.55 |
| Soybean oil | 1.20 | Lysine | 1.20 |
| CaHPO4 | 1.30 | Methionine | 0.54 |
| Limestone | 1.20 | Methionine + Cysteine | 0.88 |
| Sodium chloride | 0.30 | Threonine | 0.77 |
| L-Lysine | 0.54 | Tryptophan | 0.21 |
| DL-Methionine | 0.18 | ||
| Choline (60%) | 0.10 | ||
| Premix 1 | 0.80 | ||
| Total | 100 |
| Genes | Primer 1 | Sequence | Product Size (bp) | GenBank Accession Number |
|---|---|---|---|---|
| Occludin | F:5′-3′ | GATGGACAGCATCAACGACC | 142 | NM_205128.1 |
| R:5′-3′ | CTTGCTTTGGTAGTCTGGGC | |||
| Claudin-1 | F:5′-3′ | ACACCCGTTAACACCAGATTT | 152 | NM_001013611.2 |
| R:5′-3′ | GCATTTTTGGGGTAGCCTCG | |||
| ZO-1 | F:5′-3′ | GCCTACTGCTGCTCCTTACAACTC | 129 | XM_040680630.1 |
| R:5′-3′ | GCTGGATCTATATGCGGCGGTAAG | |||
| MUC-2 | F:5′-3′ | GTGAATGGCACTACGAGCCT | 106 | XM_040701656.2 |
| R:5′-3′ | CTGGGGTAGCAACCTTCCAG | |||
| TLR4 | F:5′-3′ | TGGATCTTTCAAGGTGCCACA | 198 | NM_001030693.2 |
| R:5′-3′ | AGTGTCCGATGGGTAGGTCA | |||
| NF-κB | F:5′-3′ | TCAACGCAGGACCTAAAGACAT | 162 | NM_205134.1 |
| R:5′-3′ | GCAGATAGCCAAGTTCAGGATG | |||
| MyD88 | F:5′-3′ | GGATGATCCGTATGGGCATGG | 171 | NM_001030962.5 |
| R:5′-3′ | ATGGACCACACACACGTTCC | |||
| IL-1β | F:5′-3′ | ACTGGGCA TCAAGGGCTA | 154 | NM_214005.1 |
| R:5′-3′ | GGTAGAAGA TGAAGCGGGTC | |||
| β-actin | F:5′-3′ | TGCGTGACATCAAGGAGAAG | 199 | L08165 |
| R:5′-3′ | TGCCAGGGTACATTGTGGTA |
| Items 2 | Group 3 | SEM 4 | p-Value | |
|---|---|---|---|---|
| CON | SF | |||
| Before LPS challenge (1 to 15 day of age) | ||||
| IW (g) | 32.93 | 32.96 | 0.069 | 0.968 |
| ADG (g) | 15.70 | 15.73 | 0.177 | 0.561 |
| ADFI (g) | 25.07 | 25.17 | 0.179 | 0.826 |
| FCR | 1.60 | 1.61 | 0.014 | 0.175 |
| FW (g) | 268.43 | 266.35 | 2.556 | 0.602 |
| Items 2 | Group 3 | SEM 4 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | LPS | SF | SF + LPS | SF | LPS | SF × LPS | ||
| During LPS challenge (16 to 20 days of age) | ||||||||
| ADG(g) | 26.58 | 23.43 | 28.03 | 25.58 | 0.759 | 0.028 | 0.001 | 0.652 |
| ADFI(g) | 49.90 | 47.54 | 51.20 | 50.31 | 0.882 | 0.079 | 0.032 | 0.414 |
| FCR | 1.88 | 2.04 | 1.83 | 1.97 | 0.062 | 0.346 | 0.022 | 0.878 |
| FW(g) | 400.26 | 386.61 | 404.10 | 396.68 | 3.177 | 0.041 | 0.003 | 0.338 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhi, K.; Gong, F.; Chen, L.; Li, Z.; Li, X.; Mei, H.; Fu, C.; Zhao, Y.; Liu, Z.; He, J. Effects of Sea-Buckthorn Flavonoids on Growth Performance, Serum Inflammation, Intestinal Barrier and Microbiota in LPS-Challenged Broilers. Animals 2024, 14, 2073. https://doi.org/10.3390/ani14142073
Zhi K, Gong F, Chen L, Li Z, Li X, Mei H, Fu C, Zhao Y, Liu Z, He J. Effects of Sea-Buckthorn Flavonoids on Growth Performance, Serum Inflammation, Intestinal Barrier and Microbiota in LPS-Challenged Broilers. Animals. 2024; 14(14):2073. https://doi.org/10.3390/ani14142073
Chicago/Turabian StyleZhi, Kexin, Fanwen Gong, Lele Chen, Zezheng Li, Xiang Li, Huadi Mei, Chenxing Fu, Yurong Zhao, Zhuying Liu, and Jianhua He. 2024. "Effects of Sea-Buckthorn Flavonoids on Growth Performance, Serum Inflammation, Intestinal Barrier and Microbiota in LPS-Challenged Broilers" Animals 14, no. 14: 2073. https://doi.org/10.3390/ani14142073
APA StyleZhi, K., Gong, F., Chen, L., Li, Z., Li, X., Mei, H., Fu, C., Zhao, Y., Liu, Z., & He, J. (2024). Effects of Sea-Buckthorn Flavonoids on Growth Performance, Serum Inflammation, Intestinal Barrier and Microbiota in LPS-Challenged Broilers. Animals, 14(14), 2073. https://doi.org/10.3390/ani14142073