Butyrate Conversion by Sulfate-Reducing and Methanogenic Communities from Anoxic Sediments of Aarhus Bay, Denmark


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sediment Sampling
2.2. Sediment Pore Water Analysis
2.3. Sediment Slurry Incubations
2.4. Analytical Methods
2.5. DNA Extraction
2.6. 16S rRNA Gene Amplicon Pyrosequencing
2.7. Analysis and Interpretation of the Pyrosequencing Data
2.8. Illumina HiSeq Analysis of Archaeal Community
2.9. Intact Polar Lipid (IPL) Extraction and HPLC-ESI/IT/MS Analysis
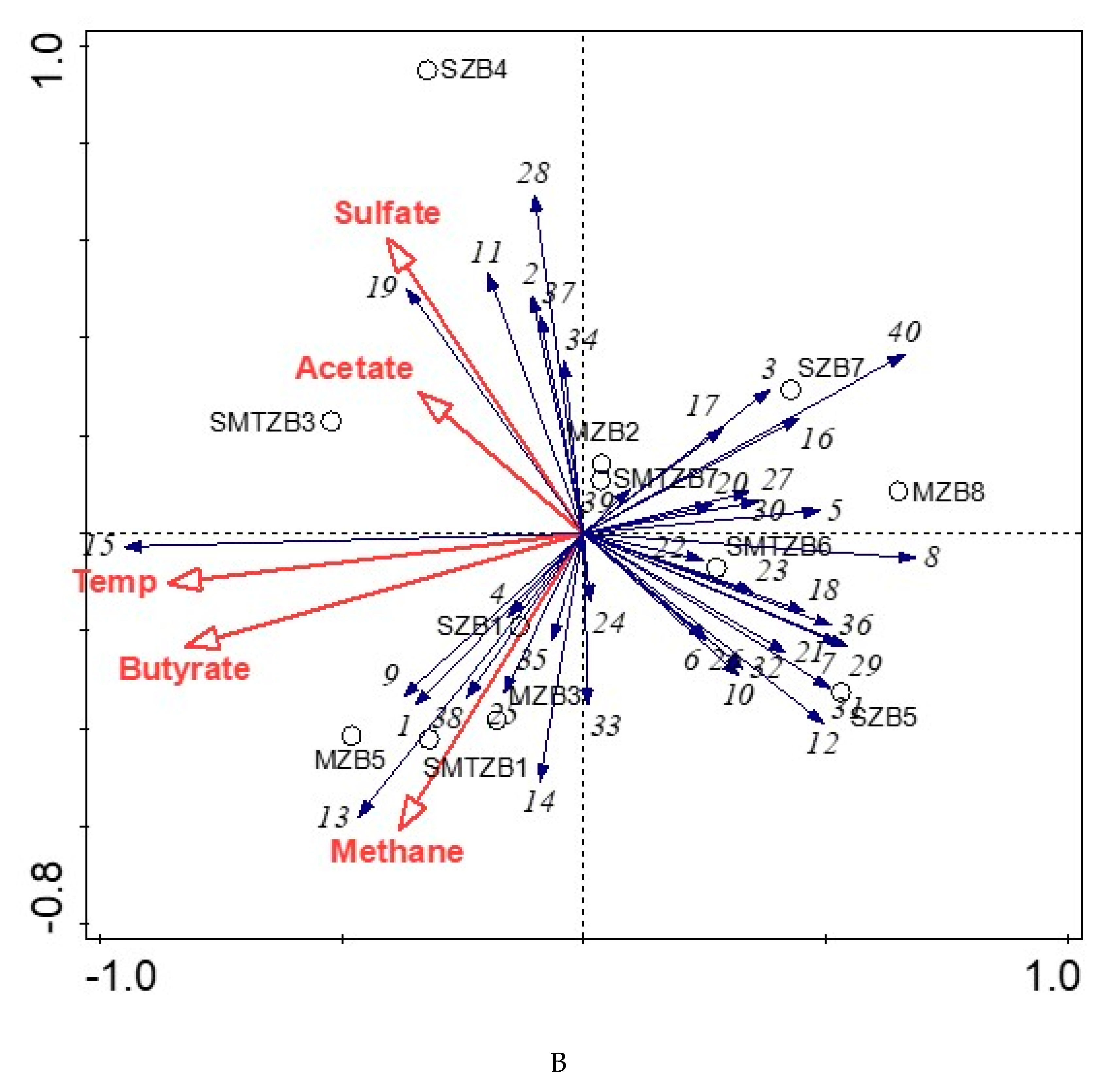
2.10. Statistical Analysis
3. Results
3.1. Sampling Site Geochemistry
3.2. Slurry Incubations
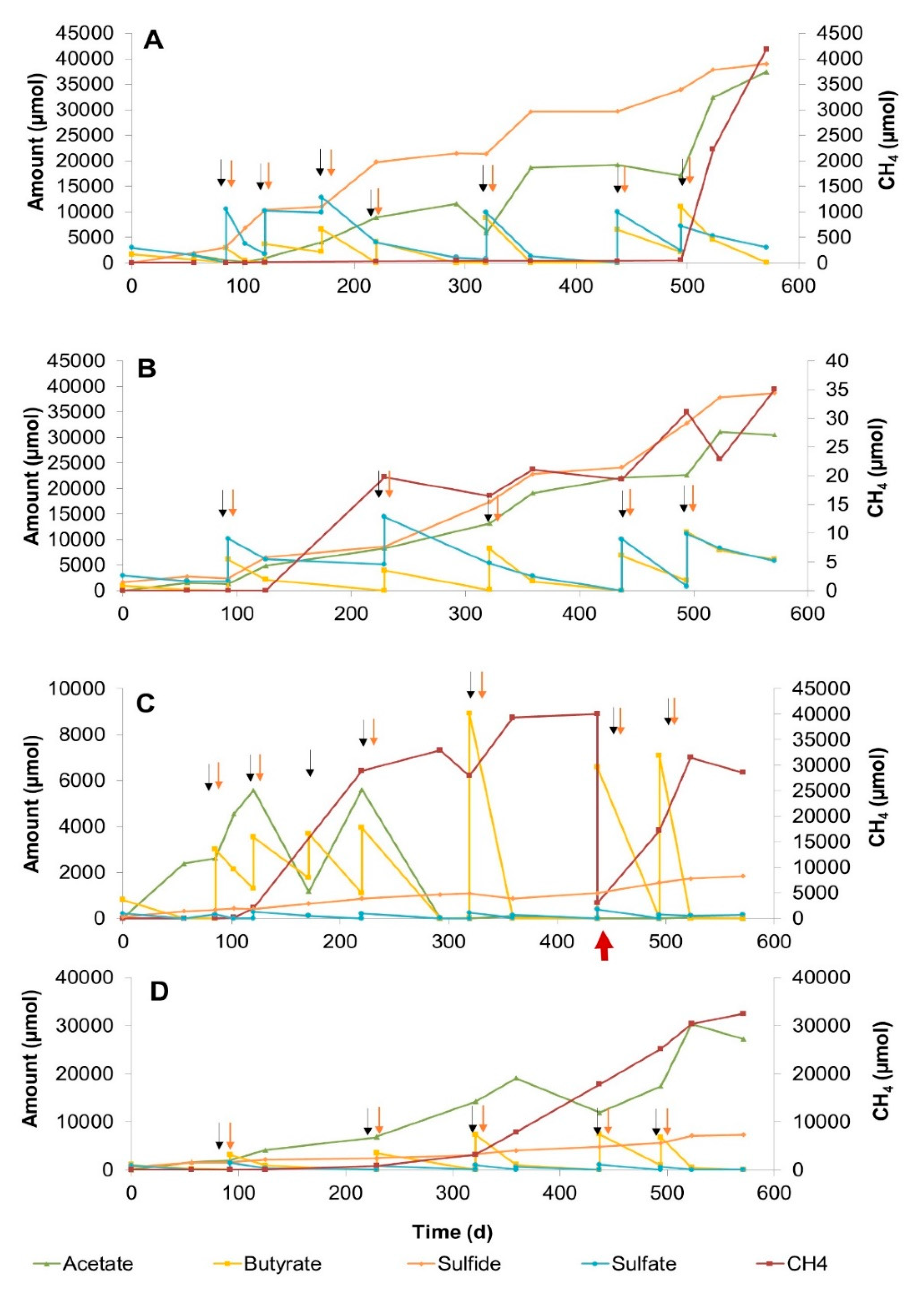
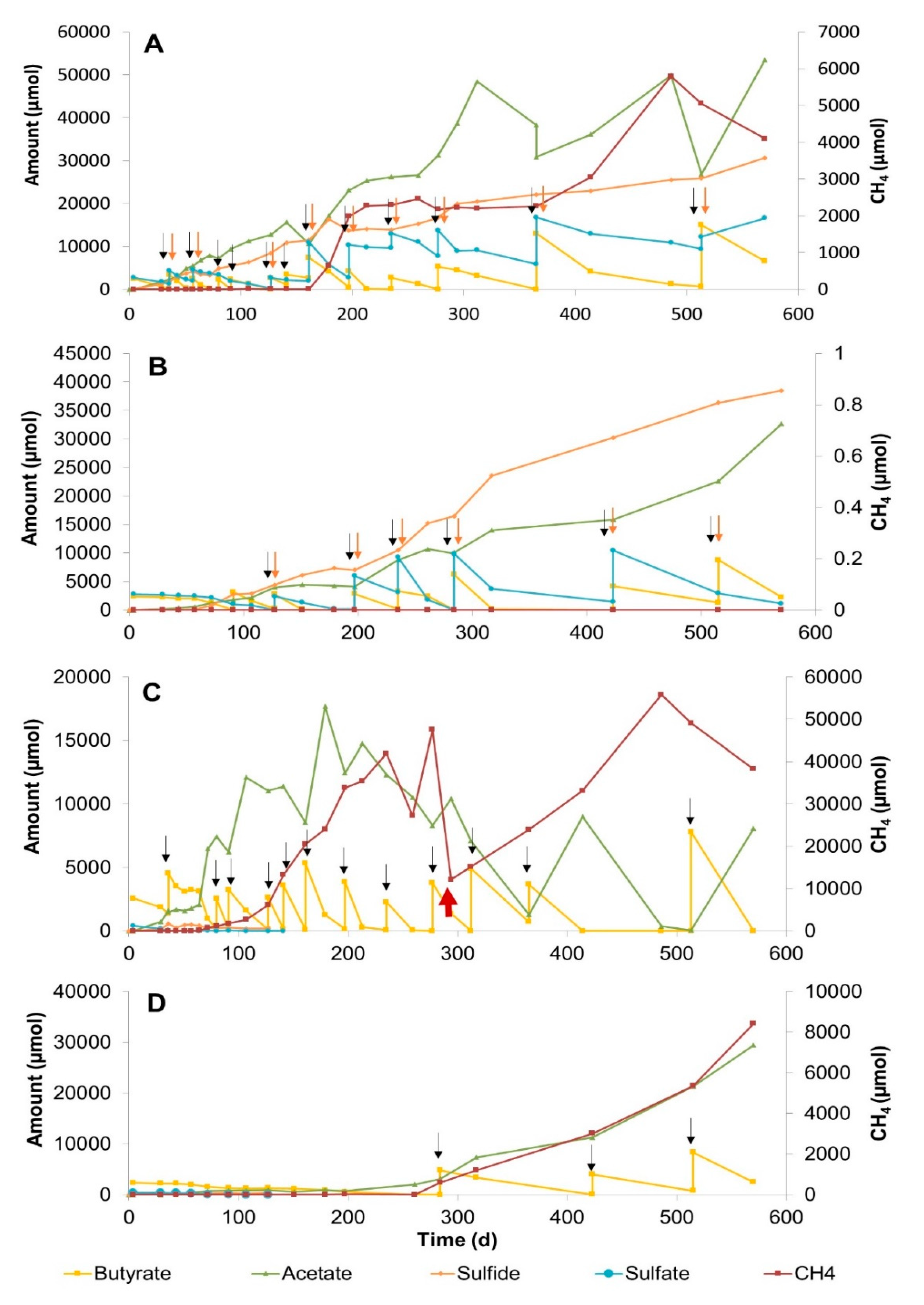
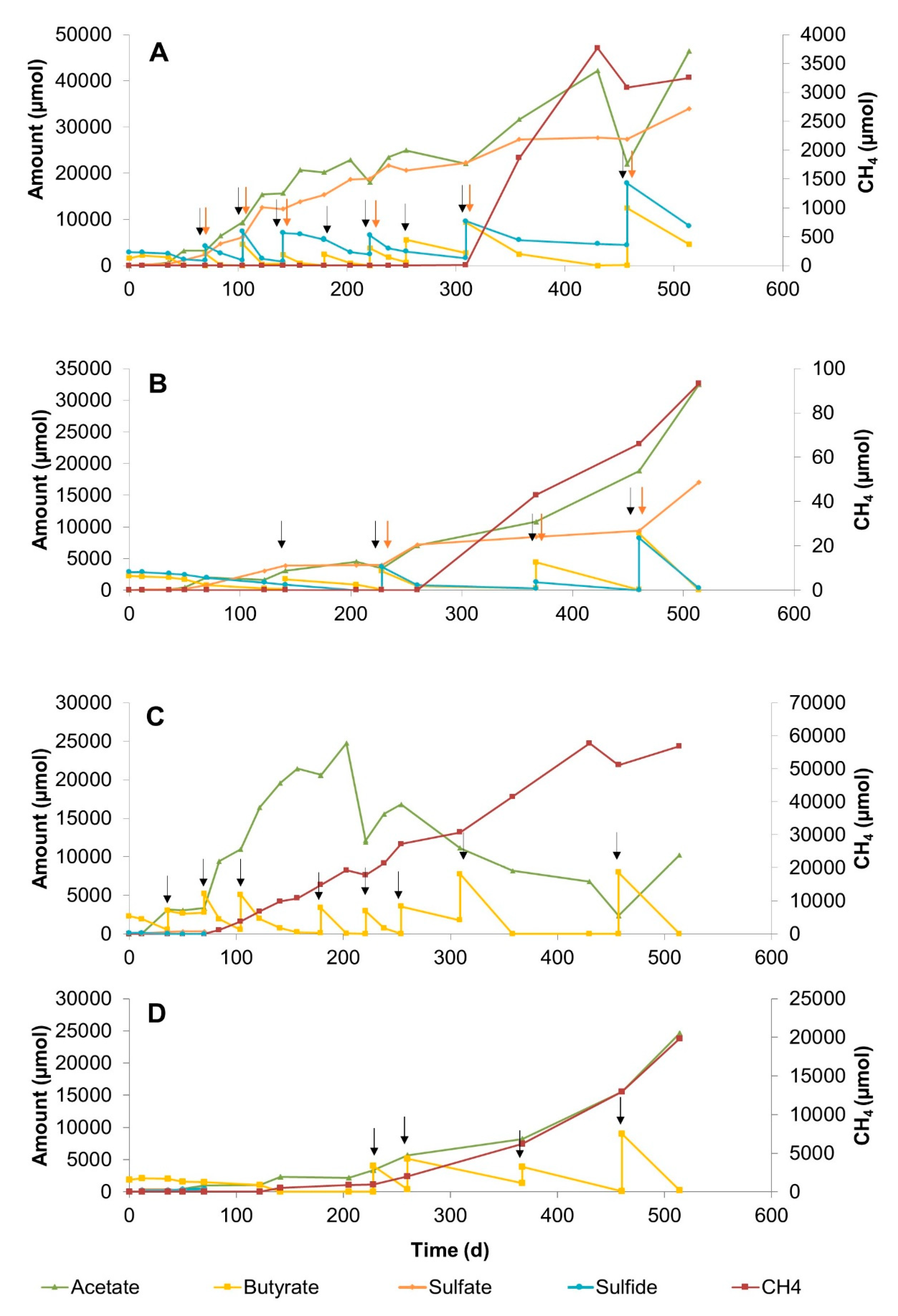
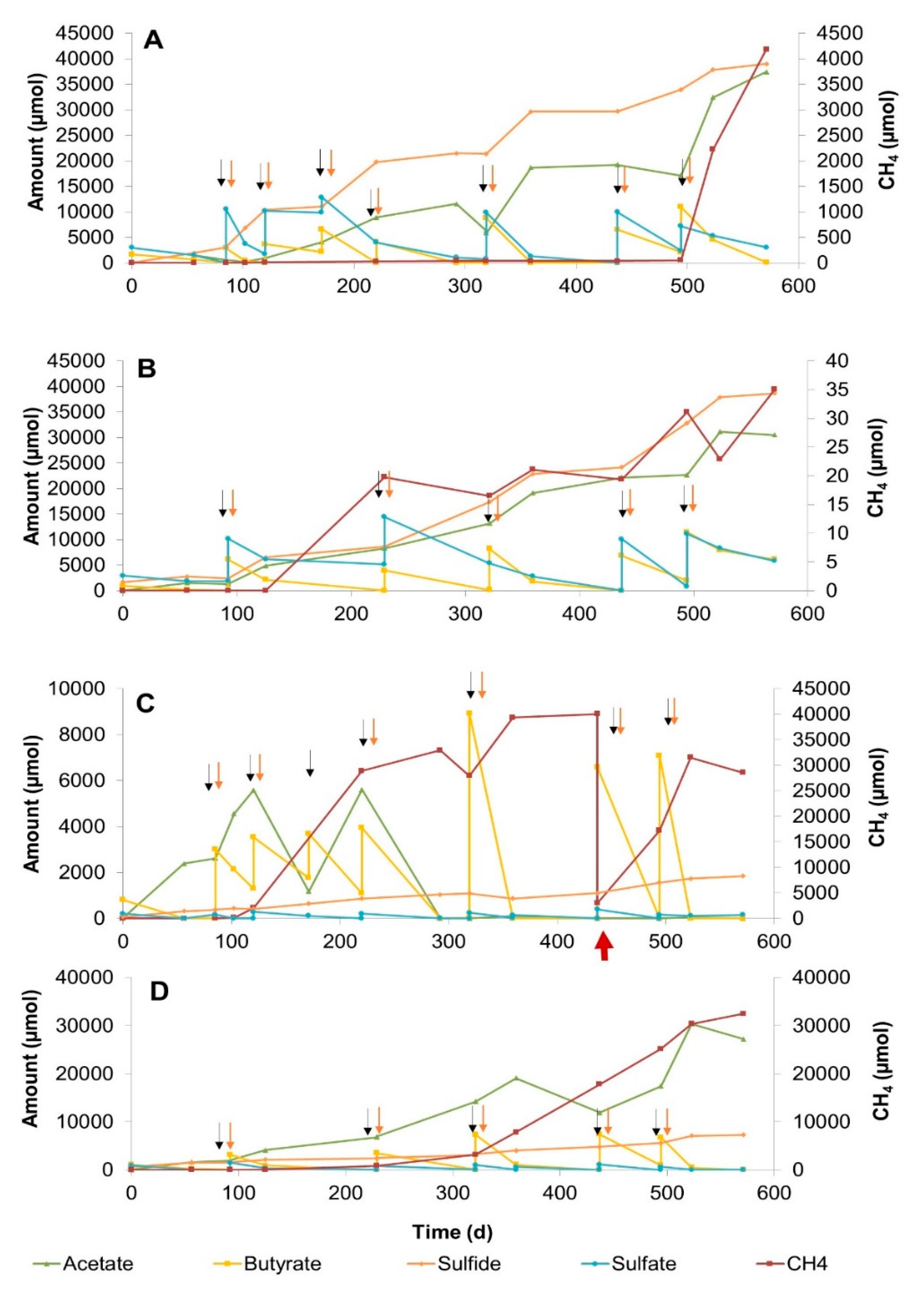
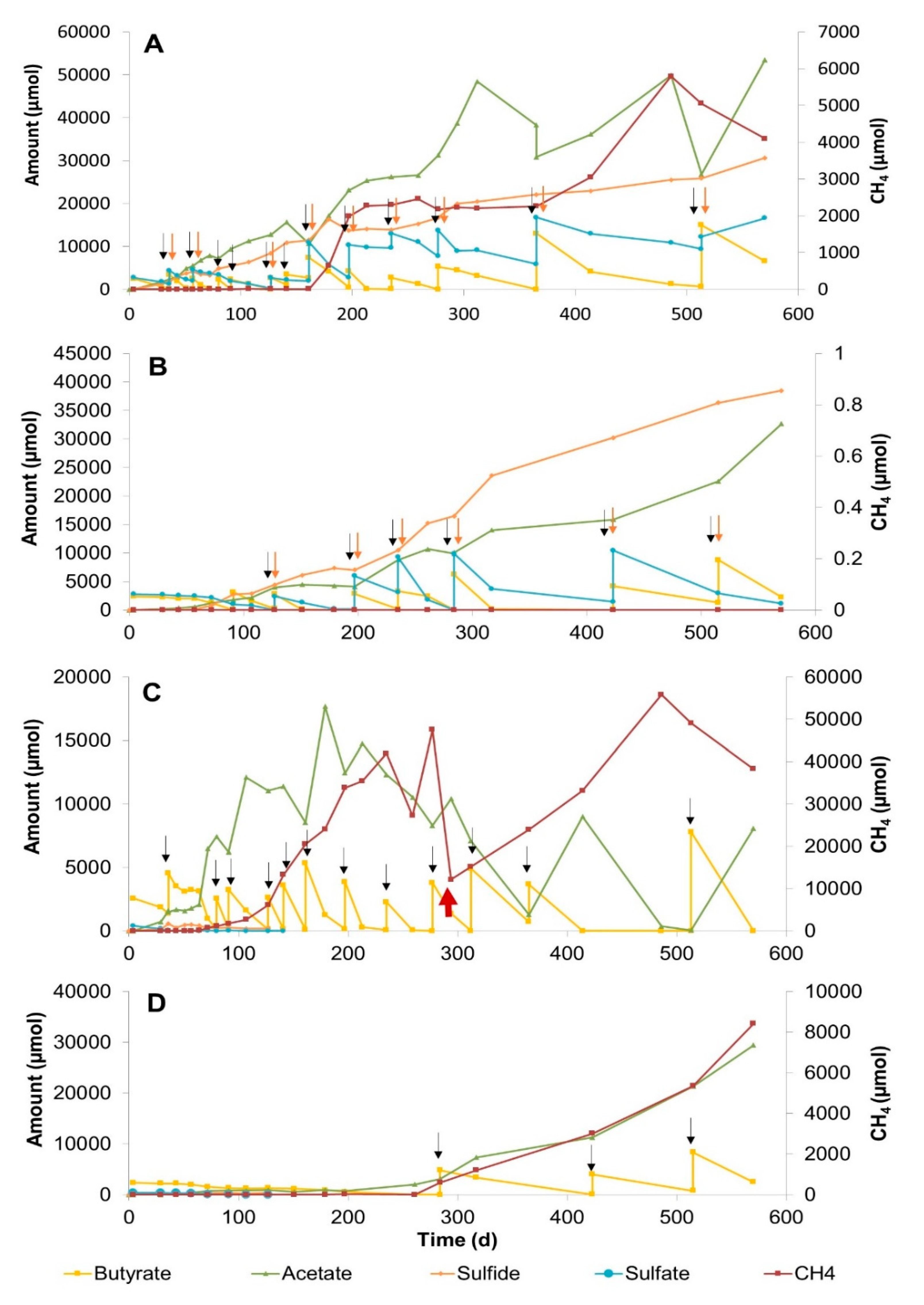
3.2.1. Sulfate Zone Sediment Slurries
3.2.2. Sulfate–Methane Transition Zone Sediment Slurries
3.2.3. Methane Zone Sediment Slurries
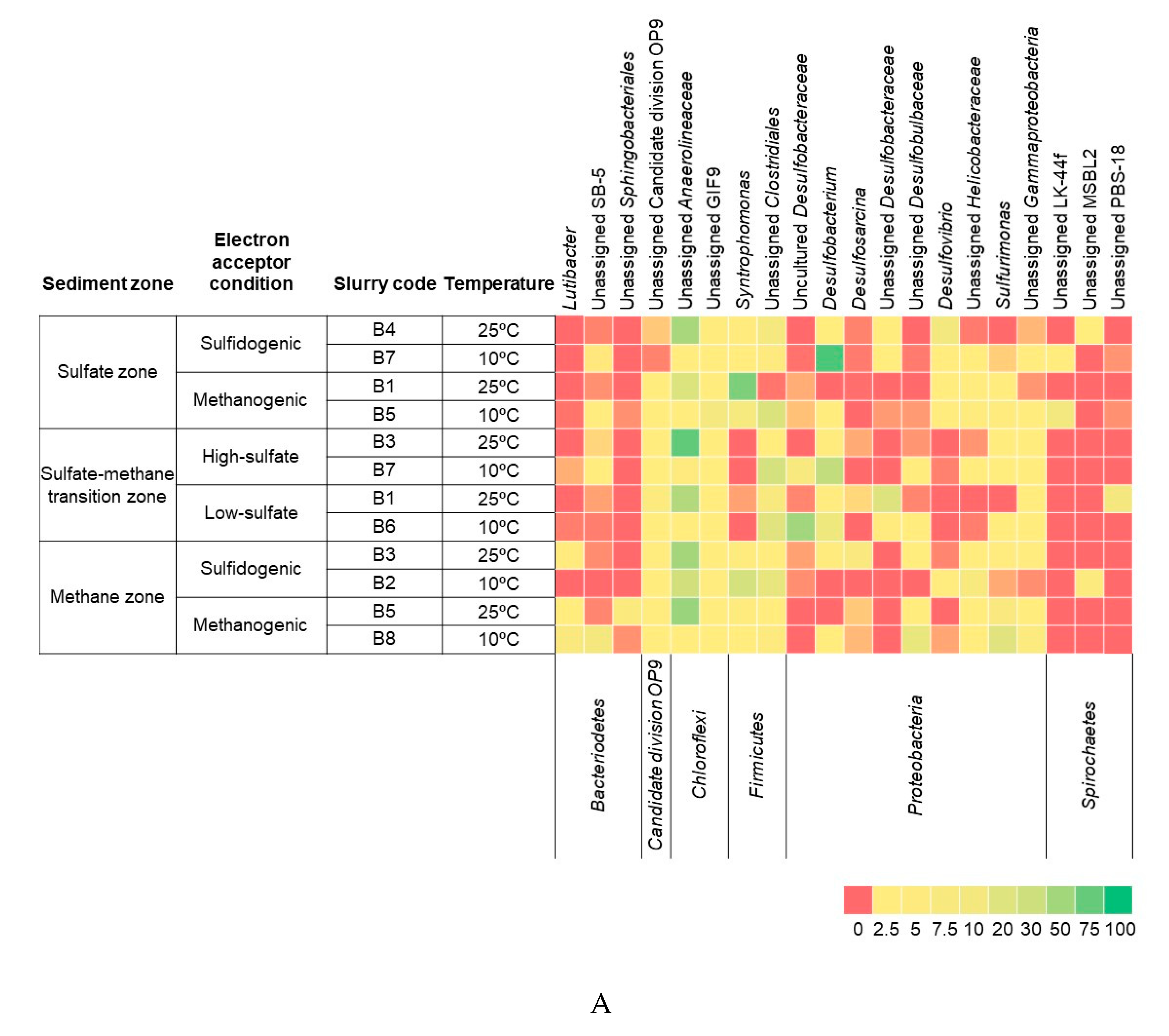
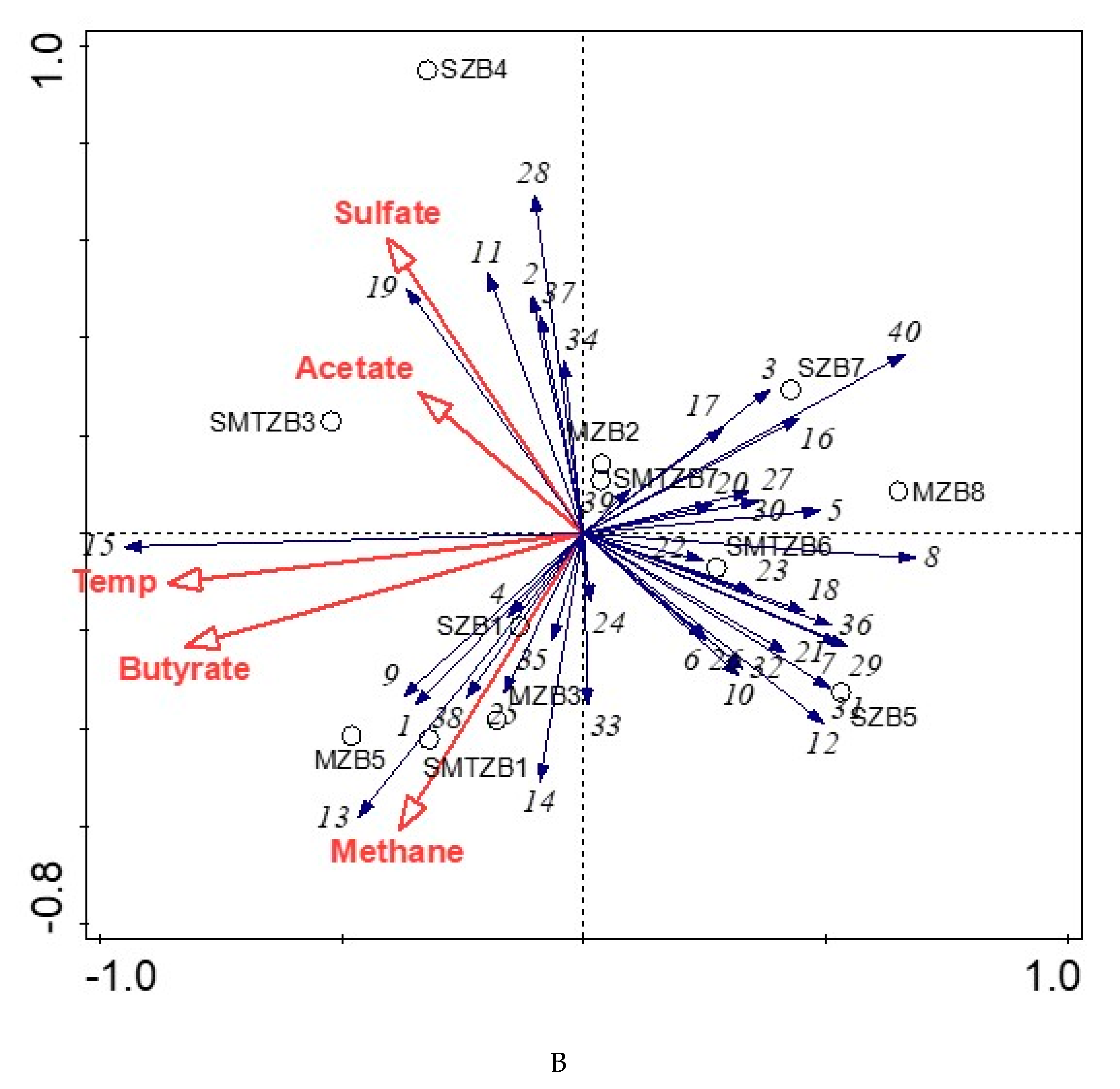
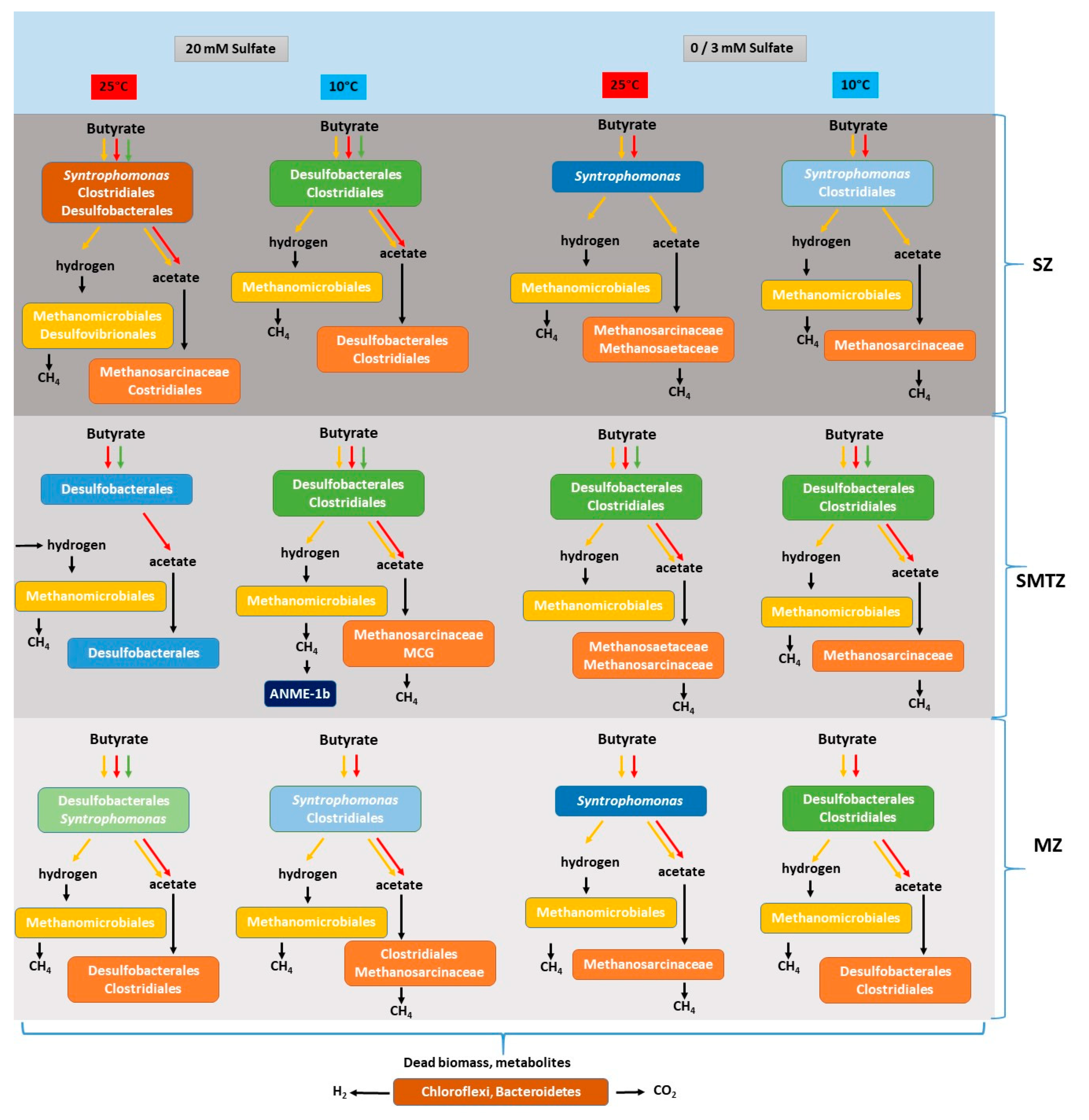
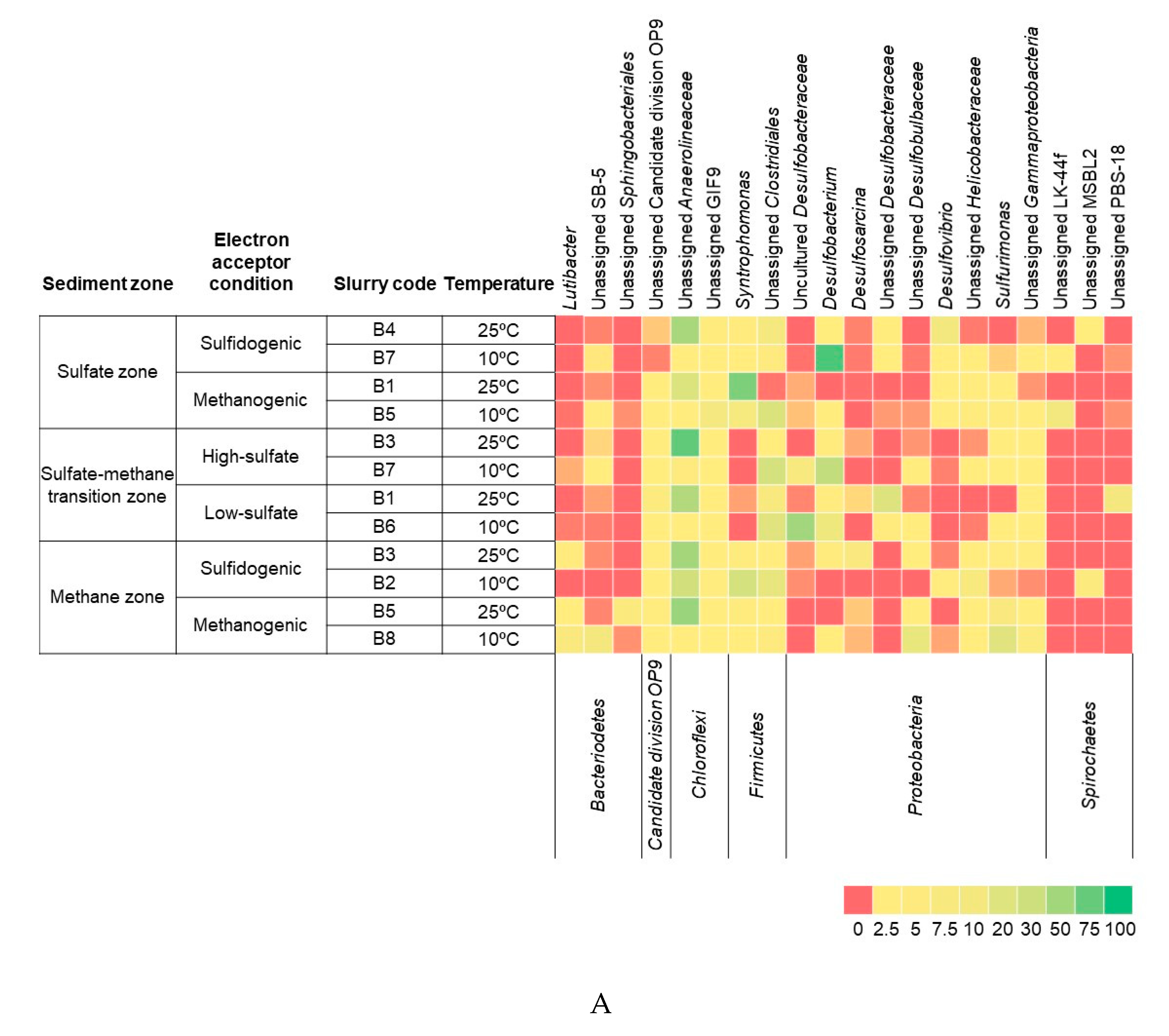
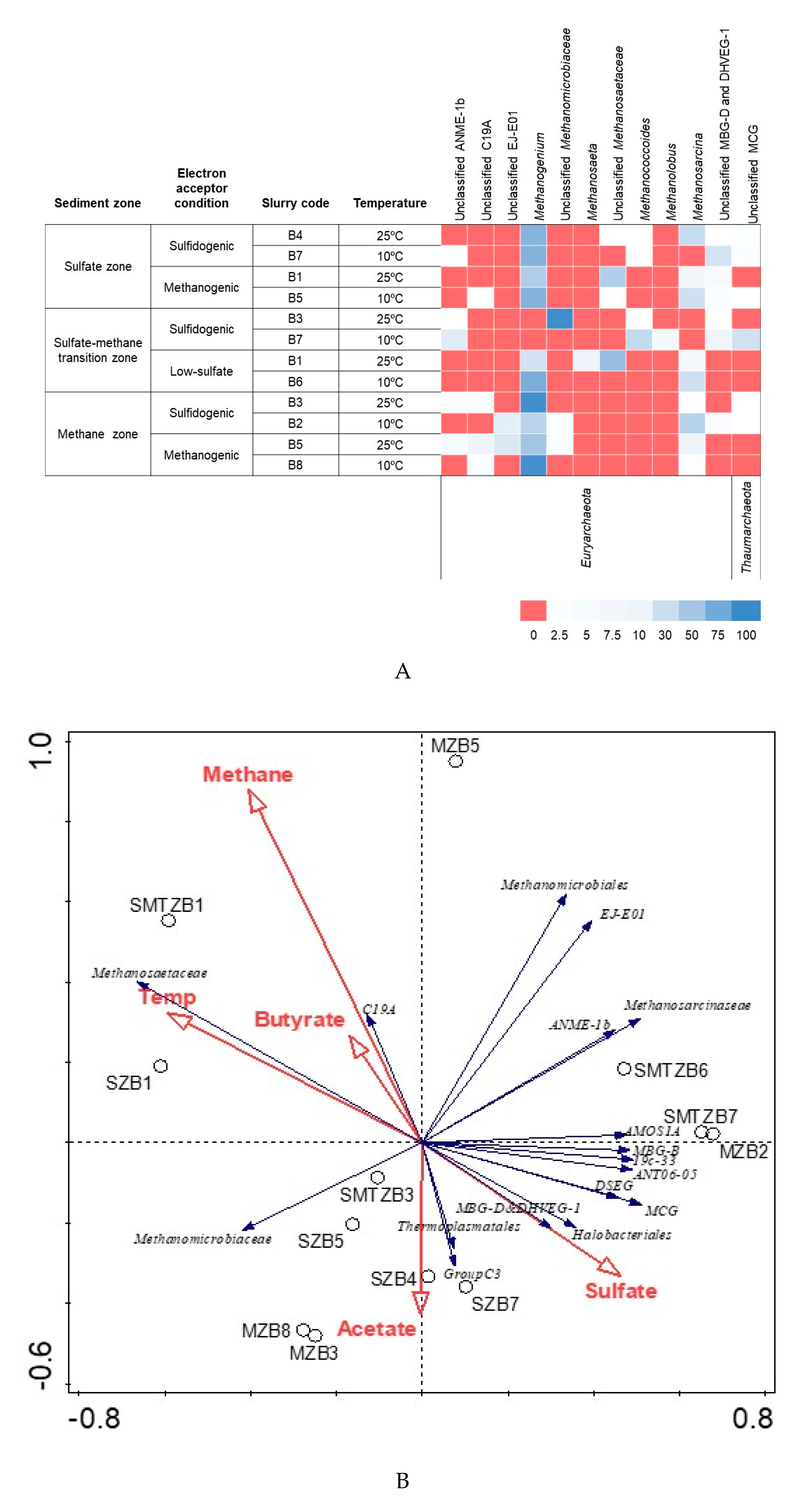
3.3. Bacterial and Archaeal Community Composition
4. Discussion
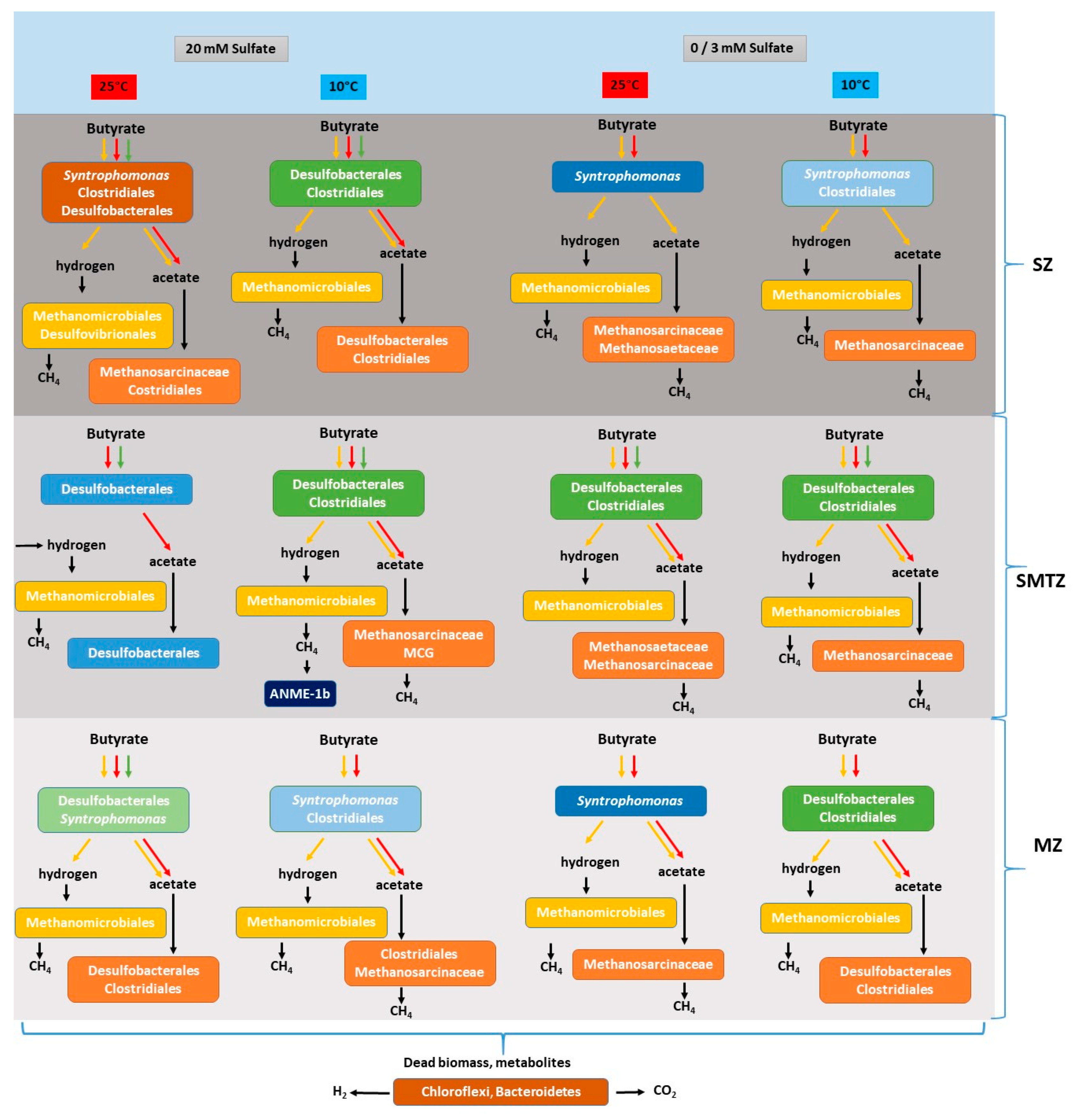
4.1. Butyrate Conversion in Aarhus Bay Sediment
4.2. The Effect of Sulfate Concentration on Butyrate Conversion Dynamics
4.3. Bacterial and Archaeal Community Structure in the Enrichment Slurries
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sequence Data Deposition
References
- Jørgensen, B.B. Bacteria and Marine Biogeochemistry; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2006; Volume 2, pp. 169–206. [Google Scholar]
- Canfield, D.; Kristensen, E.; Thamdrup, B. Heterotrophic Carbon Metabolism; Elsevier BV: Amsterdam, The Netherlands, 2005; Volume 48, pp. 129–166. [Google Scholar]
- Mitterer, R.M. Methanogenesis and sulfate reduction in marine sediments: A new model. Earth Planet. Sci. Lett. 2010, 295, 358–366. [Google Scholar] [CrossRef]
- Glombitza, C.; Egger, M.; Røy, H.; Jørgensen, B.B. Controls on volatile fatty acid concentrations in marine sediments (Baltic Sea). Geochim. Cosmochim. Acta 2019, 258, 226–241. [Google Scholar] [CrossRef]
- Jørgensen, B.B.; Findlay, A.J.; Pellerin, A. The biogeochemical sulfur cycle of marine sediments. Front. Microbiol. 2019, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Schink, B.; Stams, A.J.M. Syntrophism among Prokaryotes. In The Prokaryotes; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2013; pp. 471–493. [Google Scholar]
- Widdel, F. Microbiology and ecology of sulfate- and sulfur-reducing bacteria. In Biology of Anaerobic Microorganisms; Zehnder, A.J.B., Ed.; John Wiley and Sons Inc.: New York, NY, USA, 1988; pp. 469–585. [Google Scholar]
- Maltby, J.; Sommer, S.; Dale, A.W.; Treude, T. Microbial methanogenesis in the sulfate-reducing zone of surface sediments traversing the Peruvian margin. Biogeosciences 2016, 13, 283–299. [Google Scholar] [CrossRef] [Green Version]
- Sela-Adler, M.; Ronen, Z.; Herut, B.; Antler, G.; Vigderovich, H.; Eckert, W.; Sivan, O. Co-existence of methanogenesis and sulfate reduction with common substrates in sulfate-rich estuarine sediments. Front. Microbiol. 2017, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Oremland, R.S.; Polcin, S. Methanogenesis and sulfate reduction: Competitive and noncompetitive substrates in estuarine sediments. Appl. Environ. Microbiol. 1982, 44, 1270–1276. [Google Scholar] [CrossRef] [Green Version]
- Finke, N.; Hoehler, T.M.; Jørgensen, B.B. Hydrogen “leakage” during methanogenesis from methanol and methylamine: Implications for anaerobic carbon degradation pathways in aquatic sediments. Environ. Microbiol. 2007, 9, 1060–1071. [Google Scholar] [CrossRef]
- Leloup, J.; Loy, A.; Knab, N.J.; Borowski, C.; Wagner, M.; Jørgensen, B.B. Diversity and abundance of sulfate-reducing microorganisms in the sulfate and methane zones of a marine sediment, Black Sea. Environ. Microbiol. 2007, 9, 131–142. [Google Scholar] [CrossRef]
- Leloup, J.; Fossing, H.; Kohls, K.; Holmkvist, L.; Borowski, C.; Jørgensen, B.B. Sulfate-reducing bacteria in marine sediment (Aarhus Bay, Denmark): Abundance and diversity related to geochemical zonation. Environ. Microbiol. 2009, 11, 1278–1291. [Google Scholar] [CrossRef]
- Kendall, M.M.; Liu, Y.; Boone, D.R. Butyrate- and propionate-degrading syntrophs from permanently cold marine sediments in Skan Bay, Alaska, and description of Algorimarina butyrica gen. nov., sp. nov. FEMS Microbiol. Lett. 2006, 262, 107–114. [Google Scholar] [CrossRef]
- Kleikemper, J.; Schroth, M.; Sigler, W.V.; Schmucki, M.; Bernasconi, S.; Zeyer, J. Activity and diversity of sulfate-reducing bacteria in a petroleum hydrocarbon-contaminated aquifer. Appl. Environ. Microbiol. 2002, 68, 1516–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dar, S.A.; Kuenen, J.G.; Muyzer, G. Nested PCR-denaturing gradient gel electrophoresis approach to determine the diversity of sulfate-reducing bacteria in complex microbial communities. Appl. Environ. Microbiol. 2005, 71, 2325–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winderl, C.; Anneser, B.; Griebler, C.; Meckenstock, R.U.; Lueders, T. Depth-resolved quantification of anaerobic toluene degraders and aquifer microbial community patterns in distinct redox zones of a tar oil contaminant plume. Appl. Environ. Microbiol. 2007, 74, 792–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struchtemeyer, C.G.; Duncan, K.E.; McInerney, M.J. Evidence for syntrophic butyrate metabolism under sulfate-reducing conditions in a hydrocarbon-contaminated aquifer. FEMS Microbiol. Ecol. 2011, 76, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Plugge, C.M.; Zhang, W.; Scholten, J.C.M.; Stams, A.J.M. Metabolic flexibility of sulfate-reducing bacteria. Front. Microbiol. 2011, 2, 81. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.-Q.; Beulig, F.; Kjeldsen, K.U.; Jørgensen, B.B.; Risgaard-Petersen, N. Concurrent methane production and oxidation in surface sediment from Aarhus Bay, Denmark. Front. Microbiol. 2017, 8, 1198. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Dale, A.; Aguilera, D.R.; Regnier, P.; Fossing, H.; Knab, N.J.; Jørgensen, B.B. Seasonal dynamics of the depth and rate of anaerobic oxidation of methane in Aarhus Bay (Denmark) sediments. J. Mar. Res. 2008, 66, 127–155. [Google Scholar] [CrossRef] [Green Version]
- Van Gelder, A.H.; Aydin, R.; Alves, M.M.; Stams, A.J.M. 1,3-Propanediol production from glycerol by a newly isolated Trichococcus strain. Microb. Biotechnol. 2012, 5, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Ozuolmez, D.; Stams, A.J.M.; Plugge, C.M. Propionate converting anaerobic microbial communities enriched from distinct biogeochemical zones of Aarhus Bay, Denmark under sulfidogenic and methanogenic conditions. Microorganisms 2020, 8, 394. [Google Scholar] [CrossRef] [Green Version]
- Timmers, P.H.A.; Widjaja-Greefkes, H.C.A.; Ramiro-Garcia, J.; Plugge, C.M.; Stams, A.J.M. Growth and activity of ANME clades with different sulfate and sulfide concentrations in the presence of methane. Front. Microbiol. 2015, 6, 869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, D.; Ben Mahfoudh, K.; Wagner, M.; Loy, A. Barcoded primers used in multiplex amplicon pyrosequencing bias amplification. Appl. Environ. Microbiol. 2011, 77, 7846–7849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azman, S.; Khadem, A.F.; Plugge, C.M.; Stams, A.J.M.; Bec, S.; Zeeman, G. Effect of humic acid on anaerobic digestion of cellulose and xylan in completely stirred tank reactors: Inhibitory effect, mitigation of the inhibition and the dynamics of the microbial communities. Appl. Microbiol. Biotechnol. 2016, 101, 889–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poncheewin, W.; Hermes, G.D.A.; Van Dam, J.C.J.; Koehorst, J.J.; Smidt, H.; Schaap, P.J. NG-Tax 2.0: A semantic framework for high-throughput amplicon analysis. Front. Genet. 2020, 10, 1366. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Rütters, H.; Sass, H.; Cypionka, H.; Rullkötter, J. Phospholipid analysis as a tool to study complex microbial communities in marine sediments. J. Microbiol. Methods 2002, 48, 149–160. [Google Scholar] [CrossRef]
- Pitcher, A.; Hopmans, E.C.; Schouten, S.; Damsté, J.S.S. Separation of core and intact polar archaeal tetraether lipids using silica columns: Insights into living and fossil biomass contributions. Org. Geochem. 2009, 40, 12–19. [Google Scholar] [CrossRef]
- Sturt, H.F.; Summons, R.E.; Smith, K.; Elvert, M.; Hinrichs, K.-U. Intact polar membrane lipids in prokaryotes and sediments deciphered by high-performance liquid chromatography/electrospray ionization multistage mass spectrometry—new biomarkers for biogeochemistry and microbial ecology. Rapid Commun. Mass Spectrom. 2004, 18, 617–628. [Google Scholar] [CrossRef]
- Moore, E.K.; Hopmans, E.C.; Rijpstra, W.I.C.; Villanueva, L.; Damsté, J.S.S. Elucidation and identification of amino acid containing membrane lipids using liquid chromatography/high-resolution mass spectrometry. Rapid Commun. Mass Spectrom. 2016, 30, 739–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGarigal, K.; Stafford, S.; Cushman, S. Multivariate Statistics for Wildlife and Ecology Research; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2000. [Google Scholar]
- Muyzer, G.; Stams, A.J.M. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Genet. 2008, 6, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Pender, S.; Toomey, M.; Carton, M.; Eardly, D.; Patching, J.W.; Colleran, E.; O’Flaherty, V. Long-term effects of operating temperature and sulphate addition on the methanogenic community structure of anaerobic hybrid reactors. Water Res. 2004, 38, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Kuever, J. The Family Desulfobacteraceae; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2014; pp. 45–73. [Google Scholar]
- Jochum, L.M.; Chen, X.; Lever, M.A.; Loy, A.; Jørgensen, B.B.; Schramm, A.; Kjeldsen, K.U. Depth distribution and assembly of sulfate-reducing microbial communities in marine sediments of Aarhus Bay. Appl. Environ. Microbiol. 2017, 83, e01547-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oude Elferink, S.J.W.H.; Visser, A.; Hulshoff Pol, L.W.; Stams, A.J.M. Sulfate reduction in methanogenic bioreactors. FEMS Microbiol. Rev. 1994, 15, 119–136. [Google Scholar]
- McInerney, M.J.; Beaty, P.S. Anaerobic community structure from a nonequilibrium thermodynamic perspective. Can. J. Microbiol. 1988, 34, 487–493. [Google Scholar] [CrossRef]
- Kleinsteuber, S.; Schleinitz, K.M.; Vogt, C. Key players and team play: Anaerobic microbial communities in hydrocarbon-contaminated aquifers. Appl. Microbiol. Biotechnol. 2012, 94, 851–873. [Google Scholar] [CrossRef]
- Webster, G.; Sass, H.; Cragg, B.A.; Gorra, R.; Knab, N.J.; Green, C.J.; Mathes, F.; Fry, J.C.; Weightman, A.; Parkes, R.J. Enrichment and cultivation of prokaryotes associated with the sulphate-methane transition zone of diffusion-controlled sediments of Aarhus Bay, Denmark, under heterotrophic conditions. FEMS Microbiol. Ecol. 2011, 77, 248–263. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Sekiguchi, Y. Cultivation of uncultured chloroflexi subphyla: Significance and ecophysiology of formerly uncultured chloroflexi ‘subphylum i’ with natural and biotechnological relevance. Microbes Environ. 2009, 24, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Hug, L.A.; Castelle, C.; Wrighton, K.; Thomas, B.C.; Sharon, I.; Frischkorn, K.R.; Williams, K.H.; Tringe, S.G.; Banfield, J.F. Community genomic analyses constrain the distribution of metabolic traits across the Chloroflexi phylum and indicate roles in sediment carbon cycling. Microbiome 2013, 1, 22. [Google Scholar] [CrossRef] [Green Version]
- Rütters, H.; Sass, H.; Cypionka, H.; Rullkötter, J. Monoalkylether phospholipids in the sulfate-reducing bacteria Desulfosarcina variabilis and Desulforhabdus amnigenus. Arch. Microbiol. 2001, 176, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Rossel, P.; Lipp, J.S.; Fredricks, H.F.; Arnds, J.; Boetius, A.; Elvert, M.; Hinrichs, K.-U. Intact polar lipids of anaerobic methanotrophic archaea and associated bacteria. Org. Geochem. 2008, 39, 992–999. [Google Scholar] [CrossRef]
- Moore, E.K.; Hopmans, E.C.; Rijpstra, W.I.C.; Villanueva, L.; Dedysh, S.; Kulichevskaya, I.S.; Wienk, H.; Schoutsen, F.; Damsté, J.S.S. Novel mono-, di-, and trimethylornithine membrane lipids in northern wetland planctomycetes. Appl. Environ. Microbiol. 2013, 79, 6874–6884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, E.K.; Villanueva, L.; Hopmans, E.C.; Rijpstra, W.I.C.; Mets, A.; Dedysh, S.; Damsté, J.S.S. Abundant trimethylornithine lipids and specific gene sequences are indicative of planctomycete importance at the oxic/anoxic interface in sphagnum-dominated northern wetlands. Appl. Environ. Microbiol. 2015, 81, 6333–6344. [Google Scholar] [CrossRef] [Green Version]
- Bale, N.J.; Hopmans, E.C.; Schoon, P.L.; De Kluijver, A.; Downing, J.A.; Middelburg, J.J.; Damsté, J.S.S.; Schouten, S. Impact of trophic state on the distribution of intact polar lipids in surface waters of lakes. Limnol. Oceanogr. 2016, 61, 1065–1077. [Google Scholar] [CrossRef] [Green Version]
- Boyer, G.M.; Schubotz, F.; Summons, R.E.; Woods, J.; Shock, E.L. Carbon oxidation state in microbial polar lipids suggests adaptation to hot spring temperature and redox gradients. Front. Microbiol. 2020, 11, 229. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.-L.; Ollivier, B.; Whitman, W.B. The Order Methanomicrobiales; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2006; pp. 208–230. [Google Scholar]
- Parkes, R.; Cragg, B.; Roussel, E.; Webster, G.; Weightman, A.; Sass, H. A review of prokaryotic populations and processes in sub-seafloor sediments, including biosphere:geosphere interactions. Mar. Geol. 2014, 352, 409–425. [Google Scholar] [CrossRef]
- Jetten, M.S.M.; Zehnder, A.J.B.; Stams, A.J.M. Methanogenesis from acetate: A comparison of the acetate metabolism in Methanothrix soehngenii and Methanosarcina spp. FEMS Microbiol. Lett. 1992, 88, 181–198. [Google Scholar] [CrossRef]
- Stams, A.J.M.; Plugge, C.M.; De Bok, F.A.M.; Van Houten, B.H.G.W.; Lens, P.N.L.; Dijkman, H.; Weijma, J. Metabolic interactions in methanogenic and sulfate-reducing bioreactors. Water Sci. Technol. 2005, 52, 13–20. [Google Scholar] [CrossRef]
- Struchtemeyer, C.G.; Elshahed, M.S.; Duncan, K.E.; McInerney, M.J. Evidence for aceticlastic methanogenesis in the presence of sulfate in a gas condensate-contaminated aquifer. Appl. Environ. Microbiol. 2005, 71, 5348–5353. [Google Scholar] [CrossRef] [Green Version]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozuolmez, D.; Moore, E.K.; Hopmans, E.C.; Sinninghe Damsté, J.S.; Stams, A.J.M.; Plugge, C.M. Butyrate Conversion by Sulfate-Reducing and Methanogenic Communities from Anoxic Sediments of Aarhus Bay, Denmark. Microorganisms 2020, 8, 606. https://doi.org/10.3390/microorganisms8040606
Ozuolmez D, Moore EK, Hopmans EC, Sinninghe Damsté JS, Stams AJM, Plugge CM. Butyrate Conversion by Sulfate-Reducing and Methanogenic Communities from Anoxic Sediments of Aarhus Bay, Denmark. Microorganisms. 2020; 8(4):606. https://doi.org/10.3390/microorganisms8040606
Chicago/Turabian StyleOzuolmez, Derya, Elisha K. Moore, Ellen C. Hopmans, Jaap S. Sinninghe Damsté, Alfons J. M. Stams, and Caroline M. Plugge. 2020. "Butyrate Conversion by Sulfate-Reducing and Methanogenic Communities from Anoxic Sediments of Aarhus Bay, Denmark" Microorganisms 8, no. 4: 606. https://doi.org/10.3390/microorganisms8040606