
Effects of Polycyclic Aromatic Hydrocarbons on the Composition of the Soil Bacterial Communities in the Tidal Flat Wetlands of the Yellow River Delta of China


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Soil Sampling
2.2. Physicochemical Properties of Soil
2.3. DNA Extraction and 16 S rDNA Sequencing
2.4. Bioinformatics and Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
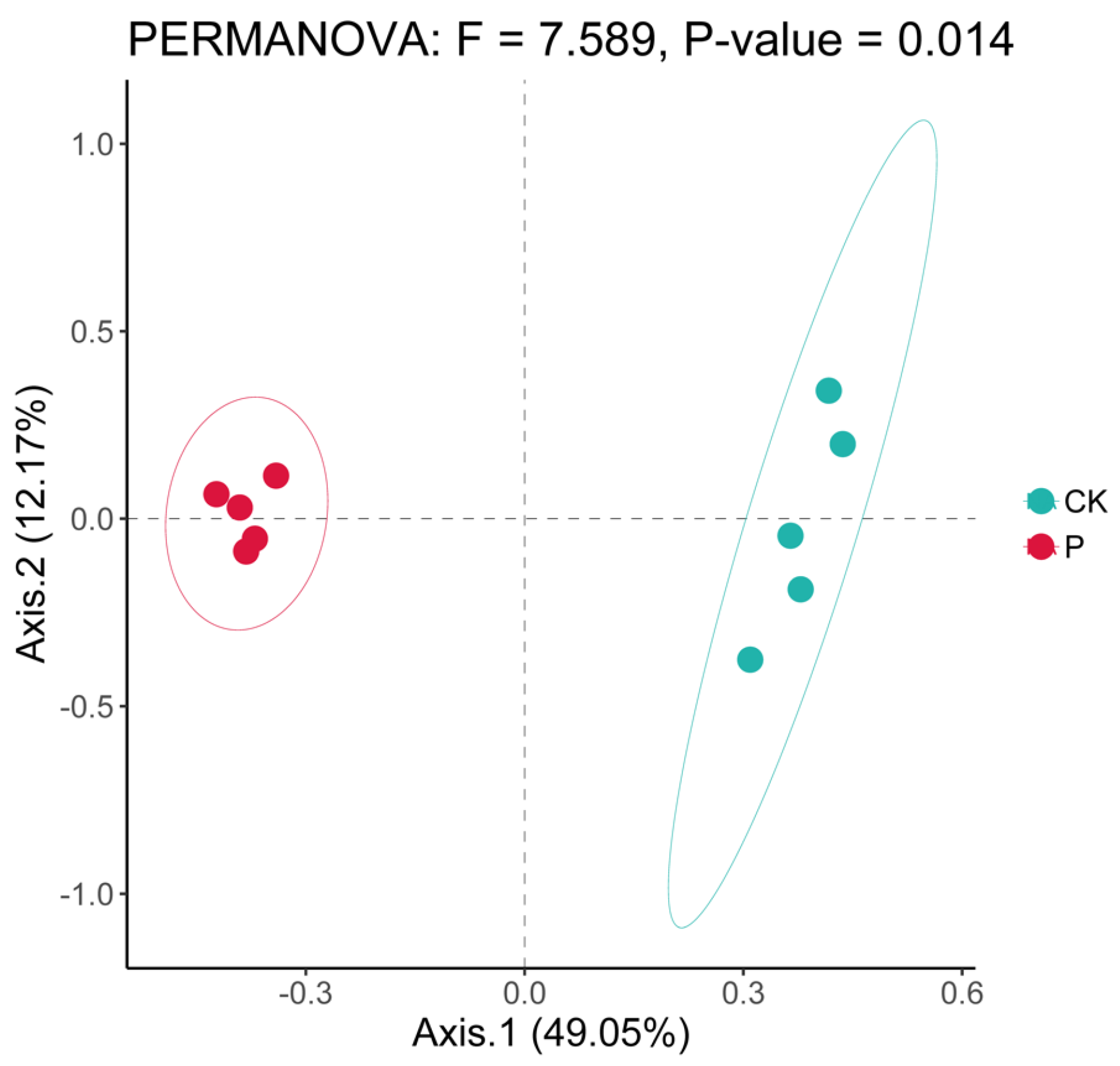
3.2. Soil Bacterial α- and β-Diversities
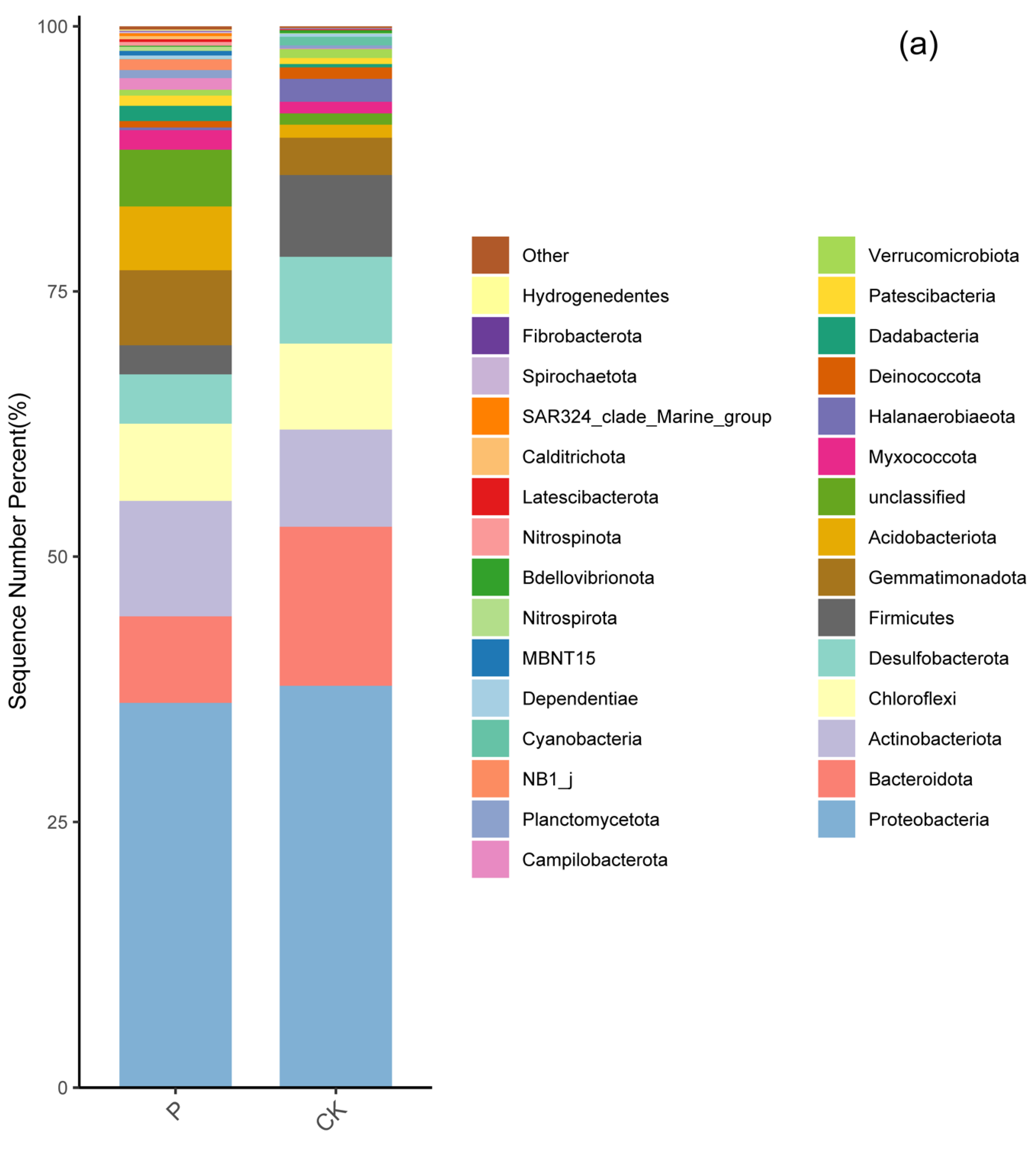
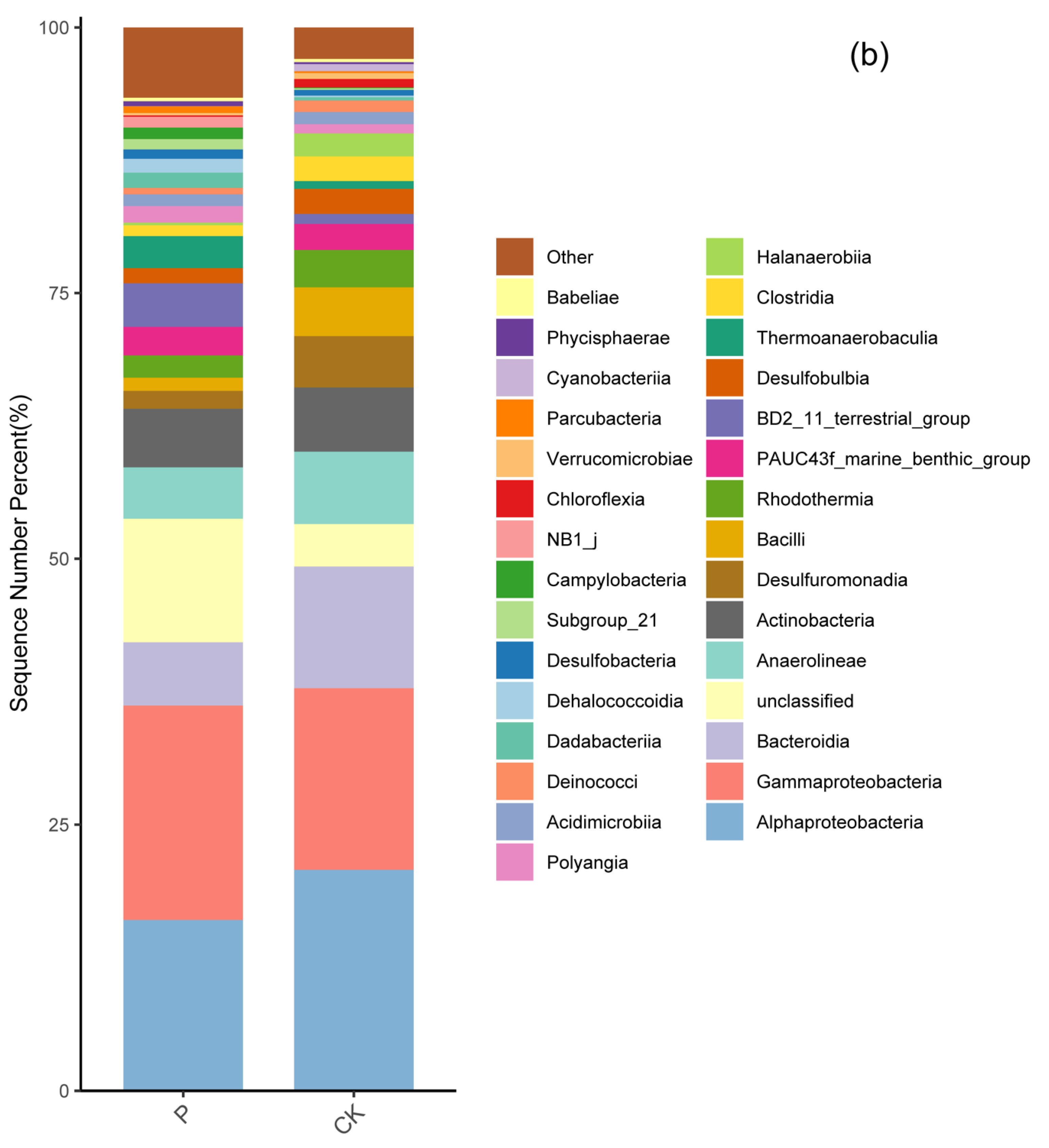
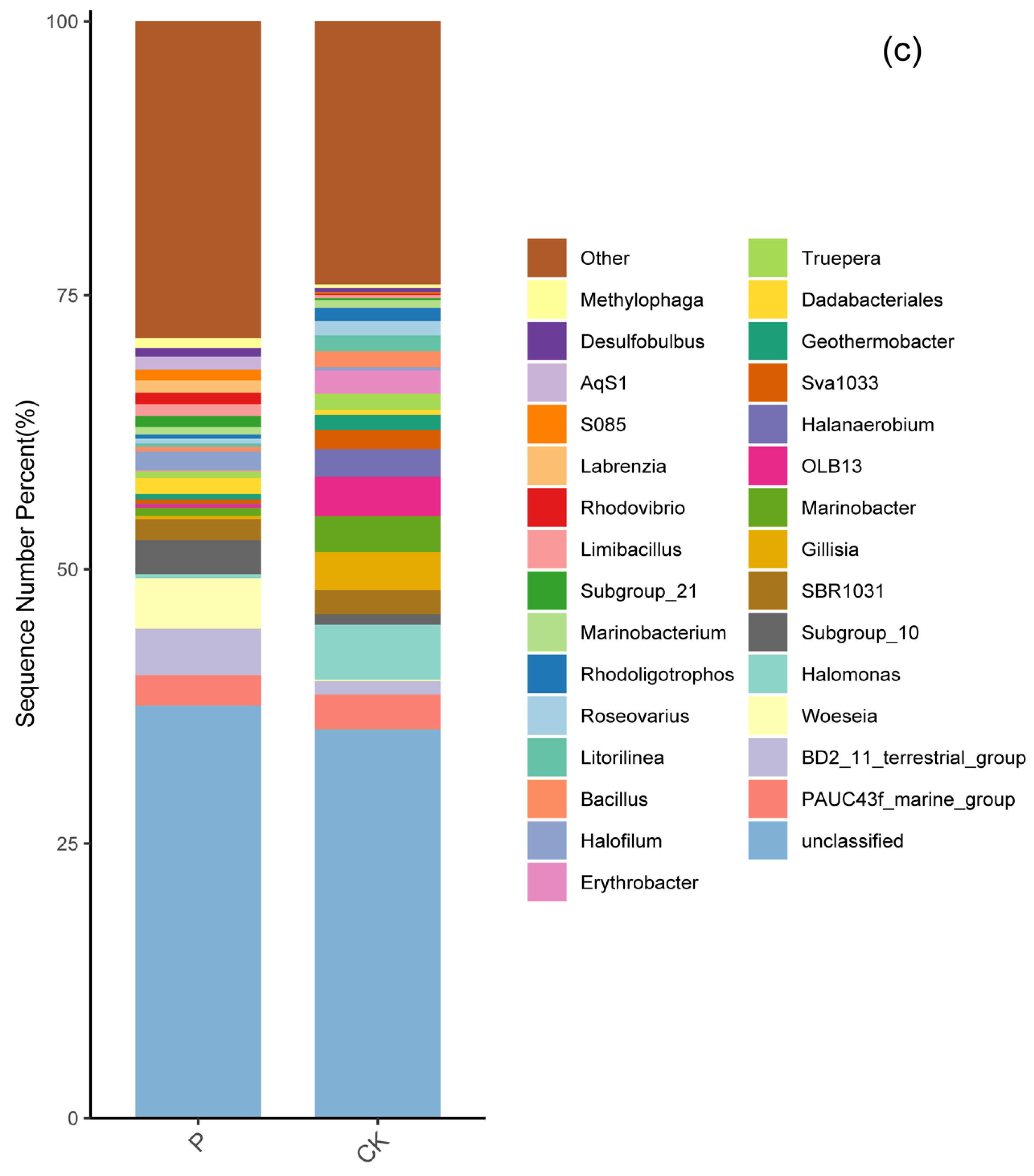
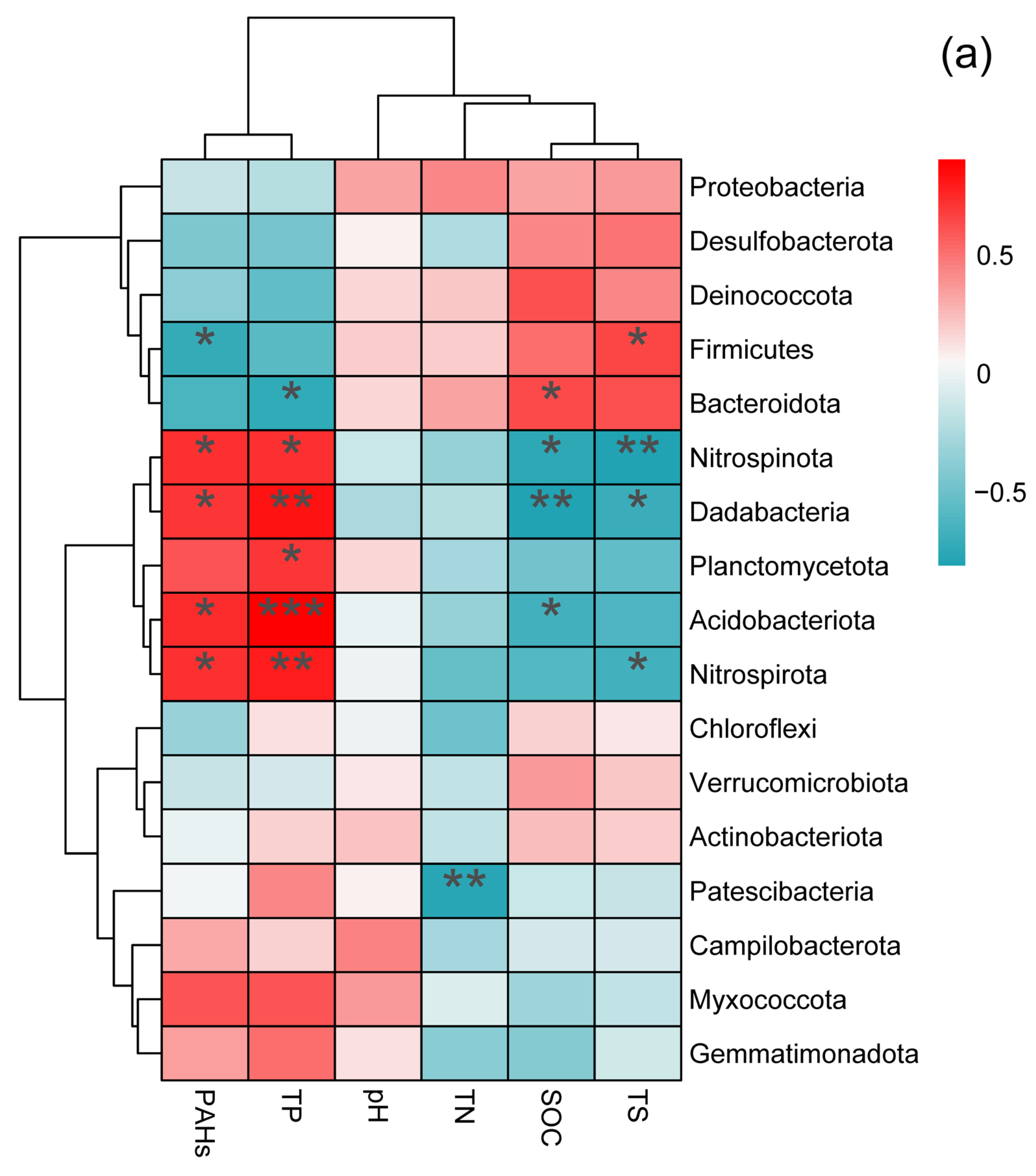
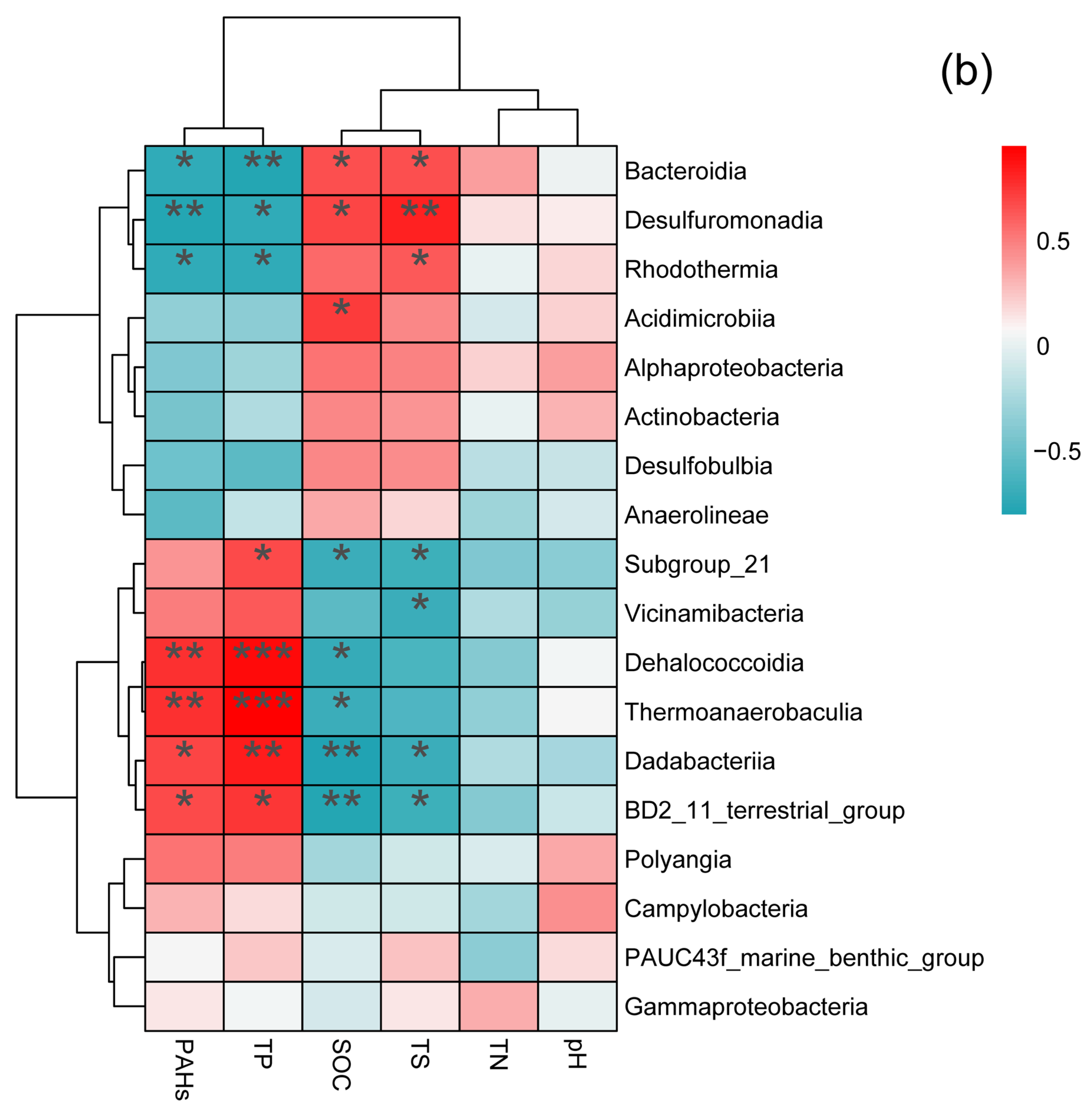
3.3. Soil Bacterial Community Composition in Two Wetlands
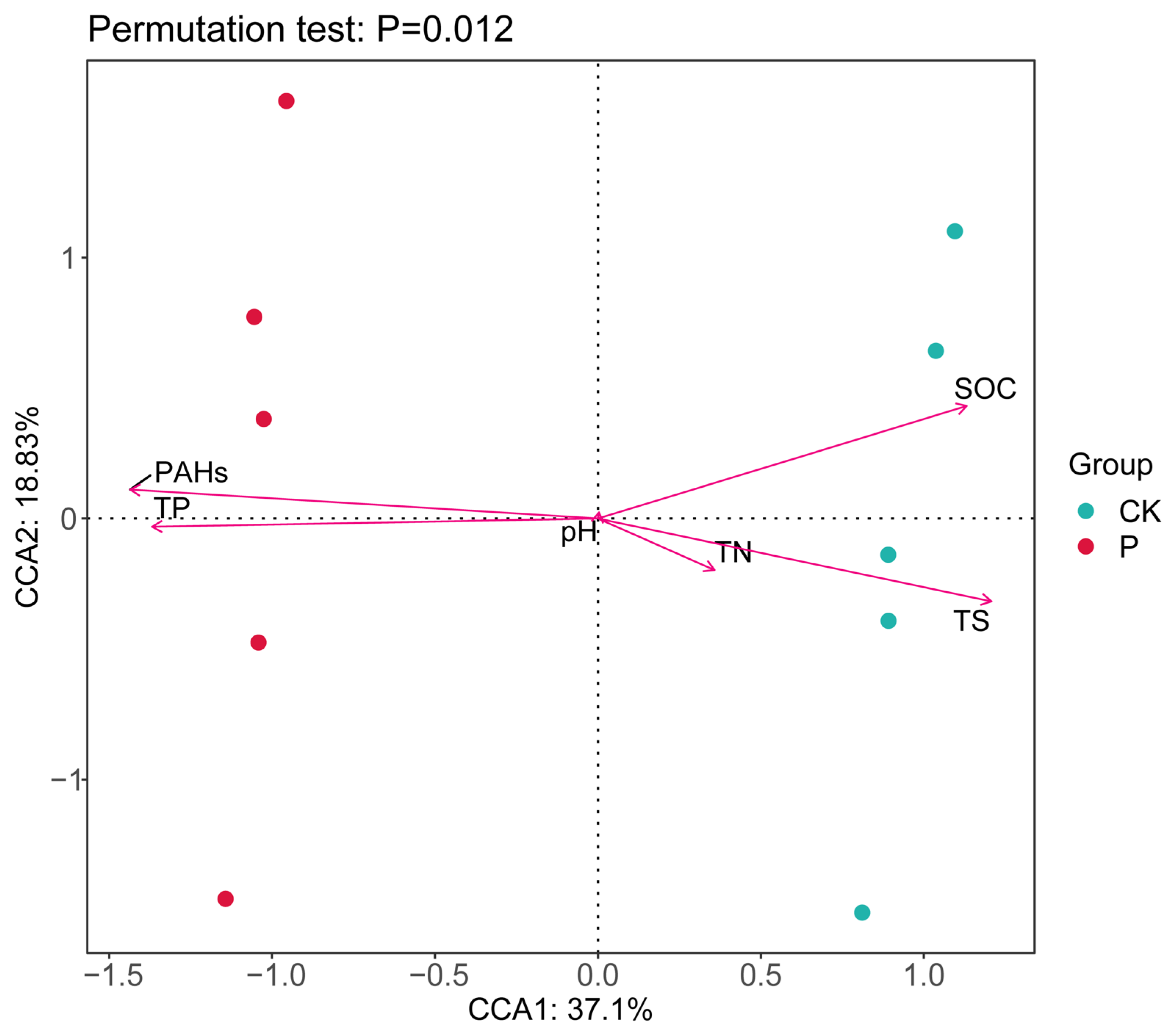
3.4. Relationship between Soil Bacterial Community Composition and Soil Physicochemical Properties in Two Wetlands in the Yellow River Delta, China
4. Discussion
4.1. Effects of PAH Pollution on Soil Bacterial Diversity
4.2. Effects of PAH Pollution on the Composition of the Soil Bacterial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sahoo, B.M.; Ravi Kumar, B.V.; Banik, B.K.; Borah, P. Polyaromatic hydrocarbons (PAHs): Structures, synthesis and their biological profile. Curr. Org. Synth. 2020, 17, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.U.; Siyi, L.; Yousaf, B.; Abbas, Q.; Hameed, R.; Zheng, C.; Wong, M.H. Emission sources and full spectrum of health impacts of black carbon associated polycyclic aromatic hydrocarbons (PAHs) in urban environment: A review. Crit. Rev. Environ. Sci. Technol. 2021, 51, 857–896. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, Y.; Zhang, N.; Wang, Y.; Ren, Y. Pollution levels, characteristics, and sources of polycyclic aromatic hydrocarbons in atmospheric particulate matter across the Hu line in China. A review. Environ. Chem. Lett. 2021, 19, 3821–3836. [Google Scholar] [CrossRef]
- Cheng, Y.; Tang, Y.T.; Nathanail, C.P. Determination of the potential implementation impact of 2016 ministry of environmental protection generic assessment criteria for potentially contaminated sites in China. Environ. Geochem. Health 2018, 40, 967–985. [Google Scholar] [CrossRef] [PubMed]
- Ranc, B.; Faure, P.; Croze, V.; Simonnot, M.O. Selection of oxidant doses for in situ chemical oxidation of soils contaminated by polycyclic aromatic hydrocarbons (PAHs): A review. J. Hazard. Mater. 2016, 312, 280–297. [Google Scholar] [CrossRef]
- Ramesh, A.; Archibong, A.E.; Hood, D.B.; Guo, Z.; Loganathan, B.G. Global environmental distribution and human health effects of polycyclic aromatic hydrocarbons. Glob. Contam. Trends Persistent Org. Chem. 2011, 63, 97–126. [Google Scholar]
- Sun, K.; Song, Y.; He, F.; Jing, M.; Tang, J.; Liu, R. A review of human and animals exposure to polycyclic aromatic hydrocarbons: Health risk and adverse effects, photo-induced toxicity and regulating effect of microplastics. Sci. Total Environ. 2021, 773, 145403. [Google Scholar] [CrossRef]
- Zhang, Z.; Rengel, Z.; Meney, K. Polynuclear aromatic hydrocarbons (PAHs) differentially influence growth of various emergent wetland species. J. Hazard. Mater. 2010, 182, 689–695. [Google Scholar] [CrossRef]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef]
- Hao, D.C.; Li, X.J.; Xiao, P.G.; Wang, L.F. The utility of electrochemical systems in microbial degradation of polycyclic aromatic hydrocarbons: Discourse, diversity and design. Front. Microbiol. 2020, 11, 557400. [Google Scholar] [CrossRef]
- Peng, R.H.; Xiong, A.S.; Xue, Y.; Fu, X.Y.; Gao, F.; Zhao, W.; Yao, Q.H. Microbial biodegradation of polyaromatic hydrocarbons. FEMS Microbiol. Rev. 2008, 32, 927–955. [Google Scholar] [CrossRef] [PubMed]
- García, M.M.; de Llasera, M.P.G. A review on the enzymes and metabolites identified by mass spectrometry from bacteria and microalgae involved in the degradation of high molecular weight PAHs. Sci. Total Environ. 2021, 797, 149035. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Singh, R.; Arora, N.K. Plant growth-promoting microbes: Diverse roles in agriculture and environmental sustainability. In Probiotics Plant Health; Springer: Singapore, 2017; pp. 71–111. [Google Scholar]
- Jamir, E.; Kangabam, R.D.; Borah, K.; Tamuly, A.; Deka Boruah, H.P.; Silla, Y. Role of soil microbiome and enzyme activities in plant growth nutrition and ecological restoration of soil health. In Microbes and Enzymes in Soil Health and Bioremediation. Microorganisms for Sustainability; Springer: Singapore, 2019; pp. 99–132. [Google Scholar]
- Dubey, A.; Malla, M.A.; Khan, F.; Chowdhary, K.; Yadav, S.; Kumar, A.; Khan, M.L. Soil microbiome: A key player for conservation of soil health under changing climate. Biodivers. Conserv. 2019, 28, 2405–2429. [Google Scholar] [CrossRef]
- Shah, K.K.; Tripathi, S.; Tiwari, I.; Shrestha, J.; Modi, B.; Paudel, N.; Das, B.D. Role of soil microbes in sustainable crop production and soil health: A review. Agric. Sci. Technol. 2021, 13, 1313–8820. [Google Scholar] [CrossRef]
- Sun, G.; Du, Y.; Yin, J.; Jiang, Y.; Zhang, D.; Jiang, B.; Hu, J. Response of microbial communities to different organochlorine pesticides (OCPs) contamination levels in contaminated soils. Chemosphere 2019, 215, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qu, C.; Bian, Y.; Gu, C.; Jiang, X.; Song, Y. New insights into the responses of soil microorganisms to polycyclic aromatic hydrocarbon stress by combining enzyme activity and sequencing analysis with metabolomics. Environ. Pollut. 2019, 255, 113312. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Leblic, M.I.; Turmero, A.; Hernández, M.; Hernández, A.J.; Pastor, J.; Ball, A.S.; Arias, M.E. Influence of xenobiotic contaminants on landfill soil microbial activity and diversity. J. Environ. Manag. 2012, 95, S285–S290. [Google Scholar] [CrossRef]
- Qin, Z.; Zhao, Z.; Adam, A.; Li, Y.; Chen, D.; Mela, S.M.; Li, H. The dissipation and risk alleviation mechanism of PAHs and nitrogen in constructed wetlands: The role of submerged macrophytes and their biofilms-leaves. Environ. Int. 2019, 131, 104940. [Google Scholar] [CrossRef]
- Ivanova, A.A.; Sazonova, O.I.; Zvonarev, A.N.; Delegan, Y.A.; Streletskii, R.A.; Shishkina, L.A.; Vetrova, A.A. Genome Analysis and Physiology of Pseudomonas sp. Strain OVF7 Degrading Naphthalene and n-Dodecane. Microorganisms 2023, 11, 2058. [Google Scholar] [CrossRef]
- Frantsuzova, E.; Bogun, A.; Solomentsev, V.; Vetrova, A.; Streletskii, R.; Solyanikova, I.; Delegan, Y. Whole Genome Analysis and Assessment of the Metabolic Potential of Gordonia rubripertincta Strain 112, a Degrader of Aromatic and Aliphatic Compounds. Biology 2023, 12, 721. [Google Scholar] [CrossRef]
- Frantsuzova, E.; Solomentsev, V.; Vetrova, A.; Travkin, V.; Solyanikova, I.; Delegan, Y. Complete Genome Sequence of Gordonia polyisoprenivorans 135, a Promising Degrader of Aromatic Compounds. Microbiol. Resour. Announc. 2023, 12, e00058. [Google Scholar] [CrossRef] [PubMed]
- Emelyanova, E.V.; Souzina, N.E.; Polivtseva, V.N.; Reshetilov, A.N.; Solyanikova, I.P. Survival and biodegradation activity of Gordonia polyisoprenivorans 135: Basics of a biosensor receptor. Appl. Biochem. Microbiol. 2017, 53, 580–586. [Google Scholar] [CrossRef]
- Wang, D.; Ma, J.; Li, H.; Zhang, X. Concentration and potential ecological risk of PAHs in different layers of soil in the petroleum-contaminated areas of the Loess Plateau, China. Int. J. Environ. Res. Public Health 2018, 15, 1785. [Google Scholar] [CrossRef]
- Dai, Y.; Liu, R.; Chen, J.; Li, N. Bioremediation of HMW-PAHs-contaminated soils by rhizosphere microbial community of Fire Phoenix plants. Chem. Eng. J. 2022, 432, 134246. [Google Scholar] [CrossRef]
- Wu, Y.; Jiang, B.; Zou, Y.; Dong, H.; Wang, H.; Zou, H. Influence of bacterial community diversity, functionality, and soil factors on polycyclic aromatic hydrocarbons under various vegetation types in mangrove wetlands. Environ. Pollut. 2022, 308, 119622. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, D.; Jin, D.; Zhang, J.; Shan, Y.; Wan, M.; Jiao, W. Soil physiochemical properties and bacterial community changes under long-term polycyclic aromatic hydrocarbon stress in situ steel plant soils. Chemosphere 2023, 334, 138926. [Google Scholar] [CrossRef]
- Mali, M.; Dell’Anna, M.M.; Mastrorilli, P.; Damiani, L.; Piccinni, A.F. Assessment and source identification of pollution risk for touristic ports: Heavy metals and polycyclic aromatic hydrocarbons in sediments of 4 marinas of the Apulia region (Italy). Mar. Pollut. Bull. 2017, 114, 768–777. [Google Scholar] [CrossRef]
- Qi, Y.; Guan, X.; He, J.; Fu, G.; Cao, M.; Zhao, C.Y.; Zhu, J.F.; Li, J.S. Distribution and Ecological Risk Assessment of Polycyclic Aromatic Hydrocarbons in Tidal Flats in Yellow River Delta, China. Environ. Sci. Technol. 2020, 43, 229–236. [Google Scholar]
- Guo, J.; Feng, H.; Roberge, G.; Feng, L.; Pan, C.; McNie, P.; Yu, Y. The negative effect of Chinese fir (Cunninghamia lanceolata) monoculture plantations on soil physicochemical properties, microbial biomass, fungal communities, and enzymatic activities. For. Ecol. Manag. 2022, 519, 120297. [Google Scholar] [CrossRef]
- Liu, X.; Li, J.; Yu, L.; Pan, H.; Liu, H.; Liu, Y.; Xu, J. Simultaneous measurement of bacterial abundance and composition in response to biochar in soybean field soil using 16S rRNA gene sequencing. Land Degrad. Dev. 2018, 29, 2172–2182. [Google Scholar] [CrossRef]
- Lu, J.; Salzberg, S.L. Ultrafast and accurate 16S rRNA microbial community analysis using Kraken 2. Microbiome 2020, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R Core Team R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Bastida, F.; Jehmlich, N.; Lima, K.; Morris, B.E.L.; Richnow, H.H.; Hernández, T.; García, C. The ecological and physiological responses of the microbial community from a semiarid soil to hydrocarbon contamination and its bioremediation using compost amendment. J. Proteom. 2016, 135, 162–169. [Google Scholar] [CrossRef]
- Sazykina, M.A.; Minkina, T.M.; Konstantinova, E.Y.; Khmelevtsova, L.E.; Azhogina, T.N.; Antonenko, E.M.; Sazykin, I.S. Pollution impact on microbial communities composition in natural and anthropogenically modified soils of Southern Russia. Microbiol. Res. 2022, 254, 126913. [Google Scholar] [CrossRef] [PubMed]
- Cébron, A.; Norini, M.P.; Beguiristain, T.; Leyval, C. Real-Time PCR quantification of PAH-ring hydroxylating dioxygenase (PAH-RHDα) genes from Gram positive and Gram negative bacteria in soil and sediment samples. J. Microbiol. Methods 2008, 73, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, G.; Törneman, N.; De Lipthay, J.R.; Sørensen, S.J. Microbial diversity and PAH catabolic genes tracking spatial heterogeneity of PAH concentrations. Microb. Ecol. 2013, 65, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, T.; Prenzler, P.D.; Howitt, J.A.; Doran, G.S. Eucalyptus saponin-and sophorolipid-mediated desorption of polycyclic aromatic hydrocarbons from contaminated soil and sediment. Environ. Sci. Pollut. Res. 2023, 30, 21638–21653. [Google Scholar] [CrossRef]
- Andreoni, V.; Gianfreda, L. Bioremediation and monitoring of aromatic-polluted habitats. Appl. Microbiol. Biotechnol. 2007, 76, 287–308. [Google Scholar] [CrossRef]
- Deng, J.; Yin, Y.; Zhu, W.; Zhou, Y. Response of soil environment factors and microbial communities to phytoremediation with Robinia pseudoacacia in an open-cut magnesite mine. Land Degrad. Dev. 2020, 31, 2340–2355. [Google Scholar] [CrossRef]
- Liu, T.; Yang, L.; Hu, Z.; Xue, J.; Lu, Y.; Chen, X.; Liu, M. Biochar exerts negative effects on soil fauna across multiple trophic levels in a cultivated acidic soil. Biol. Fertil. Soils 2020, 56, 597–606. [Google Scholar] [CrossRef]
- Su, P.H.; Lv, B.Y.; Tomy, G.T.; Xu, J.X.; Tian, W.; Hou, C.Y.; Feng, D.L. Occurrences, composition profiles and source identifications of polycyclic aromatic hydrocarbons (PAHs), polybrominated diphenyl ethers (PBDEs) and polychlorinated biphenyls (PCBs) in ship ballast sediments. Chemosphere 2017, 168, 1422–1429. [Google Scholar] [CrossRef]
- Wolf, D.C.; Cryder, Z.; Khoury, R.; Carlan, C.; Gan, J. Bioremediation of PAH-contaminated shooting range soil using integrated approaches. Sci. Total Environ. 2020, 726, 138440. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAHs | Latitude | Longitude | Vegetation |
|---|---|---|---|
| No pollution, (<100 ng/g, >0 ng/g) | 38.12249292 | 118.8743 | Suaeda salsa |
| Pollution, (>200 ng/g, 300 ng/g) | 38.11868385 | 118.7122 | Suaeda salsa |
| Wetlands | PAHs | SOC (g/kg) | TN (g/kg) | TS (%) | TP (g/kg) | pH |
|---|---|---|---|---|---|---|
| CK | 46.2 ± 1.12 b | 14.6 ± 1.14 b | 0. 48 ± 0.01 a | 0.035 ± 0.002 a | 0.47 ± 0.03 a | 7.27 ± 0.2 a |
| P | 233.7 ± 4.26 a | 18.1 ± 1.12 a | 0. 49 ± 0.01 a | 0.045 ± 0.002 a | 0.26 ± 0.01 b | 7.31 ± 0.2 a |
| Treatments | Chao1 | Faith_pd | Sobs | Shannon |
|---|---|---|---|---|
| CK | 140 ± 46.9 a | 33 ± 9.9 a | 140 ± 46.9 a | 6 ± 0.5 a |
| P | 142 ± 65.2 a | 30 ± 13.6 a | 142 ± 65.2 a | 4 ± 2.1 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Wu, Y.; Zhi, Q.; Zhang, Z.; Zhao, Y.; Fu, G. Effects of Polycyclic Aromatic Hydrocarbons on the Composition of the Soil Bacterial Communities in the Tidal Flat Wetlands of the Yellow River Delta of China. Microorganisms 2024, 12, 141. https://doi.org/10.3390/microorganisms12010141
Qi Y, Wu Y, Zhi Q, Zhang Z, Zhao Y, Fu G. Effects of Polycyclic Aromatic Hydrocarbons on the Composition of the Soil Bacterial Communities in the Tidal Flat Wetlands of the Yellow River Delta of China. Microorganisms. 2024; 12(1):141. https://doi.org/10.3390/microorganisms12010141
Chicago/Turabian StyleQi, Yue, Yuxuan Wu, Qiuying Zhi, Zhe Zhang, Yilei Zhao, and Gang Fu. 2024. "Effects of Polycyclic Aromatic Hydrocarbons on the Composition of the Soil Bacterial Communities in the Tidal Flat Wetlands of the Yellow River Delta of China" Microorganisms 12, no. 1: 141. https://doi.org/10.3390/microorganisms12010141
APA StyleQi, Y., Wu, Y., Zhi, Q., Zhang, Z., Zhao, Y., & Fu, G. (2024). Effects of Polycyclic Aromatic Hydrocarbons on the Composition of the Soil Bacterial Communities in the Tidal Flat Wetlands of the Yellow River Delta of China. Microorganisms, 12(1), 141. https://doi.org/10.3390/microorganisms12010141