Comparison of Japanese Encephalitis Force of Infection in Pigs, Poultry and Dogs in Cambodian Villages

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Description of Samples and Seroprevalence
2.2. Multivariate Analysis
2.3. Estimation of the Force of Infection (FOI) for Flaviviruses and JEV in the Sampled Animals
2.4. Force of Infection per Body Surface Area (BSA)
2.5. Comparison of FOI for Poultry in 2016 and 2018
3. Discussion
4. Materials and Methods
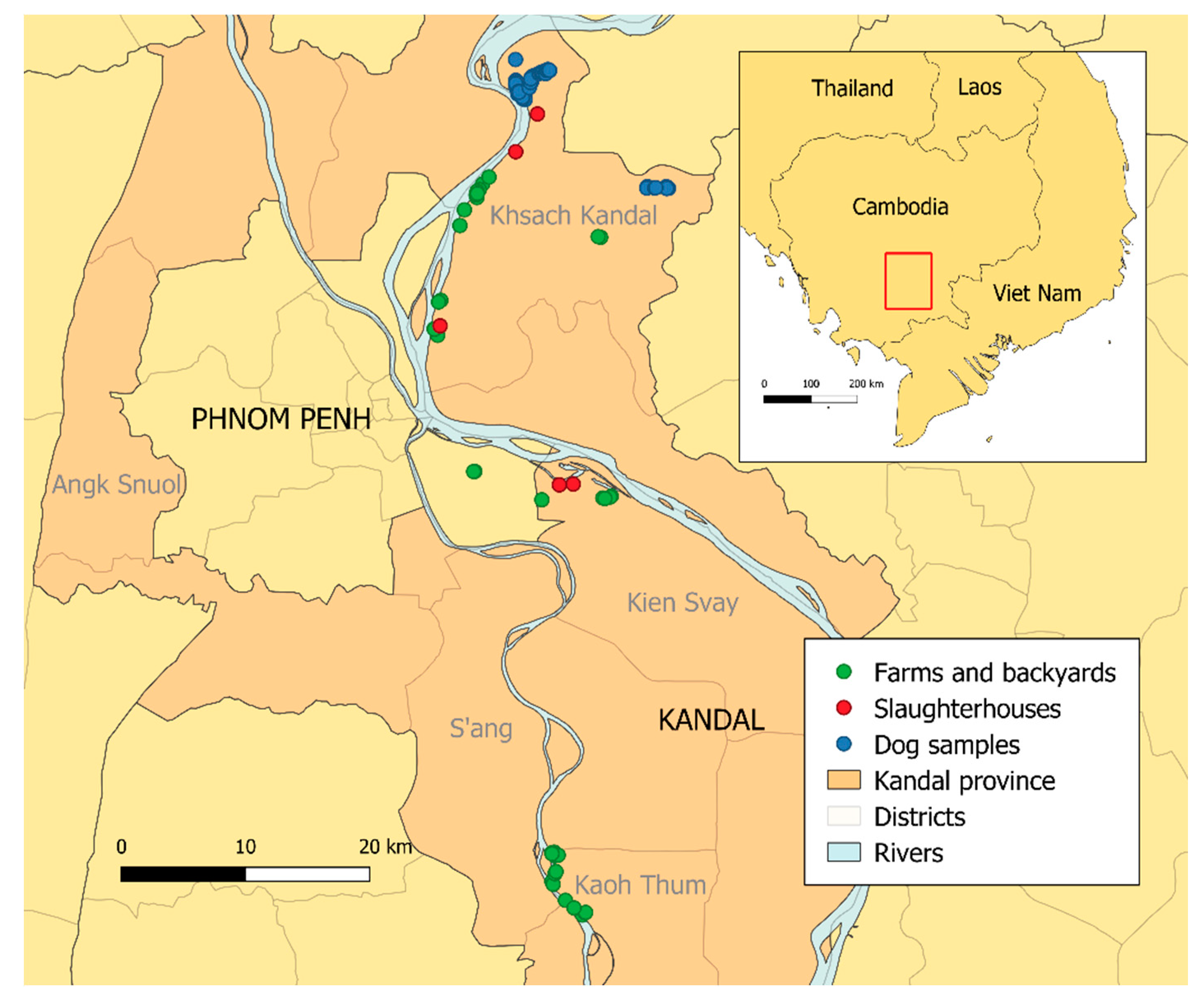
4.1. Study Area
4.2. Dog Samples
4.3. Samples of Domestic Pigs, Chickens, and Ducks
4.4. Ethic Statement
4.5. Laboratory analysis
4.5.1. Cells and Viruses
4.5.2. Hemagglutination Inhibition Assay (HIA)
4.5.3. Foci Reduction Neutralization Test (FRNT)
4.6. Statistical Analysis
4.7. Force of Infection per Unit of Body Surface Calculation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Erlanger, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, present, and future of Japanese encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tarantola, A.; Goutard, F.; Newton, P.; de Lamballerie, X.; Lortholary, O.; Cappelle, J.; Buchy, P. Estimating the Burden of Japanese Encephalitis Virus and Other Encephalitides in Countries of the Mekong Region. PLoS Negl. Trop. Dis. 2014, 8, e2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.; Hills, S.; Staples, E.; Johnson, B.; Yaich, M.; Solomon, T. Japanese Encephalitis Prevention and Control: Advances, Challenges, and New Initiatives. Emerg. Infect. 2008, 8, 93–124. [Google Scholar] [CrossRef]
- de Wispelaere, M.; Desprès, P.; Choumet, V. European Aedes albopictus and Culex pipiens Are Competent Vectors for Japanese Encephalitis Virus. PLoS Negl. Trop Dis. 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.S.; Al-Amin, H.M.; Chakma, S.; Alam, M.S.; Gurley, E.S.; Pulliam, J.R.C. Sampling Design Influences the Observed Dominance of Culex tritaeniorhynchus: Considerations for Future Studies of Japanese Encephalitis Virus Transmission. PLoS Negl. Trop Dis. 2016, 10, e0004249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, W.F.; Buescher, E.L.; McClure, H.E. Ecologic Studies of Japanese Encephalitis Virus in Japan. Am. J. Trop. Med. Hyg. 1959, 8, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Ohno, Y.; Mochizuki, M.; Iwata, H.; Okuda, M.; Maeda, K. Dogs as Sentinels for Human Infection with Japanese Encephalitis Virus. Emerg. Infect Dis. 2010, 16, 1137–1139. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Tamaru, S.; Morimoto, M.; Hayashi, T.; Shimojima, M.; Maeda, K. Experimental infection of Japanese encephalitis virus in dogs. J. Vet. Med. Sci. 2011, 73, 1241–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, G.; Lim, X.F.; Chan, S.; How, C.B.; Humaidi, M.; Yeo, G.; Mailepessov, D.; Kong, M.; Lai, Y.L.; Okumura, C.; et al. Serological evidence of continued Japanese encephalitis virus transmission in Singapore nearly three decades after end of pig farming. Parasites Vectors 2019, 12, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, W.; Kim, J.H.; Kim, J.; Lee, J.; Hwang, E.S. Changes of epidemiological characteristics of Japanese encephalitis viral infection and birds as a potential viral transmitter in Korea. J. Korean Med. Sci. 2018, 33. [Google Scholar] [CrossRef] [PubMed]
- Cleton, N.B.; Bosco-Lauth, A.; Page, M.J.; Bowen, R.A. Age-Related Susceptibility to Japanese Encephalitis Virus in Domestic Ducklings and Chicks. Am. J. Trop. Med. Hyg. 2014, 90, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Auerswald, H.; Ruget, A.S.; Ladreyt, H.; In, S.; Mao, S.; Sorn, S.; Tum, S.; Duong, V.; Dussart, P.; Cappelle, J.; et al. Serological evidence for Japanese encephalitis and West Nile virus infections in domestic birds in Cambodia. Front. Vet. Sci. 2020, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.H.; Laurenson, M.K. Identifying reservoirs of infection: A conceptual and practical challenge. Emerg. Infect. Dis. 2002, 8, 1468–1473. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.G.; Heesterbeek, J.A.P. Characterizing reservoirs of infection and the maintenance of pathogens in ecosystems. J. R. Soc. Interface 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Touch, S.; Hills, S.; Sokhal, B.; Samnang, C.; Sovann, L.; Khieu, V.; Soeung, S.C.; Toda, K.; Robinson, J.; Grundy, J. Epidemiology and burden of disease from Japanese encephalitis in Cambodia: Results from two years of sentinel surveillance. Trop. Med. Int. Health 2009, 14, 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Horwood, P.F.; Duong, V.; Laurent, D.; Mey, C.; Sothy, H.; Santy, K.; Richner, B.; Heng, S.; Hem, S.; Cheval, J.; et al. Aetiology of acute meningoencephalitis in Cambodian children, 2010–2013. Emerg. Microbes Infect. 2017, 6, e35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duong, V.; Sorn, S.; Holl, D.; Rani, M.; Deubel, V.; Buchy, P. Evidence of Japanese encephalitis virus infections in swine populations in 8 provinces of Cambodia: Implications for national Japanese encephalitis vaccination policy. Acta Trop. 2011, 120, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Cappelle, J.; Duong, V.; Pring, L.; Kong, L.; Yakovleff, M.; Prasetyo, D.B.; Peng, B.; Choeung, R.; Duboz, R.; Ong, S.; et al. Intensive Circulation of Japanese Encephalitis Virus in Peri-urban Sentinel Pigs near Phnom Penh, Cambodia. PLoS Negl. Trop. Dis. 2016, 10, e0005149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Francesco, J.; Choeung, R.; Peng, B.; Pring, L.; Pang, S.; Duboz, R.; Ong, S.; Sorn, S.; Tarantola, A.; Fontenille, D.; et al. Comparison of the dynamics of Japanese encephalitis virus circulation in sentinel pigs between a rural and a peri-urban setting in Cambodia. PLoS Negl. Trop. Dis. 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Williams, D.T.; Smith, D.W. Japanese Encephalitis Virus: The Geographic Distribution, Incidence, and Spread of a Virus with a Propensity to Emerge in New Areas. Perspect. Med. Virol. 2006, 16, 201–268. [Google Scholar]
- Kalaiyarasu, S.; Mishra, N.; Khetan, R.K.; Singh, V.P. Serological evidence of widespread West Nile virus and Japanese encephalitis virus infection in native domestic ducks (Anas platyrhynchos var domesticus) in Kuttanad region, Kerala, India. Comp. Immunol. Microbiol. Infect. Dis. 2016, 48, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ting, S.H.L.; Tan, H.C.; Wong, W.K.; Ng, M.L.; Chan, S.H.; Ooi, E.E. Seroepidemiology of neutralizing antibodies to Japanese encephalitis virus in Singapore: Continued transmission despite abolishment of pig farming? Acta Trop. 2004, 92, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Adi, A.A.A.M.; Astawa, N.; Ayu Asri Damayanti, P.; Made Kardena, I.; Gusti Made Krisna Erawan, I.; Suardana, I.W.; Putra, I.A.; Matsumoto, Y. Seroepidemiological Evidence for the Presence of Japanese Encephalitis Virus Infection in Ducks, Chickens, and Pigs, Bali-Indonesia. Bali Med. J. 2016, 5, 189. [Google Scholar] [CrossRef]
- Pant, G.R. A serological survey of pigs, horses, and ducks in Nepal for evidence of infection with Japanese encephalitis virus. Ann. N. Y. Acad. Sci. 2006, 1081, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Ruget, A.-S.; Beck, C.; Gabassi, A.; Trevennec, K.; Lecollinet, S.; Chevalier, V.; Cappelle, J. Japanese encephalitis circulation pattern in swine of northern Vietnam and consequences for swine’s vaccination recommendations. Transbound. Emerg. Dis. 2018, 65, 1485–1492. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.-X.; Li, C.-X.; Wang, G.; Zheng, Z.; Dong, Y.-D.; Zhang, Y.-M.; Xing, D.; Zhao, T.-Y. Host feeding patterns of mosquitoes in a rural malaria-endemic region in hainan island, china. J. Am. Mosq. Control Assoc. 2014, 30, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Fall, A.G.; Diaïté, A.; Lancelot, R.; Tran, A.; Soti, V.; Etter, E.; Konaté, L.; Faye, O.; Bouyer, J. Feeding behaviour of potential vectors of West Nile virus in Senegal. Parasit Vectors 2011, 4, 99. [Google Scholar] [CrossRef] [PubMed]
- Philip Samuel, P.; Arunachalam, N.; Hiriyan, J.; Tyagi, B.K. Host feeding pattern of Japanese encephalitis virus vector mosquitoes (Diptera: Culicidae) from Kuttanadu, Kerala, India. J. Med. Entomol. 2008, 45, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Reuben, R.; Thenmozhi, V.; Samuel, P.P.; Gajanana, A.; Mani, T.R. Mosquito blood feeding patterns as a factor in the epidemiology of Japanese encephalitis in southern India. Am. J. Trop. Med. Hyg. 1992, 46, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Karna, A.K.; Bowen, R.A. Experimental Evaluation of the Role of Ecologically-Relevant Hosts and Vectors in Japanese Encephalitis Virus Genotype Displacement. Viruses 2019, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocheleau, J.P.; Michel, P.; Lindsay, L.R.; Drebot, M.; Dibernardo, A.; Ogden, N.H.; Fortin, A.; Arsenault, J. Characterizing environmental risk factors for West Nile virus in Quebec, Canada, using clinical data in humans and serology in pet dogs. Epidemiol. Infect. 2017, 145, 2797–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, B.; Haskouri, H.; Lowenski, S.; Vachiery, N.; Beck, C.; Lecollinet, S. Seroprevalence of West Nile and Usutu viruses in military working horses and dogs, Morocco, 2012: Dog as an alternative WNV sentinel species? Epidemiol. Infect. 2016, 144, 1857–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, H.A.; van den Bogaard, A.E.; Nohlmans, M.K. Dogs as sentinels for human Lyme borreliosis in The Netherlands. J. Clin. Microbiol. 2001, 39, 844–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henn, J.B.; Gabriel, M.W.; Kasten, R.W.; Brown, R.N.; Theis, J.H.; Foley, J.E.; Chomel, B.B. Gray foxes (Urocyon cinereoargenteus) as a potential reservoir of a Bartonella clarridgeiae-like bacterium and domestic dogs as part of a sentinel system for surveillance of zoonotic arthropod-borne pathogens in northern California. J. Clin. Microbiol. 2007, 45, 2411–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lord, J.S.; Gurley, E.S.; Pulliam, J.R.C. Rethinking Japanese Encephalitis Virus Transmission: A Framework for Implicating Host and Vector Species. PLoS Negl. Trop. Dis. 2015, 9, e0004074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, T.T.T.; Aarnink, A.J.A.; Drucker, A.; Verstegen, M.W.A. Pig Production in Cambodia, Laos, Philippines, and Vietnam: A Review. Asian J. Agric. Dev. 2006, 3, 69–90. [Google Scholar] [CrossRef]
- FAO (2004) Review of the livestock Sector in the Mekong Countries. Available online: http://www.fao.org/docs/eims/upload/153492/lsr_mekong.pdf (accessed on 1 September 2020).
- Draaf Bretagne (2013) L’élevage en Bretagne, hier, aujourd’hui… et demain? Available online: http://draaf.bretagne.agriculture.gouv.fr/IMG/pdf/L_elevage_en_Bretagne_cle49c7b6.pdf (accessed on 1 September 2020).
- Nemeth, N.M.; Bowen, R.A. Dynamics of passive immunity to West Nile virus in domestic chickens (Gallus gallus domesticus). Am. J. Trop. Med. Hyg. 2007, 76, 310–317. [Google Scholar] [CrossRef] [PubMed]
- OIE. Use of Animals in Research and Education. In OIE Terrestrial Animal Health Code; OIR: Paris, France, 2018; Available online: http://www.oie.int/index.php?id=169&L=0&htmfile=chapitre_aw_research_education.htm (accessed on 1 September 2020).
- Fox-Lewis, A.; Hopkins, J.; Sar, P.; Sao, S.; Pheaktra, N.; Day, N.P.J.; Blacksell, S.D.; Turner, P. Seroprevalence of Dengue Virus and Rickettsial Infections in Cambodian Children. Am. J. Trop. Med. Hyg. 2019, 100, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Ly, S.; Fortas, C.; Duong, V.; Benmarhnia, T.; Sakuntabhai, A.; Paul, R.; Huy, R.; Sorn, S.; Nguon, K.; Chan, S.; et al. Asymptomatic Dengue Virus Infections, Cambodia, 2012–2013. Emerg. Infect. Dis. 2019, 25, 1354–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeki, T.; Tajima, S.; Ikeda, M.; Kato, F.; Taniguchi, S.; Nakayama, E.; Takasaki, T.; Lim, C.-K.; Saijo, M. Analysis of cross-reactivity between flaviviruses with sera of patients with Japanese encephalitis showed the importance of neutralization tests for the diagnosis of Japanese encephalitis. J. Infect. Chemother. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, D.H.; Casals, J. Techniques for hemagglutination and hemagglutination-inhibition with arthropod-borne viruses. Am. J. Trop. Med. Hyg. 1958, 7, 561–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auerswald, H.; de Jesus, A.; Seixas, G.; Nazareth, T.; In, S.; Mao, S.; Duong, V.; Silva, A.C.; Paul, R.; Dussart, P.; et al. First dengue virus seroprevalence study on Madeira Island after the 2012 outbreak indicates unreported dengue circulation. Parasit Vectors 2019, 12, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2005; ISBN 3-900051-07-0. Available online: http://www.R-project.org. (accessed on 1 September 2020).
- Lindstedt, S.L. Allometry: Body Size Constraints in Animal Design; National Academies Press (US): Washington, DC, USA, 1987. [Google Scholar]


{kind=link}
| HIA (Pos/Tested) | FRNT (Pos/Tested) | Seroprevalence Rates | |||||
|---|---|---|---|---|---|---|---|
| JEV a | DENV b | JEV and DENV c | Flavivirus (HIA) d | JEV (FRNT) e | |||
| Species | |||||||
| Pigs | 35/112 | 34/35 | 0/35 | 1/35 | 0.31 | 0.31 | |
| Chickens | 4/220 | 0/4 | 2/4 | 2/4 | 0.02 | 0.01 | |
| Ducks | 19/128 | 2/19 | 2/19 | 13/19 | 0.15 | 0.12 | |
| Dogs | 76/188 | 63/76 | 4/76 | 2/76 | 0.40 | 0.35 | |
| Districts of origin | |||||||
| Khsach Kandal | 112/335 | 84/112 | 6/112 | 15/112 | 0.33 | 0.30 | |
| Kean Svay | 18/110 | 14/18 | 2/18 | 2/18 | 0.16 | 0.15 | |
| Koh Thum | 4/168 | 1/4 | 0/4 | 1/4 | 0.02 | 0.01 | |
| Angk Snuol | 0/30 | nd | nd | nd | 0 | nd | |
| S’ang | 0/5 | nd | nd | nd | 0 | nd | |
| Age groups * | |||||||
| Pigs | 2–4 months | 9/58 | 9/9 | 0/9 | 0/9 | 0.16 | 0.16 |
| >4–5 months | 12/28 | 11/12 | 0/12 | 1/12 | 0.43 | 0.43 | |
| >5–6 months | 14/26 | 14/14 | 0/14 | 0/14 | 0.54 | 0.54 | |
| Chickens | 1–2 months | 0/129 | nd | nd | nd | 0 | nd |
| >2–3 months | 3/59 | 0/3 | 2/3 | 1/3 | 0.05 | 0.02 | |
| >3–24 months | 1/32 | 0/1 | 0/1 | 1/1 | 0.03 | 0.03 | |
| Ducks | 1–2 months | 2/65 | 1/2 | 0/2 | 0/2 | 0.03 | 0.02 |
| >2–5.5 months | 6/40 | 1/6 | 0/6 | 4/6 | 0.15 | 0.13 | |
| >5.5–15 months | 11/23 | 0/11 | 2/11 | 9/11 | 0.48 | 0.39 | |
| Dogs | 1.5–24 months | 30/103 | 21/30 | 3/30 | 1/30 | 0.29 | 0.21 |
| >24–48 months | 26/45 | 25/26 | 0/26 | 0/26 | 0.58 | 0.56 | |
| >48–108 months | 20/40 | 17/20 | 1/20 | 1/20 | 0.50 | 0.45 | |
| Flavivirus Seroprevalence (HIA) | JEV Seroprevalence (FRNT) | ||||
|---|---|---|---|---|---|
| Variable | Value | OR (95% CI) | p-Value | OR (95% CI) | p-Value |
| Species | Pigs | ref. | - | ref. | - |
| Chickens | 0.07 (0.02–0.19) | <0.0001 | 0.03 (0.01–0.12) | <0.0001 | |
| Ducks | 0.69 (0.31–1.49) | 0.34 | 0.50 (0.22–1.11) | 0.09 | |
| Dogs | 0.62 (0.31–1.25) | 0.18 | 0.38 (0.18–0.79) | 0.009 | |
| Age | quantitative | 1.12 * (1.04–1.2) | 0.003 | 1.14 * (1.06–1.23) | 0.0004 |
| Flavivirus FOI (HIA) | JEV FOI (FRNT) | |||
|---|---|---|---|---|
| FOI | 95% CI | FOI | 95% CI | |
| Pigs | 0.09 | (0.07–0.11) | 0.09 | (0.07–0.11) |
| Ducks | 0.04 | (0.03–0.06) | 0.03 | (0.02–0.04) |
| Dogs | 0.02 | (0.01–0.02) | 0.01 | (0.01–0.02) |
| Chickens | 0.007 | (0.003–0.014) | 0.003 | (0.001–0.009) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ladreyt, H.; Auerswald, H.; Tum, S.; Ken, S.; Heng, L.; In, S.; Lay, S.; Top, C.; Ly, S.; Duong, V.; et al. Comparison of Japanese Encephalitis Force of Infection in Pigs, Poultry and Dogs in Cambodian Villages. Pathogens 2020, 9, 719. https://doi.org/10.3390/pathogens9090719
Ladreyt H, Auerswald H, Tum S, Ken S, Heng L, In S, Lay S, Top C, Ly S, Duong V, et al. Comparison of Japanese Encephalitis Force of Infection in Pigs, Poultry and Dogs in Cambodian Villages. Pathogens. 2020; 9(9):719. https://doi.org/10.3390/pathogens9090719
Chicago/Turabian StyleLadreyt, Héléna, Heidi Auerswald, Sothyra Tum, Sreymom Ken, Leangyi Heng, Saraden In, Sokchea Lay, Chakriyouth Top, Sowath Ly, Veasna Duong, and et al. 2020. "Comparison of Japanese Encephalitis Force of Infection in Pigs, Poultry and Dogs in Cambodian Villages" Pathogens 9, no. 9: 719. https://doi.org/10.3390/pathogens9090719