
Four New Species of the Genus Eoneureclipsis (Trichoptera: Psychomyiidae) from China Inferred from Morphology and DNA Barcodes †



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Morphological Study
2.3. Molecular Analysis
3. Results
3.1. Taxonomy
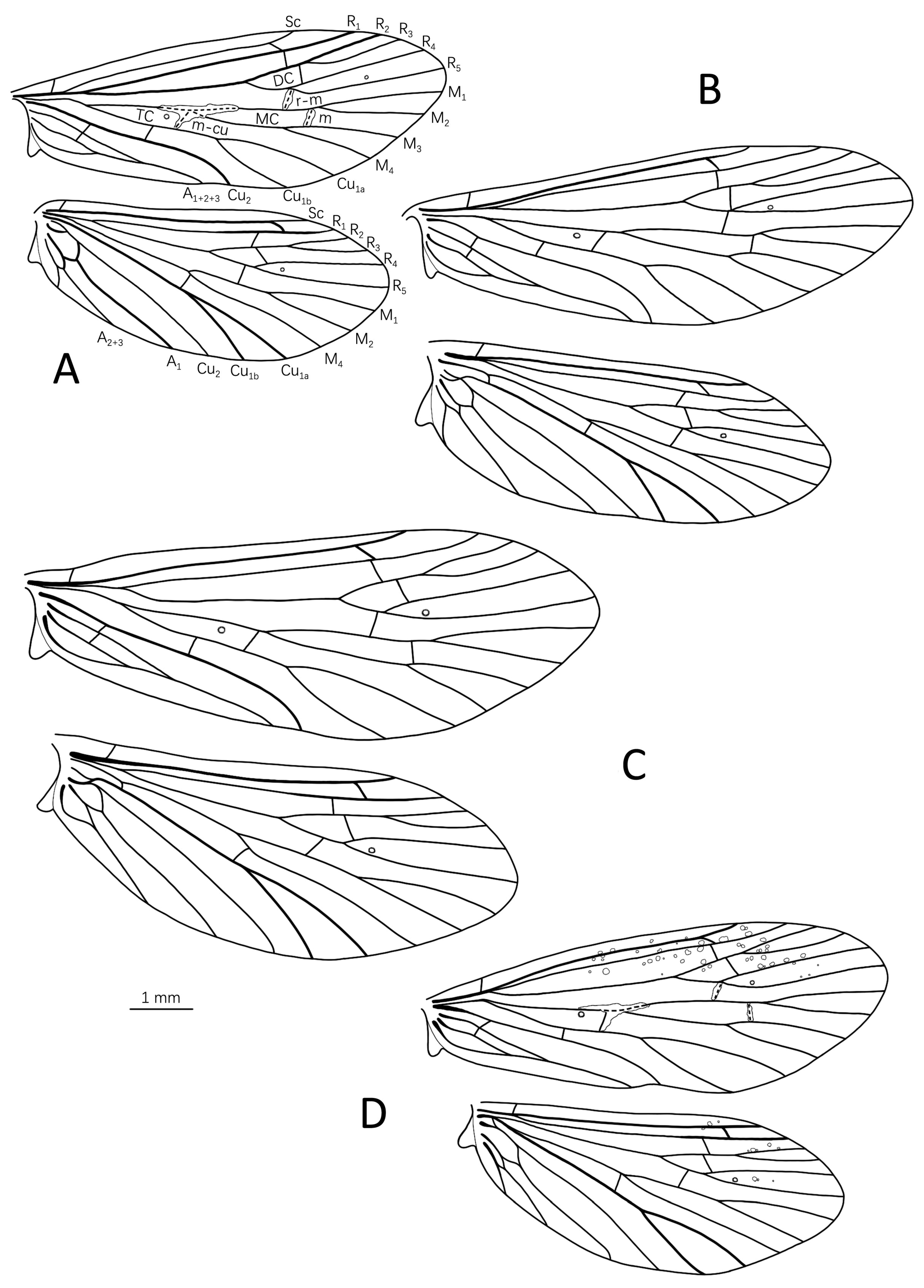
3.1.1. Eoneureclipsis jianfenglingensis Peng, Zang & Sun, sp. nov. (Figure 3A–E and Figure 7A)
3.1.2. Eoneureclipsis foraminulatus Peng, Zang & Sun, sp. nov. (Figure 4A–C and Figure 7B)
3.1.3. Eoneureclipsis spinosus Peng, Zang & Sun, sp. nov. (Figure 5A–F and Figure 7C)
3.1.4. Eoneureclipsis gei Peng, Zang & Sun, sp. nov. (Figure 6A–D and Figure 7D)
3.2. Key to Male Chinese Eoneureclipsis Species
- 1
- -
- Preanal appendages acute or narrow in lateral view ……………………………………4
- 2
- Intermediate appendages with stout spines (Figure 5B,C,E) ……E. spinosus sp. nov.
- -
- 3
- Intermediate appendages longer than phallic apparatus, slender, each curved upward subapically in lateral view [16] ……………………………………………E. sarach
- -
- Intermediate appendages of the same length as phallic apparatus, each broad subapically then abruptly tapered to acute apex in lateral view (Figure 6C) …E. gei sp. nov.
- 4
- -
- Phallic apparatus with ventral plate; intermediate appendages simple ………………5
- 5
- Phallic apparatus with dorsal process basally in lateral view [6].…………E. malchidael
- -
- 6
- Tergite IX posterior margin strongly expanded, with several long ossifying spines on each side in dorsal view (Figure 4B) …………………………….E. foraminulatus sp. nov.
- -
- Tergite IX tapering to end, without spines in dorsal view [15] …………E. hainanensis
3.3. Molecular Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kimmins, D.E. Results of the Oxford University expedition to Sarawak, 1932. Order Trichoptera. Sarawak Mus. J. 1955, 6, 374–442. [Google Scholar]
- Walker, F. Cataloque of the Specimens of Neuropterous Insects in the Collection of the British Museum, Part I: Phryganides-Perlides; British Museum: London, UK, 1852; pp. 1–192. [Google Scholar]
- Holzenthal, R.W.; Blahnik, R.J.; Prather, A.L.; Kjer, K.M. Order Trichoptera Kirby, 1813 (Insecta), Caddisflies. Zootaxa 2007, 1668, 639–698. [Google Scholar] [CrossRef]
- Morse, J.C. Trichoptera World Checklist. Available online: http://entweb.clemson.edu/database/trichopt/index.htm (accessed on 4 November 2022).
- Suwannarat, N.; Malicky, H.; Laudee, P. Four new species of caddisflies (Trichoptera: Polycentropodidae, Psychomyiidae, Hydropsychidae, Odontoceridae) from Khao Nan and Tai Rom Yen National Parks, southern Thailand. Zootaxa 2020, 4801, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Malicky, H. Beiträge zur Kenntnis chinesischer Köcherfliegen (Insecta, Trichoptera). Linz. Biol. Beiträge 2020, 52, 417–455. [Google Scholar]
- Torii, T.; Nishimoto, H. Discovery of the genus Eoneureclipsis Kimmins (Trichoptera: Psychomyiidae) from Japan. Zoosymposia 2011, 5, 453–464. [Google Scholar] [CrossRef]
- Malicky, H. Beschreibung von neuen Kocherfliegen (Trichoptera) aus Thailand und Burma. Entomol. Ber. Luzern 1989, 22, 117–126. [Google Scholar]
- Malicky, H.; Chantaramongkol, P. Weitere neue Köcherfliegen (Trichoptera) aus Thailand. Arbeit Nr. 20 über thailändische Köcherfliegen. Linz. Biol. Beiträge 1997, 29, 203–215. [Google Scholar]
- Malicky, H.; Chantaramongkol, P.; Bunlue, P.; Changthong, N.; Nawvong, J.; Nuntakwang, A.; Prommi, T.; Thamsenanupap, P.; Thapanya, D. 27 neue Köcherfliegen aus Thailand (Insecta, Trichoptera) (36. Arbeit über thailändische Köcherfliegen). Linz. Biol. Beiträge 2004, 36, 287–304. [Google Scholar]
- Thapanya, D.; Chantaramongkol, P.; Malicky, H. An updated survey of caddisflies (Trichoptera, Insecta) from Doi Suthep-Pui and Doi Inthanon National Parks, Chiang Mai Province, Thailand. Trop. Nat. Hist. 2004, 1, 21–40. [Google Scholar]
- Malicky, H. Neue Kocherfliegen (Trichoptera, Insecta) aus Vietnam. Linz. Biol. Beiträge 1995, 27, 851–885. [Google Scholar]
- Arefina-Armitage, T.I.; Armitage, B.J. New species and new country records for Vietnamese caddisflies (Insecta: Trichoptera). Insecta Mundi 2015, 0438, 1–19. [Google Scholar]
- Armitage, B.J.; Mey, W.; Arefina, T.I.; Schefter, P.W. The caddisfly fauna (Insecta: Trichoptera) of Vietnam. In Proceedings of the 11th International Symposium on Trichoptera, Shiga, Japan, 12–19 June 2003; Tanida, K., Rossiter, A., Eds.; Tokai University Press: Kanagawa, Japan, 2005; pp. 25–37. [Google Scholar]
- Mey, W. Eoneureclipsis hainanensis spec. nov. from China and its systematic position (Insecta, Trichoptera, Psychomyiidae) (pl. 14). Esperiana Band 2013, 18, 255–258. [Google Scholar]
- Malicky, H. Köcherfliegen (Trichoptera) von Taiwan, mit Neubeschreibungen. Linz. Biol. Beiträge 2014, 46, 1607–1646. [Google Scholar]
- Schmid, F. Sur quelques nouvelles psychomyiines tropicales (Trichoptera: Psychomyiidae). Le Nat. Can. 1972, 99, 143–172. [Google Scholar]
- Malicky, H. Beiträge zur Kenntnis asiatischer Trichopteren. Braueria 2009, 36, 11–58. [Google Scholar]
- Thamsenanupap, P.; Chantaramongkol, P.; Malicky, H. Description of caddis larvae (Trichoptera) from northern Thailand of the genera Himalopsyche (Rhyacophilidae), Arctopsyche (Arctopsychidae), cf. Eoneureclipsis (Psychomyiidae) and Inthanopsyche (Odontoceridae). Braueria 2005, 32, 7–11. [Google Scholar]
- Torii, T.; Nakamura, M. DNA identification and morphological description of the larva of Eoneureclipsis montanus (Trichoptera, Psychomyiidae). Zoosymposia 2016, 10, 424–431. [Google Scholar] [CrossRef]
- Xu, J.H.; Wang, B.X.; Sun, C.H. The Stenopsyche simplex species group from China with descriptions of three new species (Trichoptera: Stenopsychidae). Zootaxa 2015, 3785, 217–230. [Google Scholar] [CrossRef]
- Zang, H.; Ge, X.; Peng, L.; Sun, C.; Wang, B. A New Species of Limnephilus (Insecta: Trichoptera: Limnephilidae) from China, with Revision of the Genus Limnephilus on the Chinese Mainland. Insects 2022, 13, 653. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome coxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Ge, X.Y.; Peng, L.; Du, J.; Sun, C.H.; Wang, B.X. New species of the genus Molanna Curtis, 1834 (Trichoptera, Molannidae) in China inferred from morphology and DNA barcodes. ZooKeys 2022, 1112, 161–178. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Morse, J.C. Phylogeny and classification of Psychomyiidae (Trichoptera) genera. In Proceedings of the 8th International Symposium on Trichoptera, Minneapolis and Lake Itasca, MI, USA, 9–15 July 1995; Holzenthal, R.W., Flint, O.S., Jr., Eds.; Ohio Biological Survey: Columbus, OH, USA, 1997; pp. 271–276. [Google Scholar]
- Sharma, R.M.; Chandra, K. Checklist of Indian caddisflies (Insecta: Trichoptera). Zool. Surv. India 2009, 1–33. Available online: https://www.researchgate.net/publication/237460449_CHECKLIST_OF_INDIAN_CADDISFLIES_INSECTA_TRICHOPTERA (accessed on 15 January 2023).
- Tachet, H.; Coppa, G.; Forcellini, M. A comparative description of the larvae of Psychomyia pusilla (Fabricius 1781), Metalype fragilis (Pictet 1834), and Paduniella vandeli Décamps 1965 (Trichoptera: Psychomyiidae) and comments on the larvae of other species belonging to these three genera. Zootaxa 2018, 4402, 91–112. [Google Scholar] [CrossRef]
- Silva, A.V.S.D. Phylogeny of Xiphocentronidae Ross, 1949 and Taxonomy of Neotropical Xiphocentroninae (Trichoptera: Annulipalpia: Psychomyioidea). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2020. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
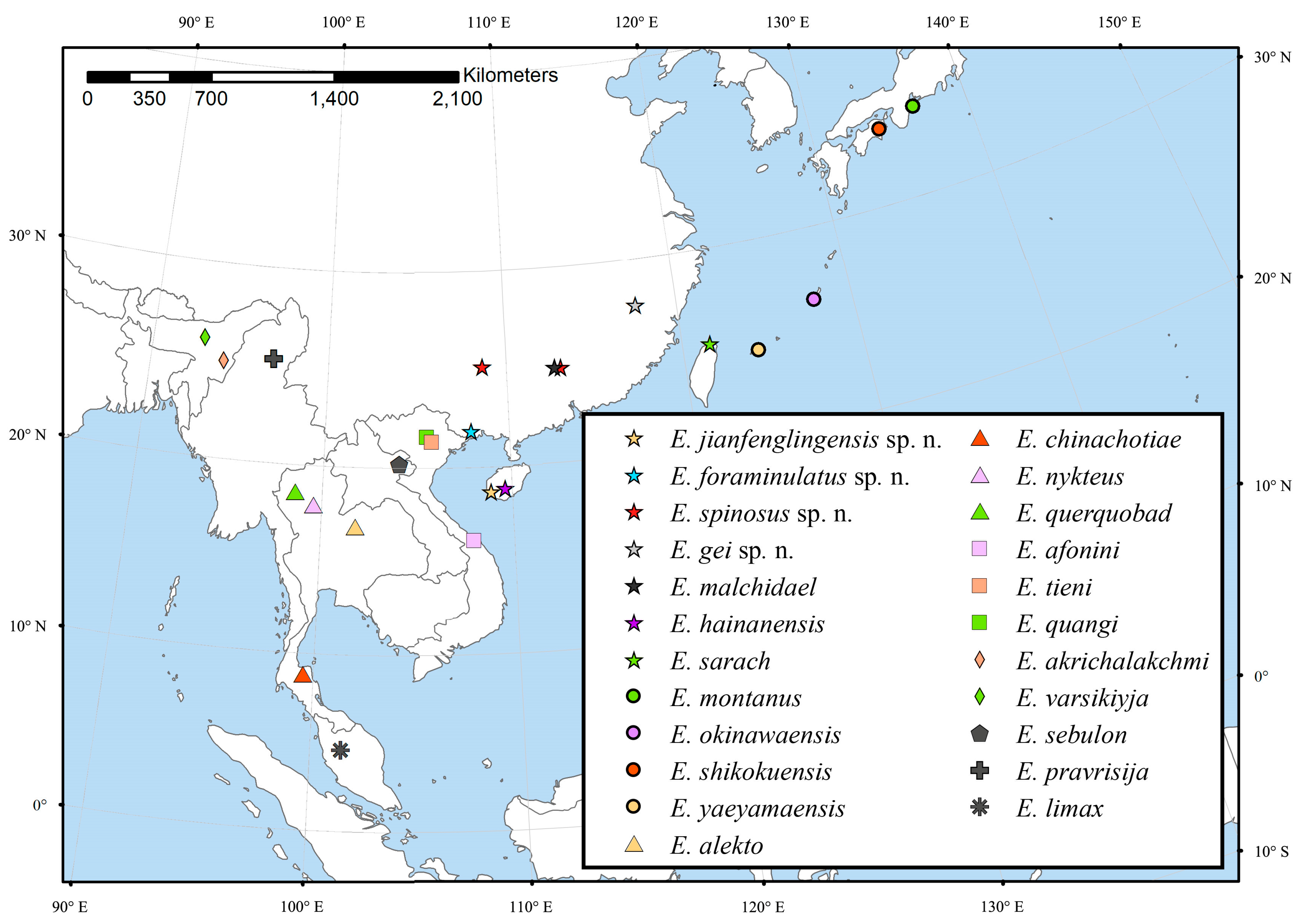
| Number | Species | Distribution |
|---|---|---|
| 1 | E. jianfenglingensis sp. nov. | China: Hainan |
| 2 | E. foraminulatus sp. nov. | China: Guangxi |
| 3 | E. spinosus sp. nov. | China: Guangxi, Guangdong |
| 4 | E. gei sp. nov. | China: Fujian |
| 5 | E. malchidael Malicky, 2020 | China: Guangdong |
| 6 | E. hainanensis Mey, 2013 | China: Hainan |
| 7 | E. sarach Malicky, 2014 | China: Taiwan |
| 8 | E. montanus Torii & Nishimoto, 2011 | Japan: Honshu |
| 9 | E. okinawaensis Torii & Nishimoto, 2011 | Japan: Okinawa |
| 10 | E. shikokuensis Torii & Nishimoto, 2011 | Japan: Shioku |
| 11 | E. yaeyamaensis Torii & Nishimoto, 2011 | Japan: Yaeyama Islands |
| 12 | E. alekto Malicky & Chantaramongkol, 1997 | Thailand |
| 13 | E. chinachotiae Malicky &Laudee, 2020 | Thailand |
| 14 | E. nykteus Malicky & Nawvong, 2004 | Thailand |
| 15 | E. querquobad Malicky & Chantaramongkol, 1989 | Thailand |
| 16 | E. afonini Arefina-Armitage & Armitage, 2015 | Vietnam |
| 17 | E. quangi Malicky, 1995 | Vietnam |
| 18 | E. tieni Malicky, 1995 | Vietnam |
| 19 | E. akrichalakchmi Schmid, 1972 | India: Manipur |
| 20 | E. varsikiyja Schmid, 1972 | India: Assam |
| 21 | E. sebulon Malicky, 2009 | Laos |
| 22 | E. pravrisija Schmid, 1972 | Myanmar |
| 23 | E. limax Kimmins, 1955 | Malaysia: Sarawak |
| Species | Number of Specimens | Collection Site | Collection Date | Molecular Sample |
|---|---|---|---|---|
| E. jianfenglingensis sp. nov. | 1 male | Jianfeng Town, Ledong County, Hainan | 16 Apr 2019 | yes |
| E. jianfenglingensis sp. nov. | 1 male | Jianfeng Town, Ledong County, Hainan | 27 Jul 2022 | no |
| E. foraminulatus sp. nov. | 1 male | Shangsi County, Fangchenggang City, Guangxi | 06 Jun 2004 | no |
| E. spinosus sp. nov. | 2 males | Huanjiang County, Hechi City, Guangxi | 15 Jun 2004 | no |
| E. spinosus sp. nov. | 1 male | Ruyuan County, Shaoguan City, Guangdong | 21–22 May 2004 | no |
| E. gei sp. nov. | 1 male | Xingcun Town, Wuyishan City, Fujian | 16 Jul 2022 | yes |
| E. hainanensis Mey, 2013 | 1 male | Jianfeng Town, Ledong County, Hainan | 28 Jul 2022 | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, L.; Zang, H.; Sun, C.; Wang, L.; Wang, B. Four New Species of the Genus Eoneureclipsis (Trichoptera: Psychomyiidae) from China Inferred from Morphology and DNA Barcodes. Insects 2023, 14, 158. https://doi.org/10.3390/insects14020158
Peng L, Zang H, Sun C, Wang L, Wang B. Four New Species of the Genus Eoneureclipsis (Trichoptera: Psychomyiidae) from China Inferred from Morphology and DNA Barcodes. Insects. 2023; 14(2):158. https://doi.org/10.3390/insects14020158
Chicago/Turabian StylePeng, Lang, Haoming Zang, Changhai Sun, Le Wang, and Beixin Wang. 2023. "Four New Species of the Genus Eoneureclipsis (Trichoptera: Psychomyiidae) from China Inferred from Morphology and DNA Barcodes" Insects 14, no. 2: 158. https://doi.org/10.3390/insects14020158
APA StylePeng, L., Zang, H., Sun, C., Wang, L., & Wang, B. (2023). Four New Species of the Genus Eoneureclipsis (Trichoptera: Psychomyiidae) from China Inferred from Morphology and DNA Barcodes. Insects, 14(2), 158. https://doi.org/10.3390/insects14020158