
Wetlands in the Pleistocene Steppe-Tundra Landscapes of Beringia, Their Insects, and the Role of Aeolian Sedimentation


Abstract

1. Introduction
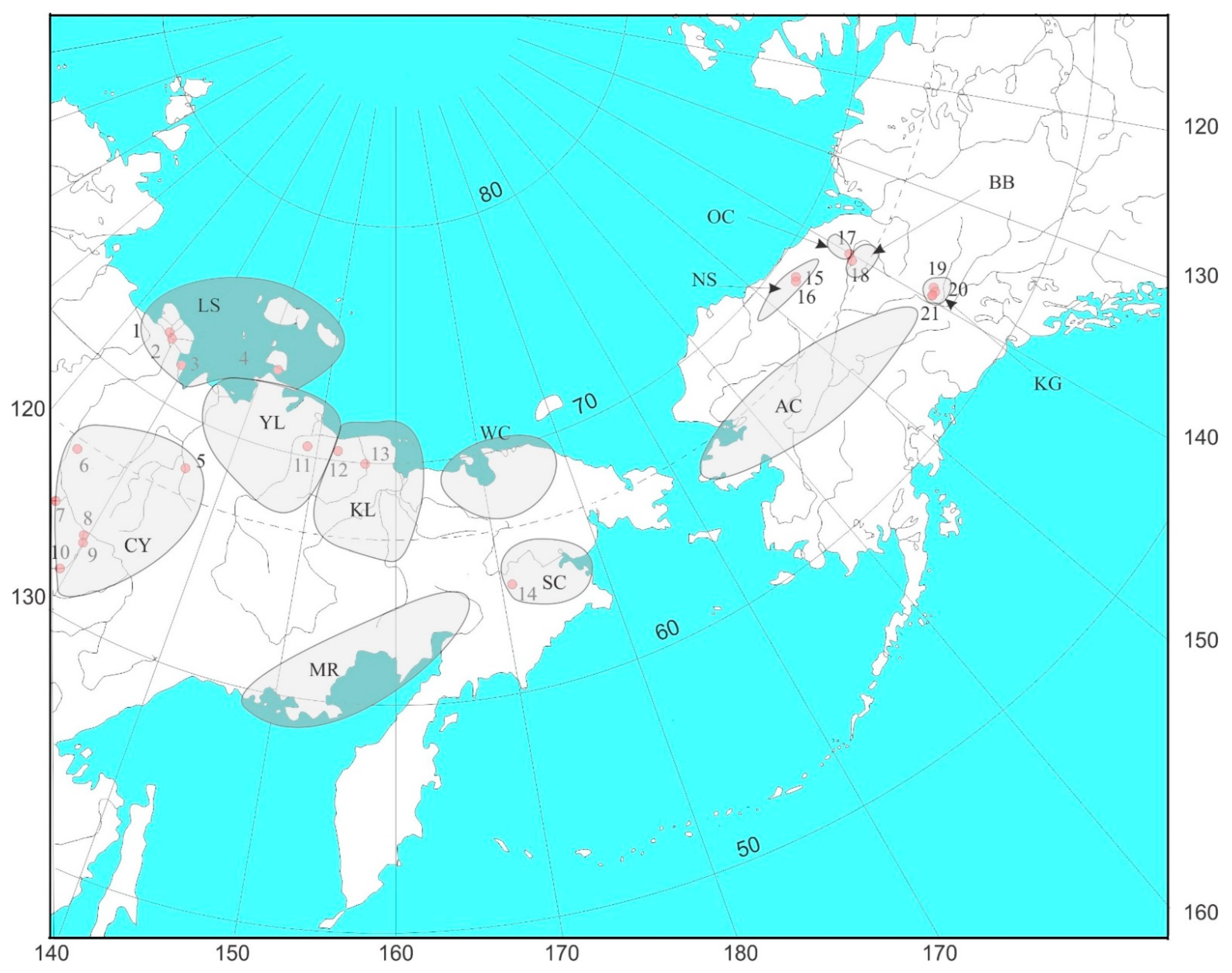
2. Materials and Methods
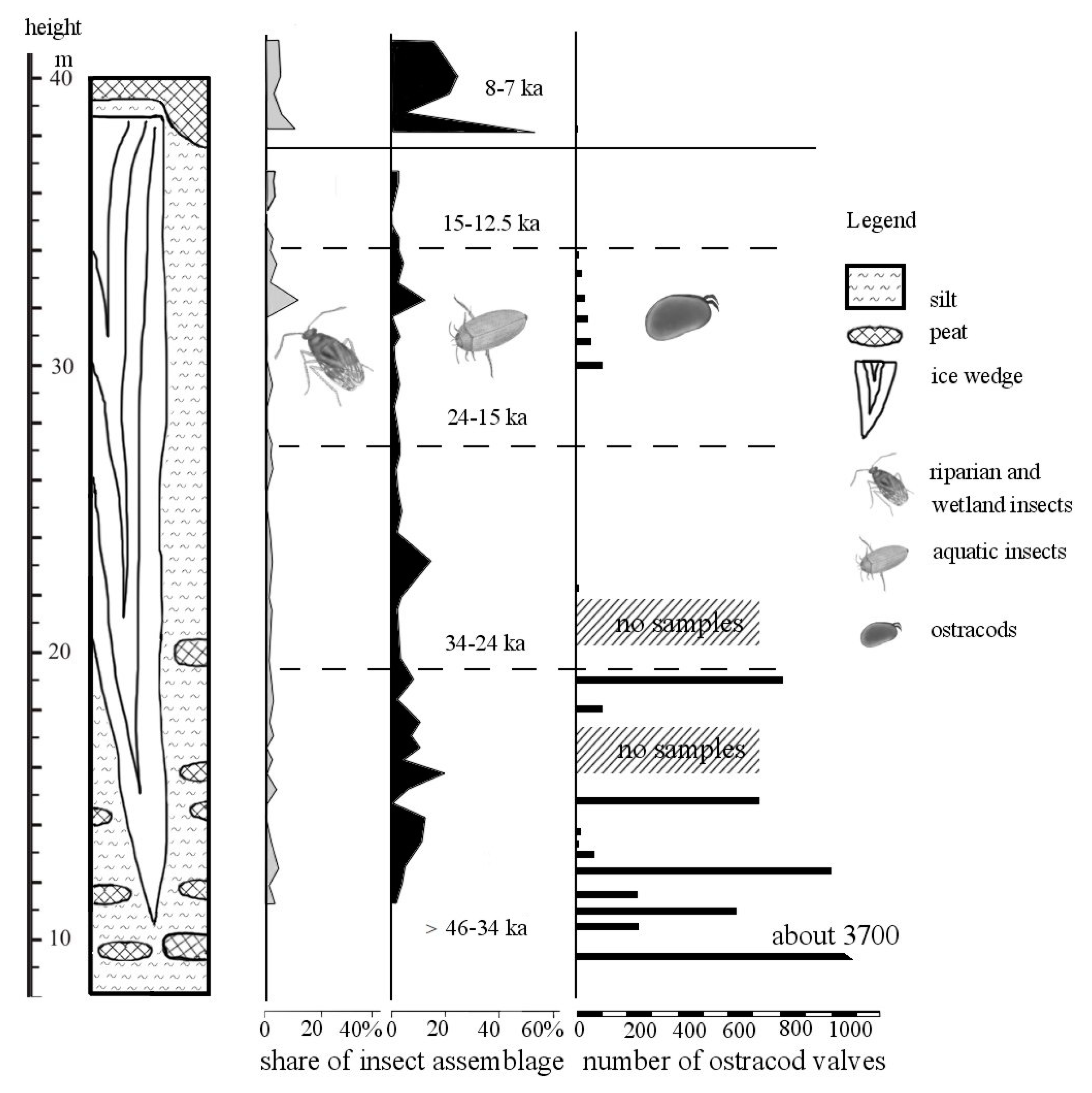
2.1. Lacustrine Deposits
2.2. Alluvium
2.3. Peatland Deposits
2.4. Wetland Deposits
2.5. Subaerial Deposits
3. Results
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zherikhin, V.V. Pattern of insect burial and conservation. In History of Insects; Rasnitsyn, A.P., Quicke, D.L.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 17–64. ISBN 978-0-306-47577-1. [Google Scholar]
- Coope, G.R. Constancy of insect species versus inconstancy of Quaternary environments. In Diversity of Insect Faunas; Mound, L.A., Waloff, N., Eds.; Blackwell Science: Oxford, UK, 1978; pp. 176–187. [Google Scholar]
- Abellán, P.; Benetti, C.J.; Angus, R.B.; Ribera, I. A review of Quaternary range shifts in European aquatic Coleoptera. Global Ecol. Biogeogr. 2011, 20, 87–100. [Google Scholar] [CrossRef]
- Fikáček, M.; Prokin, A.; Angus, R.B. A long-living species of the hydrophiloid beetles: Helophorus sibiricus from the early Miocene deposits of Kartashevo (Siberia, Russia). ZooKeys 2011, 130, 239–254. [Google Scholar] [CrossRef]
- Shur, Y.; Fortier, D.; Jorgenson, M.T.; Kanevskiy, M.; Schirrmeister, L.; Strauss, J.; Vasiliev, A.; Ward Jones, M. Yedoma Permafrost Genesis: Over 150 Years of Mystery and Controversy. Front. Earth Sci. 2022, 9, 757891. [Google Scholar] [CrossRef]
- Efremov, I.A. Taphonomy: A new branch of paleontology. Pan-Am. Geol. 1940, 74, 81–93. [Google Scholar]
- Martin, R.E. Taphonomy. A Process Approach; Cambridge Paleobiology Series; Cambridge University Press: Cambridge, UK, 1999; Volume 4. [Google Scholar]
- Kuzmina, S.A. Quaternary Insects and Environment of the Northeastern Asia. Ltd., Pleiades Publishing, Ltd. Paleontol. J. Suppl. 2015, 49, 1–189. [Google Scholar] [CrossRef]
- Kiselev, S.V.; Nazarov, V.I. Late Cenozoic Insects of Northern Eurasia. Pleiades Publishing, Ltd. Paleontol. J. Suppl. 2009, 43, 1–128. [Google Scholar] [CrossRef]
- Kuzmina, S.; Froese, D.G.; Jensen, B.J.L.; Hall, E.; Zazula, G.D. Middle Pleistocene (MIS 7) to Holocene fossil insect assemblages from the Old Crow basin, northern Yukon, Canada. Quatern. Intern. 2014, 341, 216–242. [Google Scholar] [CrossRef]
- Kuzmina, S.A.; Elias, S.A.; Kotov, A.A. Late Quaternary insects and freshwater invertebrates of the Alaskan North Slope and paleoenvironmental reconstructions in Arctic Alaska. Invert. Zool. 2019, 16, 89–125. [Google Scholar] [CrossRef]
- Andreev, A.A.; Grosse, G.; Schirrmeister, L.; Kuznetsova, T.V.; Kuzmina, S.A.; Bobrov, A.A.; Tarasov, P.E.; Novenko, E.Y.; Meyer, H.; Derevyagin, A.Y.; et al. Weichselian and Holocene palaeoenvironmental history of the Bol’shoy Lyakhovsky Island, New Siberian Archipelago, Arctic Siberia. Boreas 2009, 38, 72–110. [Google Scholar] [CrossRef]
- Sher, A.V.; Kuzmina, S.A.; Kuznetsova, T.V.; Sulerzhitsky, L.D. New insights into the Weichselian environment and climate of the Eastern-Siberian Artic, derived from fossil insects, plants, and mammals. Quatern. Sci. Rev. 2005, 24, 533–569. [Google Scholar] [CrossRef]
- Wetterich, S.; Kuzmina, S.; Andreev, A.A.; Kienast, F.; Meyer, H.; Schirrmeister, L.; Kuznetsova, T.; Sierralta, M. Palaeoenvironmental dynamics inferred from late Quaternary permafrost deposits on Kurungnakh Island, Lena Delta, Northeast Siberia, Russia. Quatern. Sci. Rev. 2008, 27, 1523–1540. [Google Scholar] [CrossRef]
- Kuzmina, S.A.; Wetterich, S.; Meyer, H. Paleoecological and sedimentological studies f Permafrost deposits in the Central Lena Delta (Kurungnakh and Samoylov Islands). Russian German Cooperation SYSTEM LAPTEV SEA The expedition 2002. Rep. Polar Res. 2003, 466, 71–81. [Google Scholar]
- Wooller, M.J.; Zazula, G.D.; Blinnikov, M.; Gaglioti, B.V.; Bigelow, N.H.; Sanborn, P.; Kuzmina, S.; La Farge, C. The detailed paleoecology of a mid-Wisconsinan interstadial (ca. 32 000 14C yr BP) vegetation surface from interior Alaska. Journ. Quatern. Sci. 2011, 26, 746–756. [Google Scholar] [CrossRef]
- Zanina, O.G.; Gubin, S.V.; Kuzmina, S.A.; Maximovich, S.V.; Lopatina, D.A. Late-Pleistocene (MIS 3-2) palaeoenvironments as recorded by sediments, palaeosols, and ground-squirrel nests at Duvanny Yar, Kolyma lowland, northeast Siberia. Quatern. Sci. Rev. 2011, 30, 2107–2123. [Google Scholar] [CrossRef]
- Kuzmina, S.; Elias, S.; Matheus, P.; Storer, J.E.; Sher, A. Paleoenvironmental reconstruction of the Last Glacial Maximum, inferred from insect fossils from a buried soil at Tempest Lake, Seward Peninsula, Alaska. Palaeogeogr. Palaeoclim. Palaeoecol. 2008, 267, 245–255. [Google Scholar] [CrossRef]
- Reyes, A.V.; Jensen, B.J.L.; Zazula, G.D.; Ager, T.A.; Kuzmina, S.; La Farge, C.; Froese, D.G. A Late–Middle Pleistocene (Marine Isotope Stage 6) vegetated surface buried by Old Crow tephra at the Palisades, interior Alaska. Quatern. Sci. Rev. 2010, 29, 801–811. [Google Scholar] [CrossRef]
- Zazula, G.D.; Froese, D.G.; Elias, S.; Kuzmina, S.; Farge, C.L.; Reyes, A.V.; Sanborn, P.T.; Schweger, C.E.; Scott Smith, C.A.; Mathewes, R.W. Vegetation buried under Dawson tephra (25,300 14C years BP) and locally diverse late Pleistocene paleoenvironments of Goldbottom Creek, Yukon, Canada. Palaeogeogr. Palaeoclim. Palaeoecol. 2006, 242, 253–286. [Google Scholar] [CrossRef]
- Kuzmina, S.A.; Basilyan, A.E. First finds of the fossil insects in the Quaternary aeolian deposit of the Central Yakutia and their role in the environmental reconstructions. Routes of evolutionary geography. Issue 2. In Proceedings of the 2nd Scientific Conference in Memory of Prof. A.A. Velichko, Moscow, Russia, 22–25 November 2021; pp. 170–173. (In Russian). [Google Scholar]
- Kiselev, S.V. Late Cenozoic Coleoptera of North-East Siberia; Nauka: Moscow, Russia, 1981. (In Russian) [Google Scholar]
- Sher, A.V.; Kuzmina, S. Beetle records/Late Pleistocene of Northern Asia. In Encyclopedia of Quaternary Science; Elias, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 246–267. [Google Scholar]
- Kuzmina, S.A. Macroentomology Analysis: Methods, Opportunities, and Examples of Reconstructions of Paleoclimatic and Paleoenvironmental Conditions in the Quaternary of the Northeastern Siberia. Contemporary Probl. Ecol. 2017, 10, 336–349, Pleiades Publishing, Ltd., 2017. Original Russian Text S.A. Kuzmina, 2017, published in Sibirskii Ekologicheskii Zhurnal, 2017, 4, 381–398. [Google Scholar] [CrossRef]
- Kuzmina, S. New approach to the Quaternary studies: QINSIB–The database of Siberian fossil insects. Quatern. Intern. 2014, 341, 283–293. [Google Scholar] [CrossRef]
- Guthrie, R.D. Origin and causes of the mammoth steppe: A story of cloud cover, woolly mammal tooth pits, buckles, and inside-out Beringia. Quat. Sci. Rev. 2001, 20, 549–574. [Google Scholar] [CrossRef]
- Sher, A.V. Pleistocene mammals and stratigraphy of the far northeast USSR and North America. Internal. Geol. Rev. 1974, 16, 1–284. [Google Scholar]
- Matthews, J.V., Jr.; Telka, A. Insect Fossils from the Yukon. In Insects of the Yukon; Danks, H.V., Downes, J.A., Eds.; Biological Survey of Canada (Terrestrial Arthropods): Ottawa, Canada, 1997; pp. 911–962. [Google Scholar]
- Zazula, G.D.; Froese, D.G.; Schweger, C.E.; Mathewes, C.E.; Beaudoin, A.B.; Telka, A.M.; Harington, C.R.; Westgate, J.A. Ice age steppe vegetation in east Beringia. Nature 2003, 423, 603. [Google Scholar] [CrossRef]
- Matthews, J.V., Jr. Tertiary and Quaternary environments: Historical background for an analysis of the Canadian insect fauna.Canada and its insect fauna. Mem. Ent. Soc. Can. 1979, 108, 31–86. [Google Scholar] [CrossRef]
- Alfimov, A.V.; Berman, D.I.; Sher, A.V. Tundra-steppe insect assemblages and reconstruction of Late Pleistocene climate in the lower reaches of the Kolyma River. Zool. Zhurn. 2003, 82, 281–300. (In Russian) [Google Scholar]
- Matthews, J.V.; Schweger, C.E.; Janssens, J.A. The last (Koy-Yukon) inter- glaciation in the northern Yukon: Evidence from unit 4 at Ch’ijee’s bluff, Bluefish basin. Géogr. Phys. Quat. 1990, 44, 341–352. [Google Scholar]
- Schirrmeister, L.; Froese, D.; Tumskoy, V.; Grosse, G.; Wetterich, S. Yedoma: Late Pleistocene Ice-Rich Syngenetic Permafrost of Beringia. In The Encyclopedia of Quaternary Science; Elias, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 3, pp. 542–552. [Google Scholar]
- Froese, D.G.; Westgate, J.A.; Sanborn, P.T.; Reyes, A.V.; Pearce, N.J.G. The Klondike goldfields and Pleistocene environments of Beringia. GSA Today 2009, 19, 4–10. [Google Scholar] [CrossRef]
- Sher, A.V. Yedoma as a store of paleoenvironmental records in Beringida. In Beringia Paleoenvironmental Workshop, Abstracts and Program; Elias, S., Brigham-Grette, J., Eds.; Ohana Productions: Nepean, ON, Canada, 1997; pp. 92–94. [Google Scholar]
- Gubin, S.V. Pedogenesis–the main component of the late Pleistocene Ice Complex forming. Krioshera Zemli 2002, 6, 82–91. (in Russian). [Google Scholar]
- Wetterich, S.; Schirrmeister, L.; Pietrzeniuk, E. Freshwater ostracodes in Quaternary permafrost deposits in the Siberian Arctic. J. Paleolimnol. 2005, 34, 363–376. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Number of | Region | |||
|---|---|---|---|---|---|
| Coleoptera | Sections | Samples | Individuals | ||
| Gyrinidae | |||||
| 1 | Gyrinus aeratus Stephens, 1835 | 1 | 3 | 10 | KL |
| 2 | G. confinis LeConte, 1868 | 1 | 1 | 1 | AC |
| 3 | G. marinus Gyllenhal, 1808 | 1 | 1 | 1 | OC |
| 4 | G. minutus Fabricius, 1798 | 5 | 6 | 7 | AC, OC; CY, KL |
| 5 | G. natator (Linnaeus, 1758) | 9 | 19 | 77 | CY, KL, SC, WC, YL |
| 6 | G. opacus C.R.Sahlberg, 1817 | 13 | 19 | 28 | OC; CY, KL, LS, YL |
| 7 | G. pectoralis LeConte, 1868 | 2 | 3 | 5 | BB, OC |
| 8 | G. pullatus Zaitsev, 1908 | 2 | 3 | 3 | BB, OC |
| 9 | G. wallisi Fall, 1922 | 1 | 1 | 1 | OC |
| 10 | Gyrinus sp. | 10 | 16 | 19 | AC, BB, OC; CY, KL, LS, SC |
| Haliplidae | |||||
| 11 | Haliplus immaculicollis Harris, 1828 | 2 | 2 | 2 | BB; KL |
| Dytiscidae | |||||
| 12 | Agabus affinis (Paykull, 1798) | 2 | 2 | 2 | OC; CY |
| 13 | A. antennatus Leech, 1939 | 5 | 5 | 17 | BB, OC |
| 14 | A. approximatus Fall, 1922 | 1 | 1 | 1 | KG |
| 15 | A. arcticus (Paykull, 1798) | 7 | 8 | 12 | AC, BB, KG, OC; CY |
| 16 | A. bifarius (Kirby, 1837) | 1 | 1 | 1 | OC |
| 17 | A. clavicornis Sharp, 1882 | 1 | 1 | 1 | KG |
| 18 | A. colymbus Leech, 1938? | 1 | 1 | 1 | OC |
| 19 | A. confinis (Gyllenhal, 1808) | 5 | 2 | 2 | OC |
| 20 | A. discolor (Harris, 1828) | 4 | 1 | 1 | OC |
| 21 | A. elongatus (Gyllenhal, 1826) | 8 | 9 | 3 | AC, KG, OC |
| 22 | A. infuscatus Aube, 1838 | 4 | 4 | 3 | BB, OC |
| 23 | A. inscriptus (Crotch, 1873) | 1 | 1 | 1 | OC |
| 24 | A. lapponicus (Thomson, 1867)? | 11 | 5 | 3 | KL, LS |
| 25 | A. moestus (Curtis, 1835) | 23 | 96 | 317 | BB, KG, NS, OC; KL, LS, SC, WC, YL |
| 26 | A. semipunctatus (Kirby, 1837) | 2 | 4 | 12 | AC, BB |
| 27 | A. seriatus (Say, 1823) | 2 | 2 | 2 | OC |
| 28 | A. serricornis (Paykull, 1799) | 3 | 6 | 21 | NS, OC; CY |
| 39 | A. strigulosus (Crotch, 1873) | 1 | 1 | 1 | OC |
| 30 | A. thomsoni (J.Sahlberg, 1871) | 14 | 19 | 65 | AC, KG, NS; CY, KL, LS, SC, WC, YL |
| 31 | A. tristis Aube, 1838 | 2 | 5 | 11 | AC, OC |
| 32 | Agabus sp. | 58 | 271 | 730 | AC, BB, KG, LS, NS, OC; CY, KL, MR, SC, WS, YL |
| 33 | Colymbetes dahuricus Aube, 1837 | 33 | 69 | 353 | AC, BB, NS, OC; KL, LS, SC, WS, YL |
| 34 | C. dolabratus (Paykull, 1798) | 15 | 48 | 141 | AC, BB, NS, OC; KL, LS, SC, WS, YL |
| 35 | C. paykulli Erichson, 1837 | 13 | 17 | 21 | CY, KL, WS, YL |
| 36 | Colymbetes sp. | 19 | 37 | 50 | AC, BB, KG, NS; CY, KL, WC |
| 37 | Dytiscus circumcinctus Ahrens, 1811 | 3 | 4 | 9 | CY, WC |
| 38 | Dytiscus sp. | 6 | 7 | 8 | BB, KG, OC; KL, LS, YL |
| 39 | Graphoderes zonatus Hoppe, 1795? | 1 | 1 | 3 | CY |
| 40 | Graphoderes sp. | 1 | 2 | 2 | WC |
| 41 | Hydroporus acutangulus Thomson, 1856? | 10 | 45 | 147 | KL, LS, SC, WC, YL |
| 42 | H. appalachius Sherman, 1913 | 2 | 2 | 3 | AC, BB |
| 43 | H. badiellus Fall, 1923 | 3 | 2 | 3 | OC |
| 44 | H. dentellus Fall, 1923 | 1 | 1 | 4 | OC |
| 45 | H. fuscipennis Schaum, 1868 | 8 | 12 | 40 | AC, BB, OC; CY, LS |
| 46 | H. lapponum (Gyllenhal, 1808) | 11 | 19 | 37 | BB, KG, NS; KL, LS, SC |
| 47 | H. morio Aube, 1838 | 12 | 23 | 59 | AC, BB, KG, OC; SC |
| 48 | H. polaris Fall, 1923 | 2 | 11 | 24 | AC, NS |
| 49 | H. puberulus LeConte, 1850? | 1 | 1 | 2 | AC |
| 50 | H. punctipennis J.Sahlberg, 1880 | 1 | 1 | 1 | YL |
| 51 | H. rufinasus Mannerheim, 1852 | 2 | 2 | 2 | BB, OC |
| 52 | H. signatus Mannerheim, 1852 | 1 | 1 | 1 | AC |
| 53 | H. tristis (Paykull 1798) | 2 | 2 | 2 | KG; CY |
| 54 | Hydroporus sp. | 59 | 222 | 538 | AC, BB, KG, NS, OC; CY, KL, LS, MG, SC, WS, YL |
| 55 | Hygrotus sp. | 2 | 2 | 3 | KG; CY |
| 56 | Ilybius erichsoni (Gemminger and Harold, 1868) | 119 | 26 | 10 | AC, BB, KG, OC |
| 57 | I. opacus (Aube, 1837) | 1 | 1 | 3 | AY |
| 58 | I. picipes (Kirby, 1837) | 1 | 1 | 1 | NS |
| 59 | I. subaeneus (Erichson, 1837) | 1 | 1 | 1 | AC |
| 60 | Ilybius sp. | 1 | 1 | 1 | CY |
| 61 | Laccophilus sp.? | 1 | 1 | 1 | KL |
| 62 | Oreodytes dauricus (Motschulsky, 1860) | 1 | 1 | 1 | CY |
| 63 | O. laevis (Kirby, 1837) | 1 | 1 | 1 | NS |
| 64 | O. recticollis (Fall, 1926) | 1 | 1 | 1 | NS |
| 65 | O. sanmarkii (C.R.Sahlberg, 1826) | 11 | 23 | 38 | NS; CY, KL, YL |
| 66 | Rhantus notatus (Fabricius, 1781) | 2 | 2 | 3 | KL, WS |
| 67 | R. frontalis (Marsham, 1802) | 1 | 1 | 1 | KG |
| 68 | Dytiscidae gen. indet. | 10 | 12 | 23 | CY, KL, LS, MR, WC, YL |
| Hydrophilidae | |||||
| 69 | Coelostoma orbicularis (Fabricius, 1775)? | 1 | 1 | 1 | KG |
| 70 | Cymbiodyta acuminata Fall, 1924 | 3 | 3 | 4 | AC, KG, OC |
| 71 | C. marginella (Fabricius, 1792) | 1 | 1 | 2 | CY |
| 72 | C. vindicata Fall, 1924 | 1 | 1 | 1 | AC |
| 73 | Cymbiodyta sp. | 3 | 3 | 5 | AC, KG, NS |
| 74 | Enochrus ochropterus (Marsham, 1802) | 1 | 1 | 1 | CY |
| 75 | Helophorus aspericollis Angus, 1973? | 1 | 2 | 2 | WC |
| 76 | H. lacustris LeConte, 1850 | 1 | 1 | 1 | AC |
| 77 | H. oblongus LeConte, 1850 | 5 | 6 | 7 | AC, KG, OC; WC |
| 78 | H. obscurellus Poppius 1907 | 7 | 9 | 16 | KL, LS, WC, YL |
| 79 | H. sempervarians Angus, 1970? | 1 | 1 | 1 | NS |
| 80 | H. sibiricus (Motschulsky, 1860) | 9 | 11 | 15 | KG; KL, LS, MR, WC |
| 81 | H. splendidus J.Sahlberg, 1880 | 78 | 200 | 445 | AC, BB, KG, NS, OC; CY, KL, LS, WC, YL |
| 82 | H. tuberculatus (Gyllenhal, 1808) | 11 | 11 | 15 | BB, KG; CY, KL, LS, MR, WC |
| 83 | Helophorus sp. | 15 | 20 | 27 | KG, NS, OC; CY, KL, LS, WC |
| 84 | Hydrobius fuscipes (Linnaeus, 1758) | 77 | 123 | 209 | AC, BB, KG, OC; CY, KL, LS, MR, SC, WC, YL |
| 85 | Hydrochus brevis Herbst, 1793? | 1 | 1 | 1 | KL |
| 86 | Hydrochara caraboides (Linnaeus, 1758) | 1 | 1 | 1 | CY |
| 87 | Laccobius sp. | 11 | 15 | 20 | BB, OC; CY, KL, LS, WC, YL |
| 88 | Hydrophilidae gen. indet. | 6 | 6 | 7 | CY, KL, LS, YL |
| Hydraenidae | |||||
| 89 | Hydraena sp. | 1 | 1 | 5 | KG |
| 90 | Limnebius sp. | 1 | 1 | 1 | CY |
| 91 | Ochthebius costatellus Reitter, 1897 | 1 | 1 | 1 | OC |
| 92 | O. cribricollis LeConte, 1850 | 1 | 1 | 1 | BB |
| 93 | O. kaszabi Janssens, 1967. | 7 | 12 | 19 | BB, KG |
| 94 | Ochthebius sp. | 4 | 5 | 8 | KG; CY, YL |
| Dryopidae | |||||
| 95 | Dryopidae gen. indet. | 1 | 1 | 1 | KL |
| Curculionidae | |||||
| 96 | Bagous limosus (Gyllenhal, 1827) | 3 | 4 | 5 | CY, LS |
| 97 | B. magister LeConte, 1876 | 1 | 1 | 5 | OC |
| 98 | Bagous sp. | 5 | 5 | 5 | LS, YL, WC |
| Heteroptera | |||||
| Veliidae | |||||
| 99 | Microvelia umbricola Wroblewsky, 1938 | 1 | 1 | 2 | KL |
| Corixidae | |||||
| 100 | Sigara fallenoidea (Hungerford, 1926) | 1 | 1 | 2 | CY |
| 101 | Sigara sp. | 11 | 22 | 39 | KG, NS; CY, KL, LS, SC, YL |
| 102 | Corixidae gen. indet. | 2 | 2 | 14 | KG |
| Ephemeroptera | |||||
| 103 | Ephemeroptera gen. indet. (nymph) | 2 | 3 | 5 | CY |
| Odonata | |||||
| 104 | Odonata gen. indet. (larvae) | 1 | 1 | 2 | CY |
| Trichoptera | |||||
| Brachycentridae | |||||
| 105 | Brachycentrus subnubilus Curtis, 1834 | 3 | 3 | 10 | CY |
| 106 | Micrasema gelidum McLachlan, 1876 | 1 | 1 | 1 | CY |
| 107 | Micrasema sp. | 2 | 2 | 4 | OC; CY |
| Limnephilidae | |||||
| 108 | Limnephilus sp. | 3 | 3 | 8 | CY |
| Molannidae | |||||
| 109 | Molanna sp. | 1 | 1 | 1 | CY |
| Apataniidae | |||||
| 110 | Apataniidae gen. indet. | 1 | 1 | 1 | CY |
| 111 | Trichoptera gen. indet. (larvae) | 26 | 105 | 520 | AC, BB, NS, OC; CY, KL, LS, SC, YL |
| Taxa | Number of | Region | |||
|---|---|---|---|---|---|
| Coleoptera | Sections | Samples | Individuals | ||
| Carabidae | |||||
| 1 | Agonum (Agonothorax) exaratum (Mannerheim, 1853) | 2 | 5 | 5 | AC; WC |
| 2 | A. (Agonothorax) impressum (Panzer, 1796) | 1 | 2 | 3 | LS |
| 3 | A. (Agonum) carbonarium Dejean, 1828 | 2 | 6 | 12 | KL, LS |
| 4 | A. (Agonum) sp. | 2 | 2 | 4 | KL, MR |
| 5 | A. (Europhilus) consimile (Gyllenhal, 1810) | 13 | 20 | 57 | AC, BB, KG, NS, OC; KL |
| 6 | A. (Europhilus) fuliginosum (Panzer, 1809) | 4 | 6 | 7 | KL, LS |
| 7 | A. (Europhilus) sexpunctatum (Linnaeus, 1758) | 4 | 5 | 5 | CY |
| 8 | A. (Europhilus) simile Kirby 1837 | 4 | 6 | 8 | BB, KG, OC |
| 9 | A. (Europhilus) sordens Kirby, 1837? | 1 | 1 | 5 | AC |
| 10 | A. (Europhilus) superioris Lindroth, 1966 | 2 | 2 | 2 | AC, OC |
| 11 | A. (Europhilus) thoreyi Dejean, 1828 | 2 | 2 | 3 | AC, BB |
| 12 | A. (Europhilus) thoreyi Dej.? | 1 | 1 | 1 | AC |
| 13 | A. (Europhilus) sp. | 8 | 19 | 37 | CY, KL, WC, YL |
| 14 | A. (Olisares) affine Kirby, 1837 | 4 | 5 | 6 | BB, KG, OC |
| 15 | A. (Olisares) ericeti quinquepunctatum Motschulsky, 1844 | 24 | 37 | 2 | AC, BB, KG, OC; KL, LS, MR, SC, WC, YL |
| 16 | A. (Olisares) harrisi LeConte, 1846 | 2 | 2 | AC, OC | |
| 17 | A. (Olisares) mutatum (Gemminger & Harold, 1868) | 2 | 2 | 3 | AC, OC |
| 18 | Agonum sp. | 16 | 27 | 43 | AC, KG; CY, KL, LS, MR, SC, WC, YL |
| 19 | Amara (Curtonotus) lacustris LeConte, 1855 | 5 | 6 | 8 | AY, BB, OC |
| 20 | Asaphidion alaskanum Wickham 1919 | 2 | 2 | 2 | NS |
| 21 | A. yukonense Wickham 1919 | 1 | 1 | 1 | NS |
| 22 | Asaphidion sp. | 2 | 5 | 12 | AY, CY |
| 23 | Bembidion (Asioperyphus) bimaculatum (Kirby, 1837) | 7 | 9 | 11 | NS, CY, KL, SC, WC, YL |
| 24 | B. (Asioperyphus) umiatense Lindroth, 1963 | 38 | 124 | 321 | AY, BB, KG, NS, OC, CY, KL, LS, SC, WC, YL |
| 25 | B. (Asioperyphus) sp. | 1 | 1 | 4 | CY |
| 26 | B. (Bembidion) quadrimaculatum (Linnaeus, 1761) | 3 | 5 | 15 | BB, KL, YL |
| 27 | B. (Bembidion) sp.? | 3 | 4 | 5 | KL, YL |
| 28 | B. (Bracteon) foveum Motschulsky, 1844 | 1 | 1 | 1 | BB |
| 29 | B. (Bracteon) lapponicum Zetterstedt, 1828 | 3 | 3 | 4 | CY, LS |
| 30 | B. (Bracteon) sp. | 2 | 2 | 2 | CY |
| 31 | B. (Dioplocampa) transparens (Gebler, 1829) | 2 | 2 | 2 | AC, BB |
| 32 | B. (Eurytrachelus) interventor Lindroth, 1963 | 1 | 1 | 1 | BB |
| 33 | B. (Hidromicrus) semistriatum (Haldeman, 1843) | 3 | 5 | 9 | OC |
| 34 | B. (Hirmoplataphus) concolor (Kirby, 1837) | 1 | 1 | 1 | OC |
| 35 | B. (Hirmoplataphus) nigrum Say, 1823 | 2 | 2 | 6 | AC, OC |
| 36 | B. (Hirmoplataphus) salebratum (LeConte, 1848) | 1 | 1 | 1 | BB |
| 37 | B. (Leuchydrium) scudderi LeConte, 1878 | 2 | 3 | 3 | BB, KG |
| 38 | B. (Notaphus) coloradense Hayward, 1897 | 1 | 2 | 2 | AC |
| 39 | B. (Notaphus) insulatum LeConte 1852 | 1 | 1 | 2 | BB |
| 40 | B. (Notaphus) nigripes (Kirby, 1837) | 2 | 2 | 3 | BB, KG |
| 41 | B. (Notaphus) patruele Dejean, 1831 | 2 | 2 | 3 | AC, OC |
| 42 | B. (Notaphus) semipunctatum Donovan, 1806 | 1 | 1 | 1 | BB |
| 43 | B. (Notaphus) varium (Olivier, 1795) | 2 | 2 | 2 | LS |
| 44 | B. (Notaphus) sp. | 2 | 2 | 3 | KG; WC |
| 45 | B. (Ocydromus) obscurellum (Motschulsky, 1845) | 1 | 1 | 1 | KG; WC |
| 46 | B. (Ocydromus) sp. | 1 | 1 | 1 | KG; WC |
| 47 | B. (Peryphanes) yukonum Fall, 1926 | 4 | 8 | 13 | BB, OC |
| 48 | B. (Peryphus) petrosum Gebler, 1833 | 8 | 10 | 13 | AC; CY, KL, LS, WC, YL |
| 49 | B. (Peryphus) sejunctum Casey, 1918? | 1 | 1 | 1 | AC |
| 50 | B. (Peryphus) sp. | 12 | 26 | 46 | KL, LS, SC, WC, YL |
| 51 | B. (Phyla) sp.? | 1 | 1 | 1 | YL |
| 52 | B. (Plataphodes) arcticum Lindroth 1963 | 21 | 44 | 79 | AC, BB, KG, NS, OC; KL, LS, WC, YL |
| 53 | B. (Plataphodes) complanulum (Mannerheim, 1853) | 1 | 1 | 5 | OC |
| 54 | B. (Plataphodes) compressum Lindroth 1963? | 1 | 1 | 1 | KG |
| 55 | B. (Plataphodes) quadrifoveolatum Mannerheim, 1843 | 1 | 1 | 1 | KG |
| 56 | B. (Plataphodes) sp. | 4 | 4 | 5 | BB, OC; SC, WC |
| 57 | B. (Plataphus) curtulatum Casey, 1918 | 1 | 1 | 2 | OC |
| 58 | B. (Plataphus) gebleri (Gebler, 1833) | 2 | 5 | 6 | LS, WC |
| 59 | B. (Plataphus) hyperboraeorum Munster, 1923 | 7 | 14 | 24 | BB, KG, NS; LS, WC |
| 60 | B. (Plataphus) kuprianovi Mannerheim, 1843 | 1 | 1 | 8 | KG |
| 61 | B. (Plataphus) planatum (LeConte, 1847) | 1 | 1 | 1 | OC |
| 62 | B. (Plataphus) quadrifoveolatum Mannerheim, 1843 | 1 | 1 | 1 | KG |
| 63 | B. (Plataphus) sp. | 10 | 15 | 32 | KG; KL, LS, SC, YL |
| 64 | B. (Pseudoperyphus) chalceum (Stephens, 1828) | 1 | 1 | 1 | BB |
| 65 | B. (Semicampa) roosvelti Pic, 1902 | 2 | 3 | 5 | BB, OC |
| 66 | B. (Trepanedoris) acutifrons LeConte, 1879? | 1 | 1 | 1 | OC |
| 67 | B. (Trepanedoris) concretum Casey, 1918 | 4 | 9 | 11 | AC, BB, KG, OC |
| 68 | B. (Trepanedoris) fortestriatum (Motschulsky 1845) | 3 | 4 | 6 | AC, OC |
| 69 | Bembidion sp. | 40 | 167 | 711 | |
| 70 | Blethisa inexspectata Goulet and Smetana, 1983 | 4 | 6 | 6 | BB, KG; KL |
| 71 | B. multipunctata (Linnaeus, 1758) | 9 | 15 | 18 | BB, OC; KL, LS, YL |
| 72 | Dyschiriodes globulosus Herbst, 1784 | 5 | 5 | 12 | AC, KG |
| 73 | D. melancholicus (Putzeys, 1866) | 43 | 107 | 191 | AC, BB, KG, NS, OC; CY, KL, LS, WC, YL |
| 74 | Dyschiriodes sp. | 7 | 11 | 15 | AC, KG; CY, LS, WC |
| 75 | Dyschirius aeneolus LeConte, 1850 | 1 | 1 | 2 | KG |
| 76 | D. integer LeConte, 1852 | 8 | 16 | 29 | BB, KG, OC |
| 77 | D. setosus LeConte, 1857? | 1 | 1 | 1 | KG |
| 78 | D. subarcticus Lindroth, 1961 | 1 | 1 | 3 | BB, KG, OC |
| 79 | Elaphrus angusticollis Putzeys, 1867 | 7 | 20 | 46 | BB, KG, OC |
| 80 | E. cupreus Duftschmid, 1812 | 1 | 1 | 1 | KL |
| 81 | E. lapponicus Gyllenhal, 1810 | 24 | 52 | 81 | BB, KG; CY, KL, LS, WC, YL |
| 82 | E. parviceps Van Dyke, 1925 | 10 | 23 | 65 | AC, KG, NS, OC |
| 83 | E. riparius (Linnaeus, 1758) | 28 | 76 | 142 | BB, OC; CY, KL, LS, SC, WC, YL |
| 84 | E. splendidus Fischer von Waldheim, 1828 | 8 | 10 | 11 | CY, KL, SC |
| 85 | E. tuberculatus Mäklin, 1878 | 1 | 1 | 2 | KG |
| 86 | Elaphrus sp. | 13 | 29 | 36 | AY, BB, KG, OC, CY, LS, WC, YL |
| 87 | Nebria frigida R.F.Sahlberg, 1844 | 13 | 24 | 36 | KL, LS, MR, WC, YL |
| 88 | N. gyllenhali (Strom, 1768) | 3 | 5 | 8 | KG |
| 89 | N. metallica Fischer von Waldheim, 1820 | 1 | 1 | 3 | KG |
| 90 | N. nivalis (Paykull, 1790) | 11 | 15 | 19 | BB, KG, OC; CY, WC, YL |
| 91 | N. rufescens (Strom, 1768) | 2 | 2 | 2 | KG; CY |
| 92 | N. sahlbergi Fischer von Waldheim, 1828 | 1 | 1 | 5 | BB |
| 93 | Nebria sp. | 1 | 1 | 1 | LS |
| 94 | Opisthius richardsoni Kirby, 1837 | 1 | 1 | 1 | KG |
| 95 | Patrobus septentrionis Dejean, 1828 | 9 | 10 | 15 | AC, BB, KG, OC; CY, WC, YL |
| 96 | Patrobus sp. | 3 | 3 | 8 | AC, BB, KG, OC; CY, WC, YL |
| 97 | Pelophila borealis (Paykull, 1790) | 15 | 17 | 24 | AC, BB, OC; KL, LS, WC, YL |
| 98 | Platynus mannerheimii (Dejean, 1828) | 1 | 2 | 3 | KG |
| 99 | Pterostichus (Argutor) patruelis (Dejean, 1831)? | 1 | 2 | 14 | AC |
| 100 | P. (Cryobius) hudsonicus LeConte, 1863. | 1 | 1 | 2 | OC |
| 101 | P. (Petrophilus) uralensis Motschulsky, 1850 | 2 | 2 | 2 | KL, LS |
| 102 | P. (Steropus) circulosus Lindroth, 1966? | 1 | 1 | 1 | NS |
| 103 | P. (Steropus) maurusiacus Mannerheim, 1825? | 2 | 4 | 15 | KL |
| 104 | Sericoda obsoleta (Say, 1823) | 1 | 1 | 1 | OC |
| 105 | S. quadripunctata (De Geer, 1774) | 5 | 5 | 7 | AC, OC, LS |
| 106 | Trechus apicalis Motschulsky, 1845 | 2 | 2 | 2 | CY, LS |
| Hydrophilidae | |||||
| 107 | Cercyon borealis Baranowski, 1985 | 1 | 1 | 1 | CY |
| 108 | Coelostoma orbiculare Fabricius, 1775 | 6 | 7 | 16 | CY, KL, YL |
| 109 | Cymbiodyta marginella (Fabricius, 1792) | 1 | 1 | 5 | OC |
| 110 | Cymbiodyta sp. | 6 | 7 | 13 | CY, KL, MR, WC, YL |
| 111 | Enochrus testaceus (Fabricius, 1801) | 1 | 1 | 1 | CY |
| 112 | Georissus crenulatus (Rossi, 1794) | 1 | 1 | 1 | CY |
| 113 | Georissus sp. | 2 | 4 | 10 | CY |
| 114 | Megasternum posticatum (Mannerheim, 1852) | 1 | 1 | 5 | KG |
| Staphylinidae | |||||
| 115 | Acylophorus pronus Erichson, 1840? | 1 | 2 | 2 | ER |
| 116 | Bledius confusus LeConte, 1877 | 1 | 1 | 1 | OC |
| 117 | B. viriosus Herman, 1983 | 1 | 1 | 10 | KG |
| 118 | B. zophus Herman, 1983 | 1 | 1 | 1 | BB |
| 119 | B. (Hesperophilus) sp. | 1 | 3 | 9 | WC |
| 120 | Bledius sp. | 4 | 4 | 6 | KG; CY, LS, MR |
| 121 | Boreaphilus henningianus C.R.Sahlberg, 1832 | 1 | 1 | 2 | KG |
| 122 | Coryphium hyperboreum (Maklin, 1880) | 2 | 3 | 3 | BB, OC |
| 123 | Erichsonius nanus (Horn, 1884)? | 1 | 1 | 1 | AC |
| 124 | Eucnecosum brachypterum (Gravenhorst, 1802) | 1 | 1 | 1 | AC |
| 125 | Geodromicus verticalis (Say, 1831) | 1 | 1 | 1 | OC |
| 126 | Holoboreaphilus nordenskioeldi (Maklin, 1878) | 16 | 37 | 188 | AY, KG, KL, MR, LS, WS |
| 127 | Kalissus nitidus LeConte, 1874 | 1 | 1 | 1 | OC |
| 128 | Microedus austinianus LeConte, 1874 | 1 | 1 | 2 | KG |
| 129 | Olophrum boreale (Paykull, 1792) | 4 | 5 | 10 | BB, KG, OC |
| 130 | O. latum Maklin, 1853 | 15 | 44 | 340 | AC, BB, KG, NS |
| 131 | O. rotundicolle (C.R.Sahlberg, 1817) | 11 | 22 | 98 | AC, ER, KG, NS, OC |
| 132 | Stenus aureolus Fauvel, 1875 | 1 | 1 | 2 | CY |
| 133 | S. fuscipes Gravenhorst, 1802 | 2 | 2 | 2 | CY |
| 134 | S. palustris Erichson, 1839 | 1 | 1 | 2 | CY |
| 135 | Stenus sp. | 50 | 112 | 248 | AC, BB, KG, NS, OC, CY, KL, LS, MR, SC, WC, YL |
| Scarabaeidae | |||||
| 136 | Aegialia kamtschatica Motschulsky, 1860 | 6 | 13 | 20 | CY, LS, WC |
| 137 | A. cylindrica (Eschscholz, 1822) | 3 | 4 | 11 | AC |
| 138 | A. lacustris LeConte, 1850 | 5 | 6 | 27 | BB, KG, OC |
| 139 | A. terminalis Brown, 1931 | 9 | 10 | 25 | AC, BB, KG |
| 140 | Aegialia sp. | 3 | 3 | 11 | AC, KG, OC |
| Scirtidae | |||||
| 141 | Cyphon variabilis (Thunberg, 1787) | 9 | 13 | 17 | AC, KG, OC, CY |
| 142 | Cyphon sp. | 14 | 18 | 35 | AC, KG; CY, KL, MR, YL |
| Heteroceridae | |||||
| 143 | Heterocerus fenestatus (Thunberg, 1784) | 1 | 1 | 1 | CY |
| 144 | Heterocerus sp. | 7 | 7 | 7 | OC; CY, KL, LS, YL |
| 145 | Lanternarius brunneus (Melsheimer, 1844) | 7 | 7 | 8 | AC, BB, KG, OC |
| 146 | Lapsus tristis (Mannerheim, 1853) | 1 | 1 | 1 | KG |
| Elateridae | |||||
| 147 | Hypnoidus rivularius (Gyllenhal, 1827) | 7 | 8 | 8 | BB, KG; CY |
| 148 | Limonius aeneoniger (DeGeer, 1774) | 2 | 2 | 2 | AC; WC |
| 149 | Limonius sp. | 1 | 1 | 1 | WC |
| 150 | Negastrius extricatus (Fall, 1926) | 1 | 1 | 1 | KG |
| 151 | Negastrius sp. | 6 | 11 | 11 | KG, OC; KL, MR, YL |
| 152 | Oedostethus femoralis LeConte, 1853 | 1 | 3 | 3 | CY |
| Coccinellidae | |||||
| 153 | Hippodamia arctica (Schneider, 1787) | 18 | 34 | 46 | BB, KG, OC; CY, KL, LS, MG, SC, WC, YL |
| 154 | Nephus bipunctatus (Kugelann, 1794) | 1 | 1 | 1 | CY |
| 155 | Scymnus sp. | 9 | 21 | 29 | CY, KL, LS, WC |
| Chrysomelidae | |||||
| 156 | Donacia cazieri Marx, 1957? | 1 | 1 | 1 | OC |
| 157 | D. crassipes Fabricius, 1775 | 1 | 2 | 2 | CY |
| 158 | D. distincta LeConte, 1851 | 2 | 2 | 2 | AC, OC |
| 159 | D. hirticollis Kirby, 1837? | 2 | 2 | 2 | BB, OC |
| 160 | D. porosicollis Lacordaire, 1845 | 1 | 1 | 1 | OC |
| 161 | D. proxima Kirby, 1837 | 1 | 1 | 1 | OC |
| 162 | D. semicuprea Panzer, 1796 | 1 | 1 | 1 | CY |
| 163 | D. splendens Jacobson, 1894 | 2 | 2 | 2 | CY |
| 164 | Donacia sp. | 22 | 29 | 41 | AC, KG, OC; CY, KL, LS, WC, YL |
| 165 | Hippuriphila canadensis Brown, 1942 | 1 | 1 | 1 | AC |
| 166 | H. mancula (LeConte, 1861) | 1 | 1 | 2 | KG |
| 167 | Hydrothassa glabra (Herbst, 1783 | 3 | 3 | 3 | LS |
| 168 | H. hannoverana (Fabricius, 1775) | 24 | 79 | 115 | KL, LS, SC, WC, YL |
| 169 | Plateumaris flavipes (Kirby, 1837) | 3 | 3 | 13 | AC, BB |
| 170 | P. frosti (Schaeffer, 1935)? | 1 | 1 | 1 | KG |
| 171 | P. robusta (Schaeffer, 1920) | 1 | 1 | 1 | OC |
| 172 | P. sericea (Linnaeus, 1760) | 1 | 1 | 1 | CY |
| 173 | Plateumaris sp. | 8 | 9 | 12 | AC, CY, KL, WC |
| 174 | Prasocuris boreella (Schaeffer, 1928) | 3 | 3 | 3 | AC, BB, KG |
| 175 | P. phellandrii (Linnaeus, 1758) | 2 | 2 | 2 | LS, WC |
| Curculionidae | |||||
| 176 | Auleutes epilobii (Paykull, 1800) | 3 | 3 | 3 | AC, BB, KG |
| 177 | A. nebulosus (LeConte, 1876) | 1 | 1 | 1 | BB |
| 178 | Erirhinus aethiops (Fabricius, 1792) | 24 | 55 | 134 | KG, NS, OC, CY, KL, LS, MR, SC, WC, YL |
| 179 | Eubrychius velutus (Beck, 1817) | 2 | 2 | 5 | CY, KL |
| 180 | Grypidius equiseti (Fabricius, 1775) | 16 | 30 | 50 | AC, BB; CY, KL, WC, YL |
| 181 | Grypus mannerheimi Faust, 1881 | 10 | 16 | 38 | CY, KL, LS, WS, YL |
| 182 | Limnobaris dolorosa (Goeze, 1777) | 1 | 1 | 1 | CY |
| 183 | Listronotus humilis (Gyllenhal, 1834) | 1 | 1 | 6 | OC |
| 184 | Lixus paraplecticus (Linnaeus, 1758) | 1 | 1 | 1 | LS |
| 185 | Notaris acridulus (Linnaeus, 1758) | 4 | 6 | 8 | CY, WC, YL |
| 186 | N. bimaculatus (Fabricius, 1792) | 49 | 222 | 675 | AY, BB, KG, OC, CY, KL, LS, SC, WC, YL |
| 187 | N. eversmanni Faust 1882 | 9 | 17 | 67 | KL, LS, SC, WC, YL |
| 188 | N. ochoticus Korotyaev, 1984 | 9 | 23 | 47 | CY, KL, LS, WC, YL |
| 189 | N. puncticollis (LeConte 1876) | 2 | 2 | 6 | BB, OC |
| 190 | Notaris sp. | 6 | 8 | 9 | AC, BB, OC; WC, YL |
| 191 | Pelenomus quadrituberculatus (Fabricius, 1787) | 3 | 3 | 3 | LS |
| 192 | P. velaris (Gyllenhal, 1827) | 17 | 37 | 77 | CY, KL, LS, SC, WC, YL |
| 193 | Pelenomus sp. | 9 | 24 | 39 | KL, LS, WC, YL, BB, KG |
| 194 | Phytobius leucogaster (Marsham, 1802) | 3 | 4 | 4 | OC, KL |
| 195 | Procas lecontei Bedel, 1879 | 3 | 4 | 5 | BB, OC |
| 196 | Rhinoncus castor (Fabricius, 1792) | 3 | 4 | 5 | KL |
| 197 | R. jakovlevi Faust, 1893 | 1 | 1 | 1 | WC |
| Heteroptera | |||||
| Saldidae | |||||
| 198 | Chiloxanthus stellatus (Curtis, 1835) | 4 | 8 | 9 | KG, OC, CY |
| 199 | Chiloxanthus sp. | 5 | 8 | 9 | AC, EA, KG, NS; KL |
| 200 | Salda littoralis (Linnaeus, 1758) | 4 | 15 | 20 | AC; LS, CY, YL |
| 201 | Salda sp. | 8 | 13 | 18 | KG; KL, LS, SC, YL |
| 202 | Saldula fucicola (J.Sahlberg, 1870) | 1 | 1 | 1 | CY |
| 203 | Saldula sp. | 5 | 8 | 13 | KG, NS; SC |
| 204 | Teloleuca pellucens (Fabricius, 1779) | 2 | 2 | 4 | CY, LS |
| 205 | Saldidae gen. indet. | 6 | 8 | 10 | BB, KG, OC |
| Age | N of Aquatic Insects | % | N of Wetland and Riparian Insects | % | N of All Insects |
|---|---|---|---|---|---|
| Late Neogene | 37 | 5 | 116 | 15 | 760 |
| Early Pleistocene | 237 | 4 | 421 | 7 | 6125 |
| Middle Pleistocene | 1392 | 3 | 1715 | 4 | 42,063 |
| Late Pleistocene (MIS5) | 359 | 6 | 471 | 8 | 5590 |
| Late Pleistocene (MIS4-2) | 1530 | 3 | 1302 | 3 | 46,047 |
| Holocene | 934 | 13 | 1058 | 15 | 6952 |
| Region, Section, Sample, Age | Recorded aq and ri Species | Habitat | MNI | N of Insects in Sample | % | |
|---|---|---|---|---|---|---|
| aq | ri | |||||
| Buried soil | ||||||
| AC, Tempest Lake, KL-03-TL-003, LP | Chiloxanthus sp. | ri | 1 | 526 | 0 | 0.2 |
| OC, CRH 44, OC-45, MP | Blethisa multipunctata (Linnaeus, 1758) | ri | 1 | 368 | 3 | 5 |
| Elaphrus angusticollis Putzeys, 1867 | ri | 4 | ||||
| E. parviceps Van Dyke, 1925 | ri | 3 | ||||
| E. riparius (Linnaeus, 1758) | ri | 2 | ||||
| Bembidion (Asioperyphus) umiatense Lindroth, 1963 | ri | 3 | ||||
| Agonum (Europhilus) simile Kirby 1837 | ri | 1 | ||||
| Hydroporus morio Aube, 1838 | aq | 4 | ||||
| Agabus confinis (Gyllenhal, 1808) | aq | 2 | ||||
| Colymbetes dolabratus (Paykull, 1798) | aq | 4 | ||||
| Helophorus oblongus LeConte, 1850 | aq | 1 | ||||
| Cyphon variabilis (Thunberg, 1787) | ri | 1 | ||||
| Hippodamia arctica (Schneider, 1787) | ri | 3 | ||||
| Notaris puncticollis (LeConte 1876) | ri | 1 | ||||
| Trichoptera gen. indet. (larvae) | aq | 1 | ||||
| KG, Lucky Lady, D-D-L-9, LP | ––––––––– | 0 | 0 | 125 | 0 | 0 |
| KL, Duvanny Yar, P-1212s, LP3 | Bembidion (Plataphus) sp. | ri | 1 | 90 | 0 | 2 |
| Notaris bimaculatus (Fabricius, 1792) | ri | 1 | ||||
| Peat | ||||||
| OC, CRH-11, OC-8, LP | Gyrinus opacus C.R.Sahlberg, 1817 | aq | 1 | 139 | 24 | 15 |
| Pelophila borealis (Paykull, 1790) | ri | 5 | ||||
| Blethisa multipunctata (Linnaeus, 1758) | ri | 1 | ||||
| Bembidion (Asioperyphus) umiatense Lindroth, 1963 | ri | 2 | ||||
| Agonum (Europhilus) consimile (Gyllenhal, 1810) | ri | 2 | ||||
| Hydroporus morio Aube, 1838 | aq | 2 | ||||
| H. lapponum (Gyllenhal, 1808) | aq | 2 | ||||
| Agabus bifarius (Kirby, 1837) | aq | 1 | ||||
| A. erichsoni (Gemminger and Harold, 1868) | aq | 3 | ||||
| A. tristis Aube, 1838 | aq | 1 | ||||
| A. arcticus (Paykull, 1798) | aq | 1 | ||||
| A. moestus (Curtis, 1835) | aq | 3 | ||||
| Colymbetes dolabratus (Paykull, 1798) | aq | 4 | ||||
| Olophrum latum Maklin, 1853 | ri | 7 | ||||
| Helophorus splendidus J.Sahlberg, 1880 | aq | 1 | ||||
| Hydrobius fuscipes (Linnaeus, 1758) | aq | 1 | ||||
| Cyphon variabilis (Thunberg, 1787) | ri | 1 | ||||
| Hippodamia arctica (Schneider, 1787) | ri | 3 | ||||
| Trichoptera gen. indet. (larvae) | aq | 14 | ||||
| KG, Lucky Lady, D-L-16, H | Elaphrus lapponicus Gyllenhal, 1810 | ri | 2 | 68 | 3 | 15 |
| E. angusticollis Putzeys, 1867 | ri | 2 | ||||
| Dyschirius integer LeConte, 1852 | ri | 1 | ||||
| Dyschiriodes melancholicus (Putzeys, 1866) | ri | 1 | ||||
| Patrobus septentrionis Dejean, 1828 | ri | 3 | ||||
| Platynus mannerheimii (Dejean, 1828) | ri | 1 | ||||
| Helophorus tuberculatus Gyll. | aq | 1 | ||||
| Hydrobius fuscipes (Linnaeus, 1758) | aq | 1 | ||||
| LS, Kurungnakh Isl., BKh -4, H | Bembidion (Peryphus) petrosum Gebler, 1833 | ri | 1 | 71 | 11 | 8 |
| Agonum (Agonum) carbonarium Dejean, 1828 | ri | 1 | ||||
| Agabus moestus (Curtis, 1835) | aq | 1 | ||||
| Hydroporus acutangulus Thomson, 1856? | aq | 2 | ||||
| Helophorus splendidus J.Sahlberg, 1880 | aq | 1 | ||||
| Hydrobius fuscipes (Linnaeus, 1758) | aq | 1 | ||||
| Holoboreaphilus nordenskioeldi (Maklin, 1878) | ri | 2 | ||||
| Stenus sp. | ri | 1 | ||||
| Hydrothassa hannoverana (Fabricius, 1775) | ri | 1 | ||||
| Trichoptera gen. indet. (larvae) | aq | 3 | ||||
| KG, Hollis Mine D-H-41, MP | Dyschiriodes melancholicus (Putzeys, 1866) | ri | 13 | 131 | 1 | 12 |
| Bembidion (Trepanedoris) concretum Casey, 1918 | ri | 1 | ||||
| B. (Notaphus) nigripes (Kirby, 1837) | ri | 1 | ||||
| Agonum (Europhilus) consimile (Gyllenhal, 1810) | ri | 1 | ||||
| Hydrobius fuscipes (Linnaeus, 1758) | aq | 1 | ||||
| Grassy sediment | ||||||
| LS, Samoylov Isl., Sam-1-3, H | Dyschiriodes melancholicus (Putzeys, 1866) | ri | 1 | 124 | 94 | 4 |
| Agonum (Europhilus) fuliginosum (Panzer, 1809) | ri | 2 | ||||
| Agabus thomsoni (J.Sahlberg, 1871) | aq | 4 | ||||
| Holoboreaphilus nordenskioeldi (Maklin, 1878) | ri | 1 | ||||
| Salda sp. | ri | 1 | ||||
| Trichoptera gen. indet. (larvae) | aq | 112 | ||||
| Alluvium | ||||||
| NS, Titaluk, T03-13, MP | Elaphrus parviceps Van Dyke, 1925 | ri | 1 | 389 | 3 | 2 |
| Bembidion (Plataphodes) arcticum Lindroth, 1963 | ri | 1 | ||||
| B. (Asioperyphus) sordidum (Kirby, 1837) | ri | 1 | ||||
| B. (Asioperyphus) umiatense Lindroth, 1963 | ri | 2 | ||||
| Asaphidion alaskanum Wickham 1919 | ri | 1 | ||||
| Pterostichus (Steropus) circulosus Lindroth, 1963 | ri | 1 | ||||
| Agabus serricornis (Paykull, 1799) | aq | 3 | ||||
| Hydroporus polaris Fall, 1923 | aq | 1 | ||||
| H. lapponum (Gyllenhal, 1808) | aq | 5 | ||||
| Colymbetes sp. | aq | 1 | ||||
| Helophorus splendidus J.Sahlberg, 1880 | aq | 2 | ||||
| KL, Alazea, AL-66-2, MP | Bembidion (Peryphus) sp. | ri | 1 | 249 | 4 | 2 |
| Agabus moestus (Curtis, 1835) | aq | 2 | ||||
| Colymbetes dolabratus (Paykull, 1798) | aq | 1 | ||||
| Hydroporus sp. | aq | 1 | ||||
| Dryopidae gen. indet. | aq | 1 | ||||
| Hydrothassa hannoverana (Fabricius, 1775) | ri | 1 | ||||
| Notaris bimaculatus (Fabricius, 1792) | ri | 2 | ||||
| Trichoptera gen. indet. (larvae) | aq | 5 | ||||
| KL, Alazea, AL-203-6, H | Gyrinus aeratus Stephens, 1835 | aq | 3 | 62 | 32 | 13 |
| Blethisa multipunctata (Linnaeus, 1758) | ri | 1 | ||||
| Elaphrus riparius (Linnaeus, 1758) | ri | 1 | ||||
| Bembidion (Asioperyphus) umiatense Lindroth, 1963 | ri | 2 | ||||
| Bembidion (Plataphus) sp. | ri | 1 | ||||
| Agabus lapponicus (Thomson, 1867) | aq | 2 | ||||
| A. serricornis (Thomson, 1867)? | aq | 2 | ||||
| A. thomsoni (J.Sahlberg, 1871) | aq | 4 | ||||
| Dytiscus sp. | aq | 1 | ||||
| Hydroporus fuscipennis Schaum, 1868 | aq | 1 | ||||
| H. lapponum (Gyllenhal, 1808) | aq | 2 | ||||
| Helophorus sibiricus (Motschulsky, 1860) | aq | 1 | ||||
| Hydrobius fuscipes (Linnaeus, 1758) | aq | 4 | ||||
| Donacia sp. | ri | 1 | ||||
| Grypidius equiseti (Fabricius, 1775) | ri | 1 | ||||
| Erirhinus aethiops (Fabricius, 1792) | ri | 1 | ||||
| WC, Amguema, Am-3, H | Bembidion (Asioperyphus) umiatense Lindroth, 1963 | ri | 5 | 143 | 4 | 10 |
| B. (Plataphodes) arcticum Lindroth, 1963 | ri | 2 | ||||
| B. (Plataphus) hyperboraeorum Munster, 1923 | ri | 3 | ||||
| Bembidion sp. | ri | 1 | ||||
| Agonum (Agonothorax) exaratum (Mannerheim, 1853) | ri | 1 | ||||
| Agabus moestus (Curtis, 1835) | aq | 2 | ||||
| Hydroporus sp. | aq | 2 | ||||
| Dytiscidae gen. indet. | aq | 1 | ||||
| Erirhinus aethiops (Fabricius, 1792) | ri | 1 | ||||
| Notaris bimaculatus (Fabricius, 1792) | ri | 1 | ||||
| Trichoptera gen. indet. (larvae) | aq | 1 | ||||
| Lacustrine | ||||||
| KL, Alazea, AL-73-1,2, EP | Colymbetes dolabratus (Paykull, 1798) | aq | 49 | 89 | 55 | 2 |
| Pelenomus velaris Gyll. | ri | 1 | ||||
| OC, REM-78-1, OC-53, MP | Elaphrus sp. | ri | 1 | 69 | 4 | 12 |
| Dyschiriodes melancholicus (Putzeys, 1866) | ri | 2 | ||||
| Agabus sp. | aq | 1 | ||||
| Gyrinus minutus Fabricius, 1798 | aq | 1 | ||||
| Hydrobius fuscipes (Linnaeus, 1758) | aq | 1 | ||||
| Notaris bimaculatus (Fabricius, 1792) | ri | 5 | ||||
| Elaphrus parviceps Van Dyke, 1925 | ri | 1 | ||||
| NS, Titaluk, T03-18, H | Agabus sp. | aq | 1 | 207 | 1 | 1 |
| Colymbetes dolabratus (Paykull, 1798) | aq | 2 | ||||
| Olophrum latum Maekl. | ri | 1 | ||||
| Chiloxanthus sp. | ri | 1 | ||||
| KL, Medvezhyi Isls, Medv-1-11, H | Gyrinus opacus C.R.Sahlberg, 1817 | aq | 1 | 90 | 20 | 14 |
| Elaphrus riparius (Linnaeus, 1758) | ri | 2 | ||||
| Bembidion (Asioperyphus) umiatense Lindroth, 1963 | ri | 4 | ||||
| Bembidion (Plataphus) sp. | ri | 2 | ||||
| Agabus moestus (Curtis, 1835) | aq | 3 | ||||
| Colymbetes dolabratus (Paykull, 1798) | aq | 6 | ||||
| Hydroporus lapponum (Gyllenhal, 1808) | aq | 2 | ||||
| Helophorus splendidus J.Sahlberg, 1880 | aq | 2 | ||||
| Hydrobius fuscipes (Linnaeus, 1758) | aq | 1 | ||||
| Stenus sp. | ri | 1 | ||||
| Notaris bimaculatus (Fabricius, 1792) | ri | 4 | ||||
| Sigara sp. | aq | 3 | ||||
| Yedoma | ||||||
| LS, Kurungnakh Isl., Bkh-02-13, LP | Hydrothassa glabra (Herbst, 1783) | ri | 1 | 297 | 1 | 1 |
| Sigara sp. | aq | 1 | ||||
| Salda sp. | ri | 1 | ||||
| Trichoptera gen. indet. (larvae) | aq | 3 | ||||
| YL, Yana, Tums1-11, LP | Bembidion (Asioperyphus) umiatense Lindroth, 1963 | ri | 2 | 119 | 4 | 3 |
| Helophorus splendidus J.Sahlberg, 1880 | aq | 4 | ||||
| Notaris sp. | ri | 1 | ||||
| Sigara sp. | aq | 1 | ||||
| LS, B. Lyakhovsky Isl, LR-18-9, LP | ––––––––– | 0 | 0 | 102 | 0 | 0 |
| KG, Quartz Creek, D-Q-11, LP | ––––––––– | 0 | 0 | 161 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuzmina, S. Wetlands in the Pleistocene Steppe-Tundra Landscapes of Beringia, Their Insects, and the Role of Aeolian Sedimentation. Water 2023, 15, 494. https://doi.org/10.3390/w15030494
Kuzmina S. Wetlands in the Pleistocene Steppe-Tundra Landscapes of Beringia, Their Insects, and the Role of Aeolian Sedimentation. Water. 2023; 15(3):494. https://doi.org/10.3390/w15030494
Chicago/Turabian StyleKuzmina, Svetlana. 2023. "Wetlands in the Pleistocene Steppe-Tundra Landscapes of Beringia, Their Insects, and the Role of Aeolian Sedimentation" Water 15, no. 3: 494. https://doi.org/10.3390/w15030494
APA StyleKuzmina, S. (2023). Wetlands in the Pleistocene Steppe-Tundra Landscapes of Beringia, Their Insects, and the Role of Aeolian Sedimentation. Water, 15(3), 494. https://doi.org/10.3390/w15030494