
Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment

Department of Hydrobiology and Protection of Ecosystems, University of Life Sciences in Lublin, ul. Dobrzańskiego 37, 20-262 Lublin, Poland
*
Author to whom correspondence should be addressed.
Water 2022, 14(23), 3826; https://doi.org/10.3390/w14233826
Submission received: 2 November 2022
/
Revised: 16 November 2022
/
Accepted: 18 November 2022
/
Published: 24 November 2022
(This article belongs to the Special Issue Monitoring of Freshwater Biodiversity and Water Quality)
Abstract
:The influence of water brownification (an increase in water color) on floating-leaved macrophytes appears to be an important issue in predicting the consequences of climate change in freshwater ecosystems. To fill the gap in the knowledge in this area, we performed a laboratory experiment aiming to test the impact of humic acid (HA) on Lemna minor sampled from an oxbow lake. Plants were exposed over seven days to four different additions of pure HA (resulting in DOC concentrations between 5.0 ± 0.1 mg dm−3 and 36.5 ± 0.4 mg dm−3). Changes in duckweed total fresh biomass, leaf area and the root length were measured, as well as the concentrations of total phosphorus, phosphates and nitrates in the water. The study did not find any significant decrease in either the biomass or leaf area of L. minor under conditions of enhanced HA content. However, we found that a moderate increase in HA in water promoted the growth of duckweed roots, thus, increasing the biomass but not the leaf area. In contrast, a large increase in HA in the water suppressed the growth of roots in L. minor, which was also in coincidence with less nutrient depletion in the water.
1. Introduction
Both observed and projected climatic changes seem to be the most important processes which will affect Earth’s ecosystems in the nearest future as they are comparable in magnitude to the largest global changes of the past 65 million years [1]. Although some biological effects of climate change—such as shifts in species range—are acknowledged [2], long-term ecological effects of a more complex nature are also expected [3,4]. However, some effects of these changes will have a more discrete nature. For example, long-term changes in precipitation frequency and intensity have been reported worldwide since the beginning of the 21st century [5,6,7,8]. It is recognized that precipitation is a major factor affecting the content of dissolved organic carbon (DOC) in freshwaters, including a colored fraction of it (CDOC) [9,10,11,12]. Thus, changes in precipitation patterns may result in a phenomenon which can be commonly named “water browning” or “water brownification”, as the main part of CDOC consists of brown and yellow humic substances [13]. Although changes in water color in freshwaters may be linked to various factors [14,15,16,17], a lot of reports have shown considerable CDOC increase (water browning) in the freshwaters of Northern and Central Europe [18,19,20,21].
It has been broadly demonstrated that CDOC in freshwaters, especially in lakes, plays various considerable roles, influencing a wide range of processes which include, among others, changes in: underwater light climate, pH, contaminant toxicity and nutrient availability (review in [22]). Recent developments in this topic have shown the complex and multifactorial nature of these effects (summarized in [17]). Looking for a more ecosystem-oriented approach, the browning of lakes may generally lead to regime shifts from clear-water, autotrophic, benthic-dominated ecosystems to brown-water, heterotrophic, pelagic-dominated ecosystems. These processes are driven by the synergistic effect of allochthonous DOC promoting heterotrophs as an energy source at the base of planktonic food web and suppressing autotrophs via light extinction and intensified nutrient competition [17,23,24,25]. Enhanced light extinction results in the worsening of underwater light conditions, which, first of all, reduces the abundance of submersed macrophytes [26], which, in consequence, may result in further lake ecosystem deterioration. The effect on floating-leaved macrophytes—such as nympheids and duckweeds—is less probable, as they are not dependent on light penetrating the water column.
It is known, however, that the most favorable conditions for duckweed growth occur at high phosphorus and nitrogen loadings and increasing temperature [27,28,29,30]—two factors related to the impact of climate change on freshwater ecosystems. Duckweeds are able to achieve very fast vegetative reproduction [31,32], which may result in their massive development in small water reservoirs, such as oxbow lakes, temperate ponds, ditches and mid-field water holes [33,34,35]. The creation of floating dense mats by duckweeds causes negative consequences for small ecosystems; the suppression of underwater primary producers (phytoplankton, submersed macrophytes) leads to the development of anoxic conditions and deterioration in the fish and macroinvertebrate communities [36]. What is most important is that this phenomenon may be persistent, and it is suggested that duckweed’s massive development is a self-stabilizing state [37].
Thus, the influence of water brownification on free-floating macrophytes appears to be an important issue in predicting consequences in freshwater ecosystem functioning under the conditions of a changing climate. What is interesting is that, to date, the problem has received scant attention in the research literature. Some data can be found in a study concerning the impact of humic acid on the toxicity of herbicides, where Lemna minor was used as a test organism [38]. To fill this knowledge gap, we performed this study and aimed to answer the basic question—whether water brownification impacts in any way duckweed population in small freshwater lakes. To verify this, we performed a laboratory experiment in which we tested the impact of pure humic acid on Lemna minor plants sampled from a small oxbow lake.
2. Material and Methods
Lemna minor plants were sampled in the middle of June from a small and shallow oxbow lake situated in the floodplain of the Bystrzyca River in Eastern Poland (Lublin Upland, European Plain, 51°16′23″ E; 22°37′51″ E). Then, the plants were flushed with tap water and, afterwards, stored for acclimatization for five days in the water sampled from the same oxbow lake but filtered through a planktonic net of mesh size 25 µm to remove planktonic organisms. During the acclimatization, the plants were illuminated with fluorescent lighting with a color temperature of 6500 K (daylight), which gave an irradiance of 45 μmol photons m−2 s−1 12 h day−1 with a day/night cycle of 16:8 (which corresponds to the length of the day in Central Poland in June). The culture was kept at 24 °C during the day and 22 °C during the night (which corresponds to temperatures occurring in June/July in the eastern part of Poland). The influence of humic acid (HA) on duckweed was tested in 20 glass cylinders with a capacity of ca. 0.5 L (internal diameter 8 cm) placed in the growing cabinet for eight days in the same conditions as during plant acclimatization.
The same amount of oxbow lake water (0.4 dm−3) was poured into each tank after being filtrated through a plankton net (25 μm). Four cylinders served as controls (marked as c). The other 16 served as treatments with the addition of humic acid (Sigma Aldrich, USA) in four various amounts; thus, each experimental cylinder with HA addition had four replications (Figure S1). The substance was first dissolved in water by adding 1.5 g of pure HA powder to 300 mL of lake water (previously filtered by a GF/C fiber glass filter), then homogenizing in an ultrasound homogenizer (20 min at 200 W and 45 kHz) and filtered through a paper filter under vacuum thereafter. The obtained solution had a concentration of total organic carbon (TOC) of 330 mg dm−3. Various amounts of this solution were afterwards added to treatment cylinders: 10 mL, 35 mL, 80 mL and 170 mL (marked as h1, h2, h3 and h4, respectively).
A similar number (mean 0.085 ± 0.002 g FW; for details, see Table 1) of healthy-looking duckweed plants were placed into all cylinders. Before putting the plants into them, 50 specimens were randomly chosen and the length of their roots measured (mean 12.65 ± 1.74 mm). After putting the Lemna fronds into the tanks, each jar was photographed with the use of a digital camera against the background of a millimeter paper sheet, which was further used to measure the leaf surface area with the use of Easy Leaf Area software [39]. The same procedure was performed on the last day of the experiment. After this, all fronds were removed from the jars, and the length of the roots of ten randomly chosen specimens from each jar was measured. Then, they were dried on filter paper and weighed to obtain the fresh weight (mg FW).
The physical and chemical water parameters conductivity and pH were measured using the YSI 556 MPS electrode. Total organic carbon (TOC) was determined using a PASTEL UV analyzer. Phosphorus compounds (TP and P-PO4) were measured using a spectrophotometer UV-1610 Schimadzu (with molybdate method after mineralization with a mixture of K2S2O8 and H2SO4) [40]. Nitrate concentration was determined with the use of an FIA analyzer (photometric via reduction to nitrite and azo dye formation) (flow analysis method according to [41]).
The effect of water brownification (TOC concentration) on fresh biomass, leaf surface area and length of roots of L. minor was verified using analysis of variance (ANOVA). Post hoc Duncan’s test was used to compare significant (p < 0.05) differences between means. Additionally, ANOVA was used to compare concentrations of TP, P-PO4 and N-NO3 between experimental treatments. The analysis was performed using STATISTICA 13.3 software.
3. Results
The water sampled from the oxbow lake, which was used in the experiment at the beginning (D01), had moderate mineralization (EC = 364 μS cm−1), low TOC content (4.0 ± 0.1 mg dm−3) and slightly alkaline reaction (pH 8.7). After the addition of various amounts of HA solution to the experimental cylinders (h1–h4), TOC concentrations were increased to 5.0 ± 0.1 mg dm−3, 8.8 ± 0.3 mg dm−3, 21.6 ± 0.4 mg dm−3 and 36.5 ± 0.4 mg dm−3, respectively. Control treatments, in which water from the oxbow lake was poured, had a concentration of TOC as mentioned above. After seven days of the experiment, water mineralization, as well as pH, remained on a similar level (Table 2.). In the case of TOC concentration, an increase in the mean values was observed between the start and the end of the experiment. Differences in the final TOC concentration in the treatments within replications were rather low—except for two of them (h3 and h4, standard deviation 5.0 mg dm−3 and 9.8 mg dm−3, respectively).
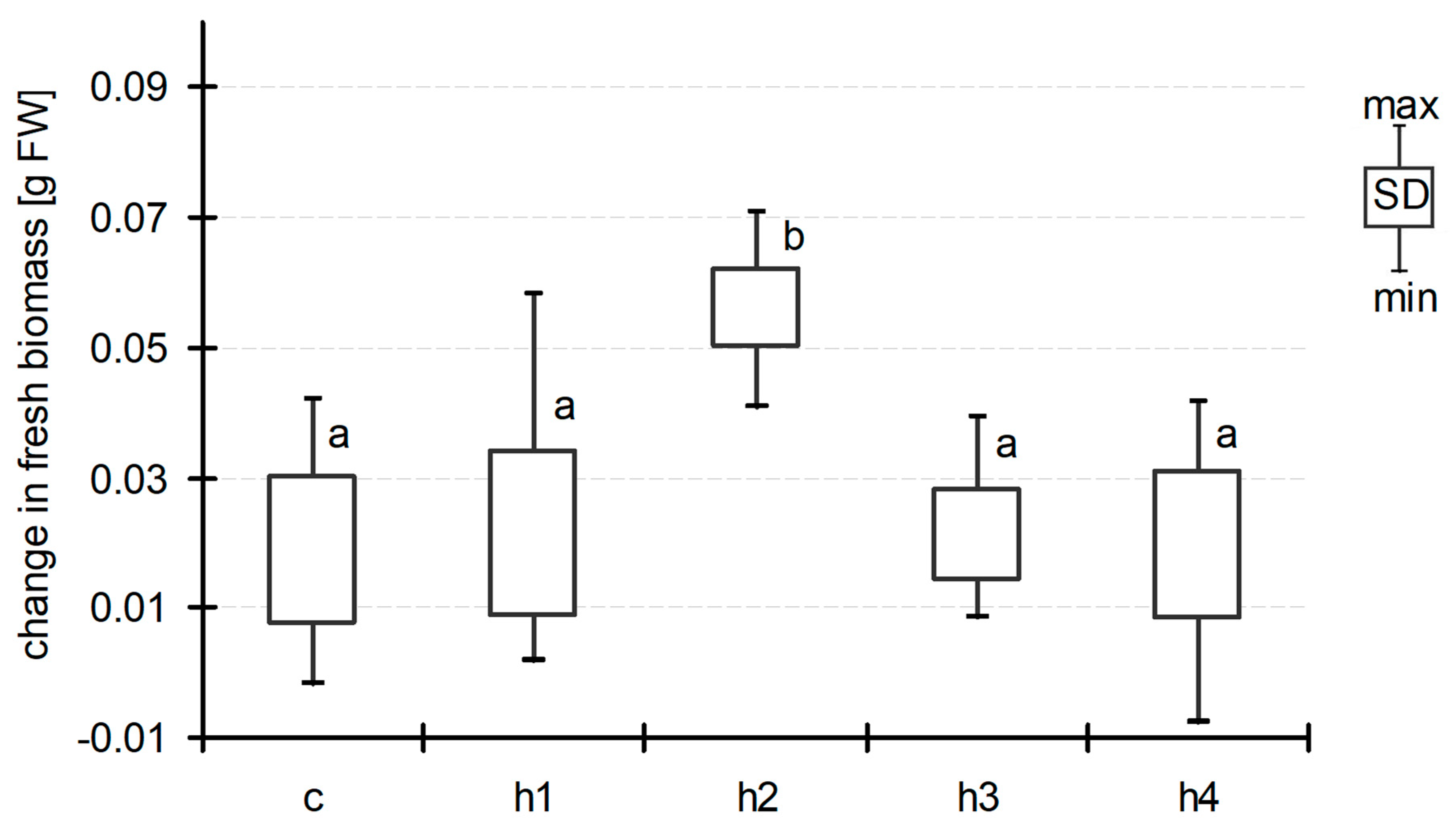
Generally, we observed an increase in Lemna biomass during exposition in all treatments including the control ones. The starting biomass in all treatments and the control was very similar and ranged between 0.085 g and 0.086 g FW per jar (on average). At the end of experiment, in the control and treatments h1, h3 and h4, the increase in frond biomass was about 0.02 g per 7 days (0.018 ± 0.022, 0.021 ± 0.025, 0.021 ± 0.013 and 0.019 ± 0.022, respectively, Figure 1), suggesting that humic acid addition had no impact on Lemna biomass. In the h2 treatment (TOC concentration 21.6 ± 0.4 mg dm−3 at D0), the biomass increase was at least twice as great and amounted to 0.056 ± 0.013 g per 7 days. Analysis of variance did not show significant differences between treatments at D07, although post hoc Duncan’s test demonstrated that the mean biomass of L. minor in h2 varied significantly from that in all the other treatments (Figure 1). Humic acid addition also affected the increase in leaf area in all treatments (0.8–0.9 cm2 per 7 days as compared to control jars) with the exception of h3, but the observed differences were not significant (ANOVA at p < 0.05) (Figure 2). A more explicit reaction to the HA addition after seven days of exposition was seen in the case of Lemna root lengths (Figure 3). We did not observe differences between that of h1 and control jars (22.5 ± 3.6 mm and 21.6 ± 7.9 mm, respectively). In the h2 treatment, the length of the roots increased (31.0 ± 2.2 mm), while, in the h3 and h4 treatments with the highest addition of HA, one could observe a decrease in this parameter (18.4 ± 2.9 mm and 14.0 ± 2.3 mm). The differences were statistically significant (ANOVA, F = 57.05, p < 0.001).
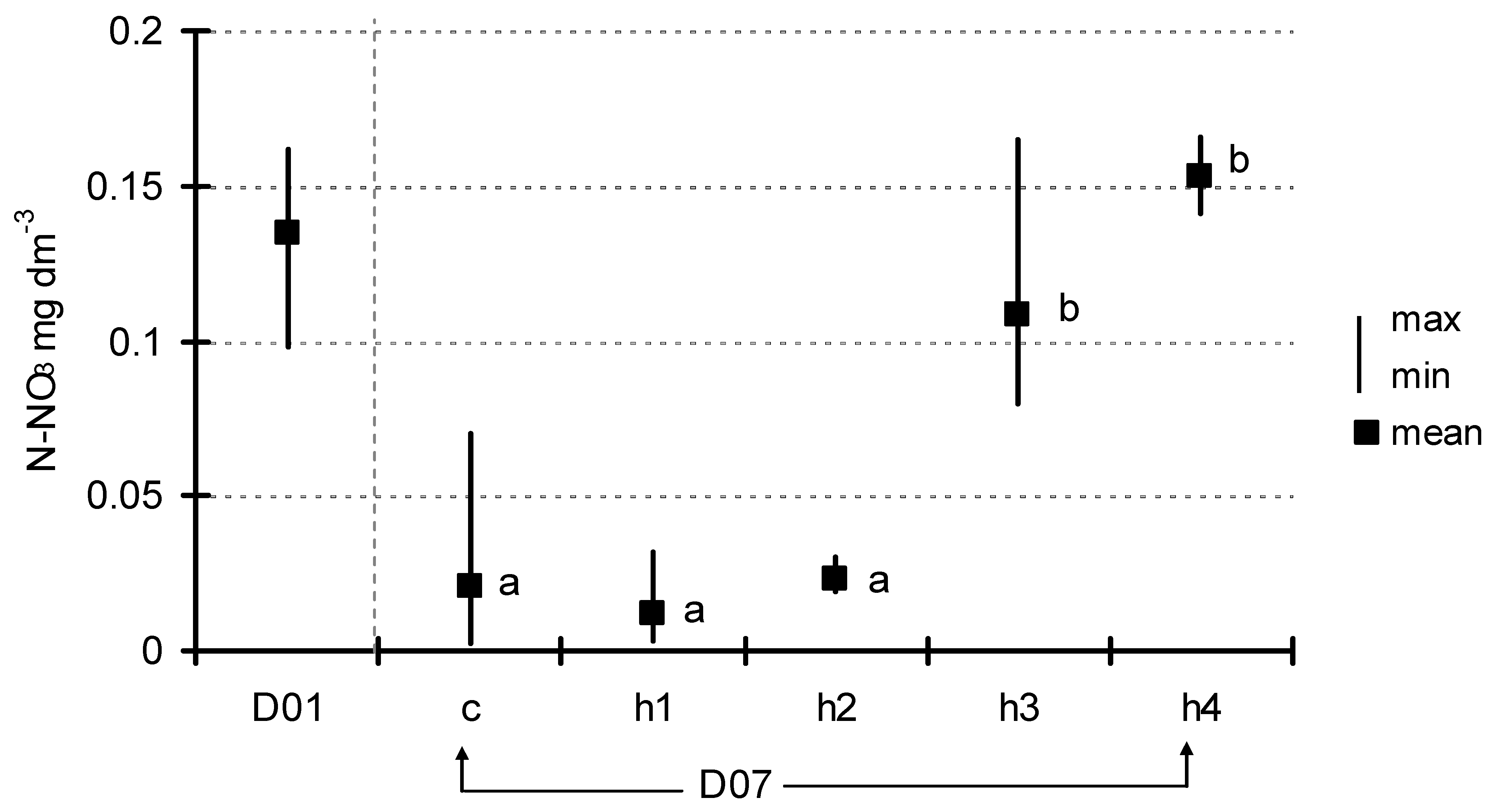
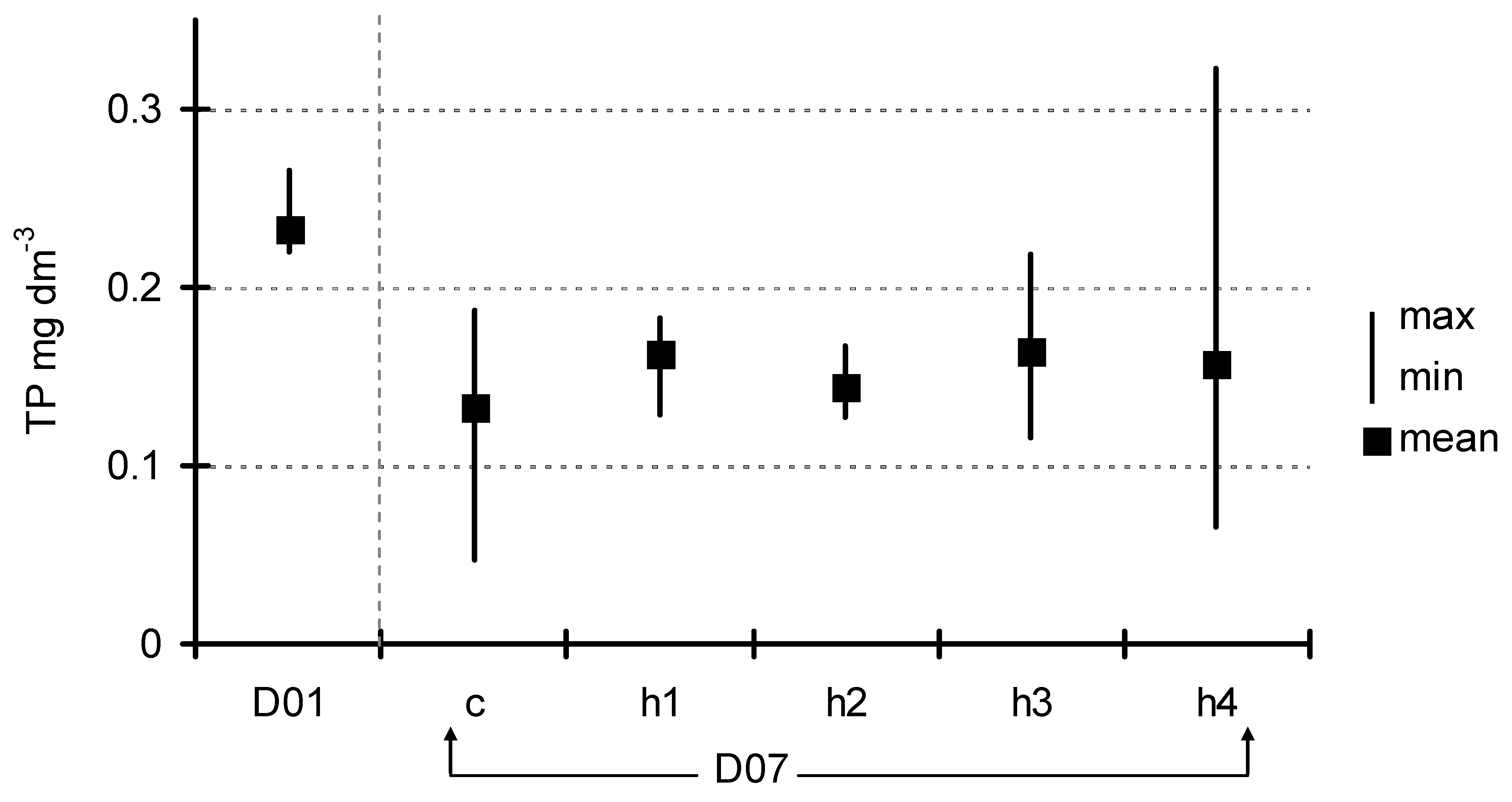
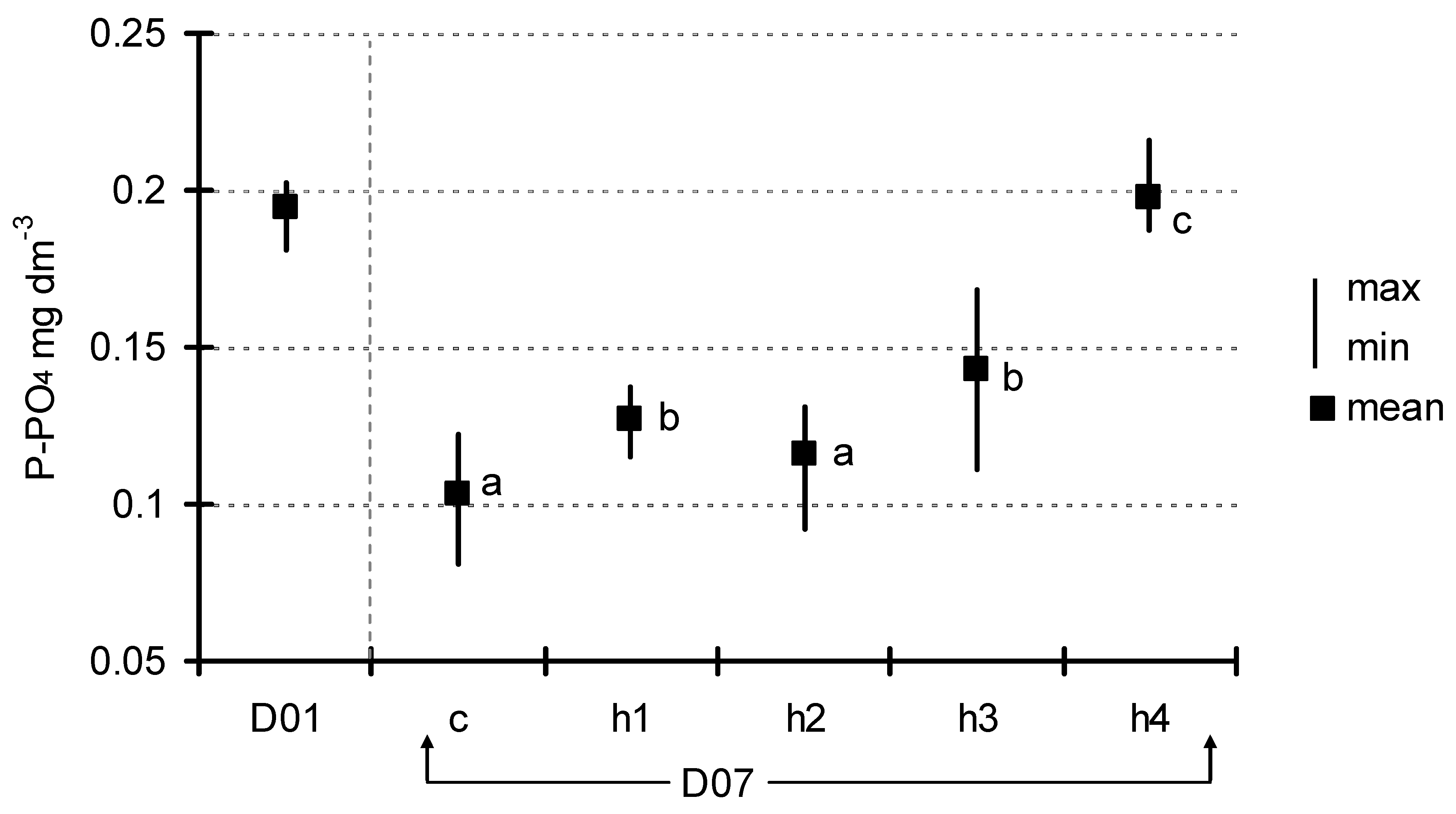
After seven days of exposition, the concentration of basic nutrients in the water below the floating plant cover was also measured. The concentration of total phosphorus (Figure 4) decreased from 0.23 ± 0.037 mg dm−3 (D01) to 0.13 ± 0.06–0.16 ± 0.05 mg dm−3 (D07) in the control and the treatments, but these differences were not significant (ANOVA at p < 0.05). A decrease in the concentration after seven days of exposition was generally observed in phosphates; however, one could observe a more diversified response to the HA addition (Figure 5). Analysis of variance showed significant differences (F = 9.85, p < 0.001) which appeared among three groups of experimental jars: c/h2, h1/h3 and h4. The most visible decrease in P-PO4 concentration was observed in the control as well as in the h2 treatments (from 0.19 ± 0.009 mg dm−3 at the start to 0.10 ± 0.02–0.11 ± 0.02 mg dm−3). The second group (h1 and h3 treatments) showed a smaller level of decrease (to the range of 0.12 ± 0.01–0.14 ± 0.02 mg dm−3), while, in h4 (the most HA rich), the concentration of phosphates did not change significantly over the course of the experiment (0.19 ± 0.01 mg dm−3 at the end). The trend in N-NO3 concentration was somewhat similar (Figure 6). Nitrates content was almost unchanged in the h3 and h4 treatments, while an almost twofold decrease was observed in the c, h1 and h2 treatments (from 0.135 ± 0.02 mg dm−3 at D01 to the range of 0.011 ± 0.01–0.022 ± 0.004 mg dm−3 at D07). The difference between the first-mentioned group (h3/h4) and the other three treatments (c/h1/h2) was strongly statistically significant (ANOVA, F = 13.01, p < 0.001).
4. Discussion
Freshwater brownification (increase in water color) is a phenomenon not yet sufficiently understood, but it is usually linked with climate change. The increase in temperature, enhanced biodegradation of soil organic matter and changes in precipitation (droughts and floods occurring more frequently) all lead to the increased production and export of dissolved organic matter from the catchments to surface waters [42]. The main part of dissolved organic matter (60–90%) consists of colorful humic acids [43,44]; thus, an increased load of these substances leads to the colorization of freshwaters. One of the consequences is the deterioration of underwater light climate, which is expected to affect submersed macrophytes [26]. However, this effect is not applied to floating-leaved macrophytes (such as duckweeds) as they are not limited by light penetrating the water column. Thus, in this study, we posed the question as to whether water brownification affects the growth parameters of small duckweed from the genus Lemna in any other way.
Boavida and Wetzel [45] assessed the average DOC content of freshwaters worldwide to be 6.5 mg dm−3. More recent studies from the northern hemisphere (over 7500 studied lakes) reported that the median DOC concentration was 5.71 mg dm−3, ranging from 0.1 mg dm−3 to 332 mg dm−3, while the mean concentration was 7.58 ± 0.19 mg dm−3. However, in 87% of studied lakes, the DOC was between 1 and 20 mg dm−3. Lakes between 20 and 40 mg dm−3 were few (4.2%), and only 0.4% of the lakes had DOC concentrations above 40 mg dm−3 [46]. Thus, it can be stated that the concentration of dissolved organic matter in our experimental cylinders was environmentally relevant (especially in the h1, h2 and h3 treatments where DOC concentrations at D01 were 5.0 ± 0.1 mg dm−3, 8.8 ± 0.3 mg dm−3 and 21.6 ± 0.4 mg dm−3, respectively). Nevertheless, the fourth treatment, with the highest concentration (D01: 36.5 ± 0.4 mg dm−3) was also kept in the natural range.
In compliance with expectations, the study did not find any significant decrease in either the biomass or leaf area of L. minor under conditions of enhanced HA content. It is in agreement with the report dealing with the impact of humic acid on the toxicity of herbicides, where Lemna minor was used as a test organism [38]. Any changes in both duckweed fresh biomass and number of fronds after HA addition were noted in that study, but they used very low concentrations of HA in the experiment (2 mg dm−3). However, our results show a few other interesting findings. First, humic acid addition increased the biomass of L. minor at moderate concentrations (8.8 ± 0.3 −10.1 ± 0.3 mg DOC dm−3). Second, there was no reaction concerning the leaf area of Lemna. Third, a significant increase in root length was noted with the use of moderate DOC concentration (the same as mentioned above). Moreover, some inhibiting effects in the growth of roots were seen in the case of the highest HA addition (36.5 ± 0.4–38.0 ± 9.8 mg dm−3). The same was also observed in the changes of phosphates and nitrates contents in the water. The highest HA addition did not cause the depletion of these nutrients as was shown in the other treatments.
Taking all findings together, we conclude that:
a. A moderate increase in natural-colored, dissolved organic compounds in water (in the form of HA) promotes the growth of duckweed roots, thus, increasing the biomass but not the leaf area;
b. A large increase in HA in water may suppress the growth of roots in L. minor, which is also in coincidence with less nutrient depletion in the water.
The mechanisms by which DOC may impact macrophyte communities in freshwaters (in addition to the deterioration of light conditions) are manifold, although far from being clearly understood [42,47]. Several studies showed indirect effects related to the modulations of the bioavailability of nutrients, including the binding of orthophosphates or inhibition of phosphatase activity in lake water [45,48]. The binding capacity of HA with heavy metals, such as copper or cadmium, is also favorable for plants—it protects macrophytes against bioaccumulation of this elements in their tissues, thus, preventing negative effects in the form of lowering chlorophyll content [49].
It was hypothesized that HA acts in rather an indirect way due to its large molecular mass being too big to penetrate bio-membranes [50]. However, some empirical studies have shown that HA may be taken up by plants [51]. Direct influence on some macrophytes was evidenced, especially on hornwort (Ceratophyllum demersum). The exposure of the species fronds revealed that fulvic acids directly interfere within photosynthetic oxygen production and electron transport [50]. Furthermore, the addition of different humic acids strongly increased the guaiacol peroxidase activity in C. demersum [52]. Humic acid uptake to plant cells can also cause the formation of reactive oxygen species, which, in consequence, may damage the cells, as was demonstrated in charophyte Nitellopsis obtusa after 24 h exposition under a very low concentration of DOC (1.1 mg dm−3) [53].
Our major finding, however, is an unexpected “positive” effect of moderate HA concentration: enhanced growth of Lemna roots. Duckweeds extend their root length in response to phosphate starvation [54], although there have also been contradictory reports [55]. Additionally, nitrate deficiency is supposed to promote roots elongation [56]. Decreased nutrient uptake by roots and their elongation might occur when rich epiphytic communities develop within the macrophyte rhisosphere [57]. Unexpected enhanced growth of plant roots was also observed after a seven-day exposition of L. minor on alumina nanoparticles. The proposed mechanism of action was “stress-induced morphogenic response” (SIMR), where a key role is played by reactive oxygen species and the phytohormone auxin [58]. A similar mechanism, but concerning humic acids, was proposed by Steinberg et al. [59], who suggested that mild HA stresses may be beneficial to specific plant organs due to the increased expression of anti-stress genes, resulting in multi-stress resistance (cited from [42]).
Modulations of nutrient bioavailability do not explain the enhanced root growth of L. minor under moderate HA addition. We observed phosphate and nitrate shortages in both the control and HA treatments. The inhibiting effect of the highest HA concentration on root growth is hard to clarify as well and may be conditioned by the complicated and unrecognized nature of DOC and freshwater interactions. The relationships between the DOC in freshwaters and its controlling variables are often nonlinear as these relationships potentially involve many interactions [60]. It can probably be seen in the nonlinear reactions of lake ecosystems or organisms to increased DOC loading. For example, a study by Alles (1996) showed that DOC concentrations ranging from 0 to 2 mg dm−3 lowered the species diversity of diatoms. In medium concentrations (3–10 mg dm−3), this effect was reduced, but, in higher concentrations (above 10 mg dm−3), the effect appeared again [61].
Our results confirmed the need for further studies concerning DOC–freshwater interactions on the level of biota response to enhanced water brownification. Studies on DOC impact on duckweeds should focus on: a. a qualitative and quantitative analysis of epiphytic communities which usually develop on exposed roots; b. their role in nutrient bioavailability for plants; c. uptake of humic acids by plant tissues; and d. the role of humic acids in growth regulation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14233826/s1, Figure S1: Experimental cylinders with Lemna minor after seven-day exposition (from left to right: control [c], h1, h2, h3 and h4 treatments).
Author Contributions
Conceptualization, W.P., M.T.-K. and M.N.; Methodology, W.P., M.T.-K. and M.N.; Software, W.P.; Statistical Analysis, M.T.-K.; Investigation, W.P., M.T.-K. and M.N.; Writing—Original Draft Preparation, W.P.; Writing—Review and Editing, M.T.-K. and M.N., Visualization, W.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Diffenbaugh, N.S.; Singh, D.; Mankin, J.S. Unprecedented climate events: Historical changes, aspirational targets, and national commitments. Sci. Adv. 2018, 4, eaao3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Falconi, L. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Pe’er, G.; Singer, A.; Gonzalez, A. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, D.P.; Gianoli, E.; Morris, W.F.; Bozinovic, F. Ecological and evolutionary impacts of changing climatic variability. Biol. Rev. 2017, 92, 22–42. [Google Scholar] [CrossRef] [Green Version]
- Kunkel, K.E. North American trends in extreme precipitation. Nat. Hazards 2003, 29, 291–305. [Google Scholar] [CrossRef]
- Gallego, M.C.; García, J.A.; Vaquero, J.M.; Mateos, V.L. Changes in frequency and intensity of daily precipitation over the Iberian Peninsula. J. Geophys. Res. Atmos. 2006, 111, D24105. [Google Scholar] [CrossRef]
- Chou, C.; Chen, C.A.; Tan, P.H.; Chen, K.T. Mechanisms for global warming impacts on precipitation frequency and intensity. J. Clim. 2012, 25, 3291–3306. [Google Scholar] [CrossRef]
- Ban, N.; Schmidli, J.; Schär, C. Heavy precipitation in a changing climate: Does short-term summer precipitation increase faster? Geophys. Res. Lett. 2015, 42, 1165–1172. [Google Scholar] [CrossRef]
- Hongve, D.; Riise, G.; Kristiansen, J.F. Increased colour and organic acid concentrations in Norwegian forest lakes and drinking water–a result of increased precipitation? Aquat. Sci. 2004, 66, 231–238. [Google Scholar] [CrossRef]
- Weyhenmeyer, G.A.; Willén, E.; Sonesten, L. Effects of an extreme precipitation event on water chemistry and phytoplankton in the Swedish Lake Mälaren. Boreal Environ. Res. 2004, 9, 409–420. [Google Scholar]
- Arvola, L.; Rask, M.; Ruuhijärvi, J.; Tulonen, T.; Vuorenmaa, J.; Ruoho-Airola, T.; Tulonen, J. Long-term patterns in pH and colour in small acidic boreal lakes of varying hydrological and landscape settings. Biogeochemistry 2010, 101, 269–279. [Google Scholar] [CrossRef]
- Pęczuła, W. Variability of coloured dissolved organic carbon in seepage humic lakes as related to lake morphometry, catchment characteristics and precipitation. Ecohydrology 2015, 8, 1229–1238. [Google Scholar] [CrossRef]
- de Wit, H.A.; Valinia, S.; Weyhenmeyer, G.A.; Futter, M.N.; Kortelainen, P.; Austnes, K.; Vuorenmaa, J. Current browning of surface waters will be further promoted by wetter climate. Environ. Sci. Technol. Lett. 2016, 3, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Delpla, I.; Jung, A.V.; Baures, E.; Clement, M.; Thomas, O. Impacts of climate change on surface water quality in relation to drinking water production. Environ. Int. 2009, 35, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.D.; Freeman, C.; Cork, L.G.; Thomas, D.N.; Reynolds, B.; Billett, M.F.; Garnett, M.H.; Norris, D. Evidence against recent climate-induced destabilisation of soil carbon from 14C analysis of riverine dissolved organic matter. Geophys. Res. Lett. 2007, 34, L07407. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.D.; Jones, T.G.; Burden, A.; Ostle, N.; Zieliński, P.; Cooper, M.D.; Peacock, M.; Clark, J.M.; Oulehle, F.; Cooper, D.; et al. Acidity controls on dissolved organic carbon mobility in organic soils. Glob. Chang. Biol. 2012, 18, 3317–3331. [Google Scholar] [CrossRef] [Green Version]
- Creed, I.F.; Bergström, A.K.; Trick, C.G.; Grimm, N.B.; Hessen, D.O.; Karlsson, J.; Senar, O.E. Global change-driven effects on dissolved organic matter composition: Implications for food webs of northern lakes. Glob. Chang. Biol. 2018, 24, 3692–3714. [Google Scholar] [CrossRef]
- Evans, C.D.; Monteith, D.T.; Cooper, D.M. Long-term increases in surface water dissolved organic carbon, Observations, possible causes and environmental impacts. Environ. Pollut. 2005, 137, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Monteith, D.T.; Stoddard, J.L.; Evans, C.D.; de Wit, H.A.; Forsius, M.; Hogasen, T.; Wilander, A.; Skjelkvale, B.L.; Jeffries, D.S.; Vuorenmaa, J.; et al. Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 2007, 450, 537–540. [Google Scholar] [CrossRef]
- Worrall, F.; Harriman, R.; Evans, C.D.; Watts, C.D.; Adamson, J.; Neal, C.; Stevens, P. Trends in dissolved organic carbon in UK rivers and lakes. Biogeochemistry 2004, 70, 369–402. [Google Scholar] [CrossRef]
- Hejzlar, J.; Dubrovský, M.; Buchtele, J.; Růžička, M. The apparent and potential effects of climate change on the inferred concentration of dissolved organic matter in a temperate stream (the Malše River, South Bohemia). Sci. Total Environ. 2003, 310, 143–152. [Google Scholar] [CrossRef]
- Williamson, C.E.; Morris, D.P.; Pace, M.L.; Olson, O.G. Dissolved organic carbon and nutrients as regulators of lake ecosystems, Resurrection of a more integrated paradigm. Limnol. Oceanogr. 1999, 44, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Hessen, D.O.; Andersen, T.; Lyehe, A. Carbon metabolism in a humic lake: Pool sires and cycling through zooplankton. Limnol. Oceanogr. 1990, 35, 84–99. [Google Scholar] [CrossRef]
- Ask, J.; Karlsson, J.; Persson, L.; Ask, P.; Byström, P.; Jansson, M. Terrestrial organic matter and light penetration: Effects on bacterial and primary production in lakes. Limnol. Oceanogr. 2009, 54, 2034–2040. [Google Scholar] [CrossRef]
- Solomon, C.T.; Jones, S.E.; Weidel, B.C.; Buffam, I.; Fork, M.L.; Karlsson, J.; Saros, J.E. Ecosystem consequences of changing inputs of terrestrial dissolved organic matter to lakes: Current knowledge and future challenges. Ecosystems 2015, 18, 376–389. [Google Scholar] [CrossRef]
- Sřndergaard, M.; Phillips, G.; Hellsten, S.; Kolada, A.; Ecke, F.; Mäemets, H.; Oggioni, A. Maximum growing depth of submerged macrophytes in European lakes. Hydrobiologia 2013, 704, 165–177. [Google Scholar] [CrossRef]
- Landolt, E.; Kandeler, R. Phytochemistry, Physiology, Application, Bibliography. In The Family of Lemnaceae—A Monographic Study; Veroeffentlichungen des Geobotanischen Instituts der ETH Stiftung Ruebel: Zürich, Switzerland, 1988; Volume 2. [Google Scholar]
- Portielje, R.; Roijackers, R.M.M. Primary succession of aquatic macrophytes in experimental ditches in relation to nutrient input. Aquat. Bot. 1995, 50, 127–140. [Google Scholar] [CrossRef]
- Szabó, S.; Roijackers, R.; Scheffer, M.; Borics, G. The strength of limiting factors for duckweed during algal competition. Arch. Hydrobiol. 2005, 164, 127–140. [Google Scholar] [CrossRef]
- Kufel, L.; Strzałek, M.; Konieczna, A.; Izdebska, K. The effect of Stratiotes aloides L. and nutrients on the growth rate of Lemna minor L. Aquat. Bot. 2010, 92, 168–172. [Google Scholar] [CrossRef]
- Vermaat, J.E.; Hanif, M.K. Performance of common duckweed species (Lemnaceae) and the waterfern Azolla filiculoides on different types of waste water. Water Res. 1998, 32, 2569–2576. [Google Scholar] [CrossRef]
- Frederic, M.; Samir, L.; Louise, M.; Abdelkrim, A. Comprehensive modeling of mat density effect on duckweed (Lemna minor) growth under controlled eutrophication. Water Res. 2006, 40, 2901–2910. [Google Scholar] [CrossRef] [PubMed]
- Mehra, A.; Farago, M.E.; Banerjee, D.K. A study of eichhornia crassipes growing in the overbank and floodplain soils of the River Yamuna in Delhi, India. Environ. Monit. Assess. 2000, 60, 25–45. [Google Scholar] [CrossRef]
- Brendonck, L.; Maes, J.; Rommens, W.; Dekeza, N.; Nhiwatiwa, T.; Barson, M.; Marshall, B. The impact of water hyacinth (Eichhornia crassipes) in a eutrophic subtropical impoundment (Lake Chivero, Zimbabwe). II. Species diversity. Arch. Hydrobiol. 2003, 158, 389–405. [Google Scholar] [CrossRef]
- Kufel, L.; Strzałek, M.; Przetakiewicz, A. Plant response to overcrowding–Lemna minor example. Acta Oecol. 2018, 91, 73–80. [Google Scholar] [CrossRef]
- Driever, S.M.; van Nes, E.H.; Roijackers, R.M. Growth limitation of Lemna minor due to high plant density. Aquat. Bot. 2005, 81, 245–251. [Google Scholar] [CrossRef]
- Scheffer, M.; van Nes, E. Shallow lakes theory revisited: Various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 2007, 584, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Mihajlović, V.; Tomić, T.; Tubić, A.; Molnar Jazić, J.; Ivančev Tumbas, I.; Šunjka, D.; Teodorović, I. The impact of humic acid on toxicity of individual herbicides and their mixtures to aquatic macrophytes. Environ. Sci. Pollut. Res. 2019, 26, 23571–23582. [Google Scholar] [CrossRef]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated digital image analysis for rapid and accurate measurement of leaf area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef]
- PN-EN ISO 6878; Jakość Wody—Oznaczanie Fosforu—Metoda Spektrometryczna z Molibdenianem Amonu. Polski Komitet Normalizacyjny: Warszawa, Poland, 2006.
- DIN EN ISO 13 395; Water Quality—Determination of Nitrite Nitrogen and Nitrate Nitrogen and the Sum of both by Flow Analysis (CFA and FIA) and Spectrometric Detection. Beuth Verlag GmbH: Berlin, Germany, 1996.
- Reitsema, R.E.; Meire, P.; Schoelynck, J. The future of freshwater macrophytes in a changing world: Dissolved organic carbon quantity and quality and its interactions with macrophytes. Front. Plant Sci. 2018, 9, 629. [Google Scholar] [CrossRef]
- Wetzel, R.G. Detrital dissolved and particulate organic carbon functions in aquatic ecosystems. Bull. Mar. Sci. 1984, 35, 503–509. [Google Scholar]
- Sachse, A.; Henrion, R.; Gelbrecht, J.; Steinberg, C.E.W. Classification of dissolved organic carbon (DOC) in river systems: Influence of catchment characteristics and autochthonous processes. Org. Geochem. 2005, 36, 923–935. [Google Scholar] [CrossRef]
- Boavida, M.J.; Wetzel, R.G. Inhibition of phosphatase activity by dissolved humic substances and hydrolytic reactivation by natural ultraviolet light. Freshw. Biol. 1998, 40, 285–293. [Google Scholar] [CrossRef]
- Sobek, S.; Tranvik, L.J.; Prairie, Y.T.; Kortelainen, P.; Cole, J.J. Patterns and regulation of dissolved organic carbon: An analysis of 7,500 widely distributed lakes. Limnol. Oceanogr. 2007, 52, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Pflugmacher, S.; Tidwell, L.F.; Steinberg, C.E. Dissolved humic substances can directly affect freshwater organisms. Acta Hydrochim. Hydrobiol. 2001, 29, 34–40. [Google Scholar] [CrossRef]
- Wetzel, R.G. Gradient-dominated ecosystems: Sources and regulatory functions of dissolved organic matter in freshwater ecosystems. Hydrobiologia 1992, 229, 181–198. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Z.; Cheng, S.; Wu, Z. Influence of humic acids on the accumulation of copper and cadmium in Vallisneria spiralis L. from sediment. Environ. Earth Sci. 2010, 61, 1207–1213. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Pietsch, C.; Rieger, W.; Steinberg, C.E. Dissolved natural organic matter (NOM) impacts photosynthetic oxygen production and electron transport in coontail Ceratophyllum demersum. Sci. Total Environ. 2006, 357, 169–175. [Google Scholar] [CrossRef]
- Steinberg, C.E.; Kamara, S.; Prokhotskaya, V.Y.; Manusadžianas, L.; Karasyova, T.A.; Timofeyev, M.A.; Menzel, R. Dissolved humic substances–ecological driving forces from the individual to the ecosystem level? Freshw. Biol. 2006, 51, 1189–1210. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Spangenberg, M.; Steinberg, C.E.W. Dissolved Organic Matter (DOM) and effects on the aquatic macrophyte Ceratophyllum demersum in relation to photosynthesis, pigmentpattern and activity of detoxication enzymes. J. Appl. Bot. 1999, 73, 184–190. [Google Scholar]
- Grigutytė, R.; Nimptsch, J.; Manusadžianas, L.; Pflugmacher, S. Response of oxidative stress enzymes in charophyte Nitellopsis obtusa exposed to allochthonous leaf extracts from beech Fagus sylvatica. Biologija 2009, 55, 3–4. [Google Scholar] [CrossRef]
- Reid, M.S.; Bieleski, R.L. Response of Spirodela oligorrhiza to phosphorus deficiency. Plant Physiol. 1970, 46, 609–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Bucio, J.; Hernández-Abreu, E.; Sánchez-Calderón, L.; Nieto-Jacobo, M.F.; Simpson, J.; Herrera-Estrella, L. Phosphate availability alters architecture and causes changes in hormone sensitivity in the Arabidopsis root system. Plant Physiol. 2002, 129, 244–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linkohr, B.I.; Williamson, L.C.; Fitter, A.H.; Leyser, H.M.O. Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. Plant J. 2002, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Roijackers, R.; Szabo, S.; Scheffer, M. Experimental analysis of the competition between algae and duckweed. Arch. Hydrobiol. 2004, 160, 401–412. [Google Scholar] [CrossRef]
- Juhel, G.; Batisse, E.; Hugues, Q.; Daly, D.; van Pelt, F.N.; O’Halloran, J.; Jansen, M.A. Alumina nanoparticles enhance growth of Lemna minor. Aquat. Toxicol. 2011, 105, 328–336. [Google Scholar] [CrossRef]
- Steinberg, C.E.W.; Meinelt, T.; Timofeyev, M.A.; Bittner, M.; Menzel, R. Humic substances (review series). Part 2: Interactions with Organisms. Environ. Sci. Pollut. Res. 2008, 15, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Toming, K.; Kotta, J.; Uuemaa, E.; Sobek, S.; Kutser, T.; Tranvik, L.J. Predicting lake dissolved organic carbon at a global scale. Sci. Rep. 2020, 10, 8471. [Google Scholar] [CrossRef]
- Alles, E. Fließgewässerversauerung im Schwarzwald. In Ökologische Bewertung Auf Der Basis Benthischer Diatomeen; Oberirdische Gewässer, Gewässerökologie (Vol. 51); Landesanstalt für Umweltschutz Baden-Württemberg: Karlsruhe, Germany, 1996. [Google Scholar]
Figure 1.
Change in biomass of Lemna minor between D01 and D07 (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).
Figure 1.
Change in biomass of Lemna minor between D01 and D07 (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).
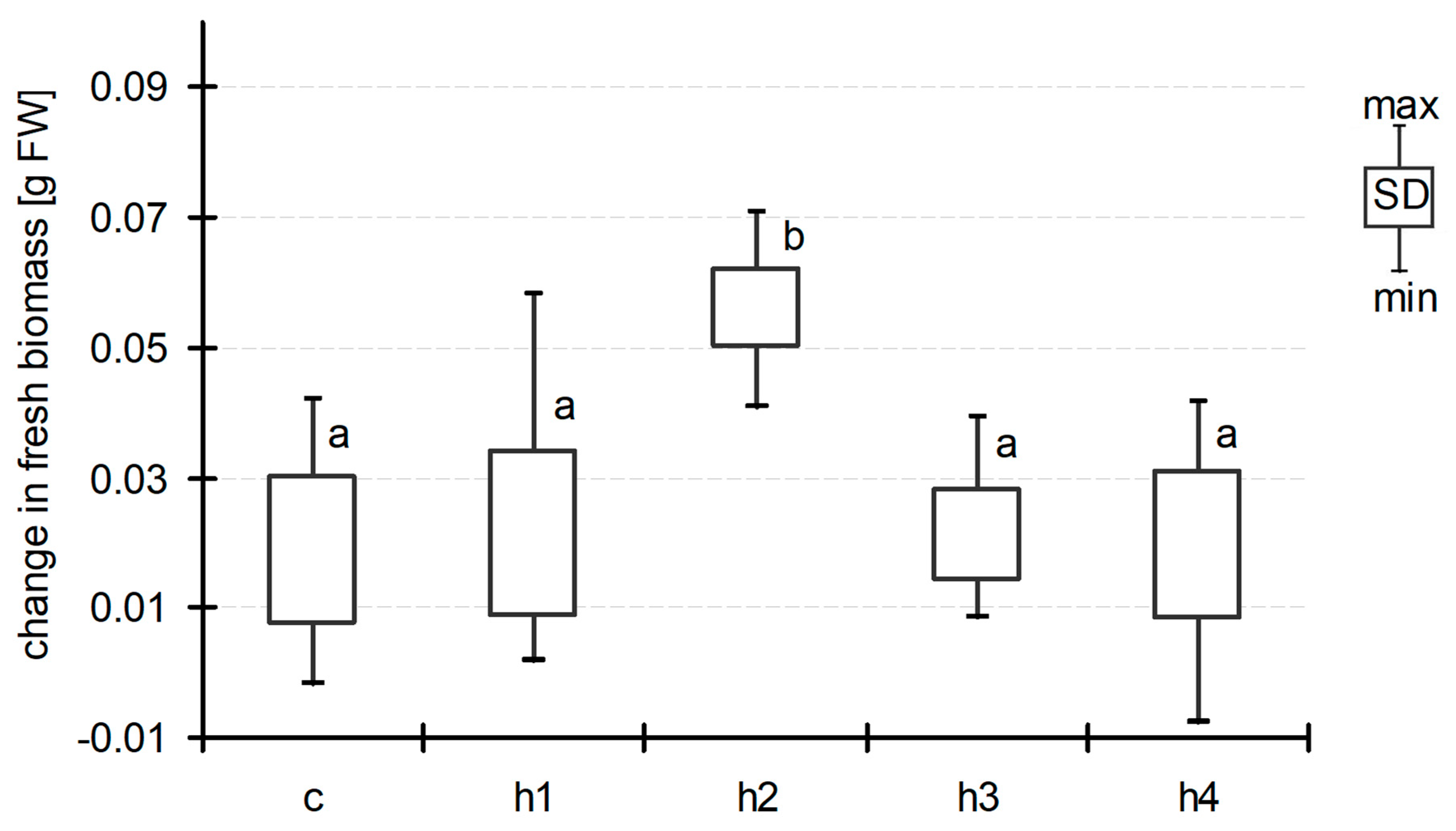
Figure 2.
Change in leaf area of Lemna minor between D01 and D07.

Figure 3.
The length of roots of Lemna minor at the end of the experiment (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).
Figure 3.
The length of roots of Lemna minor at the end of the experiment (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).
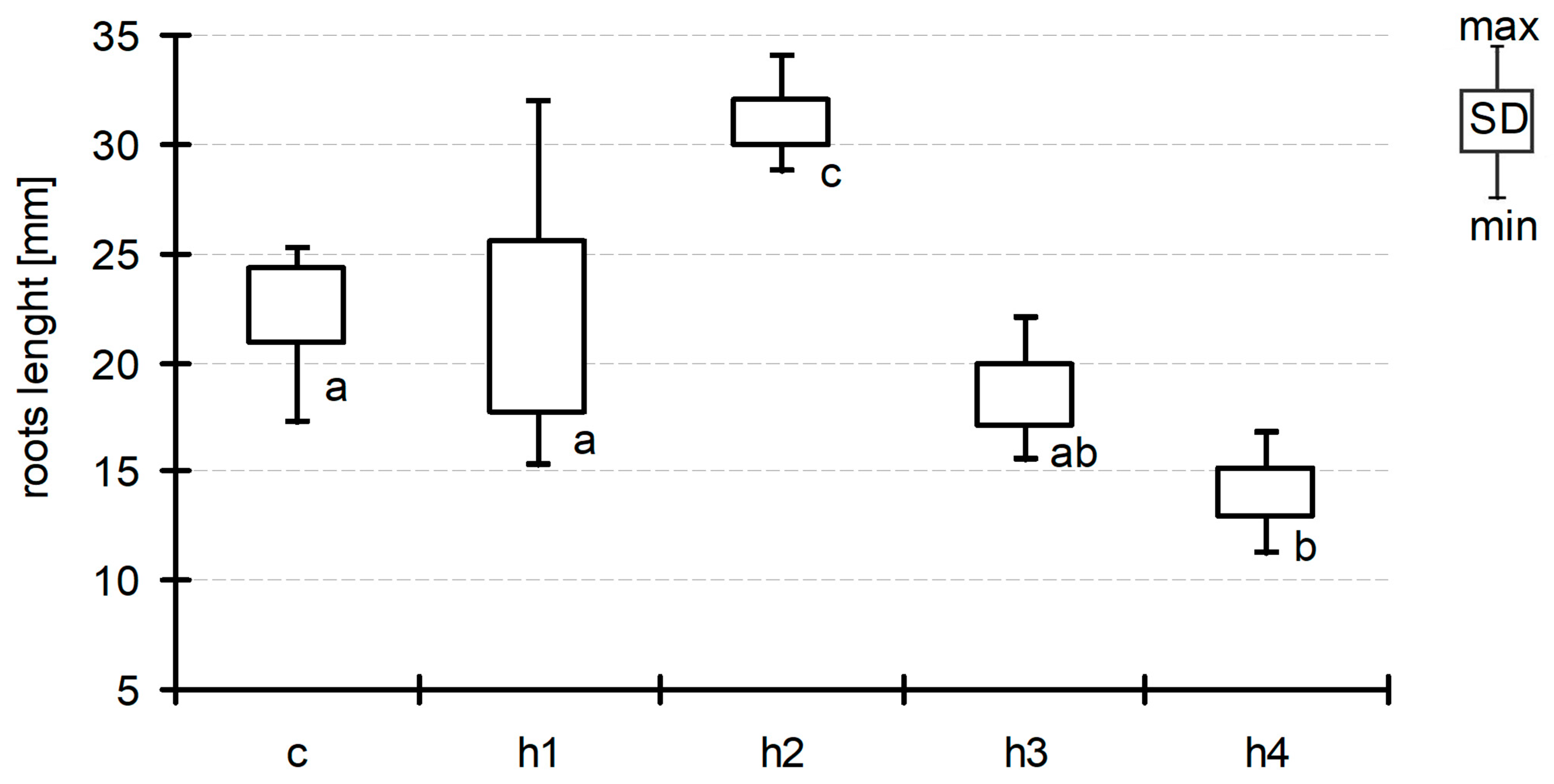
Figure 4.
Change in TP concentration between D01 and D07. Observed differences were not significant (ANOVA at p < 0.05).
Figure 4.
Change in TP concentration between D01 and D07. Observed differences were not significant (ANOVA at p < 0.05).
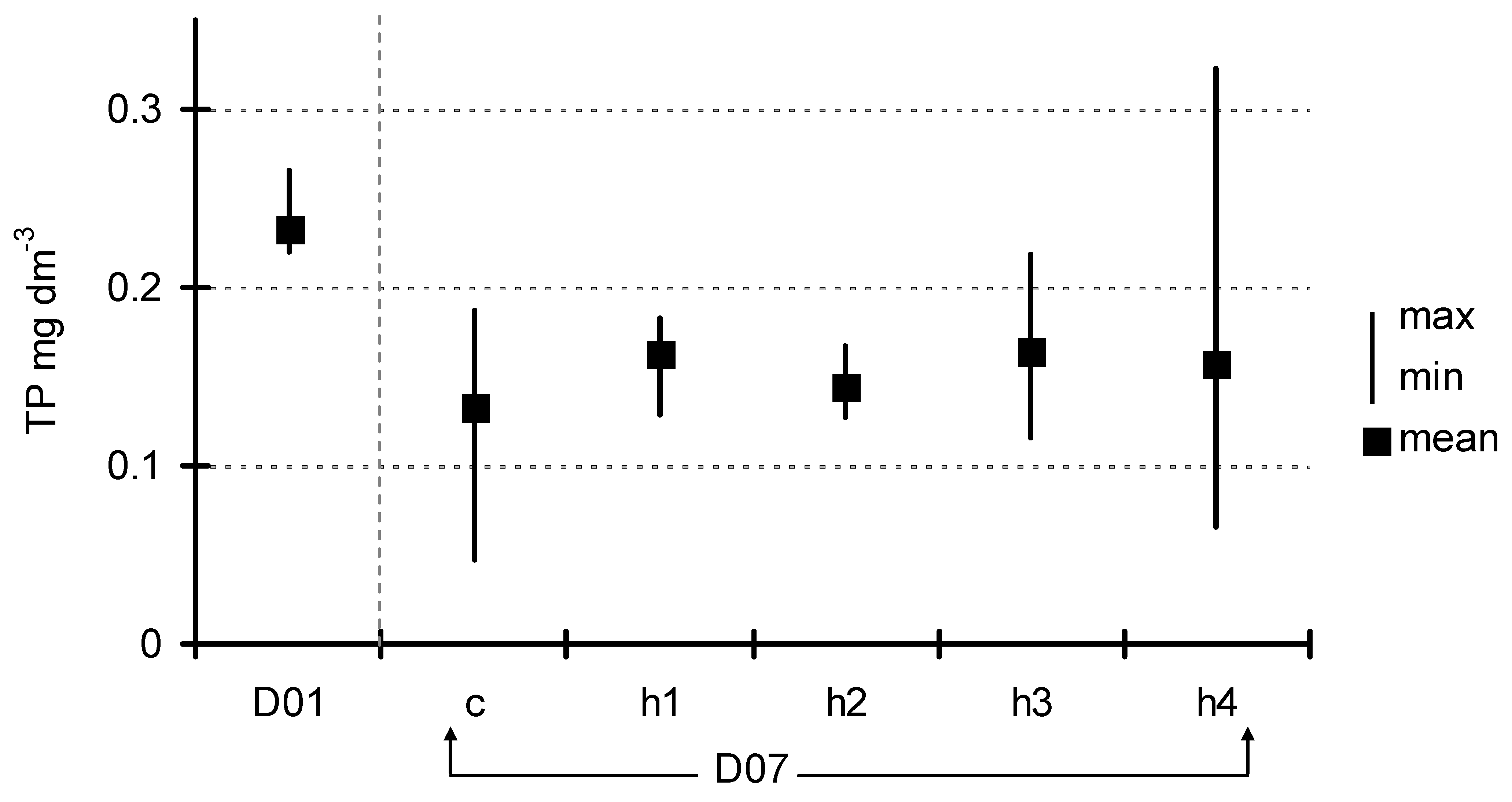
Figure 5.
Change in P-PO4 concentration between D01 and D07 (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).
Figure 5.
Change in P-PO4 concentration between D01 and D07 (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).
Figure 6.
Change in N-NO3 concentration between D01 and D07 (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).
Figure 6.
Change in N-NO3 concentration between D01 and D07 (means with the same letters did not differ significantly; post hoc Duncan’s test at p < 0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biomass (g FW) of Lemna minor in control and treatments at the beginning of the experiment.
Table 1.
Biomass (g FW) of Lemna minor in control and treatments at the beginning of the experiment.
| Treatment | c | h1 | h2 | h3 | h4 |
|---|---|---|---|---|---|
| mean | 0.086 | 0.086 | 0.086 | 0.085 | 0.085 |
| SD | 0.004 | 0.001 | 0.003 | 0.003 | 0.002 |
Table 2.
Basic parameters of water at the beginning (D01) and at the end (D07) of the experiment.
| D01 | D07 | |||||
|---|---|---|---|---|---|---|
| c | h1 | h2 | h3 | h4 | ||
| EC (μS cm−1) | 364 | 371.5 ± 15.3 | 355 ± 7.6 | 358.7 ± 9.1 | 368.7 ± 25.6 | 357.5 ± 7.0 |
| pH | 8.7 | 8.8–8.9 | 8.8–8.9 | 8.7–8.8 | 8.6–8.7 | 8.5–8.6 |
| TOC (mg dm−3) | 4.0 ± 0.1–36.5 ± 0.4 | 4.9 ± 0.1 | 6.7 ± 0.4 | 10.1 ± 0.3 | 22.9 ± 5.0 | 38.0 ± 9.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pęczuła, W.; Tarkowska-Kukuryk, M.; Niedźwiecki, M. Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment. Water 2022, 14, 3826. https://doi.org/10.3390/w14233826
AMA Style
Pęczuła W, Tarkowska-Kukuryk M, Niedźwiecki M. Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment. Water. 2022; 14(23):3826. https://doi.org/10.3390/w14233826
Chicago/Turabian StylePęczuła, Wojciech, Monika Tarkowska-Kukuryk, and Michał Niedźwiecki. 2022. "Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment" Water 14, no. 23: 3826. https://doi.org/10.3390/w14233826
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.