
Spatial Distribution of Cladocera in a Stratified Palaearctic Lake
Department of Hydrobiology and Protection of Ecosystems, University of Life Sciences, B. Dobrzańskiego 37, 20-262 Lublin, Poland
Water 2022, 14(17), 2667; https://doi.org/10.3390/w14172667
Submission received: 4 July 2022
/
Revised: 18 August 2022
/
Accepted: 24 August 2022
/
Published: 29 August 2022
(This article belongs to the Section Biodiversity and Functionality of Aquatic Ecosystems)
Abstract
:The spatial distribution of 39 species of Chydoridae, representing seven families, was examined in a stratified Palaearctic lake in Poland (Lake Piaseczno). Thirty-one of the species living in that lake were found to exist exclusively in the littoral zone. No species were found exclusively in the pelagic zone; in this zone, eight species were identified, but all of these were also quite abundant in the littoral zone. Almost all species of Chydoridae and all species of Macrothricidae were found only in littoral, whereas Daphnidae and Sididae families comprised species either restricted to the littoral or occurring in both pelagic and littoral zones. Two Bosminidae species, in turn, displayed quite a consistent distribution in both biotic zones, but differed in their habitat preferences. On the other hand, species of Polyphemidae and Leptodoridae exhibited littoral and littoral-pelagic distributions, respectively. Few species occurred in one habitat only. Most species, despite showing clear preferences for a distinct habitat (or habitats), were also present at lower densities throughout the littoral zone, or the whole lake, in the case of limnetic species. Archetype analysis showed that cladocerans were distributed along the entire gradient of macrophyte characteristics and habitat depth. Among Chydoridae, which displayed the strongest attachment to habitats overgrown by macrophytes, few species showed above 50% similarity to the archetype representing species living among submerged macrophytes with the highest plant per volume infested. Furthermore, many littoral species, including these of the Macrothricidae family, P. pediculus and S. mucronata, avoided staying in habitats with particularly complicated spatial architecture. Regarding their distribution in relation to depth, cladocerans could be grouped into species found in the whole littoral zone, species settling in relatively shallow parts of the littoral zone, species found mainly in habitats 3–6 m deep and species settling throughout the lake. Archetype analysis showed negligible similarity between cladocerans and the archetype representing species living in macrophyte-free habitats in the deepest parts of the lake. The study provides quantitative support showing that cladocerans display a distribution threshold at one time in one lake. Although this group of aquatic invertebrates is widely suspected of being ecologically flexible and tolerant to environmental gradients, individual species showed high spatial segregation according to depth and habitat architecture.
1. Introduction
Cladocera represent an important part of freshwater microfauna, living in all types of waters. The spatial distribution of cladocerans is structured by the spatial diversity of aquatic ecosystems. However, such distribution is additionally a spatial factor that diversifies lake ecosystems, because it can have a significant influence on biogene transfer, predators’ feeding activity and trophic relationships in food webs. In shallow lakes lacking a typical pelagic zone, cladocerans may coexist spatially. Deep, stratified lakes display spatial separation in littoral and pelagic zones resulting in characteristic spatial distribution of littoral and limnetic species. In each zone different habitats coexist, resulting in further differentiation in terms of the spatial distribution of cladocerans. In the littoral zone, the architecture of macrophytes in an important factor in diversifying habitats in a horizontal profile. A high diversity of habitats coincides with species diversity and organism density, weakens predator–prey relationships and leads to numerous scenarios of trophic interactions [1,2]. As a result, habitats settled by species of disparate ecological niches occur in the littoral zone. The pelagic zone is more homogeneous than the littoral zone. However, spatial gradients of temperature, light intensity, oxygen concentration, pH and concentration of biogenes can be observed in stratified lakes. Consequently, while photosynthesis is confined to the illuminated layers of water, organic matter accumulates in deeper waters; here the community respiration rate is enhanced and the oxygen concentration decreases. Moreover, most planktivorous fish require a minimum amount of visible light to search for their prey, whereas the activity of poikilothermic predators correlates with water temperature. Therefore, predation pressure on limnetic cladocerans is generally restricted to the upper illuminated and warm part of the water column [3]. The spatial distribution of cladocerans has tended to be investigated in shallow lakes, whereas few studies have been conducted in deep stratified lakes. Moreover, previous studies have generally focused on the influence of habitat variations in the littoral zone on the species structure of epiphytic fauna, rather than the littoral-benthic species of Cladocera [4,5,6]. Similarly, studies on the spatial distribution of Cladocera in pelagic zones have mainly concerned daily migrations in the vertical profile instead of observing their habitat choices within the lake [7,8,9]. In response, the aim of the present study was to identify the spatial distribution of cladocerans in a stratified lake. The specific questions asked were: (i) Do Cladocera show clear habitat preferences? (ii) Do cladocerans display family-specific patterns of distribution? and (iii) Do habitat traits determine the distribution of Cladocera? By comparing the spatial distribution of cladoceran families and species, this paper sheds light on the underlying drivers and mechanisms shaping their community assembly in a stratified lake.
2. Materials and Methods
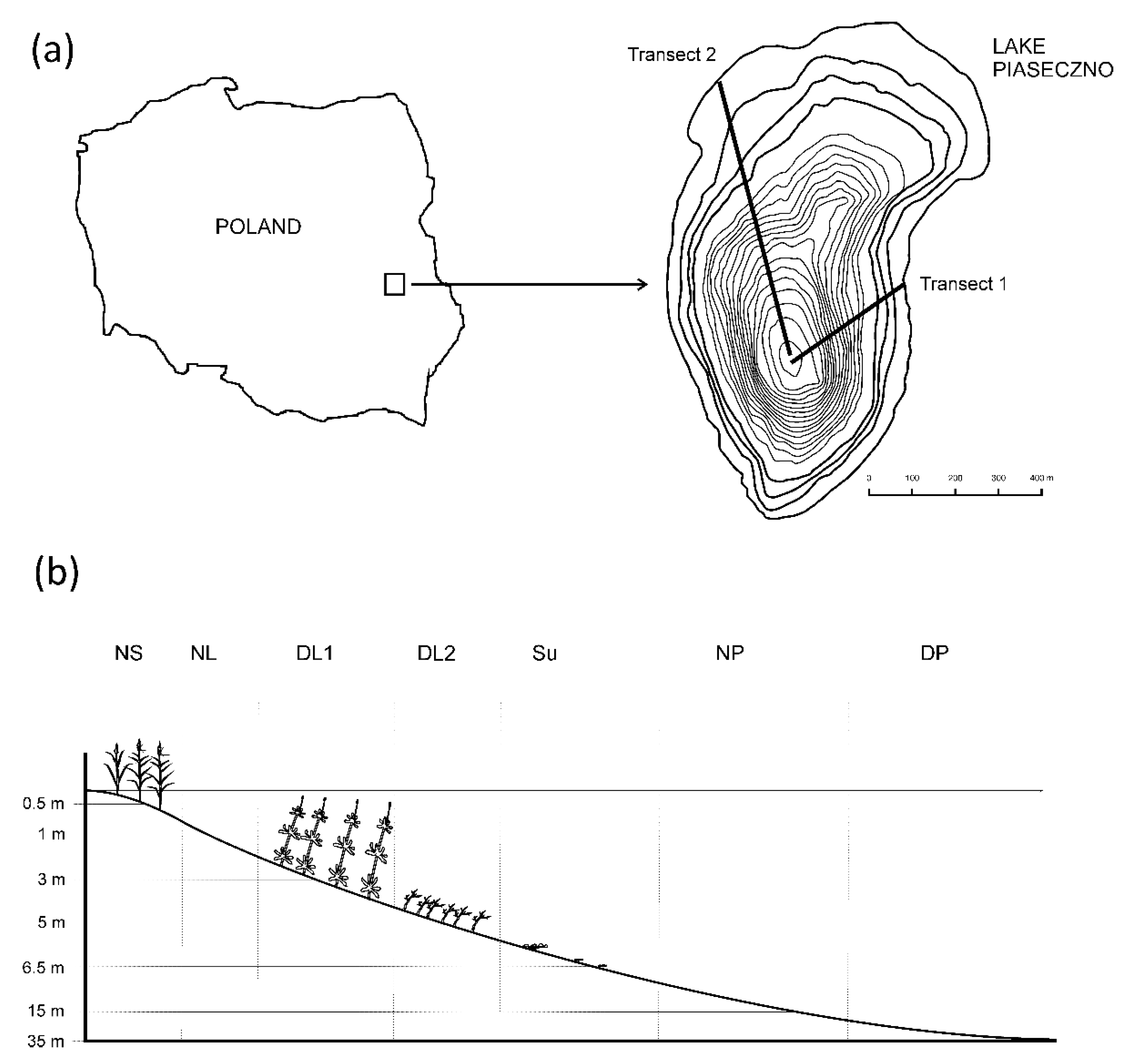
Field and Laboratory Analyses. The study was conducted in the mesotrophic Lake Piaseczno (51°23′03″ N, 23°01′46″ E), situated in eastern Poland. The lake area is 83.2 ha and its maximum depth reaches 38.8 m. The littoral zone reaches a depth of 6 m due to high water transparency and the pelagic zone exhibits a vertical temperature stratification. Samples were collected once a month from April to November in different habitats located along two transects (Figure 1), placed in the lake’s eastern and north-western parts, which were similar in depth and vegetation structure and stretched from the shore to the deepest part of the lake. In the littoral zone, five different habitats were distinguished: the near shore, overgrown by the common reed Phragmites australis (hereinafter NS); the near littoral, i.e., a macrophyte-free region with a sandy bottom (NL); the deep littoral, overgrown by water milfoil Myriophyllum alterniflorum (DL1) and macroalga Nitella flexilis (DL2); and the sublittoral, where the bottom is characterized by separate clusters of moss Drepanocladus sp. (Su). In each habitat, the number of samples collected increased with depth (Table 1). In the pelagic zone, the samples were taken in a non-stratified manner near the pelagial (NP) and in a stratified manner in the deep pelagial (DP). Similar to the case of the littoral zone, samples in the pelagial were collected from different depths in the water column (Table 1). Cladocerans were collected around noon using a 5 L sampler. Double samples, 10 dm3 in volume each, were sieved through a 40 µm mesh net and preserved with 5% buffered formalin treated with 5% glycerin. The cladocerans were classified to the species level and counted (ind.·dm−3) under a microscope at 100× magnification with the use of a Sedgewick-Rafter counting cell. A permutational multivariate analysis of variance using distance matrices was applied to test the statistical significance of the differences in the spatial variability of cladocerans. The differences in the densities of cladocerans were insignificant between the transects, thus the numerical data of density presented in this paper are the mean values for the two transects. To estimate the spatial architecture of the habitats, investigations of the macrophytes were carried out in April, May, June, August and October. Macrophyte density was expressed as plant per volume infested (PVI), calculated as the product of per cent coverage and height divided by water depth [10]. These data were subsequently used to estimate the impact of habitat traits on the distribution of Cladocera.
The influence of habitat traits on spatial distribution of Cladocera. The habitat traits examined were described using different types of variables: continuous (habitat depth, PVI for submerged macrophytes) and categorical (biological zone, macrophyte occurrence and macrophyte type). For each analysis, 2–6 modalities were created within each trait (Table S1 in Supplementary material), and each species was included in only one modality per trait based on its highest relative density. In order to identify the distribution of cladocerans, archetypal analysis was used [11]. This analysis was chosen because it represents observations in a multivariate data set as convex combinations of extreme points. As a result, it gives a simple and useful way of looking at multivariate data. In this analysis, each species of Cladocera was represented in a data set as a mixture of ‘individuals of pure type’ or ‘archetypes’. These are a small number of (not necessarily observed) extreme points in a set of multivariate observations and the data are expressed as a probabilistic mixture of archetypes [5,6]. In this study, continuous traits were scaled, to ensure equal weights. The values of the archetypes were determined by running the algorithm for k = 1, n + 1, by performing iterations and by calculating the residual sum of squares (RSS) for each one [6]. A small value of k with a low RSS was chosen, according to the ‘elbow criterion’. The R library ‘archetypes’ was used for this analysis [12].
3. Results
3.1. Cladocerans in Lake Piaseczno: General Information
In total, 39 species of cladocerans were found. The majority—24 species—represented the Chydoridae family, while five species belonged to the Daphnidae family and four to the Macrothricidae family. The Bosminidae and Sididae families were represented by two species each and the Leptodoridae and Polyphemidae families by one species each.
3.2. Spatial Distribution of Cladocera
All of the species of Chydoridae were found in the littoral zone, while only one species—Chydorus sphaericus—occurred in the pelagic zone as well. Chydorids exhibited a very patchy distribution in the littoral zone. Alona rectangula was found in NS and DL2, whereas Pleuroxus truncatus occurred in NS. Alonella excisa was found mainly in NL and DL1, whereas Acroperus elongatus, Monospilus dispar and Rhynhotalona falcata occurred mostly in NS and NL. The bulk of Alonella nana, Pseudochydorus globossus, Graptoleberis testudinaria, Alona costata, Alona guttata, Anchistropus emarginatus, Acroperus harpae, Alonella exigua and Camptocersus rectirostris were found in DL1. Eurycercus lamellatus shifted in very similar densities in DL1, DL2 and Su. Leydygia acanthocercoides and Pleuroxus aduncus were found mainly in DL1 and DL2, while Alona affinis occurred in DL1 and Su. Pleuroxus laevis was found in DL2, while Pleuroxus trigonellus was found in Su. Pleuroxus uncinatus and Alona intermedia occurred mainly in Su. Chydorus sphaericus, the sole species occurring in both the littoral and the pelagic zones, reached the highest densities in NP and DP (Figures S1–S24 in Supplementary Material).
The Macrothricidae family was found exclusively in the littoral zone. Macrothricidae displayed clear habitat preferences, as Drepanothrix dentata and Bunops serricaudata preferred to stay in Su, Macrothrix rosea was found mainly in NS, whereas Lathonura rectirostris shifted among DL1, DL2 and Su (Figures S25–S28 in Supplementary Material).
Among the five species of Daphnidae found in Lake Piaseczno, Simocephalus vetulus and Scapholeberis mucronata occurred exclusively in the littoral zone, whereas the remaining three species—Ceriodaphnia quadrangula, Daphnia cucullata and Daphnia longispina—occurred in both biotic zones. In the case of the two species found exclusively in the littoral zone, S. mucronata occurred only in NS and NL, whereas S. vetulus preferred to stay in DL1 and DL2. Among the other three species found in both the littoral and the pelagic zones, C. quadragula preferred to stay in DL1, whereas D. cucullata and D. longispina preferred NP and DP (Figures S29–S33 in Supplementary Material).
The two species of Bosminidae found in Lake Piaseczno settled in both the littoral and the pelagic zones, although their densities were higher in the latter. The species differed in terms of their habitat preferences, as Bosmina longirostris stayed in the highest densities in widely contrasting habitats (NS and DP), whereas Bosmina coregoni preferred pelagic habitats (NP and DP) (Figures S34 and S35 in Supplementary Material).
Sididae were represented by two species; Sida crystallina occurred in the littoral zone and preferred NS and DL1, whereas Diaphanosoma brachyurum, although settled in both biotic zones, was most abundant in NP (Figures S36 and S37 in Supplementary Material).
The Polyphemidae family was represented by the sole species Polyphemus pediculus. It was observed only in the littoral zone, where it occurred in a very low density and preferred to stay in NS (Figure S38 in Supplementary Material).
The only representative of the Leptodoridae family—Leptodora kindtii—was observed in both zones, but achieved significantly higher density in the pelagic zone. Analyses of habitat preferences indicated that it preferred NP (Figure S39 in Supplementary Material).
3.3. Archetype Similarity
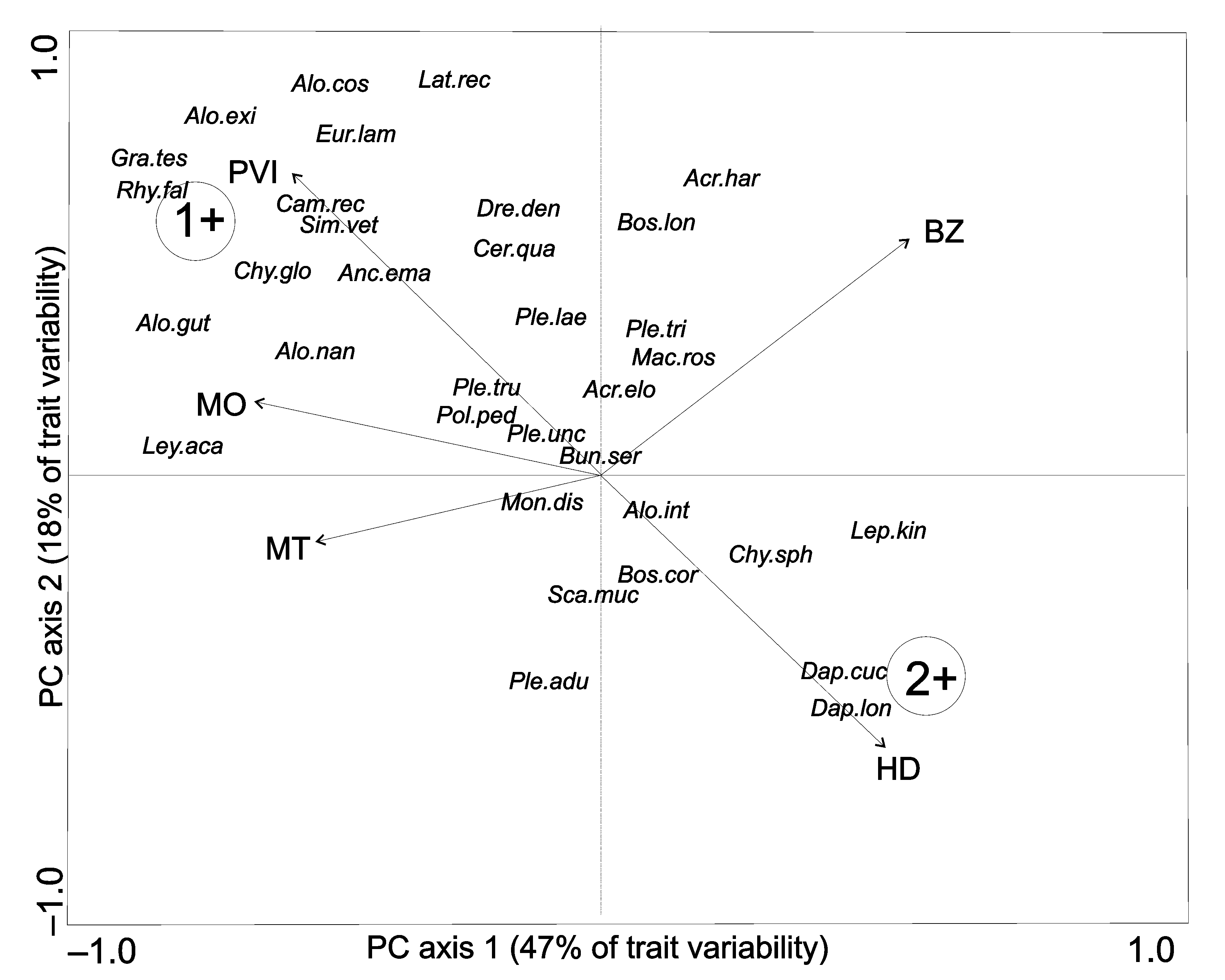
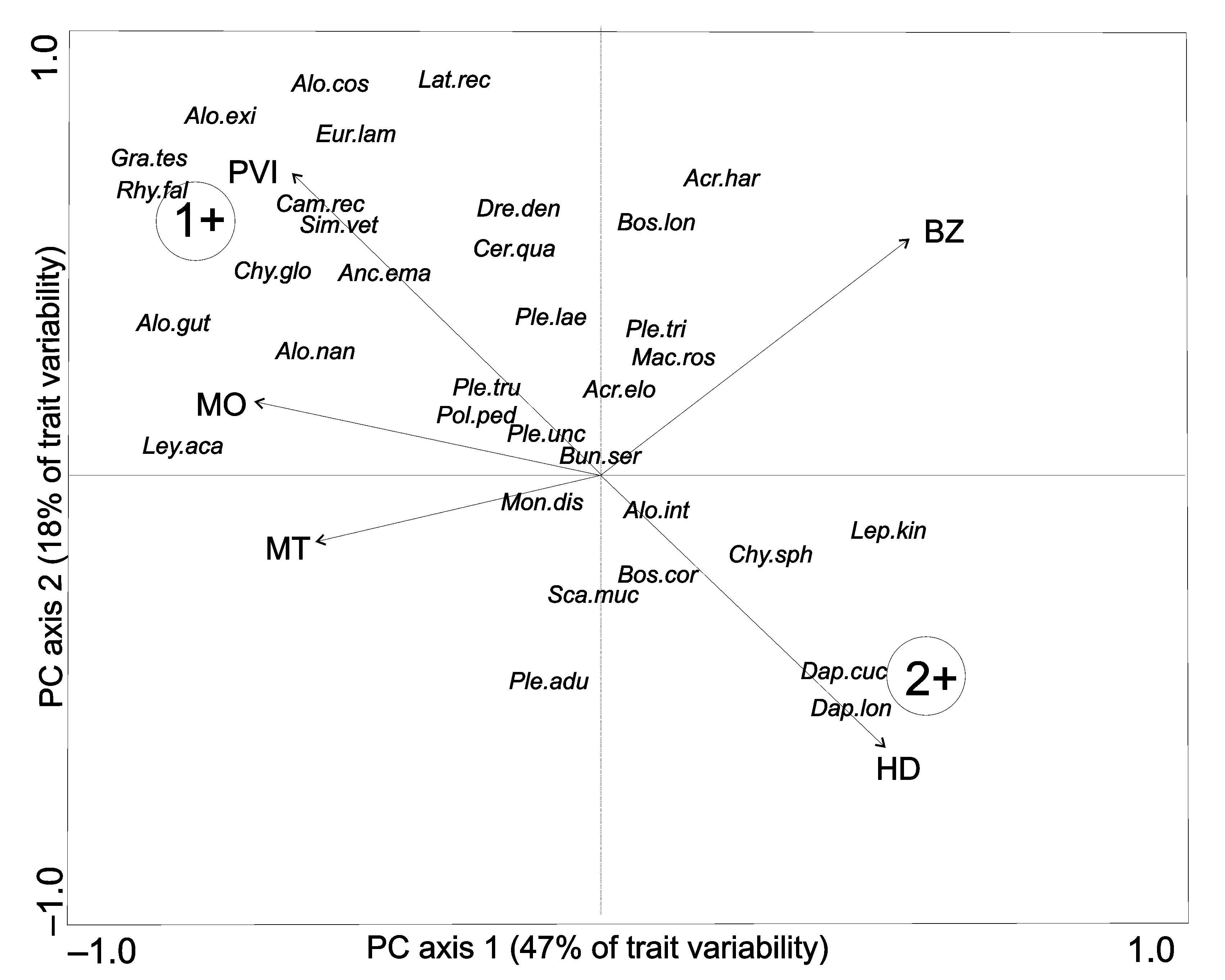
Using the ‘elbow criterion’, the optimal number of archetypes (habitat traits associated with species distribution) was two. Principal components analysis (PCA) biplot of the five traits used indicated that the first two PCs accounted for more than half of the trait variability. The first PC (47% of trait variability) was based mainly on macrophyte occurrence, macrophyte type and PVI, while the second (18% of trait variability) was based mainly on biotic zone and depth. The two archetypes depicted represent two hypothetical species existing in the multivariate set of 39 species, each possessing a combination of traits representing its distribution. The first archetype species (Figure 2, 1+) occurred in the littoral zone among submerged macrophytes with the highest PVI. The second archetype species (Figure 2, 2+) stayed in macrophyte-free habitats with the greatest depth. The proportional affinity of each species to the two archetypes identified indicates that half of the Chydoridae species were associated with archetype-1 (Table S2 in Supplementary Material). Among this group, A. costata, G. testudinaria, A. exigua and C. globossus shared more than 80% similarity with archetype-1. The remaining chydorids shared less than 30% similarity with each archetype (Figure 2, Figure S40 in Supplementary Material). In the Macrophricidae family, only L. rectirostris showed a clear association with archetype-1, whereas the remaining macrothricids were weakly associated with each archetype (Figure 2, Table S2 and Figure S41 in Supplementary Material). Among Daphnidae, C. quadrangula and S. vetulus were associated with archetype-1, sharing 33% and 69% similarity, respectively. Daphnia cucullata and D. longispina, respectively, shared 50% and 54% similarly with archetype-2, whereas M. dispar was similar to neither archetype-1 nor archetype-2 (Figure 2, Table S2 and Figure S42 in Supplementary Material). The other families of Cladocera—including Bosminidae, Leptodoridae, Polyphemidae and Sididae—exhibited little similarity to each archetype (Figure 2, Table S2 and Figures S43–S46 in Supplementary Material).
4. Discussion
Do Cladocera show clear habitat preferences? Of the 39 species of cladocerans living in Lake Piaseczno, 31 existed exclusively in the littoral zone. However, few species occurred in one habitat only, including P. truncatus, A. intermedia and A. costata of the Chydoridae family as well as M. rosea and B. serricaudata of the Macrothricidae family. Other littoral species, despite showing clear preferences for a distinct habitat (or habitats), were also distributed in lower densities throughout the littoral zone. In general, the total density of cladocerans was higher in macrophyte thickets and proceeded in the following order: DL1 > NS > DL2 > Su > NL. No species were found exclusively in the pelagic zone. In this zone, eight species were identified, a typical number for the pelagic zone of deep lakes [13]; however, these species were also quite abundant in the littoral zone. Some authors have hypothesized that limnetic cladocerans are particularly abundant in habitats overgrown with macrophytes [2,14,15]. In Lake Piaseczno, only C. quadrangula, despite being present in both zones, preferred to stay in DL1, i.e., the habitat overgrown with water milfoil. Other limnetic species showed a heterogeneous distribution in the pelagic zone, as daphnids and bosminids preferred DP, whereas Leptodora, Diaphanosoma and C. sphaericus stayed mainly in NP. Comparing Lake Piaseczno with other deep reservoirs, the spatial distribution of limnetic species appears to differ in relation to the characteristics of the aquatic ecosystem in question. In some ecosystems, C. sphaericus occurs exclusively in the habitats of the littoral zone [15], whereas daphnids and bosminids are evenly distributed in the littoral and pelagic zones [14].
Do cladocerans display a family-specific distribution? In this study, almost all of the species of Chydoridae and all of the species of Macrothricidae were found exclusively in the littoral zone. Previous studies regarding the ecology of Chydoridae have shown that their distribution is commonly linked with the occurrence of aquatic plants [16,17,18]. Indeed, the largest number of species was found in DL1, which was overgrown with water milfoil and exhibited 89% PVI and a dissected structure. However, some species of Chydoridae preferred littoral habitats that were less infested by macrophytes. Different habitat preferences may be the result of the chydorids’ wide range of specializations. Chydoridae are generally described as scrapers, but they actually represent various feeding mechanisms. Extreme cases are A. emarginatus (which is a carnivore of cnidarians), P. globossus (which scavenges on carrion) [19] and R. falcata (which extracts food from the mud in which it lives) [20]. These three species, although not tightly connected with the occurrence of aquatic plants, displayed clear habitat preferences concordant with the distribution of other Chydorids. Similarly, habits of Macrothricidae are diverse and permit the exploitation of a wide variety of ecological niches. According to this study’s findings, Macrothricidae species, although found exclusively in the littoral zone, clearly avoided habitats overgrown by dense macrophytes. This might have resulted from their weak locomotive abilities, enabling them to move among more spatially complex habitats, given that most macrothricids neither swim nor creep efficiently [21].
The families of Daphnidae and Sididae comprised species either restricted to the littoral zone or occurring in both zones. S. vetulus occurred only in the littoral zone and preferred to stay in DL1 and DL2, habitats overgrown by water milfoil and macroalgae. This is in agreement with other studies noting that this species inhabits macrophyte-rich littoral zones [22,23], although its habitat preferences do not result from its feeding mechanism, as it is able to use diverse feeding sources [24]. The other Daphnidae species are found exclusively in the littoral zone. S. mucronata is hyponeustic and positively phototactic [25], and is known to occur in small and shallow water bodies [26]. Indeed, even in the deep Lake Piaseczno, it occurred exclusively in NS and NL, the shallowest habitats of the littoral zone. In the Sididae family, represented by two species, the distribution of S. crystallina was restricted to the littoral zone exclusively. Some previous papers have reported that S. crystallina is largely confined to the littoral zone by its attachment to aquatic macrophytes, but unlike most plant-associated species, it is a phytoplankton filter feeder [27]. Sida crystallina clearly preferred two habitats in Lake Piaseczno: NS and DL1. These habitats, although both overgrown with macrophytes, displayed a distinct spatial architecture, as NS was the shallowest habitat and was covered with the emergent macrophyte common reed, whereas DL1 was quite deep and was overgrown with the submerged macrophyte water milfoil. Two Bosminidae species, in turn, displayed quite a consistent distribution in both biotic zones, although they differed in their habitat preferences. Indeed, as mentioned above, B. coregoni preferred to stay in DP, whereas B. longirostris was found in two widely contrasting habitats (NS and DP), a finding that supports previous works describing it as a cosmopolitan taxon with an ability to tolerate a range of environmental conditions [28,29,30].
Do habitat traits determine the distribution of Cladocera? Among the five habitat traits included in the archetype analysis, macrophyte characteristics and habitat depth clearly had the greatest influence on the distribution of cladocerans. Previous studies have shown that littoral complexity influences the availability of cladocerans to predators exploring the littoral zone; however, such availability also results from the types of predators in question as well as the size and the motility of the prey [31,32,33,34]. This explains why cladocerans were distributed along the whole gradient of macrophyte characteristics. Among Chydoridae, which displayed the strongest attachment to habitats overgrown by macrophytes, few species showed above 50% similarity to the archetype representing species living among submerged macrophytes with the highest PVI. This observation is consistent with other reports that Chydoridae can display patterns of separation according to depth even in morphologically homogeneous lakes [35]. Many littoral species—including those of the Macrothricidae family, P. pediculus and S. mucronata—avoided staying in habitats with a more complicated spatial architecture, as can also be explained by their habits as described above.
Depth was the other factor influencing the distribution of cladocerans. Regarding the distribution of cladocerans in relation to habitat depth, four groups could be distinguished: (i) species settling in relatively shallow parts of the littoral zone (A. elongatus, A. excisa, A. exigua, A. guttata, A. rectangula, M. dispar, M. rosea, P. pediculus, P. truncatus, R. falcata and S. mucronata); (ii) species found mainly in areas located 3–6 m deep (A. costata, A. intermedia, B. serricaudata, C. rectirostris, D. dentata, E. lamellatus, G. testudinaria, L. rectirostris, P. laevis, P. uncinatus and S. vetulus); (iii) species found in the whole littoral zone (A. affinis, A. harpae, A. nana, P. aduncus, P. globosus, P. trigonellus and S. crystallina); and (iv) species settling throughout the lake (B. coregoni, B. longiristris, C. quadrangula, C. sphaericus, D. brachyurum, D. cucullata, D. longispina and L. kindtii). Some studies have reported that lake depth influences the community structure of many cladocerans [36,37]. Furthermore, even in morphologically homogeneous lakes, cladocerans can display patterns of separation according to depth, which are forced upon them by the thermal properties, ultraviolet (UV) exposure, food resources and predators associated with these varying depths [35]. Interestingly, the archetype analysis showed a low level of similarity between cladocerans and the archetype representing species living in macrophyte-free habitats with the greatest depth. Rather, cladocerans were distributed throughout the depth range, including those species occurring in relatively high densities in deeper habitats, like Daphnia, Bosmina, Diaphanosoma and Leptodora.
5. Conclusions
The spatial structure and the depth of habitats proved to be significant factors influencing the distribution of cladocerans in both the littoral and the pelagic zones. Inter- and intra-lake differences in these traits, observed in the biotic zones of deep stratified lakes, are frequently ignored in field studies on the distribution of aquatic biota. Nevertheless, this study suggests that variations in the diversity and the abundance of cladocerans should be considered at small spatial scales in such lakes. Otherwise, we cannot fully ascertain the species structure in both the littoral and the pelagic zones, this often being falsely perceived as quite homogeneous in stratified lakes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14172667/s1.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Grabowski, J.H.; Hughes, A.R.; Kimbro, D.L. Habitat complexity influences cascading effects of multiple predators. Ecology 2008, 89, 3413–3422. [Google Scholar] [CrossRef] [PubMed]
- Stefanidis, K.; Papastergiadou, E. Influence of hydrophyte abundance on the spatial distribution of zooplankton in selected lakes in Greece. Hydrobiologia 2010, 656, 55–65. [Google Scholar] [CrossRef]
- Zaret, T.M. Predation and Freshwater Communities; Yale University Press: New Haven, CT, USA; London, UK, 1980. [Google Scholar]
- Schramm, H.L., Jr.; Jirka, K.J.; Hoyer, M.V. Epiphytic Macroinvertebrates on Dominant Macrophytes in Two Central Florida Lakes. J. Freshw. Ecol. 1987, 4, 151–161. [Google Scholar] [CrossRef]
- Paterson, M. The distribution of microcrustacea in the littoral zone of a freshwater lake. Hydrobiologia 1993, 263, 173–183. [Google Scholar] [CrossRef]
- Cheruvelil, K.S.; Soranno, P.A.; Madsen, J.D.; Roberson, M.J. Plant architecture and epiphytic macroinvertebrate communities: The role of an exotic dissected macrophyte. J. North Am. Benthol. Soc. 2002, 21, 261–277. [Google Scholar] [CrossRef]
- Matsumura-Tundisi, T.; Tundisi, J.G.; Tavares, L.S. Diel migration and vertical distribution of Cladocera in Lake D. Helvécio (Minas Gerais, Brazil). Hydrobiologia 2014, 113, 299–306. [Google Scholar] [CrossRef]
- Adamczuk, M. Predation follows competition in depth selection behaviour of Cladocera in a deep lake (E Poland). Biol. Lett. 2009, 46, 29. [Google Scholar] [CrossRef]
- Battes, K.P.; Momeu, L. Diel vertical distribution of planktonic microcrustaceans (Crustacea: Cladocera, Copepoda) in a natural shallow lake from Transylvania, Romania. J. Limnol. 2014, 73, 34–44. [Google Scholar] [CrossRef]
- Canfield, J.E.; Shireman, J.V.; Haller, W.T.; Watkins, G.E.; Maceina, M.J. Prediction of Chlorophyll a Concentrations in Florida Lakes: Importance of Aquatic Macrophytes. Can. J. Fish. Aquat. Sci. 1984, 41, 497–501. [Google Scholar] [CrossRef]
- Cutler, A.; Breiman, L. Archetypal analysis. Technometrics 1994, 36, 338–347. [Google Scholar] [CrossRef]
- Eugster, M.J.A.; Leisch, F.R. From Spider-Man to Hero-Archetypal Analysis in R. J. Stat. Softw. 2009, 30, 1–23. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Species richness of pelagic Cladocera of large lakes in the eastern hemisphere. Hydrobiologia 2000, 434, 41–54. [Google Scholar] [CrossRef]
- Jeppesen, E.; Lauridsen, T.L.; Kairesalo, T.; Perrow, M.R. Impact of Submerged Macrophytes on Fish-Zooplankton Interactions in Lakes. In The Structuring Role of Submerged Macrophytes in Lakes; Jeppesen, E., Søndergaard, M., Søndergaard, M., Christoffersen, K., Eds.; Springer: New York, NY, USA, 1998; pp. 91–114. [Google Scholar] [CrossRef]
- Geraldes, A.M.; Boavida, M.J. Do Littoral Macrophytes Influence Crustacean Zooplankton Distribution? Limnetica 2004, 23, 57–63. [Google Scholar] [CrossRef]
- Hann, B.J. Occurrence and distribution of littoral Chydoridae (Crustacea, Cladocera) in Ontario, Canada, and taxonomic notes on some species. Can. J. Zoöl. 1981, 59, 1465–1474. [Google Scholar] [CrossRef]
- Chengalath, R. The distribution of chydorid Cladocera in Canada. Hydrobiologia 1987, 145, 151–157. [Google Scholar] [CrossRef]
- Tremel, B.; Frey, S.E.; Yan, N.D.; Somers, K.M.; Pawson, T.W. Habitat specificity of littoral Chydoridae (Crustacea, Branchiopoda, Anomopoda) in Plastic Lake, Ontario, Canada. Hydrobiologia 2000, 432, 195–205. [Google Scholar] [CrossRef]
- Van Damme, K.; Dumont, H.J. Limb morphology of the carnivorous anomopods Anchistropus emarginatus Sars, 1862 and Pseudochydorus globosus (Baird, 1843) (Crustacea: Branchiopoda: Anomopoda). Ann. Limnol.-Int. J. Limnol. 2007, 43, 271–284. [Google Scholar] [CrossRef]
- Fryer, G. Evolution and adaptive radiation in the chydoridae (Crustacea: Cladocera): A study in comparative functional morphology and ecology. Philos. Trans. R. Soc. Lond. Ser. B 1968, 254, 221–283. Available online: https://www.jstor.org/stable/2416820 (accessed on 9 February 2022).
- Fryer, G. Evolution and adaptive radiation in the Macrothricidae (Crustacea: Cladocera): A study in comparative functional morphology and ecology. Philos. Trans. R. Soc. Lond. Ser. B 1974, 269, 137–274. [Google Scholar] [CrossRef]
- Amoros, C.; Chessel, D. Populations of Cladocera (Crustacea), as describers of the hydrological functioning of stagnant waterways. Ann. Limnol.-Int. J. Limnol. 1985, 21, 227–240. [Google Scholar] [CrossRef]
- Frey, D.G. The taxonomy and biogeography of the Cladocera. Hydrobiologia 1987, 145, 5–17. [Google Scholar] [CrossRef]
- Bec, A.; Desvilettes, C.; Véra, A.; Fontvieille, D.; Bourdier, G. Nutritional value of different food sources for the benthic Daphnidae Simocephalus vetulus: Role of fatty acids. Arch. Hydrobiol. 2003, 156, 145–163. [Google Scholar] [CrossRef]
- De Meester, L. Phototactic behaviour of hyponeustic Crustacea: Scapholeberinae (Cladocera). Biol. Jaarb. Dodonaea 1988, 56, 78–85. [Google Scholar]
- De Meester, L.; Maas, S.; Diercken, K.; Dumont, H. Habitat selection and patchiness in Scapholeberis: Horizontal distribution and migration of S.mucronata in a small pond. J. Plankton Res. 1993, 15, 1129–1139. [Google Scholar] [CrossRef]
- Fairchild, G.W. Movement and Microdistribution of Sida Crystallina and Other Littoral Microcrustacea. Ecology 1981, 62, 1341–1352. [Google Scholar] [CrossRef]
- Deevey, E.S.; Deevey, G.B. The American species of Eubosmina seligo (Crustacea, Cladocera). Limnol. Oceanogr. 1971, 16, 201–218. [Google Scholar] [CrossRef]
- Lieder, U. Revision of the genus Bosmina Baird, 1845 (Crustacea, Cladocera). Int. Rev. Ges. Hydrobiol. 1983, 68, 121–139. [Google Scholar] [CrossRef]
- Adamczuk, M. Past, present, and future roles of small cladoceran Bosmina longirostris (O. F. Müller, 1785) in aquatic ecosystems. Hydrobiologia 2016, 767, 1–11. [Google Scholar] [CrossRef]
- Manatunge, J.; Asaeda, T. Optimal foraging as the criteria of prey selection by two centrarchid fishes. Hydrobiologia 1999, 391, 223–240. [Google Scholar] [CrossRef]
- Messyasz, B.; Kuczyńska-Kippen, N. Periphytic algal communities: A comparison of Typha angustifolia L. and Chara tomentosa L. beds in three shallow lakes (West Poland). Pol. J. Ecol. 2006, 54, 15–27. [Google Scholar]
- Zingel, P.; Nõges, P.; Tuvikene, L.; Feldmann, T.; Järvalt, A.; Tõnno, I.; Agasild, H.; Tammert, H.; Luup, H.; Salujõe, J.; et al. Ecological processes in macrophyte—And phytoplankton dominated shallow lakes. Proc. Est. Acad. Sci. Biol. Ecol. 2006, 55, 280–307. [Google Scholar]
- Adamczuk, M. The effect of habitat complexity on the contribution of some littoral–benthic Cladocera to the pelagic food web. Mar. Freshw. Res. 2013, 64, 1049–1057. [Google Scholar] [CrossRef]
- Nevalainen, L. Distribution of benthic microcrustaceans along a water depth gradient in an Austrian Alpine lake—Sedimentary evidence for niche separation. Limnologica 2012, 42, 65–71. [Google Scholar] [CrossRef]
- Korhola, A.; Olander, H.; Blom, T. Cladoceran and chironomid assemblages as quantitative indicators of water depth in subarctic Fennoscandian lakes. J. Paleolimnol. 2000, 24, 43–54. [Google Scholar] [CrossRef]
- Amsinck, S.L.; Strzelczak, A.; Bjerring, R.; Landkildehus, F.; Lauridsen, T.L.; Christoffersen, K.; Jeppesen, E. Lake depth rather than fish planktivory determines cladoceran community structure in Faroese lakes—Evidence from contemporary data and sediments. Freshw. Biol. 2006, 51, 2124–2142. [Google Scholar] [CrossRef]
Figure 1.
Location of the study area (a) and sampling sites (b). Habitat codes: NS—near shore, NL—near littoral, DL1—deep littoral 1, DL2—deep littoral 2, Su—sublittoral, NP—near pelagial, DP—deep pelagial.
Figure 1.
Location of the study area (a) and sampling sites (b). Habitat codes: NS—near shore, NL—near littoral, DL1—deep littoral 1, DL2—deep littoral 2, Su—sublittoral, NP—near pelagial, DP—deep pelagial.
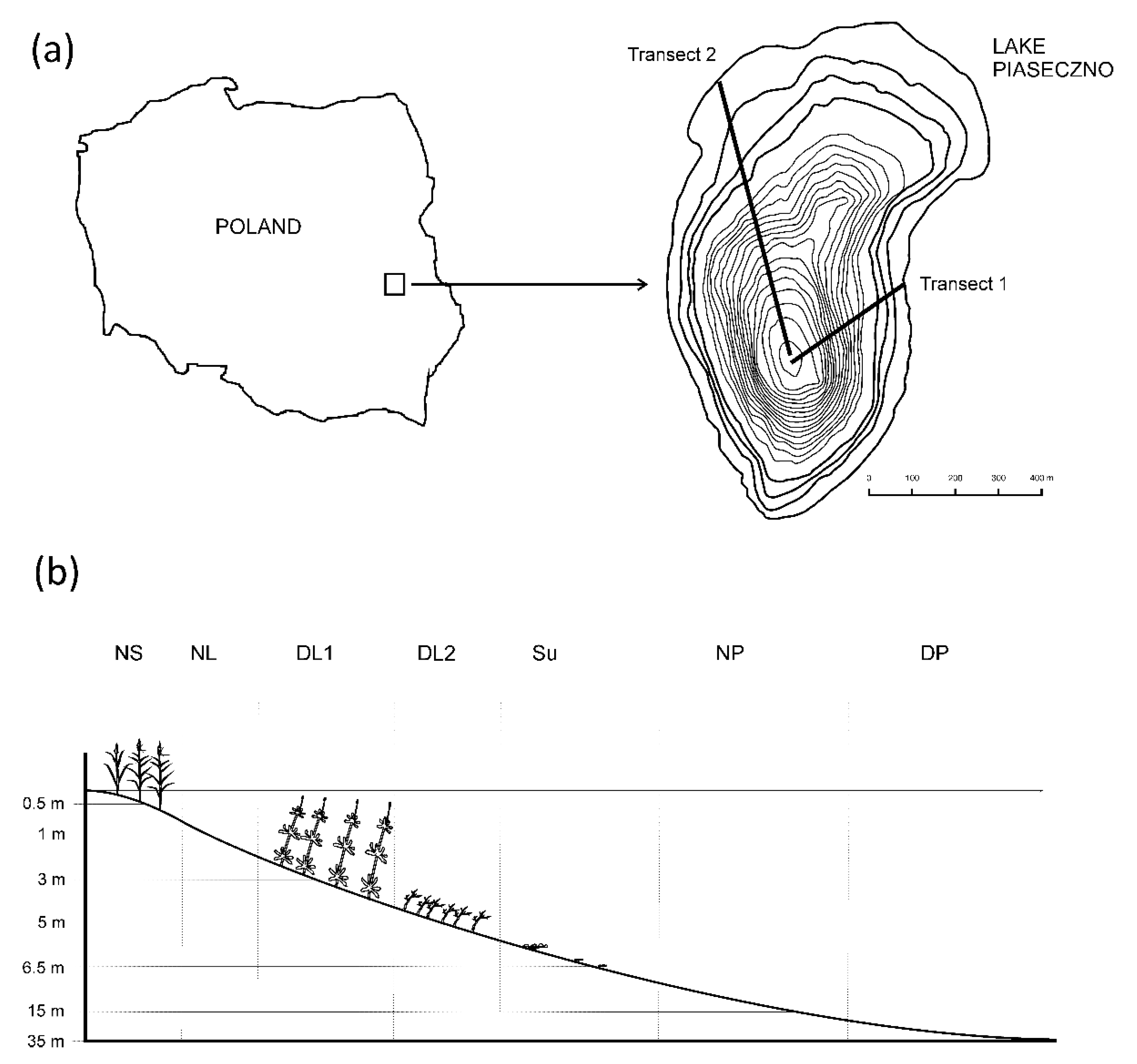
Figure 2.
Biplot of the PCA performed on five traits. Trait codes: PVI—plant volume infested, MO—macrophyte occurrence, MT—macrophyte type, BZ—biological zone, HD—habitat depth. The resulting archetypes are visualized as 1+ (Archetype 1) and 2+ (Archetype 2). Species codes: Acr.elo—Acroperus elongatus, Acr.har—Acroperus harpae, Alo.cos—Alona costata, Alo.gut—Alona guttata, Alo.rec—Alona rectangula, Alo.exi—Alonella exigua, Alo.exc—Alonella excisa, Alo.nan—Alonella nana, Anc.ema—Anchistropus emarginatus, Alo.aff—Alona affinis, Alo.int—Alona intermedia, Cam.rec—Camptocercus rectirostris, Chy.sph—Chydorus sphaericus, Eur.lam—Eurycercus lamellatus, Gra.tes—Graptoleberis testudinaria, Ley.aca—Leydygia acanthocercoides, Mon.dis—Monospilus dispar, Ple.adu—Pleuroxus aduncus, Ple.lae—Pleuroxus laevis, Ple.tri—Pleuroxus trigonellus, Ple.tru—Pleuroxus truncatus, Ple.unc—Pleuroxus uncinatus, Pse.glo—Pseudochydorus globossus, Rhy.fal—Rhynchotalona falcata, Bun.ser—Bunops serricaudata, Dre.den—Drepanothrix dentata, Lat.rec—Lathonura rectirostris, Mac.ros—Macrothrix rosea, Cer.qua—Ceriodaphnia quadrangula, Dap.cuc—Daphnia cucullata, Dap.lon—Daphnia longispina, Sca.muc—Scapholeberis mucronata, Sim.vet—Simocephalus vetulus, Bos.cor—Bosmina coregoni, Bos.lon—Bosmina longirostris, Dia.bra—Diaphanosoma brachyurum, Sid.cry—Sida crystallina, Pol.ped—Polyphemus pediculus, Lep.kin—Leptodora kindtii.
Figure 2.
Biplot of the PCA performed on five traits. Trait codes: PVI—plant volume infested, MO—macrophyte occurrence, MT—macrophyte type, BZ—biological zone, HD—habitat depth. The resulting archetypes are visualized as 1+ (Archetype 1) and 2+ (Archetype 2). Species codes: Acr.elo—Acroperus elongatus, Acr.har—Acroperus harpae, Alo.cos—Alona costata, Alo.gut—Alona guttata, Alo.rec—Alona rectangula, Alo.exi—Alonella exigua, Alo.exc—Alonella excisa, Alo.nan—Alonella nana, Anc.ema—Anchistropus emarginatus, Alo.aff—Alona affinis, Alo.int—Alona intermedia, Cam.rec—Camptocercus rectirostris, Chy.sph—Chydorus sphaericus, Eur.lam—Eurycercus lamellatus, Gra.tes—Graptoleberis testudinaria, Ley.aca—Leydygia acanthocercoides, Mon.dis—Monospilus dispar, Ple.adu—Pleuroxus aduncus, Ple.lae—Pleuroxus laevis, Ple.tri—Pleuroxus trigonellus, Ple.tru—Pleuroxus truncatus, Ple.unc—Pleuroxus uncinatus, Pse.glo—Pseudochydorus globossus, Rhy.fal—Rhynchotalona falcata, Bun.ser—Bunops serricaudata, Dre.den—Drepanothrix dentata, Lat.rec—Lathonura rectirostris, Mac.ros—Macrothrix rosea, Cer.qua—Ceriodaphnia quadrangula, Dap.cuc—Daphnia cucullata, Dap.lon—Daphnia longispina, Sca.muc—Scapholeberis mucronata, Sim.vet—Simocephalus vetulus, Bos.cor—Bosmina coregoni, Bos.lon—Bosmina longirostris, Dia.bra—Diaphanosoma brachyurum, Sid.cry—Sida crystallina, Pol.ped—Polyphemus pediculus, Lep.kin—Leptodora kindtii.

{kind=link}
{kind=link}
Table 1.
The characteristics of the habitats and the number of samples taken in each habitat during the study period.
Table 1.
The characteristics of the habitats and the number of samples taken in each habitat during the study period.
| Habitat Code | Approximate Distance from the Shore (m) | Depth (m) | Depth Levels of Sampling (m) | Total Number of Samples Taken in Each Transect during the Study Period | ||
|---|---|---|---|---|---|---|
| Transect 1 | Transect 2 | |||||
| Near shore | NS | 1 | 1 | 0.7 | 0.3 | 8 |
| Near littoral | NL | 10 | 13 | 1 | 0.1, 1 | 16 |
| Deep littoral 1 | DL1 | 40 | 57 | 3 | 0.1, 1, 3 | 24 |
| Deep littoral 2 | DL2 | 85 | 103 | 5 | 0.1, 1, 3, 4.9 | 32 |
| Sublittoral | Su | 135 | 206 | 6.5 | 0.1, 1, 3, 5, 6.4 | 40 |
| Near pelagial | NP | 225 | 314 | 15 | 0.1, 1, 3, 5, 7, 9, 11, 13, 15 | 72 |
| Deep pelagial | DP | 415 | 858 | 35 | 0.1, 1, 3, 5, 7, 9, 11, 13, 15, 19, 25, 30, 35 | 104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Adamczuk, M. Spatial Distribution of Cladocera in a Stratified Palaearctic Lake. Water 2022, 14, 2667. https://doi.org/10.3390/w14172667
AMA Style
Adamczuk M. Spatial Distribution of Cladocera in a Stratified Palaearctic Lake. Water. 2022; 14(17):2667. https://doi.org/10.3390/w14172667
Chicago/Turabian StyleAdamczuk, Małgorzata. 2022. "Spatial Distribution of Cladocera in a Stratified Palaearctic Lake" Water 14, no. 17: 2667. https://doi.org/10.3390/w14172667
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.