
Artemia spp. (Crustacea, Anostraca) in Crimea: New Molecular Genetic Results and New Questions without Answers
by
 , and
, and
Anastasia Lantushenko
1, 
Yakov Meger
1,
Alexandr Gadzhi
1,
Elena Anufriieva
1,2,* and
Nickolai Shadrin
1,2 1
Sevastopol State University, 33 Universitetskaya Street, 299053 Sevastopol, Russia
2
A.O. Kovalevsky Institute of Biology of the Southern Seas of RAS, 2 Nakhimov Ave., 299011 Sevastopol, Russia
*
Author to whom correspondence should be addressed.
Water 2022, 14(17), 2617; https://doi.org/10.3390/w14172617
Submission received: 10 July 2022
/
Revised: 2 August 2022
/
Accepted: 23 August 2022
/
Published: 25 August 2022
(This article belongs to the Special Issue Ecosystems of Inland Saline Waters)
Abstract
:Many works have been devoted to the study of the molecular genetic diversity of Artemia in different regions; however, there are regions such as Crimea, the largest peninsula in the Black Sea, which has seen few studies. Artemia specimens from several Crimean hypersaline lakes were analyzed using the mitochondrial marker cytochrome oxidase C (COI). The analyzed individuals from bisexual populations formed clades with the species A. salina, A. urmiana, A. sinica, and A. monica (=A. franciscana). A. sinica and A. monica had not been recorded in Crimea previously. In Lake Adzhigol, the three species A. urmiana, A. sinica, and A. monica were found at the same time, which has not been noted anywhere before. In the Crimean lakes, a total of 10 haplotypes were found, six of them for the first time: Once for A. monica, once for A. sinica, and four for A. salina. Those haplotypes may be regarded as endemic to Crimea. In the 1990s, experiments were carried out in Lake Yanyshskoe using mainly purchased cysts of Artemia, so A. monica and A. sinica were introduced into Crimea and could then have easily been spread by birds to other Crimean lakes.
1. Introduction
Other than in Antarctica, Artemia spp. are the most common and abundant animals in hypersaline waters worldwide [1,2]. They belong to Anostraca, the most primitive and ancient group among living crustaceans and have one of the most advanced osmoregulation systems among all animals, which allows them to exist in an extremely wide range of salinity [3,4]. Due to this, they play a key multidimensional role in most ecosystems of hypersaline waters of the planet [1,5,6,7,8]. The existence of several water bird species depends on the development of Artemia populations [9,10,11]. Artemia biomass and its cysts are of great commercial value [1,12]. These crustaceans are also considered convenient test objects in ecotoxicology [13,14], as well as model species to study various issues in different branches of biology [3,4,15,16].
It is therefore not difficult to understand the existing theoretical and practical interest in the study of Artemia, including the study of its diversity and the factors that determine it [5,6,8,17]. Recent studies show that, along with parthenogenetic populations, there are five species of bisexual Artemia in the world: A. salina (Linnaeus, 1758), A. urmiana Günther, 1890, A. monica Verrill, 1869 (=A. franciscana Kellogg, 1906), A. sinica Cai, 1989, and A. persimilis Piccinelli and Prosdocimi, 1968. Regarding the species A. monica and A. franciscana, there is currently no consensus, as some researchers believe that both species are valid while others believe that this is one species.
Many works have been devoted to the study of the molecular genetic diversity of Artemia in different regions [5,18,19]; however, there are still some practically unexplored regions. One of these is Crimea, the largest peninsula in the Black Sea (27,000 km2). The existence of Artemia in Crimea, thanks to P. Pallas, was already determined in the 18th century [20]. In the 19th century, there were four different species described, including the species A. salina, A. arietina Fischer, 1851, A. milhausenii Fischer de Waldheim, 1834, and A. koeppeniana Fischer, 1851 [21,22]. A. arietina is now recognized as a variety of A. salina, and A. milhausenii and A. koeppeniana are recognized as synonyms of A. urmiana [8].
In the second half of the 19th century, it was experimentally shown that salinity causes a high level of Artemia morphological variability [23,24,25], and, proceeding from this, all Artemia species in Crimea were reduced to one species, A. salina [26]. A revision of the diversity of Artemia in Crimea using electron microscopy showed that bisexual brine shrimp on the peninsula mainly belong to the species A. salina, but several males in Lake Sasyk-Sivash belonged to another species [27]. Later, another species was found in Lake Koyashskoe, identified by morphological characteristics as A. urmiana [28], which was confirmed using molecular genetic methods [29]. Previously, based on morphological similarity, it was suggested that A. mulhausinii corresponds to A. urmiana described from Lake Urmia [30]. Crimea was regarded as unique due to having a relatively small territory, and its hypersaline lakes host at least two bisexual Artemia species and their parthenogenetic populations [30]. So, the conclusion was made that Crimea may be considered a remnant of the center of the Artemia biodiversity origin near the ancient Tethys Ocean [30,31]. To date, it is known that Artemia populations exist in Crimea in more than fifty water bodies, including Bay Sivash, the world’s largest Artemia habitat (2560 km2), which are represented by two bisexual native species and parthenogenetic populations of different ploidy [7,32,33,34]. Nevertheless, the existence of only one bisexual species, A. urmiana, was confirmed using the molecular genetic approach [29].
The main objectives of this study are (1) to analyze Artemia specimens from different lakes of Crimea using the mitochondrial marker cytochrome oxidase C (COI), and (2) to test the hypotheses about the existence of at least two bisexual Artemia species on the peninsula and the possibility of coexistence of two bisexual Artemia species in one water body.
2. Materials and Methods
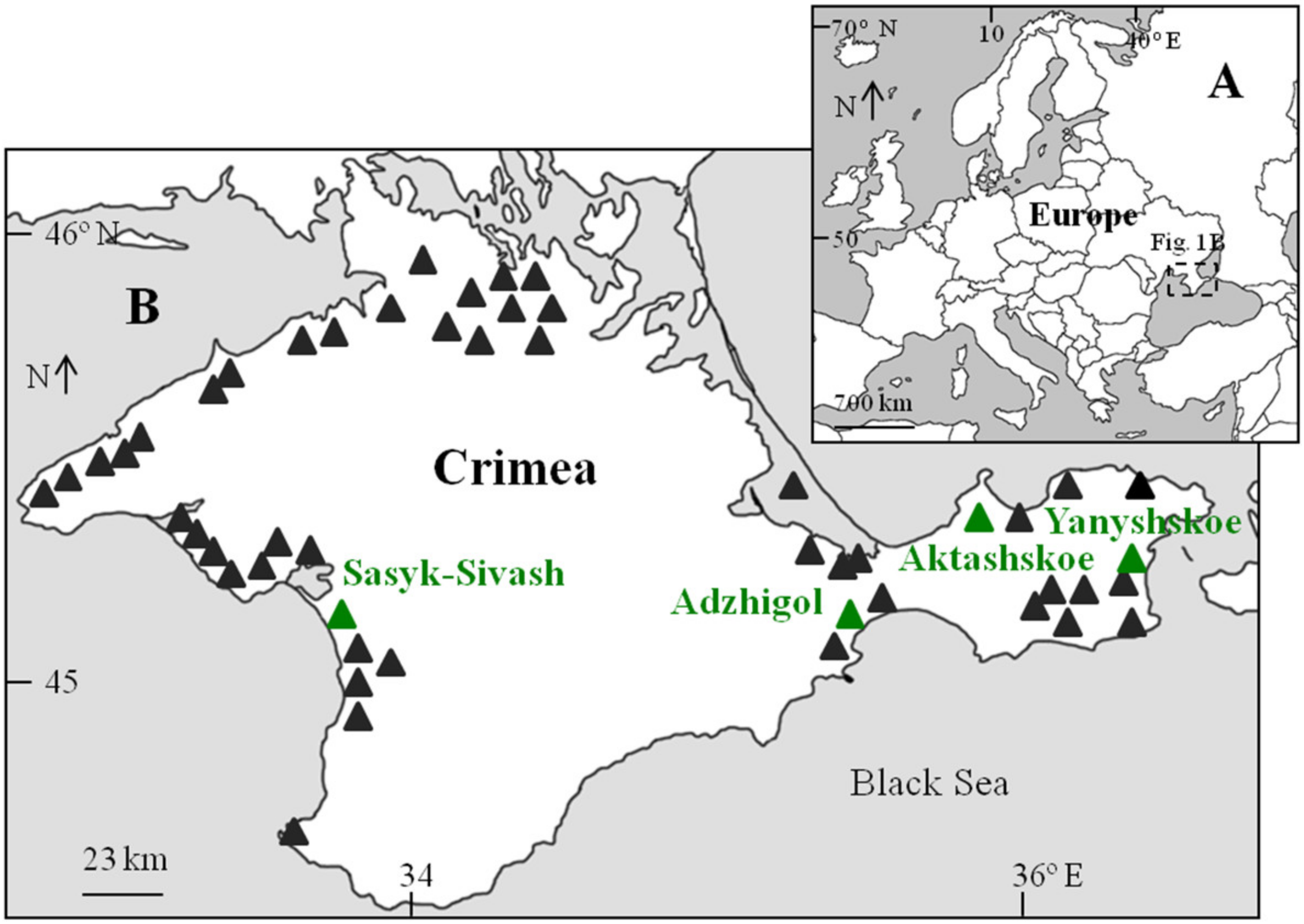
On the Crimean Peninsula, there are many hypersaline water bodies (Figure 1), which differ in size, ranges of salinity fluctuations, and biological diversity [35,36].
All lakes are shallow (up to 1.5 m deep), polymyxic, and characterized by high seasonal and long-term variability. In this study, 14 specimens of bisexual Artemia from three lakes were analyzed, the general characteristics of which are given in Table 1. Sampling was carried out by standard methods, by filtering water through a plankton net [34,35]. Live crustaceans were delivered to the laboratory. Simultaneously with sampling in lakes, water salinity and temperature were measured using a WZ212 portable refractometer (Kelilong Electron Co. Ltd., Fuan, China) and a PHH-830 electronic pH meter (OMEGA Engineering Inc., Norwalk, CT, USA), respectively.
Before DNA isolation, crustaceans were placed overnight in distilled water, then the intestines were removed from each individual, and DNA was isolated from each sample. DNA isolation was performed using a DNA-Extran 2 reagent kit (Sintol, Russia) according to the manufacturer’s instructions. Quantitative determination of the obtained genomic DNA and assessment of its purity were carried out on an Inplen nanophotometer (Inplen, Munich, Germany) using gel electrophoresis in 1% agarose gel. The PCR reaction was carried out using primer pairs jgLCOI490 and jgHCO2198 for the COI gene [37]. The PCR reaction was carried out in a volume of 25 µL using ScreenMix reagents (Evrogen, Moscow, Russia) and consisted of the following steps: 94 °C—2 min, 30 cycles (94 °C—1 min, 48 °C—1 min, 72 °C—1 min), and a final elongation of 5 min at 72 °C. The sequencing of the obtained fragments was carried out on the NANOFOR-05 sequencer (Sintol, Moscow, Russia) at the Center for Collective Use “Molecular Structure of Matter” of the Sevastopol State University. The generated DNA sequences were stored at GenBank (accession numbers ON872198-ON872211) and compared with those available in the National Center for Biotechnology Information (NCBI) database. The analysis used a large dataset containing bisexual and parthenogenetic Artemia sequences from all geographic locations. GenBank codes for sequences previously obtained by other researchers [6,38,39,40,41,42,43,44] and used in the work are presented in Table A1. Phylogenetic reconstruction was performed using a Bayesian Inference approach implemented in MrBayes version 3.2.6 [45]. When constructing a phylogenetic tree, the HQ972028 Daphnia tenebrosa sequence for the CO1 gene was used as an outgroup.
3. Results
The analyzed individuals from bisexual populations formed clades with the species A. salina, A. urmiana, A. sinica, and A. monica (Figure 2). In Lake Sasyk-Sivash, among the analyzed individuals, only A. salina was found, while in Lake Aktashskoe, representatives of two species were found, A. urmiana and A. monica, and in Lake Adzhigol, three species of A. urmiana, A. sinica, and A. monica. Contemporaneously, bisexual individuals of A. urmiana from lakes Adzhigol and Aktashskoe formed a common clade with parthenogenetic populations. In the Crimean lakes, a total of 10 haplotypes were found during this study (Table 2). Two haplotypes (H2 and H3) were shared between parthenogenetic and bisexual individuals of A. urmiana, (3W2, 3W3, and 4W2). The rather high nucleotide variability of the COI genes of samples from Crimea was noted: A. salina specimens (H7-10) from Sasyk-Sivash Lake formed separate haplotypes, and A. sinica samples from Adzhigol Lake were also not included in the haplotypes of previously studied samples and formed a separate (H6) group. Two sequences of A. monica from Lake Adzhigol form joint haplotypes with other A. monica (H4 and H5), and one is allocated to a separate H1 group.
4. Discussion
The obtained data show the presence of four bisexual Artemia species in Crimea, and such value is very high for so small an area as Crimea. In general, in the hypersaline water bodies of the Western Mediterranean region, all these species have also been previously noted but in a much larger area [8]. This fact supports the suggestion that Crimea may be regarded as one of Artemia biodiversity hotspots [30]. Individuals of three bisexual species were simultaneously found in Lake Adzhigol, something that has never been noted anywhere before.
How and when did A. monica and A. sinica appear in Crimea? This is a question that can hardly be answered unambiguously. A. monica (=A. franciscana) is known as a highly invasive species, having its native range in the Americas and currently found in Australia, Asia, Europe, and Africa [6,19,46,47,48]. Displacing native bisexual and parthenogenetic populations of Artemia, this species is rapidly expanding its presence on all continents except Antarctica. The main vector of distribution of A. monica is the widespread use of cysts, which were initially harvested mainly in American water bodies, in aquaculture of fish and shrimp [12,19]. Pond cultivation of A. monica has begun in several regions [12,19], which significantly accelerates their expansion into new territories. After a species enters a new region, its cysts within it are rapidly spread by birds over thousands of kilometers [9,49,50]. In the 1990s, experiments were carried out in Lake Yanyshskoe using mainly purchased cysts of A. monica, so the species could have entered Crimea, and later could have been easily spread by birds to other Crimean lakes.
However, one of the finds makes it doubtful that the species could have been brought to Crimea only by humans, and other scenarios for its entry into Crimea are unbelievable. Near Lake Adzhigol, where the species was found, at a distance of 2–3 km, there is another lake, Kuchuk-Adzhigol (salinity 5–7 g L−1), where three species of cyclops from Southeast Asia were previously found, transported here by birds [35]. Both of these lakes are intensively used by some aquatic bird species making various migrations, and this fact does not preclude the idea that Artemia cysts were also transported here from outside Crimea by birds. Nevertheless, looking at Figure 2 the authors also can assume an earlier migration of the species into Crimea (tens to hundreds of thousands of years ago). In this case, it is impossible to imagine any other way for the species to enter Crimea, except by an accidental introduction by birds.
Once in a new region, A. monica begins to change rapidly, adapting to the conditions of the new region [18,19,51,52]. The rapid variability and adaptability of A. monica under new conditions are facilitated by the fact that the species has different alternative gene expression patterns [16,53]. So, the existence of the alternative patterns provides the possibility to shift from one homeostatic strategy to another in a novel environment, and this may enhance the invasiveness and fitness of the species in the new habitat. Based on this, it can be assumed that, most likely, the species was brought to Crimea rather recently by humans or birds.
It is highly likely that one of those two scenarios can be assumed for A. sinica. Its cysts could be among those purchased and used in Lake Yanyshskoe by fish farmers. However, the possibility of transport by birds cannot be ruled out. At the same time, of course, it is difficult to imagine that the same birds, within the framework of one migration, brought cysts directly from China to Crimea. One can easily imagine that the transport was carried out in the form of a kind of relay race by different birds, for example, through Transbaikalia (between China and Crimea), where A. sinica was also found [54]. Relatively recent finds of A. sinica, thanks to molecular genetic studies, in the West Mediterranean [8], allow the authors to suggest other possible ways for the species to enter Crimea.
Ten Artemia haplotypes were found in Crimea and 77 haplotypes globally [6,38,39,40]. Among the 10 haplotypes found in Crimea, 6 were found for the first time: One for A. monica, one for A. sinica, and four for A. salina. Those haplotypes may be regarded as endemic to Crimea. This fact may also be explained in two ways: First, the introduction of A. monica and A. sinica occurred before the 1990s, or second, those species evolved quickly in Crimea as was shown for other regions [19].
Where did A. monica and A. sinica appear earlier in the Mediterranean or Crimea? How did each species first enter Europe? At present, there are no answers to these questions; new, deeper studies of both the genetic structure of local populations and bird migrations are needed. Another question, which is likely difficult to answer without answering the previous ones is how long have all four bisexual species coexisted, and will they continue to coexist for a long time in Crimea? The large number and variety of hypersaline water bodies, as well as their high seasonal and interannual variability, only suggest a possibility of long-term coexistence. New comprehensive studies on the Crimean Artemia populations are needed to find answers to these questions.
Author Contributions
Conceptualization, N.S.; methodology, A.L., N.S. and E.A.; field investigation and sampling, N.S., E.A. and A.L.; sample processing, species identification, molecular genetics study E.A., A.L., Y.M. and A.G.; data formal analysis, N.S., A.L., Y.M. and E.A.; writing—original draft preparation, N.S. and A.L.; writing—review and editing, N.S., A.L., E.A., Y.M. and A.G. All authors have read and agreed to the published version of the manuscript.
Funding
Field research was carried out within the framework of the state program of the A.O. Kovalevsky Institute of Biology of the Southern Seas of RAS (121041500203-3), and the molecular genetic part of the work and manuscript writing were carried out within the program “Prioritet-2030” of Sevastopol State University (strategic project No. 3, 121121700318-1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used in this study are available upon request from the corresponding author.
Acknowledgments
The authors are grateful to Bindy Datson for her selfless work in improving the English of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
Table A1.
The list of COI haplotypes for all Artemia populations analyzed in the present study (Figure 2). The Crimea populations studied in this work are highlighted in bold.
Table A1.
The list of COI haplotypes for all Artemia populations analyzed in the present study (Figure 2). The Crimea populations studied in this work are highlighted in bold.
| Haplotypes | Species | GenBank Number | Geographical Locality | Reference |
|---|---|---|---|---|
| H1 | A. franciscana | ON872200 | Crimea (Aktashskoe) | this work |
| H2 | A. urmiana | ON872198 | Crimea (Aktashskoe) | this work |
| A. parthenogenetica | KF691333-337,343,345,357,358,359,361, | Iran | [6] | |
| A. parthenogenetica | KF691530-532 | Turkmenistan | [6] | |
| H3 | A. urmiana | ON872199, ON872202 | Crimea (Aktashskoe, Adzhjigol) | this work |
| A. parthenogenetica | KF691148-153,166-172,187-189,208-212,224-226,233-236,238,287-290, | China | [6] | |
| A. parthenogenetica | KF691338-342,344,346,348 | Iran | [6] | |
| A. parthenogenetica | KF691373-375 | Iraq | [6] | |
| A. parthenogenetica | KF691391-434 | Kazakhstan | [6] | |
| A. parthenogenetica | KF691442-448 | Pakistan | [6] | |
| A. parthenogenetica | KF691455,456,458-461,465,467-475,477,478,480,485-491,493,495-497 | Russia | [6] | |
| A. parthenogenetica | KF691534 | Turkmenistan | [6] | |
| A. parthenogenetica | KF691548-553,555 | Uzbekistan | [6] | |
| H4 | A. franciscana | ON872204 | Crimea (Adzhjigol) | this work |
| A. franciscana | KF691154-156,174-175,176,179,181,184,185,186,190,192,206,222,223,231,232,252,255,258,261,264,266,267,281,292,303,315, | China | [6] | |
| A. franciscana | KF691351,353,355, | Iran | [6] | |
| A. franciscana | KF691439-441 | Pakistan | [6] | |
| A. franciscana | KF691508 | Sri Lanka | [6] | |
| A. franciscana | KF691568 | Vietnam | [6] | |
| H5 | A. franciscana | ON872205 | Crimea (Adzhjigol) | this work |
| A. franciscana | KF691191,205,239,240,242-244,250,251,253,259,260,262,263,278,280,294,295,296,297,304,305,307,308,309-314, | China | [6] | |
| A. franciscana | KF691378-382 | Iraq | [6] | |
| A. franciscana | KF691449-454 | Portugal | [6] | |
| A. franciscana | KF691503-507 | Sri Lanka | [6] | |
| A. franciscana | KF691556-567 | Vietnam | [6] | |
| H6 | A. sinica | ON872201, ON872203 | Crimea (Adzhjigol) | this work |
| H7 | A. salina | ON872208 | Crimea (Sasyk-Sivash) | this work |
| H8 | A. salina | ON872209, ON872210, ON872211 | Crimea (Sasyk-Sivash) | this work |
| H9 | A. salina | ON872206 | Crimea (Sasyk-Sivash) | this work |
| H10 | A. salina | ON872207 | Crimea (Sasyk-Sivash) | this work |
| H11 | A. franciscana | DQ119645 | USA | [40] |
| H12 | A. sinica | DQ119650 | Mongolia | [40] |
| H13 | A. urmiana | JX512748,755,758,762,766,769,771,774,776,778,780,783,788,790,795,796,803,804,808 | Iran (Urmia) | [38] |
| H14 | A. parthenogenetica | DQ426826 | Spain | [40] |
| H15 | A. salina | DQ426831,853,857 | Spain | [40] |
| H16 | A. salina | DQ426832, KF691502 | Spain | [40] |
| H17 | A. salina | DQ426833 | Spain | [40] |
| H18 | A. salina | DQ426834,856 | Spain | [40] |
| H19 | A. salina | DQ426836 | Spain | [40] |
| H20 | A. salina | DQ426837 | Spain | [40] |
| H21 | A. salina | DQ426841 | Spain | [40] |
| H22 | A. salina | DQ426845 | Spain | [40] |
| H23 | A. salina | DQ426846 | Spain | [40] |
| H24 | A. salina | DQ426847 | Spain | [40] |
| H25 | A. salina | DQ426848 | Spain | [40] |
| H26 | A. salina | DQ426858 | Spain | [40] |
| H27 | A. sinica | EF615592 | China | [40] |
| H28 | A. salina | EU543444 | Spain | [40] |
| H29 | A. salina | EU543445 | Spain | [40] |
| H30 | A. salina | EU543448 | Spain | [40] |
| H31 | A. salina | EU543452 | Morocco | [40] |
| H32 | A. salina | EU543453 | Morocco | [40] |
| H33 | A. salina | EU543456 | Tunisia | [40] |
| H34 | A. salina | EU543457 | Tunisia | [40] |
| H35 | A. salina | EU543467 | Algeria | [40] |
| H36 | A. salina | EU543468 | Algeria | [40] |
| H37 | A. salina | EU543470 | Egypt | [40] |
| H38 | A. salina | EU543480 | Italy | [40] |
| H39 | A. salina | EU543481 | Italy | [40] |
| H40 | A. salina | EU543485 | South Africa | [40] |
| H41 | A. salina | GU248381 | Italy | [41] |
| H42 | A. sinica | HM998990 | China | [42] |
| H43 | D. tenebrosa | HQ972028 | - | - |
| H44 | A. urmiana | JX512751 | Iran (Urmia) | [38] |
| H45 | A. urmiana | JX512756,764, | Iran (Urmia) | [38] |
| H46 | A. urmiana | JX512775,791,805 | Iran (Urmia) | [38] |
| H47 | A. urmiana | JX512777,801 | Iran (Urmia) | [38] |
| H48 | A. urmiana | JX512782 | Iran (Urmia) | [38] |
| H49 | A. urmiana | JX512792 | Iran (Urmia) | [38] |
| H50 | A. franciscana | KF691137-141 | Canada | [6] |
| H51 | A. franciscana | KF691143-147 | Cape Verde | [6] |
| H52 | A. parthenogenetica | KF691159,257 | China | [6] |
| H53 | A. franciscana | KF691160-165,173,177,178,180,182,207,227-230, 237,241,256,279,282-286,291,293 | China | [6] |
| A. franciscana | KF691328-332 | India | [6] | |
| A. franciscana | KF691347,349,354,356, | Iran | [6] | |
| A. franciscana | KF691376,377,381,383, | Iraq | [6] | |
| A. franciscana | KF691384-390 | Jamaica | [6] | |
| A. franciscana | KF691535,537,538,543,544,546 | USA | [6] | |
| H54 | A. parthenogenetica | KF691183 | China | [6] |
| A. parthenogenetica | KF691462 | Russia | [6] | |
| H55 | A. franciscana | KF691196,219 | China | [6] |
| H56 | A. parthenogenetica | KF691199-204,265,268, | China | [6] |
| H57 | A. sinica | KF691270,271 | China | [6] |
| H58 | A. sinica | KF691272 | China | [6] |
| H59 | A. sinica | KF691274,276,277,300,302 | China | [6] |
| H60 | A. sinica | KF691275,299 | China | [6] |
| H61 | A. franciscana | KF691306 | China | [6] |
| H62 | A. franciscana | KF691320,322 | Columbia | [6] |
| H63 | A. parthenogenetica | KF691360,367,369-372 | Iran | [6] |
| H64 | A. franciscana | KF691435,437,438 | Mexico | [6] |
| H65 | A. parthenogenetica | KF691457 | Russia | [6] |
| H66 | A. parthenogenetica | KF691464,466,476 | Russia | [6] |
| H67 | A. parthenogenetica | KF691479,482,484,492,494 | Russia | [6] |
| H68 | A. parthenogenetica | KF691481,483 | Russia | [6] |
| H69 | A. parthenogenetica | KF691520,522-525,528,529 | Turkey | [6] |
| H70 | A. franciscana | KF691536,540,542,545 | USA | [6] |
| A. franciscana | KJ863431,438,439,443,455,460,466,467,471,474,479,481,482,487,489 | - | [38] | |
| H71 | A. franciscana | KF691539 | Canada | [6] |
| A. franciscana | KJ863454 | USA | [38] | |
| H72 | A. urmiana | KF707695 | Iran | [43] |
| H73 | A. sinica | KF707886-889 | China | [43] |
| H74 | A. franciscana | KJ863484 | USA | [38] |
| H75 | A. parthenogenetica | KU053797-802 | Djarylpach | [39] |
| A. parthenogenetica | KU053803-807 | Sakskoye | [39] | |
| A. parthenogenetica | KU053808-818 | Dzharylhach | [39] | |
| H76 | A. parthenogenetica | KU053811,814 | Ukraine | [39] |
| H77 | A. sinica | LC195586 | Mongolia | [44] |
References
- Abatzopoulos, T.J.; Beardmore, J.; Clegg, J.S.; Sorgeloos, P. Artemia: Basic and Applied Biology; Springer Science & Business Media: Dordrecht, The Netherlands, 2013; 286p. [Google Scholar]
- Sainz-Escudero, L.; López-Estrada, E.K.; Rodríguez-Flores, P.C.; García-París, M. Settling taxonomic and nomenclatural problems in brine shrimps, Artemia (Crustacea: Branchiopoda: Anostraca), by integrating mitogenomics, marker discordances and nomenclature rules. PeerJ 2021, 9, e10865. [Google Scholar] [CrossRef] [PubMed]
- Gajardo, G.O.; Beardmore, J.A. Coadaptation: Lessons from the brine shrimp Artemia, “the aquatic Drosophila” (Crustacea; Anostraca). Rev. Chil. Hist. Nat. 2001, 74, 65–72. [Google Scholar] [CrossRef]
- Gajardo, G.M.; Beardmore, J.A. The brine shrimp Artemia: Adapted to critical life conditions. Front. Physiol. 2012, 3, 185. [Google Scholar] [CrossRef] [PubMed]
- Baxevanis, A.D.; Maniatsi, S.; Kouroupis, D.; Marathiotis, K.; Kappas, I.; Kaiser, H.; Abatzopoulos, T.J. Genetic identification of South African Artemia species: Invasion, replacement and co-occurrence. J. Mar. Biolog. Assoc. 2014, 94, 775–785. [Google Scholar] [CrossRef]
- Eimanifar, A.; Van Stappen, G.; Marden, B.; Wink, M. Artemia biodiversity in Asia with the focus on the phylogeography of the introduced American species Artemia franciscana Kellogg, 1906. Mol. Phylogenet. Evol. 2014, 79, 392–403. [Google Scholar] [CrossRef]
- Shadrin, N.; Yakovenko, V.; Anufriieva, E. Suppression of Artemia spp. (Crustacea, Anostraca) populations by predators in the Crimean hypersaline lakes: A review of the evidence. Int. Rev. Hydrobiol. 2019, 104, 5–13. [Google Scholar] [CrossRef]
- Sainz-Escudero, L.; López-Estrada, E.K.; Rodríguez-Flores, P.C.; García-París, M. Brine shrimps adrift: Historical species turnover in Western Mediterranean Artemia (Anostraca). Biol. Invasions 2022, 24, 2477–2498. [Google Scholar] [CrossRef]
- Green, A.J.; Sánchez, M.I.; Amat, F.; Figuerola, J.; Hontoria, F.; Ruiz, O.; Hortas, F. Dispersal of invasive and native brine shrimps Artemia (Anostraca) via waterbirds. Limnol. Oceanogr. 2005, 50, 737–742. [Google Scholar] [CrossRef]
- Muñoz, J.; Amat, F.; Green, A.J.; Figuerola, J.; Gomez, A. Bird migratory flyways influence the phylogeography of the invasive brine shrimp Artemia franciscana in its native American range. PeerJ 2013, 1, e200. [Google Scholar]
- Redón, S.; Gajardo, G.; Vasileva, G.P.; Sánchez, M.I.; Green, A.J. Explaining variation in abundance and species diversity of avian cestodes in brine shrimps in the Salar de Atacama and other Chilean wetlands. Water 2021, 13, 1742. [Google Scholar] [CrossRef]
- Van Stappen, G.; Sui, L.; Hoa, V.N.; Tamtin, M.; Nyonje, B.; de Medeiros Rocha, R.; Sorgeloos, P.; Gajardo, G. Review on integrated production of the brine shrimp Artemia in solar salt ponds. Rev. Aquac. 2020, 12, 1054–1071. [Google Scholar] [CrossRef]
- Nunes, B.S.; Carvalho, F.D.; Guilhermino, L.M.; Van Stappen, G. Use of the genus Artemia in ecotoxicity testing. Environ. Pollut. 2006, 144, 453–462. [Google Scholar] [CrossRef] [PubMed]
- El Fels, L.; Hafidi, M.; Ouhdouch, Y. Artemia salina as a new index for assessment of acute cytotoxicity during co-composting of sewage sludge and lignocellulose waste. Waste Manag. 2016, 50, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Lenormand, T.; Nougué, O.; Jabbour-Zahab, R.; Arnaud, F.; Dezileau, L.; Chevin, L.M.; Sánchez, M.I. Resurrection ecology in Artemia. Evol. Appl. 2018, 11, 76–87. [Google Scholar] [CrossRef]
- De Vos, S.; Rombauts, S.; Coussement, L.; Dermauw, W.; Vuylsteke, M.; Sorgeloos, P.; Clegg, J.S.; Nambu, Z.; Van Nieuwerburgh, F.; Norouzitallab, P.; et al. The genome of the extremophile Artemia provides insight into strategies to cope with extreme environments. BMC Genom. 2021, 22, 635. [Google Scholar] [CrossRef]
- Kappas, I.; Baxevanis, A.D.; Maniatsi, S.; Abatzopoulos, T.J. Porous genomes and species integrity in the branchiopod Artemia. Mol. Phylogenet. Evol. 2009, 52, 192–204. [Google Scholar] [CrossRef]
- Asem, A.; Eimanifar, A.; Li, W.; Wang, P.Z.; Brooks, S.A.; Wink, M. Phylogeography and population genetic structure of an exotic invasive brine shrimp, Artemia Leach, 1819 (Crustacea: Anostraca), in Australia. Aust. J. Zool. 2019, 66, 307–316. [Google Scholar] [CrossRef]
- Thirunavukkarasu, S.; Karunasagaran, G.; Munuswamy, N. Morphometric and phylogenetic analysis of morphotypes in Artemia franciscana Kellogg, 1906 (Crustacea: Anostraca). Reg. Stud. Mar. Sci. 2022, 54, 102411. [Google Scholar] [CrossRef]
- Anufriieva, E.V.; Shadrin, N.V.; Shadrina, S.N. History of research on biodiversity in Crimean hypersaline waters. Arid Ecosyst. 2017, 7, 52–58. [Google Scholar] [CrossRef]
- Fedchenko, G.P. The deposited lump salt and salt lakes of the Caspian and Azov sea basins. Izv. Imper. O-va Lyubit. Estestvozn. Antropol. Etnogr. 1870, 5. (In Russian) [Google Scholar]
- Kulagin, N.M. Fauna of Crimean salt lakes. Izv. Imper. O-va Lyubit. Estestvozn. Antropol. Etnogr. 1888, 50, 430–444. (In Russian) [Google Scholar]
- Schmankewitsch, M.W.J. On the relations of Artemia salina and Artemia mühlhausenii, and on the genus Branchipus. J. Nat. Hist. 1876, 17, 256–258. [Google Scholar] [CrossRef]
- Gajewski, N. Uber die Variabilitat bei Artemia salina. Int. Rev. Hydrobiol. 1922, 10, 139–159. [Google Scholar] [CrossRef]
- Bond, R.M. Observations on Artemia “franciscana” Kellogg, especially on the relation of environment to morphology. Int. Rev. Hydrobiol. 1933, 28, 117–125. [Google Scholar] [CrossRef]
- Voronov, P.M. Reproduction of Artemia salina in saline waters of the Crimea. Zool. Zhurnal 1973, 52, 945–947. (In Russian) [Google Scholar]
- Mura, G.R.; Nagorskaya, L.I. Notes on the distribution of the genus Artemia in the former USSR countries (Russia and adjacent regions). J. Biol. Res. 2005, 4, 139–150. [Google Scholar]
- Shadrin, N.V.; Batogova, E.A.; Belmonte, D.; Moscatello, S.; Litvinchuk, L.F.; Shadrina, S.N. Artemia urmiana Gunther, 1890 (Anostraca, Artemiidae) in Lake Koyashskoe (Crimea, Black Sea) is the first find outside Lake Urmia (Iran). Mor. Ekol. Zhurn. 2008, 7, 30–31. (In Russian) [Google Scholar]
- Abatzopoulos, T.J.; Amat, F.; Baxevanis, A.D.; Belmonte, G.; Hontoria, F.; Maniatsi, S.; Moscatello, S.; Mura, G.; Shadrin, N.V. Updating geographic distribution of Artemia urmiana Günther, 1890 (Branchiopoda: Anostraca) in Europe: An integrated and interdisciplinary approach. Int. Rev. Hydrobiol. 2009, 94, 560–579. [Google Scholar] [CrossRef]
- Belmonte, G.; Moscatello, S.; Batogova, E.A.; Pavlovskaya, T.; Shadrin, N.V.; Litvinchuk, L.F. Fauna of hypersaline lakes of the Crimea (Ukraine). Thalass. Salentina 2012, 34, 11–24. [Google Scholar]
- Shadrin, N.; Anufriieva, E.; Galagovets, E. Distribution and historical biogeography of Artemia Leach, 1819 (Crustacea: Anostraca) in Ukraine. Int. J. Artemia Biol. 2012, 2, 30–42. [Google Scholar]
- Shadrin, N.V.; Anufriieva, E.V.; Amat, F.; Eremin, O.Y. Dormant stages of crustaceans as a mechanism of propagation in the extreme and unpredictable environment in the Crimean hypersaline lakes. Chin. J. Oceanol. Limnol. 2015, 33, 1362–1367. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Anufriieva, E.V. Size polymorphism and fluctuating asymmetry of Artemia (Branchiopoda: Anostraca) populations from the Crimea. J. Sib. Fed. Univ. Biol. 2017, 10, 114–126. [Google Scholar] [CrossRef]
- Anufriieva, E.; Shadrin, N. The long-term changes in plankton composition: Is Bay Sivash transforming back into one of the world’s largest habitats of Artemia sp. (Crustacea, Anostraca)? Aquac. Res. 2020, 51, 341–350. [Google Scholar] [CrossRef]
- Anufriieva, E.; Hołyńska, M.; Shadrin, N. Current invasions of Asian cyclopid species (Copepoda: Cyclopidae) in Crimea, with taxonomical and zoogeographical remarks on the hypersaline and freshwater fauna. Ann. Zool. 2014, 64, 109–130. [Google Scholar] [CrossRef]
- Golubkov, S.M.; Shadrin, N.V.; Golubkov, M.S.; Balushkina, E.V.; Litvinchuk, L.F. Food chains and their dynamics in ecosystems of shallow lakes with different water salinities. Russ. J. Ecol. 2018, 49, 442–448. [Google Scholar] [CrossRef]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Eimanifar, A.; Wink, M. Fine-scale population genetic structure in Artemia urmiana (Günther, 1890) based on mtDNA sequences and ISSR genomic fingerprinting. Org. Divers. Evol. 2013, 13, 531–543. [Google Scholar] [CrossRef]
- Eimanifar, A.; Asem, A.; Djamali, M.; Wink, M. A note on the biogeographical origin of the brine shrimp Artemia urmiana Gunther, 1899 from Urmia Lake, Iran. Zootaxa 2016, 4097, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, J.; Gómez, A.; Green, A.J.; Figuerola, J.; Amat, F.; Rico, C. Phylogeography and local endemism of the native Mediterranean brine shrimp Artemia salina (Branchiopoda: Anostraca). Mol. Ecol. 2008, 17, 3160–3177. [Google Scholar] [CrossRef]
- Maniatsi, S.; Kappas, I.; Baxevanis, A.D.; Farmaki, T.; Abatzopoulos, T.J. Sharp phylogeographic breaks and patterns of genealogical concordance in the brine shrimp Artemia franciscana. Int. J. Mol. Sci. 2009, 10, 5455–5470. [Google Scholar] [CrossRef]
- Maniatsi, S.; Baxevanis, A.D.; Kappas, I.; Deligiannidis, P.; Triantafyllidis, A.; Papakostas, S.; Bougiouklis, D.; Abatzopoulos, T.J. Is polyploidy a persevering accident or an adaptive evolutionary pattern? The case of the brine shrimp Artemia. Mol. Phylogenet. Evol. 2011, 58, 353–364. [Google Scholar] [CrossRef]
- Maccari, M.; Amat, F.; Gomez, A. Origin and genetic diversity of diploid parthenogenetic Artemia in Eurasia. PLoS ONE 2013, 8, e83348. [Google Scholar]
- Naganawa, H.; Mura, G. Two new cryptic species of Artemia (Branchiopoda, Anostraca) from Mongolia and the possibility of invasion and disturbance by the aquaculture industry in East Asia. Crustaceana 2017, 90, 1679–1698. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Ruebhart, D.R.; Cock, I.E.; Shaw, G.R. Invasive character of the brine shrimp Artemia franciscana Kellogg 1906 (Branchiopoda: Anostraca) and its potential impact on Australian inland hypersaline waters. Mar. Freshw. Res. 2008, 59, 587–595. [Google Scholar] [CrossRef]
- Vikas, P.A.; Sajeshkumar, N.K.; Thomas, P.C.; Chakraborty, K.; Vijayan, K.K. Aquaculture related invasion of the exotic Artemia franciscana and displacement of the autochthonous Artemia populations from the hypersaline habitats of India. Hydrobiologia 2012, 684, 129–142. [Google Scholar] [CrossRef]
- Scalone, R.; Rabet, N. Presence of Artemia franciscana (Branchiopoda, Anostraca) in France: Morphological, genetic, and biometric evidence. Aquat. Invasions 2013, 8, 67–76. [Google Scholar] [CrossRef]
- Khomenko, S.V.; Shadrin, N.V. Iranian endemic Artemia urmiana in hypersaline Lake Koyashskoe (Crimea, Ukraine): A preliminary discussion of introduction by birds. Branta. Trans. Azov Black Sea Ornithol. Station 2009, 12, 81–91. [Google Scholar]
- Muñoz, J.; Gómez, A.; Figuerola, J.; Amat, F.; Rico, C.; Green, A.J. Colonization and dispersal patterns of the invasive American brine shrimp Artemia franciscana (Branchiopoda: Anostraca) in the Mediterranean region. Hydrobiologia 2014, 726, 25–41. [Google Scholar] [CrossRef]
- Kappas, I.; Abatzopoulos, T.J.; Van Hoa, N.; Sorgeloos, P.; Beardmore, J.A. Genetic and reproductive differentiation of Artemia franciscana in a new environment. Mar. Biol. 2004, 146, 103–117. [Google Scholar] [CrossRef]
- Ogello, E.O.; Nyonje, B.M.; Van Stappen, G. Genetic differentiation of Artemia franciscana (Kellogg, 1906) in Kenyan coastal saltworks. Int. J. Adv. Res. 2014, 2, 1154–1164. [Google Scholar]
- Lee, J.; Cho, B.C.; Park, J.S. Transcriptomic analysis of brine shrimp Artemia franciscana across a wide range of salinities. Mar. Genom. 2022, 61, 100919. [Google Scholar] [CrossRef] [PubMed]
- Shadrin, N.; Anufriieva, E. Review of the biogeography of Artemia Leach, 1819 (Crustacea: Anostraca) in Russia. Int. J. Artemia Biol. 2012, 2, 51–61. [Google Scholar]
Figure 1.
Distribution of the hypersaline lakes in Crimea, including the location of the studied lake. ((A)—the European scale, (B)—the Crimean scale).
Figure 1.
Distribution of the hypersaline lakes in Crimea, including the location of the studied lake. ((A)—the European scale, (B)—the Crimean scale).
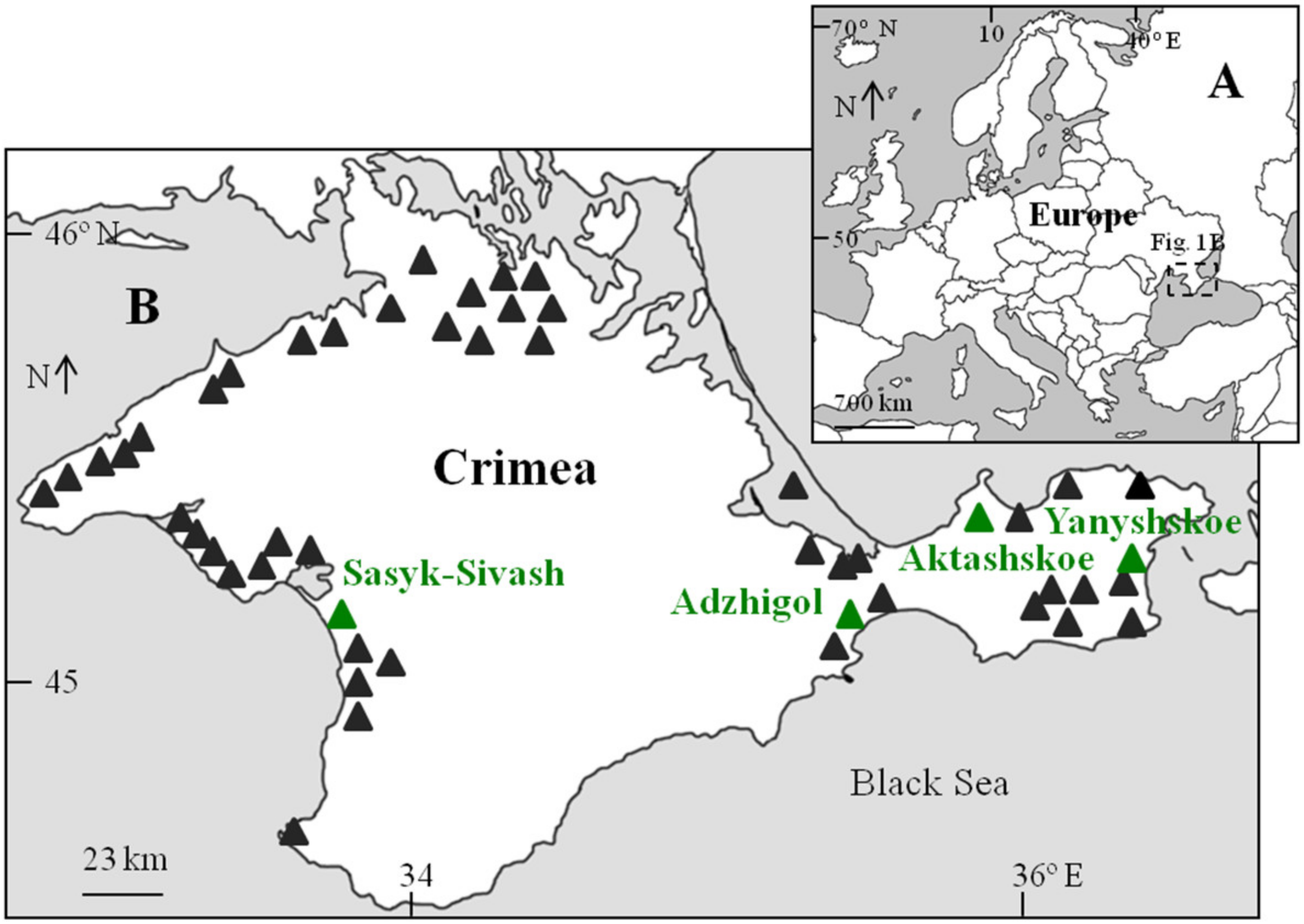
Figure 2.
Phylogenetic relationship of analyzed Artemia revealed by Bayesian analysis based on cytochrome c oxidase subunit I (COI). The bootstrap values at the nodes higher than 75% are shown next to the branches.
Figure 2.
Phylogenetic relationship of analyzed Artemia revealed by Bayesian analysis based on cytochrome c oxidase subunit I (COI). The bootstrap values at the nodes higher than 75% are shown next to the branches.
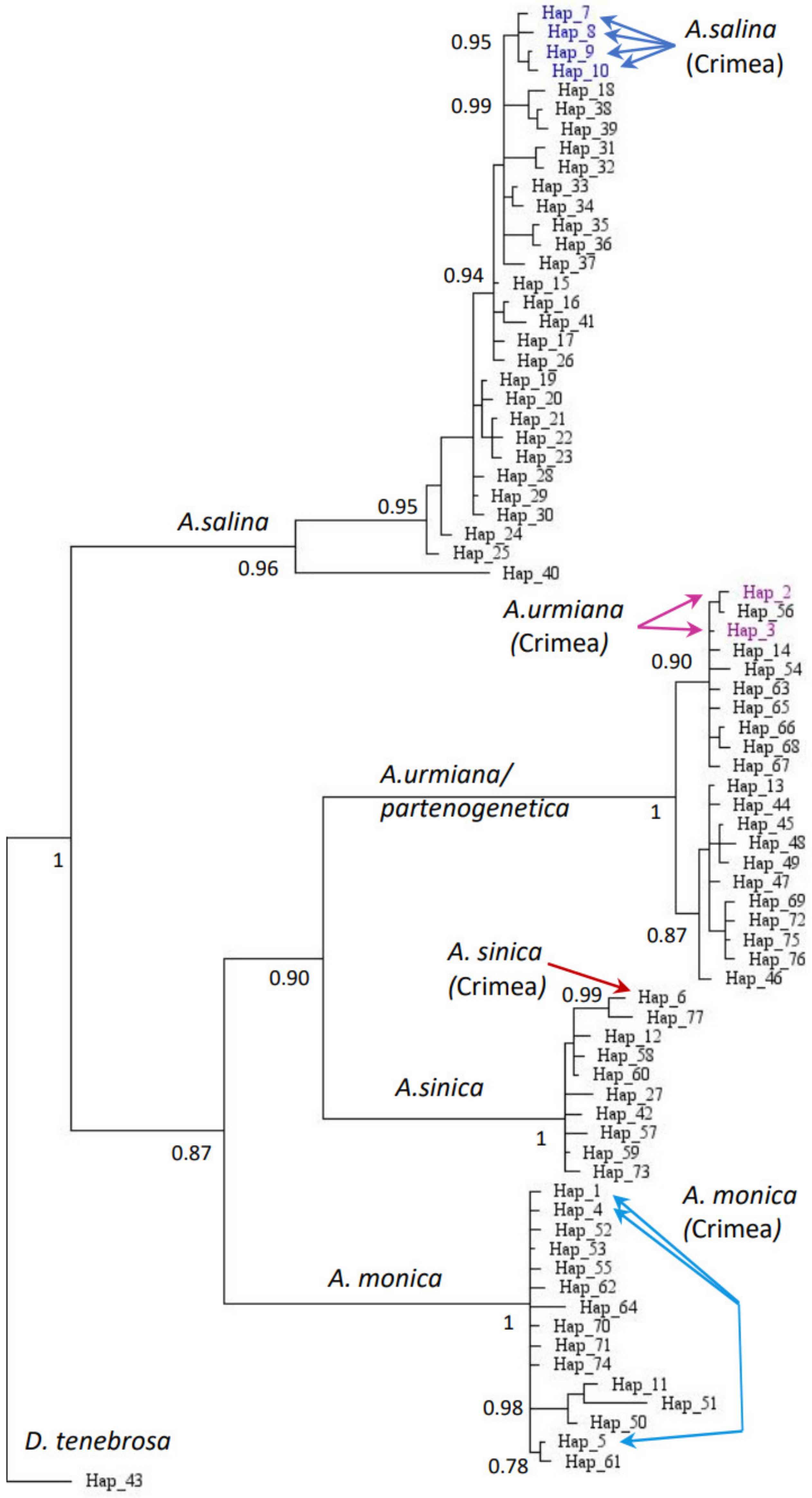
Table 1.
General characteristics of the studied Crimean hypersaline lakes where Artemia was taken.
| Lake | Coordinates | Area, km2 | Date of Sampling | Salinity (during Sampling), g L−1 | Temperature, °C | Number of Analyzed Individuals (Female/Male) | Total Artemia Abundance, Ind. m−3 |
|---|---|---|---|---|---|---|---|
| Aktashskoe | 45°22′31″ N 35°49′45″ E | 26.8 | 1 July 2021 | 173 | 28.5 | A. urmiana—2 f A. monica—1 m | 34,820 |
| Adzhjigol | 45°06′32″ N 35°27′58″ E | 0.6 | 1 July 2021 | 50 | 34.5 | A. monica—2 m A. sinica—2 f A. urmiana—1 f | 902,960 |
| Sasyk-Sivash | 45°09′21″ N 33°31′09″ E | 75.3 | 3 July 2021 | 245 | 36.0 | A. salina—4 m/2 f | 220 |
Table 2.
Artemia haplotypes found in the Crimean hypersaline lakes.
| Species | Abbreviation/Haplotypes | Lake | GenBank Number |
|---|---|---|---|
| A. monica (=A. franciscana) | 3M2/H1 | Aktashskoe | ON872200 |
| A. urmiana | 3F2/H2 | Aktashskoe | ON872198 |
| A. urmiana | 3F3/H3; 4F2/H3 | Aktashskoe, Adzhigol | ON872199 ON872202 |
| A. monica (=A. franciscana) | 4M1/H4 | Adzhigol | ON872204 |
| A. monica (=A. franciscana) | 4M2/H5 | Adzhigol | ON872205 |
| A. sinica | 4F1/H6; 4F3/H6 | Adzhigol | ON872201 ON872203 |
| A. salina | 5M1/H7 | Sasyk-Sivash | ON872208 |
| A. salina | 5M2/H8,5M3/H8,5M5/H8 | Sasyk-Sivash | ON872209 ON872210 ON872211 |
| A. salina | 5F1/H9 | Sasyk-Sivash | ON872206 |
| A. salina | 5F2/H9 | Sasyk-Sivash | ON872207 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lantushenko, A.; Meger, Y.; Gadzhi, A.; Anufriieva, E.; Shadrin, N. Artemia spp. (Crustacea, Anostraca) in Crimea: New Molecular Genetic Results and New Questions without Answers. Water 2022, 14, 2617. https://doi.org/10.3390/w14172617
AMA Style
Lantushenko A, Meger Y, Gadzhi A, Anufriieva E, Shadrin N. Artemia spp. (Crustacea, Anostraca) in Crimea: New Molecular Genetic Results and New Questions without Answers. Water. 2022; 14(17):2617. https://doi.org/10.3390/w14172617
Chicago/Turabian StyleLantushenko, Anastasia, Yakov Meger, Alexandr Gadzhi, Elena Anufriieva, and Nickolai Shadrin. 2022. "Artemia spp. (Crustacea, Anostraca) in Crimea: New Molecular Genetic Results and New Questions without Answers" Water 14, no. 17: 2617. https://doi.org/10.3390/w14172617
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.