
Dry-Season Fog Water Utilization by Epiphytes in a Subtropical Montane Cloud Forest of Southwest China
by
 , and
, and
Lu-Lu Liu
1,2,3,†, 
Bin Yang
1,2,†,
Hua-Zheng Lu
1,2,
Yi Wu
1,2,
Xian-Jing Meng
4,
Yong-Jiang Zhang
5 and
and
Liang Song
1,2,6,* 1
CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla 666303, China
2
Core Botanical Gardens, Chinese Academy of Sciences, Mengla 666303, China
3
University of Chinese Academy of Sciences, Beijing 100049, China
4
Thermo Fisher Scientific, Shanghai 201206, China
5
School of Biology and Ecology, University of Maine, Orono, ME 04469, USA
6
Ailaoshan Station for Subtropical Forest Ecosystem Studies, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Puer 676209, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2021, 13(22), 3237; https://doi.org/10.3390/w13223237
Submission received: 11 October 2021
/
Revised: 8 November 2021
/
Accepted: 10 November 2021
/
Published: 15 November 2021
(This article belongs to the Special Issue Impacts of Climate Change on Plant Water Use, Carbon Balance, Nutrient Economy, and Their Interactions)
Abstract
:Fog water is generally considered to be an important water source for epiphytes in cloud forests because they cannot directly access ground-level water sources. However, the water use proportions of potential water sources and water use efficiency of epiphytes in the subtropical montane cloud forests (MCF) remain to be further explored. In this study, we investigated the water use pattern in the dry season and the intrinsic water use efficiency () of four epiphyte groups (i.e., epiphytic lichens, epiphytic bryophytes, epiphytic ferns, and epiphytic seed plants) using stable isotope (δ2H, δ18O, and δ13C) techniques. Our results indicated that the water sources of epiphytes were significantly different among groups and species. The contribution proportions of fog water to epiphytic lichens, epiphytic bryophytes, epiphytic ferns, and epiphytic seed plants were 83.2%, 32.7%, 38.8% and 63.7%, respectively. Epiphytic lichens and epiphytic seed plants mainly depended on fog water whereas the epiphytic bryophytes and epiphytic ferns relied on both fog water and humus. This may be due to their differences in morphological and structural traits (e.g., thallus or leaves, rhizoid or roots). Additionally, the difference in was also significant among epiphyte groups and species, which could be related to their different water acquisition patterns. In conclusion, our study reveals the differentiation of water utilization in epiphytes and confirms the importance of fog water for epiphytes during the dry season.
1. Introduction
In the context of global climate change, the frequency, duration, and intensity of drought will increase in many regions of the world [1,2]. There is a widespread concern that such drought events will weaken the current forest carbon sink and threaten the biodiversity of various forest ecosystems [3]. Epiphytes, which grow on living or residual host trees, are one of the most diverse groups in the forest ecosystem. These plants play an important role in maintaining biodiversity and biomass and promoting the processes of nutrient and water cycling [4,5]. Epiphytes lack direct access to the ground soil and rely heavily on atmospheric resources (e.g., water, nutrients), which makes them exceptionally sensitive to changes in environmental conditions and good biological indicators of global climate change [6,7]. Understanding the water use strategies of different epiphytes is critical for correctly predicting the impacts of predicted changing rainfall patterns on them.
Montane cloud forest (MCF) is generally characterized by persistent, frequent incidences of fog and low cloud cover at the canopy level, which usually harbors an abundant epiphyte community [8,9,10]. The role of epiphytes in the water cycle of MCF has been studied in the Neo-tropical regions (e.g., Costa Rica, Guatemala, Mexico and Ecuador) [11,12,13,14]. These studies indicate that epiphytes contribute substantially to canopy dynamics by stabilizing canopy organic soils (i.e., humus) that affect the interception of atmospheric moisture (e.g., fog and rainfall). In the subtropical MCF of Southwest China, also known as the montane moist evergreen broadleaved forest, the diversity of epiphytes is extremely high (>600 species, including epiphytic lichens, epiphytic bryophytes, epiphytic ferns, and epiphytic seed plants) [15,16]. However, a six-month-long dry season lasts from November to the subsequent April, implying that the epiphytes need to overcome seasonal drought stress [7,17]. Moreover, the epiphytic habitats are usually harsh and heterogeneous with significant variations in water availability at small spatiotemporal scales, and short-term drought occurs even in rainy seasons [17,18]. Mechanisms on adaptation strategies of epiphytes to forest canopies with limited and unstable water availability remain one of the most captivating questions in plant ecology [19,20].
Multiple morphological and structural traits allow epiphytes to cope with the water deficits, such as early dormant, water storage stems and leaves, rhizomes, and air moisture absorption [21,22,23]. Recent studies confirm that epiphytes could take up atmospheric water (e.g., water vapor, rainwater, and fog water) directly throughout their surface structures, especially for epiphytic lichens that lack roots and an outer waxy cuticle [24,25]. Atmospheric water enhances the growth of epiphytes by alleviating the desiccation degrees and lengthening their photosynthetic activities [26]. Among them, Liu et al. [13] indicated that the Ficus tinctoria in the epiphytic stage would utilize a combination of fog water and rainwater present in canopy humus. Similarly, Wu et al. [27] found that fog water is a crucial supplemental source (contributing ratio ~87%) to maintain the water budgets of epiphytes during the dry season. To adapt to the water deficits, epiphytes was suggested to exhibit higher intrinsic water use efficiency () than the ground-rooted plants [28]. However, far too little attention has been paid to the quantitative relationship between the potential water sources and the water among different groups of epiphytes.
Epiphytes are usually divided into different groups (namely epiphytic lichens, epiphytic bryophytes, epiphytic ferns, and epiphytic seed plants) based on their systematic positions, life cycles, and biological characteristics. Epiphytic lichens are complex life forms that live in a symbiotic relationship with algae and fungi, which can attach to tree trunks only with the help of thalli base plates [29,30]. Epiphytic bryophytes, with no true roots (only rhizoid) and vascular systems (only rib), usually grow on barks or thick mats of humus [31,32,33]. As vascular plants, however, both epiphytic ferns and epiphytic seed plants have developed true roots and networks of the vascular system, allowing efficient water uptake, water and nutrient transport, and photosynthates [34,35], although roots and vascular systems in the former are much simpler than the latter. Due to these differences in structure, water uptake and transport, different epiphytic groups are expected to show differences in their water sources and water use efficiencies, which has not been well-quantified. Early findings suggested that the of epiphytic bryophytes were lower than the vascular plants [36]. Although the water use of several epiphyte species has been studied, investigating the water use characteristics of different epiphyte groups remains scarce and challenging.
With the potential aggravation of drought predicted by future climate models [37], epiphytes will face more severe water stress in the dry season due to their high sensitivity to water fluctuations [7,38]. Understanding how different epiphytes obtain water and maintain normal life activities in the dry season is urgently needed to predict their potentially different responses to changes in the rainfall pattern. To fill the knowledge gap mentioned above, 12 dominant species belonging to four epiphyte groups were selected from a subtropical MCF in SW China to study their water use at the peak of the dry season. The specific objectives of this study are to quantify the water sources of different epiphyte groups in the dry season based on stable isotope techniques (δ2H and δ18O), and explore the intrinsic water use efficiency () of various epiphyte groups using δ13C. We expected that the water use patterns of epiphytes show group-specific and interspecific differentiations, and fog water plays a crucial role in maintaining the water absorption of epiphytes during the dry season.
2. Materials and Methods
2.1. Study Site
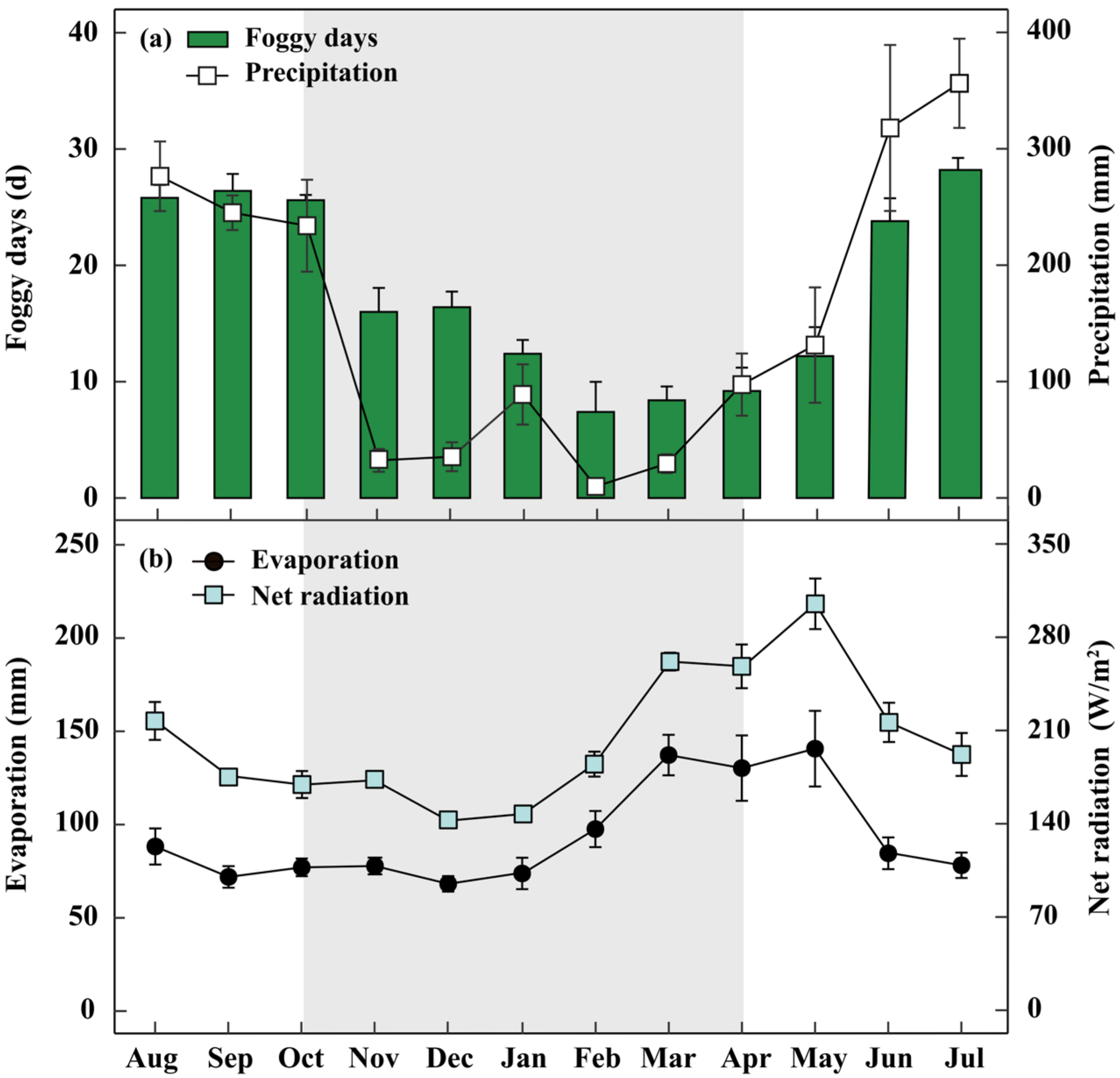
The study was conducted at the Ailaoshan Station for Subtropical Forest Ecosystem Studies (ASSFE, 23°35′–24°44′ N, 100°54′–101°01′ E), located at the Ailao Mountain, Jingdong County, Yunnan Province, SW China [39]. Co-affected by the western monsoon from India-Pakistan and the southeast monsoon, the seasonal distribution of precipitation is uneven, with only approximately 16% of the precipitation occurring in the dry season (November to April) (Figure 1a). The mean annual relative humidity (RH) was 84%, and the mean air temperature was 11.6 °C [9]. However, fogs are frequent in this research region (212 days per year). The foggy days (Visibility < 1000 m, Rainfall = 0 mm) in the dry season are 12 days per month on average [40]. Meanwhile, the average annual evaporation and annual net radiation are 1126 mm and 2444 W/m2, respectively (Figure 1b).
These meteorological data were obtained from the Ailaoshan weather station, which is located near the study site. All data were collected using a suite of micro-meteorological sensors, including air temperature (HMP45, VaisalaInc., Helsinki, Finland), radiation (CNR-1, Kipp and Zonen Inc., Delft, The Netherlands), and rainfall amount (52203, RM Young Inc., Traverse City, MI, USA). An evaporation pan (E-601, Weinasa Inc., Sichuan, China) with 0.62 m in diameter and 0.69 m in height was used to monitor daily evaporation. The water depth in E-601 was 0.6 m and the surface of the water is at ground level. Further details about the site are provided in Song et al. [9] and Zhang, et al. [41]. This region has abundant epiphytic communities, including more than 217 species of epiphytic lichens [42], 176 species of bryophytes [16], 93 species of ferns, and 125 species of seed plants [43,44].
2.2. Isotopic Sampling
Hydrogen (δ2H) and oxygen (δ18O) stable isotopes are widely used to trace the water sources of plants, which rest on the fact that the isotopic composition of plant non-photosynthetic tissues remains unchanged during the root water uptake process and long-distance water transport, until it reaches leaves or non-suberized stems [45,46]. Moreover, the stable isotope of carbon (δ13C) is an effective approach to reflect the intrinsic water use efficiency () of epiphytes [47]. This method is based on the positive correlation between bulk leaf δ13C of plants and their [48].
Because the epiphytes can absorb atmospheric water inputs and humus simultaneously through leaves, roots, and rhizoid (except epiphytic lichens) [13,27], we assumed that sampling water of epiphytes was a combination of rainwater, canopy humus, and intercepted fog. Epiphytes may be subjected to long-term drought stress during the rainless dry season due to their high dependences on atmospheric water inputs. Also, different epiphytic groups may show differences in responding to the seasonal water deficits. Therefore, we studied four groups of epiphytes at the peak of the dry season (from 10 to 23 January 2019) to infer their potential responses to changes in the rainfall pattern. Species used included two epiphytic lichens (Nephromopsis pallescens, Lobaria retigera), four epiphytic bryophytes (Homaliodendron montagneanum, Plagiochila assamica, Bazzania himlayana, Thuidium cymbifolium), four epiphytic ferns (Asplenium indicum, Lepisorus loriformis, Hymenophyllum polyanthos, Loxogramme chinensi), and two epiphytic seed plants (Aeschynanthus buxifolius, Agapetes mannii). All the species are C3 plants, and thus we can use carbon isotopic ratio δ13C to compare their [35].
For different groups of epiphytes, the samples of epiphytic seed plants (n = 4) and epiphytic ferns (n = 4) were collected from the non-photosynthetic basal culm tissue, while the samples of epiphytic bryophytes (n = 4) and epiphytic lichens (n = 4) were collected from shoots and thalli separately. All samples of the epiphyte species were collected from different host trees in the morning (9:00–11:00) from 10 to 23 January, 2019. Considering that the epiphytes grow at different heights of the host tree, 5–10 non-photosynthetic tissues, shoots, or thalli of epiphytes were combined into a single sample and four replicates for each species were collected from the trunk (<3 m) of each host tree. If the numbers of the individuals on the target host trees were insufficient, the epiphyte samples were collected from the neighboring 3–5 trees (within the distance of 5 m of the host tree). After each sampling, liquid water, humus or litter on the surface of the epiphyte samples were gently cleaned with a filter paper one by one carefully. Then the samples (~10 g per sample) were retained and instantly put into 10 mL screw-cap glass vials, sealed with parafilm, frozen (−4 °C) in the portable fridge. After getting back from the field, all samples were moved to the refrigerator instantly until water extraction using cryogenic vacuum distillation [45]. While collecting the plant samples, the humus samples (n = 4) were collected at the sampling site of epiphyte species, except for epiphytic lichens that grow directly on the bark of the host trees. The collection process of humus samples was the same as epiphyte samples.
Fog water and rainwater were collected at the Ailaoshan weather station throughout the period of the experiment for water source determination and throughout 2018 and 2019 for the analysis of the local meteoric water line (LMWL). Fog water was collected using a self-made V-shaped collector (Figure S1a). The collector was set up at 1 m height and a circular rain shield (0.6 m in diameter) was mounted on the top of the fog collector to prevent vertical precipitation (i.e., rainwater) from entering the collector [49]. Fog water samples were collected during a dense fog event between 8:00 to 9:00 before the isotope fractionation happens from re-evaporation [50,51]. Meanwhile, the rainwater was collected using a cylindrical collector measuring 0.65 m in height and 0.2 m in diameter (Figure S1b), with a 15 cm diameter funnel-shaped draining to a 1 L polyethylene bottle. All rainwater samples were collected immediately after a rainfall event or early in the morning following overnight rainfall. During the experiment period, the fog water and rainwater samples were collected daily on days with fog and rain events (a total of seven fog events and five rainfall events occurred) in the study site. All the fog and rain samples were stored immediately in 2 mL screw-cap glass vials, sealed with parafilm, and frozen (–4 °C) in the refrigerator until water extraction using cryogenic vacuum distillation. In addition, samples for all rainfall events between March 2018 to March 2019 were collected after the precipitation events (8:00) to analyze the LMWL.
For each repetition of epiphyte species for δ13C analysis, all healthy and fully expanded mature leaves (10–20) were collected from a host tree on the dates of plant tissue collection. The selected epiphyte samples were collected on the same day of sample collection for water source analysis on the same day. The humus or litter on the surface of the leaves were gently cleaned superficially with a filter paper, oven-dried at 70 °C for 48 h to a constant mass, homogenized and ground to fine powder to pass through a 100–mesh sieve and stored immediately in 2 mL screw-cap glass vials, sealed with parafilm, then subsampled until δ13C analysis in the laboratory [52].
2.3. Isotope Measurements
The liquid water from plant samples and humus (0.5–1 mL per sample) was extracted using the ultra-low temperature (−196 °C) automatic vacuum condensation and extraction system (LI-2100, Lica United Technology Limited Inc., Beijing, China). All samples were filtered using an injection syringe fitted with a filter (pore size 0.22 μm) during this process. Then, the δ2H and δ18O of liquid water (including plant tissues water, humus, fog water and rainwater) were determined using the DELTA-V-Advantage isotope ratio mass spectrometer (Thermo Fisher Scientific, Bremen, Germany) combined with a high-temperature conversion elemental analyzer. To avoid any “memory effect”, each sample was analyzed four times with the last three injections used for calculations [53]. The determination of δ13C in leaf samples was carried out using a flash combustion elemental analyzer (Flash EA) coupled with the DELTA-V-Advantage isotope ratio mass spectrometer (Thermo Fisher Scientific, Bremen, Germany) in the Thermo Fisher Scientific, Shanghai, China. The precision of the δ2H, δ18O, and δ13C measurements were 1‰, ± 0.2‰, and ± 0.15‰, respectively.
Isotope composition (δ2H, δ18O, and δ13C) of unknown samples were addressed to the Vienna-Standard Mean Ocean Water (V-SMOW) and the Vienna-standard Pee Dee Belemnite (V-PDB):
where Rsample is the isotope ratio (2H/1H, 18O/16O) of a water sample, RVSMOW is the isotope ratio (2H/1H, 18O/16O) of the Vienna Standard Mean Ocean Water (VSMOW) standard, Rleaf is the isotope ratio (13C/12C) of plant bulk leaf, RPDB is the isotope ratio (13C/12C) of the Vienna Pee Dee Belemnite (VPDB) standard.
2.4. Statistical Analysis
The MixSIAR model was used to evaluate the relative contribution of potential water sources to the water used by plants [54]. This Bayesian mixing model combines a variety of sources of uncertainty including multiple sources, spatiotemporal variability, and isotope fractionation [55,56]. Regardless of whether multiple sources are logically related to each other, or whether the isotopic signatures were significantly different, the model can accurately calculate the contribution range of each source to the mixture by priori or posteriori aggregation methods [57]. In this study, input data of the MixSIAR model were the individual isotope values (δ2H and δ18O) of epiphytes (i.e., non-photosynthetic tissues, shoots, or thalli of epiphytes in this study, n = 4) and the average and standard deviation (SD) of potential water sources (i.e., fog water: n = 7; humus: n = 4; rainwater: n = 5). The discrimination data were set to zero for both δ2H and δ18O because isotopic fractionation does not occur during plant water uptake process [45]. The humus samples were collected one-to-one correspondence to the epiphyte samples during the sampling process. In consequence of a lack of humus accumulation in their habitats, the humus was excluded from the potential water sources for the epiphytic lichens.
All statistical analysis was performed using the statistical platform R3.6.3 [58]. After checking the normality and the homogeneity of variances using the Shapiro-Wilk test and Bartlett’s test respectively, the assumptions of one-way Analysis of Variance for some data (i.e., the δ2H, δ18O and δ13C of epiphytes, and the δ18O of potential water sources) could not be satisfied even after transformation. Thus, a nonparametric Kruskal-Wallis rank-sum test, followed by the pair-wise Wilcoxon rank-sum test, were used to test for differences of δ2H, δ18O, and δ13C in epiphytes, and the δ2H and δ18O in potential water sources [59]. To test the effects of δ2H and δ18O on δ13C () in different groups of epiphytes (i.e., epiphytic lichens, epiphytic bryophytes, epiphytic ferns, and epiphytic seed plants) linear mixed model (LMM) was used with the R package ‘lme4’ [60], in which the δ2H and δ18O of different groups were treated as fixed effects. The species of each group was incorporated as a random effect to account for the potential influence of different species across the result.
3. Results
3.1. Isotopic Compositions of Water Sources and Epiphytes
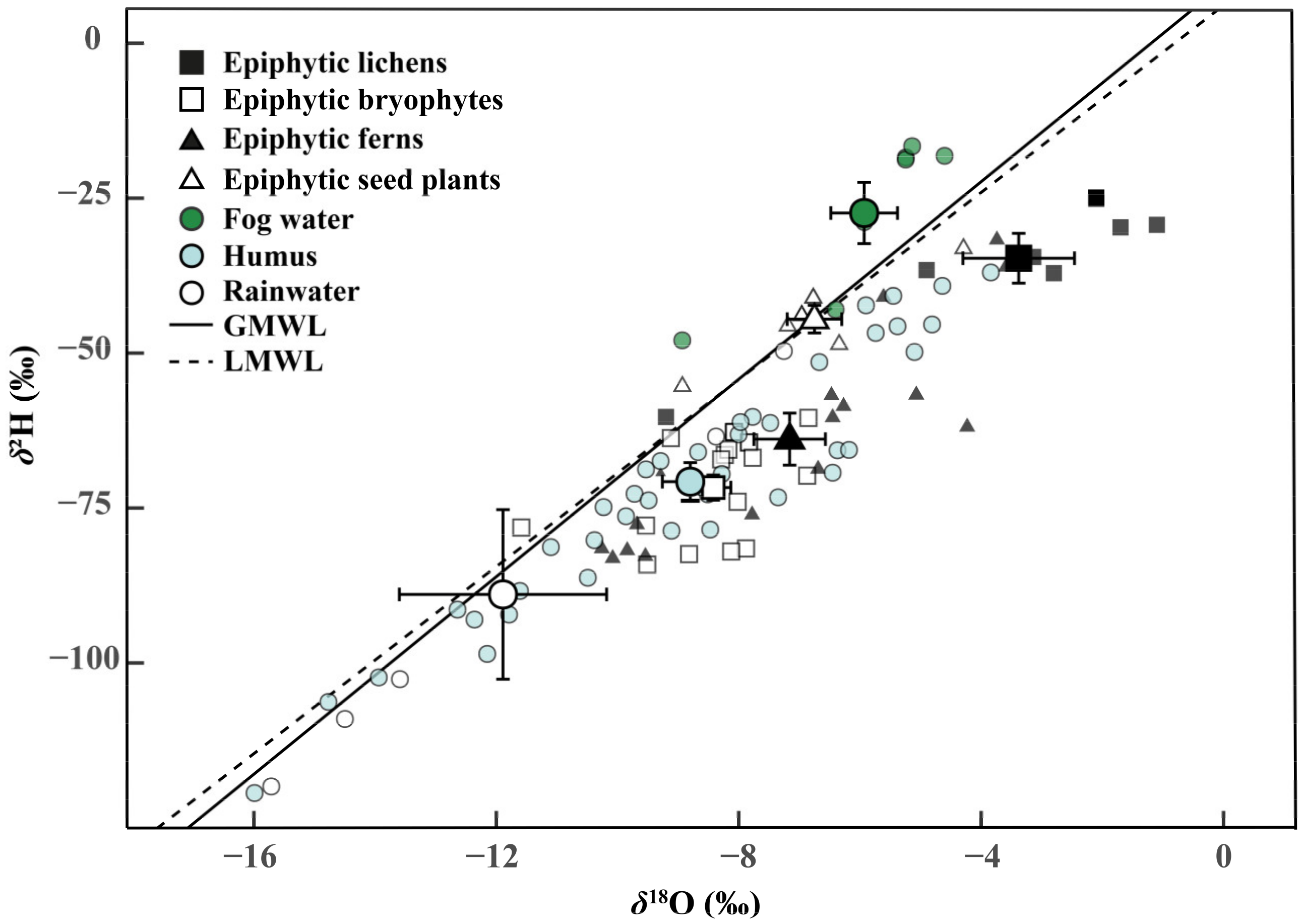
The isotope compositions (δ2H and δ18O) of epiphytes and their potential water sources were presented to appraise the isotopic fractionation processes during the peak dry season (Figure 2). As a reference, the local meteoric water line (LMWL: δ2H = 6.23 + 7.55 δ18O, R2 = 0.86, p < 0.001) was shown based on the rainfall data throughout 2018 and 2019. Compared with the global meter water line (GMWL: δ2H = 10 + 8 δ18O), the slope of LMWL was slightly lower than GMWL. The mean δ2H and δ18O of fog water were above the LMWL and GMWL, indicating that the fog water experienced lower evaporative enrichment than the canopy humus and rainwater. Among the four groups of epiphytes, the isotopic signatures of epiphytic lichens and epiphytic seed plants were close to the fog water. In addition, the δ2H and δ18O of epiphytic bryophytes and epiphytic ferns were identical to humus.
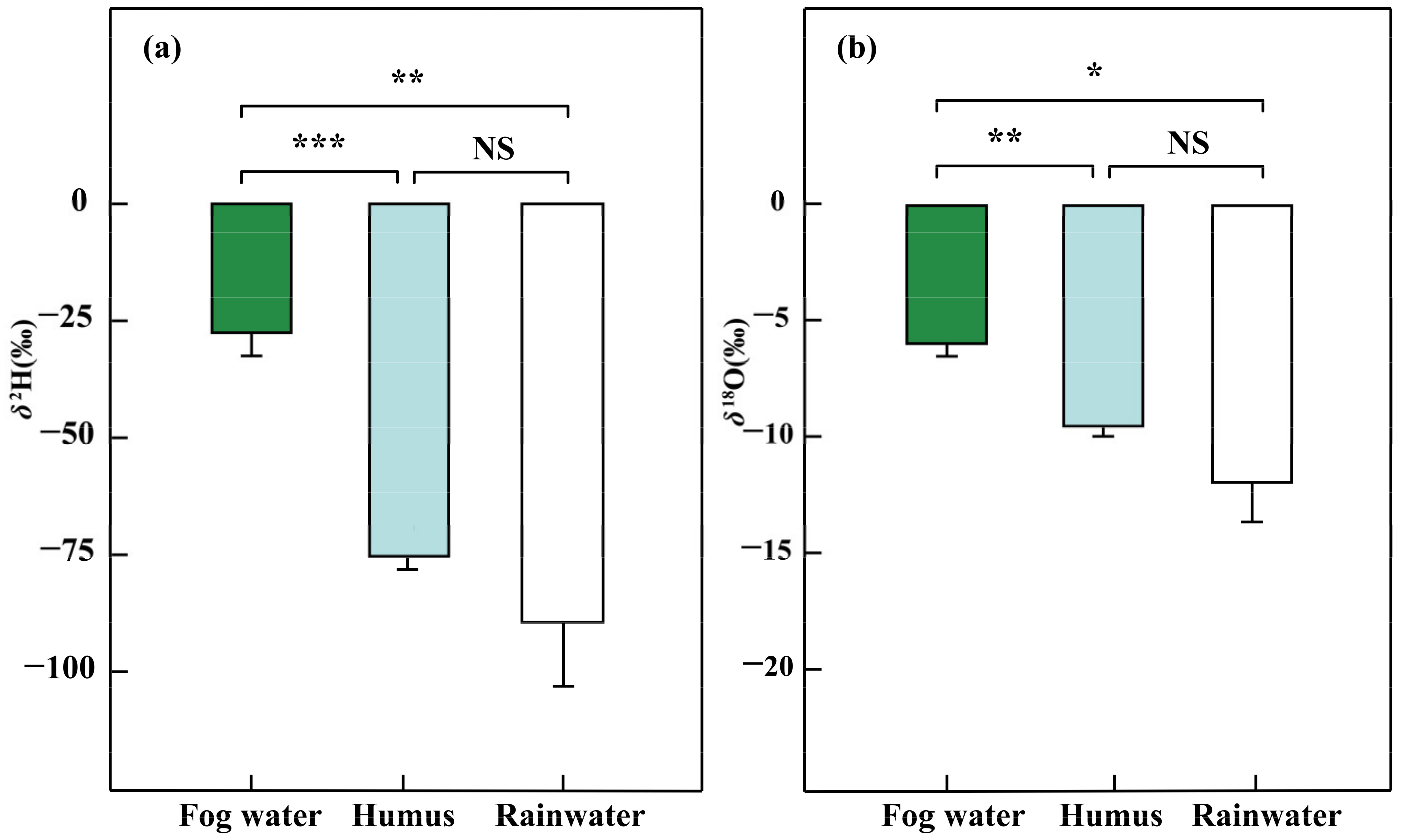
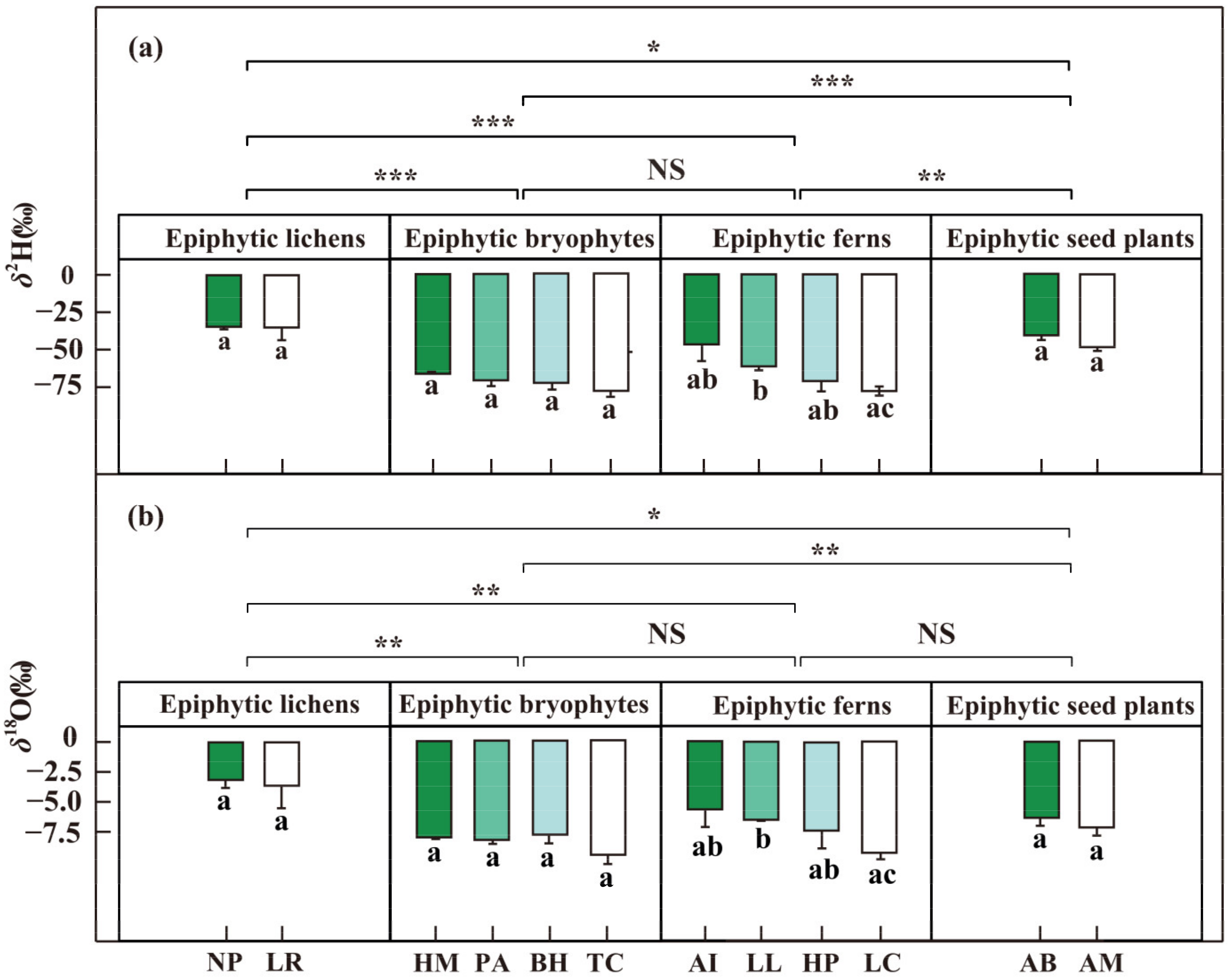
The δ2H and δ18O of fog water were higher (p < 0.05) than those of humus and rainwater (Figure 3 and Table S1). However, no significant difference was found between the humus and rainwater. Specifically, the average δ2H and δ18O values were −27.4 ± 4.9‰ and −5.93 ± 0.55‰ for fog water, −70.8 ± 3.1‰ and −8.80 ± 0.46‰ for humus, and −88.9 ± 13.7‰ and −11.89 ± 1.71‰ for rainwater. The δ2H and δ18O of epiphytic lichens were significantly higher than epiphytic bryophytes (p < 0.01), epiphytic ferns (p < 0.01), and epiphytic seed plants (p < 0.05) (Figure 4). Meanwhile, we also found a significant difference in δ2H and δ18O between epiphytic bryophytes and epiphytic seed plants (p < 0.01). There was no significant difference between the epiphytic bryophytes and the epiphytic ferns. The average δ2H and δ18O values were −34.7 ± 4.0‰ and −3.38 ± 0.92‰ for epiphytic lichens, −71.7 ± 2.0‰ and −8.42 ± 0.29‰ for epiphytic bryophytes, and −63.9 ± 4.2‰ and −7.16 ± 0.59‰ for epiphytic ferns, and −44.5 ± 2.2‰ and −6.75 ± 0.45‰ for epiphytic seed plants. There were also interspecific differences (p < 0.05) among the epiphytic ferns. The δ2H and δ18O values of epiphytic ferns ranged from −77.33‰ to −46.46‰ and from −9.22‰ to −5.66‰, respectively.
3.2. Partitioning of Water Sources for Epiphytes
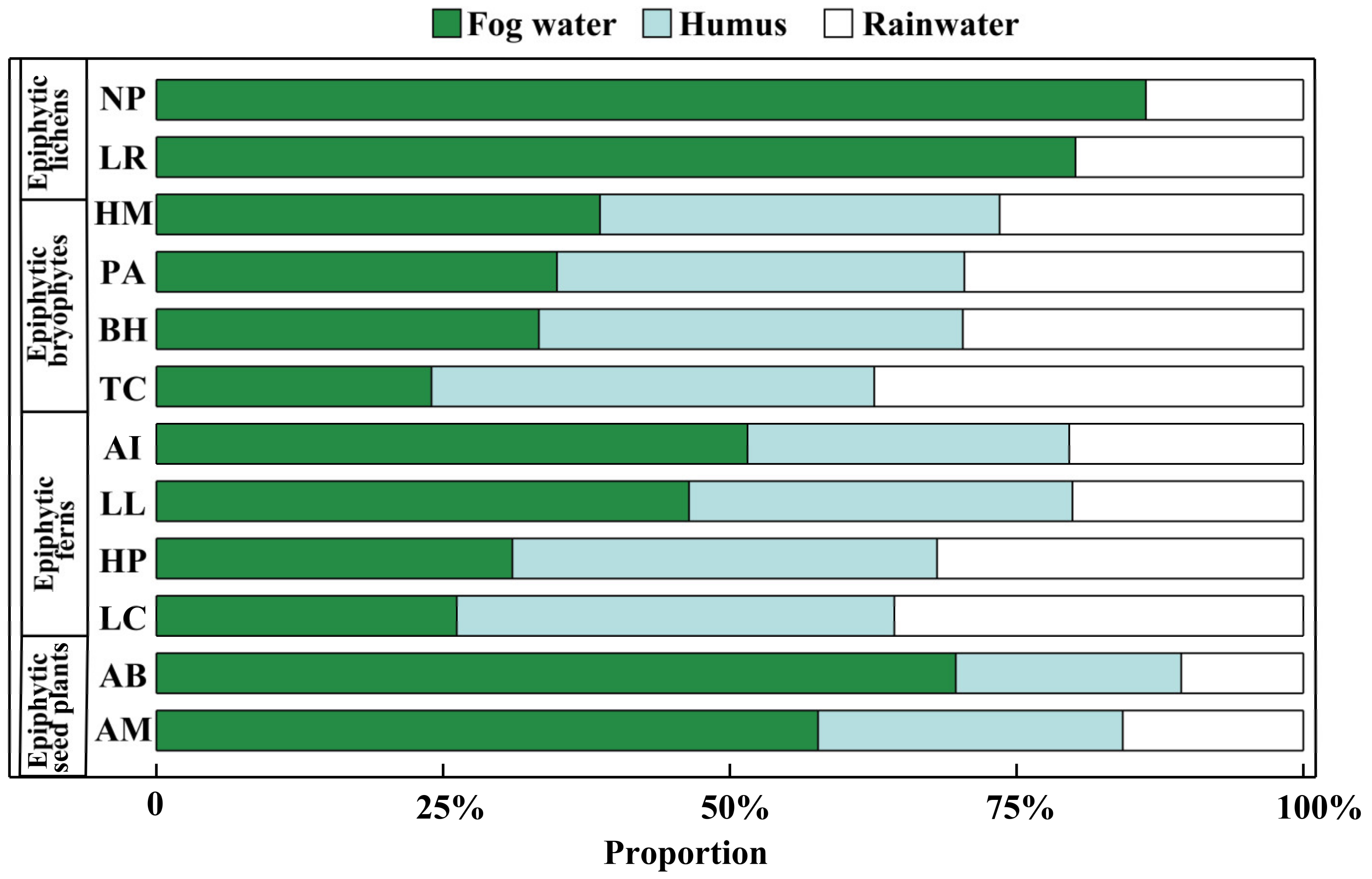
The MixSIAR model showed that all the epiphytes could use fog water as their water sources (Figure 5). Because the epiphytic lichens had only two potential water sources (see Section 2.4), the contributions of fog water to Nephromopsis pallescens (NP) and Lobaria retigera (LR) were up to 86.3 ± 9.9% and 80.1 ± 15.0%, respectively. With three potential water sources (i.e., fog water, humus, and rainwater), the epiphytic bryophytes (HM: Hamaliodendron montagneanum; PA: Plagiochila assamica; BH: Bazzania himlayana; TC: Thuidium cymbifolium) acquired 24.0 ± 11.7% to 38.7 ± 10.2% of their water from fog water. Meanwhile, the epiphytic bryophytes of PA, BH, and TC could absorb 35.6 ± 15.2% to 38.6 ± 16.3% of their water from humus. Similarly, 46.4 ± 12.6% to 51.5 ± 16.6% of the water utilized by the epiphytic ferns of Asplenium indicum (AI) and Lepisorus loriformis (LL) was from fog water. Besides this, the contribution of fog water toward Hymenophyllum polyanthos (HP) and Loxogramme chinensis (LC) accounted for 31.0 ± 13.2% and 26.2 ± 11.7%, respectively. However, the other two epiphytic ferns (HP: Hymenophyllum polyanthos; LC: Loxogramme chinensis) used humus as their major (37.0 ± 17.7% to 38.1 ± 15.9%) water source (p < 0.05). On the contrary, the epiphytic seed plants Aeschynanthus buxifolius (AB) and Agapetes mannii (AM) were highly dependent on fog water. The contribution percentage of fog water to AB and AM ranged from 57.7 ± 11.1% to 69.7 ± 12.0%.
3.3. Leaf Carbon Isotope Ratios
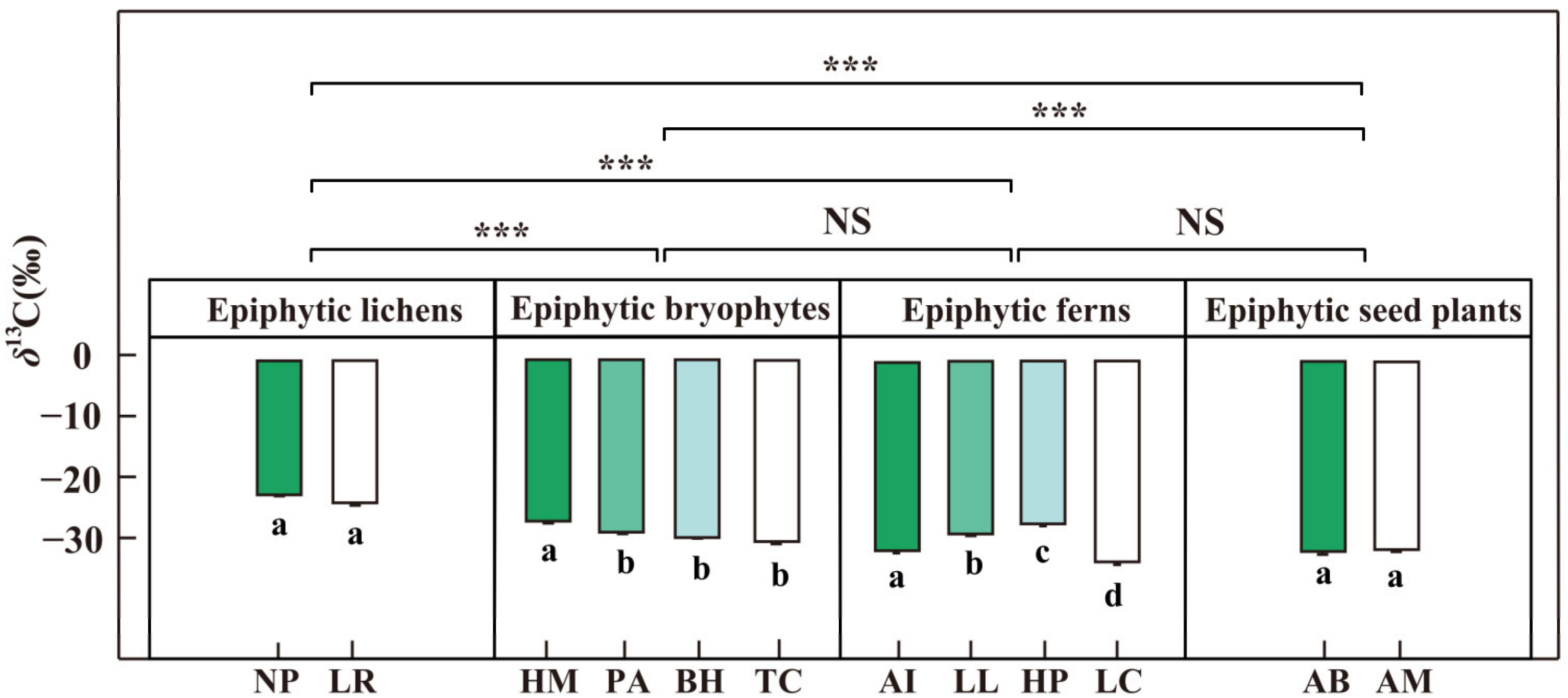
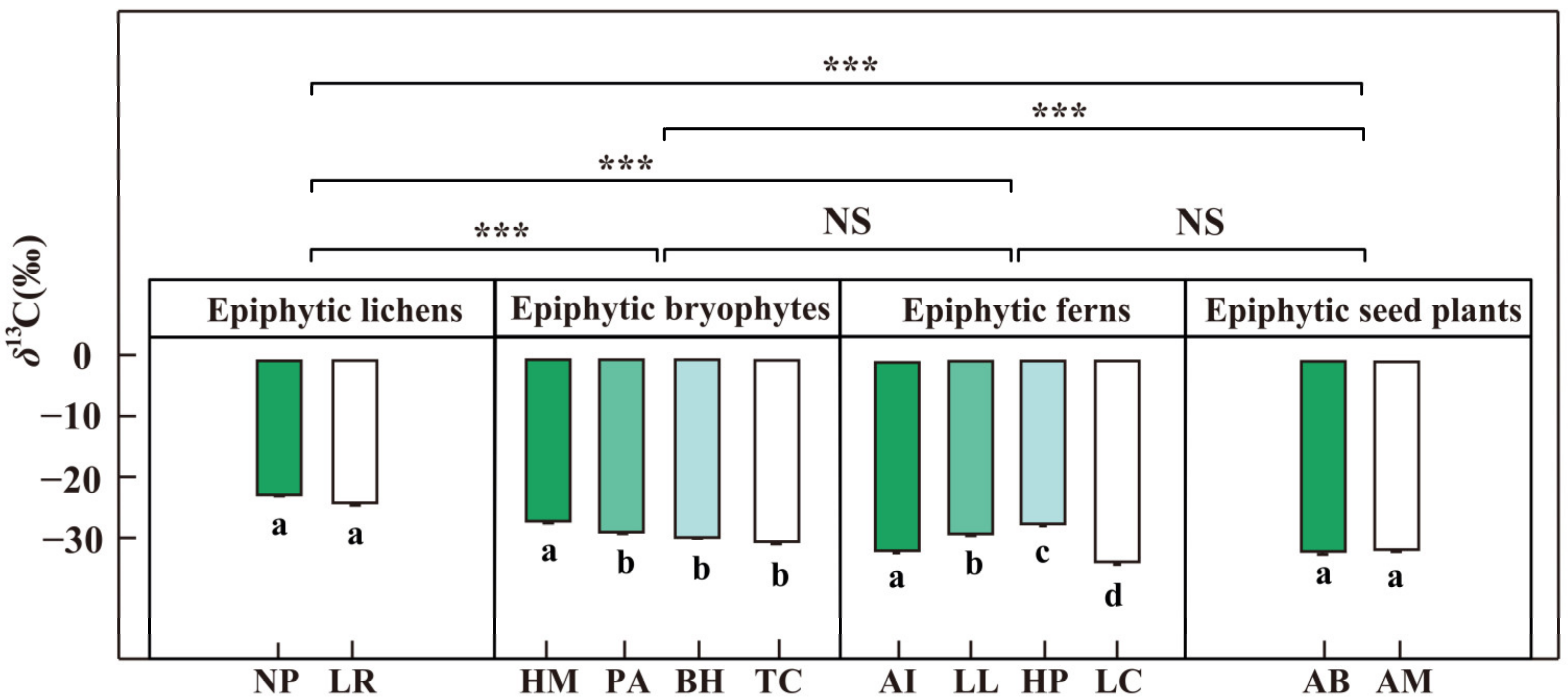
The δ13C of epiphytic lichens were significantly higher (p < 0.001) than the other epiphyte groups (Figure 6 and Table S1), but there was no difference among epiphytic bryophytes, epiphytic ferns, and epiphytic seed plants. The average δ13C values were −23.77 ± 0.34 ‰ for epiphytes lichens, −29.59 ± 0.36‰ for epiphytic bryophytes, −30.96 ± 0.66‰ for epiphytic ferns, and −32.29 ± 0.26 ‰ for epiphytic seed plants. Meanwhile, the δ13C values of epiphytic lichens were positively correlated with the δ2H (p < 0.05) and interactions of δ2H × δ18O in water sources (Table 1). The δ13C values of epiphytic bryophytes were different significantly (p < 0.05) among the four species; the average values of δ13C in Hamaliodendron montagneanum (HM: −27.57 ± 0.36‰) were significantly higher than other epiphytic bryophytes: −29.46 ± 0.29‰ for Plagiochila assamica (PA), −30.38 ± 0.23‰ for Bazzania himlayana (BH), and −30.96 ± 0.39‰ for Thuidium cymbifolium (TC). There were also interspecific differences (p < 0.05) among the four species of epiphytic ferns, which ranged from −27.81 ± 0.31‰ to −34.33 ± 0.40‰. These results indicated that the group-specific and interspecific differentiation of (i.e., δ13C) were obvious among epiphytes.
4. Discussion
Our study reveals the importance of fog in maintaining the water balance of epiphytes from a subtropical montane cloud forest in the dry season as well as the differentiation in water acquisition pattern of different epiphyte groups. The contribution of fog water to the water sources could be as high as 83.2% in epiphytic lichens. While epiphytic lichens and epiphytic seed plants mainly relied on fog water, the epiphytic bryophytes and epiphytic ferns were depending on both fog water and humus. We also found significant differences in among epiphytic groups and species. The clear differentiation of water utilization among epiphytes provides a possible explanation for their coexistence. Our results also provide important implications for their responses to projected climate change patterns including a decline in fog persistence in the region.
4.1. Water Conditions in the Subtropical MCF
In our study area, we found that the slope of LMWL (δ2H = 6.23 + 7.55 δ18O, R2 = 0.86, p < 0.001) was slightly lower than that in GMWL (δ2H = 10 + 8 δ18O). This result indicated that, in the dry season, there is a certain degree of fractionation during the rainfall process, which could be seen in other montane forests [61]. Moreover, the δ2H and δ18O of fog water were located above the LMWL, suggesting that fog water was more enriched compared with canopy humus and rainwater. Earlier studies also indicate that the isotopic signature of fog tends to be more enriched in heavier isotopes δ2H and δ18O than rainwater due to different condensation temperatures and histories [62]. In contrast, the water isotopes of canopy humus represent a mixture of evaporation and atmospheric water inputs (i.e., fog water and rainwater), which leads to the isotope signatures of δ2H and δ18O in humus falling in between fog water and rainwater [46].
4.2. Water Partitioning of the Four Epiphyte Groups
Generally, epiphytes could obtain fog water through root uptake of coalesced water that drips into the soil or by direct foliar water uptake (FWU) [63]. However, there are large differences among different groups and species in the ability of FWU. Although both epiphytic lichens and epiphytic bryophytes were probably some of the earliest land plants, they showed distinct structural differences. These differences can most likely be attributed to the evolution in plant morphology and structure. Epiphytic lichens are a symbiosis composed of fungus and algae [30]. Epiphytic lichens found in our study region can only attach to tree trunks and mainly absorb fog water (80.1% to 86.3%) during the dry season. However, it should be noted that in some other regions (e.g., the pacific NW of the USA), lichens thrive in moist habitats with a bit of humus development [64], so the water sources of lichens from different forest ecosystems need to be analyzed further. Epiphytic bryophytes are true multicellular organisms with rhizomes [31,32]. They could absorb water from fog water (24.0% to 38.7%), rainwater (26.5% to 37.4%) and humus (35.6% to 38.6%). Our study also found that epiphytic seed plants absorb more fog water than epiphytic ferns. The epiphytic seed plants Aeschynanthus buxifolius (AB) and Agapetes mannii (AM) make full use of fog water (ranged from 57.7 ± 11.1% to 69.7 ± 12.0%) during the dry season (Figure 5). This is most probably because epiphytic seed plants have complete roots, stomata, and the vascular system, which could absorb water sources more flexibly than epiphytic ferns [33,65]. There was also an obvious interspecific differentiation in water acquisition among the epiphyte species (Figure 4). Wu et al. [27] pointed out that there was a divergence of water sources among four epiphytic orchids (B. delitescens, C. sumatranus, C. viscosa, and P. weinmanniifolia). Some studies further found that the leaf morphology and anatomy (e.g., leaf thickness and stomata) of epiphytes may account for a significant part of the variation in water uptake at the interspecific level [66,67].
It is obvious that the epiphytes in this study are C3 species based on the carbon isotope data. The average δ13C values of all epiphytes were below −20‰ (Figure 6), which is commonly interpreted as evidence for C3-photosynthesis [17]. Meanwhile, we found obvious group-specific and interspecific differentiation in (i.e., δ13C) (Figure 6 and Table 1), which could be related to their different water acquisition patterns. Through the LMM model, we further found that the δ13C of epiphytic lichens were correlated with their δ2H / δ18O (Table 1). Thus, the difference in water acquisition patterns of epiphytes probably has an effect on . This relationship has also been found in other studies [36,48]. Moreno-Gutierrez et al., [48] indicated that the large variation in of coexisting species was affected by the stability of water sources. Because the habitats lack humus accumulation, epiphytic lichens in our study area can only absorb the unstable atmospheric water inputs (i.e., fog water and rainwater). Therefore, they exhibited higher compared to other groups to cope with the relatively unstable water input during the dry season (Figure 5 and Figure 6). The apparent differentiation of water utilization and may facilitate the coexistence of different epiphytes in the canopy with the intermittent water supply.
The present study confirmed that all epiphytes can use fog water in various degrees (Figure 5). For the epiphytic lichens, we found that fog water accounted for 80.1~86.3% of their water acquisitions. The average contribution proportions of fog water to epiphytic bryophytes, epiphytic ferns, and epiphytic seed plants were 32.7%, 38.8%, and 63.7%, respectively. The dependence on fog water of epiphytes was possible because of the frequent fog events in the MCF [8,68]. Gehrig-Downie et al. [8] found that the frequent input of fog water enhances the growth of four epiphyte groups (bryophytes, lichens, pteridophytes, angiosperms) growth by shortening the desiccation periods. Several early studies in cloud montane forests also found that the canopy drip of throughfall could exceed the rainfall by 1.2~3.2 times, which confirmed the importance of fog as an extra input of water [8,51]. For epiphytes, fog water is more important because there are no vascular connections to their host plants (or ground) [24,27]. Zheng and Feng [69] pointed out that the fog water absorptions of the epiphytic orchids C. occultata and S. dawsonianus were higher than that in other non-epiphytes. In addition, some studies also suggested that fog water moving horizontally can be intercepted and accumulated on the epiphytic surfaces [24,70]. For instance, Song et al. [9] showed that fan bryophytes (H. flabellatum, P. arbuscula, and P. assamica) could intercept and absorb the fog water because of the flattened surfaces.
Notably, here in the present study, the δ2H and δ18O fog and rainfall water collected in open areas were used to represent the values of fog and rainwater used by epiphytes. This may raise some methodological concerns, such as the difference in physical location of the trees and rain/fog collectors, and the sampling timing [71,72]. However, there is no or neglectable evaporation and isotope discrimination when the fog moves from an open area to the forest and from the top canopy to the site of the epiphytes because the relative humidity (RH) is equal to or close to 100% when there is fog [50]. There might be isotope fractionation when the rainwater is intercepted by canopy leaves, evaporates for a while after the rain, and then drops to epiphytes [72,73]. However, the contribution of this leaf-intercepted water should be very limited as the remaining water on the leaf surface after the rainfall event will most likely be lost through evaporation or be directly absorbed by the tree leaves (FWU) [74]. In addition, the δ2H and δ18O of humus are changing overtime after a rain or fog event due to evaporation. Nevertheless, our study suggested that the effect of evaporation on canopy humus is limited because the isotope signatures of humus are close to (p > 0.05) that of rainwater (Figure 3). Actually, the isotopic composition (δ2H and δ18O) of canopy humus varies slightly within the forest. In order to reduce the spatial uncertainty, the humus samples we collected were one-to-one correspondence to the epiphyte samples during the experiment period. Therefore, we collected humus while we were collecting epiphyte samples, which should be the best and most practical approach because the isotope signatures of humus collected should be close to that of the water absorbed by epiphytes from humus during collection. These potential technical uncertainties apply to all water source determination studies using δ2H and δ18O [56,75], and can be further tested with a study quantifying the spatial-temporal variation of δ2H and δ18O in a high sampling resolution [76].
4.3. Ecological Implications
High fog dependence of epiphytes indicates that predicted declines in fog persistence due to climate change may negatively impact their performance and survival. Fog water has been suggested to be an important water input for epiphytes in forest ecosystems [51,63]. The present study further confirmed this and indicated a significant differentiation of fog water utilization among epiphytes. With the intensification of global warming [2], rising cloud bases will intensify the solar radiation received by MCF, which may further decrease the fog frequency and duration [38]. The reduction of fog events and/or duration will probably result in a high mortality of fog-dependent species (especially epiphytes) under projected climate change scenarios [51]. Therefore, the epiphyte community in MCF may experience a change in composition and functioning. Previous results also suggested that decreases in fog persistence will negatively affect the productivity and longevity of epiphytes [62,77]. For example, Nadkarni and Solano [77] found that the decline of fog persistence significantly increased leaf mortality, reduced leaf production, and shortened the longevity of epiphytes. Because epiphytes play crucial roles in hydrological and nutrient cycles, and promote the survival of other plants and animals in the canopy habitat [7,12], the potential deterioration of epiphytes due to decreasing fog persistence may have cascading effects on the whole forest ecosystem. Therefore, more conservation attention should be paid to epiphytes in montane forest ecosystems under climate change.
5. Conclusions
Our results indicated that epiphytic lichens and epiphytic seed plants depended mainly on fog water whereas the epiphytic bryophytes and epiphytic ferns relied on both fog water and humus. Also, there were obvious variations in among groups and species, and the of epiphytic lichens were appreciably higher than the other epiphyte groups. The variation in was probably related to the differentiation of water resources and water use strategies. Despite the fact that water sources of epiphytes were significantly different among groups and species, this study clearly showed that all epiphytes rely on fog water heavily in the dry season. Thus, our results suggest a dim future for epiphytes as fog persistence is predicted to decline. Conservation efforts may be needed to retain the high diversity of epiphytes in this region. The response of these epiphytes to increasingly severe drought also needs to be studied to better understand their drought response mechanisms and to develop mitigation strategies.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/w13223237/s1, Figure S1: Photograph of self-made V-shaped fog collector (0.3 m × 1 m) (a) and cylindrical rain collector (0.2 m × 0.65 m) (b) at the study site, Table S1: The δ2H, δ18O and δ13C (‰) (mean ± SE) of water sources and epiphytes from different groups in a subtropical montane cloud forest in the dry season. The abbreviations of species names are the same as in Figure 4.
Author Contributions
Writting—original draft preparation, L.-L.L. and B.Y.; writing—review and editing, L.-L.L., L.S., H.-Z.L., Y.W., X.-J.M. and Y.-J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32171529, 32171557, 31670452), the Yunnan Natural Science Foundation (202101AT070059, 202001AS070044), the Chinese Academy of Sciences “Light of West China” program, the candidates of the Young and Middle Aged Academic Leaders of Yunnan Province (2019HB040), and the Yunnan High Level Talents Special Support Plan (YNWR-QNBJ-2020-066), and the Lancang-Mekong Cooperation (LMC) Special Fund (Biodiversity Monitoring and Network Construction along Lancang-Mekong River Basin Project).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the Ailaoshan Station for Subtropical Forest Ecosystem Studies for granting permission and facilitating this research and the Central Laboratory of Xishuangbanna Tropical Botanical Garden for the assistance in analyzing the isotope abundance. We are also grateful to Chun-Mei Li, Yan-Qin Dao, Cheng-Chang Luo, Jin-Hua Qi, Yue-Yan Zhang, Yue Luo, Zheng-Tian Wu, and Fan-Shuo Zhang for their assistance during the field surveying and sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Ault, T.R. On the essentials of drought in a changing climate. Science 2020, 368, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Corlett, R.T. The Impacts of Droughts in Tropical Forests. Trends Plant Sci. 2016, 21, 584–593. [Google Scholar] [CrossRef]
- Wanek, W.; Pörtl, K. Phyllosphere nitrogen relations: Reciprocal transfer of nitrogen between epiphyllous liverworts and host plants in the understorey of a lowland tropical wet forest in Costa Rica. New Phytol. 2005, 166, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.F.; Chen, H. Mechanisms Regulating Epiphytic Plant Diversity. Crit. Rev. Plant Sci. 2012, 31, 391–400. [Google Scholar] [CrossRef]
- Hsu, R.C.-C.; Tamis, W.L.M.; Raes, N.; De Snoo, G.R.; Wolf, J.H.D.; Oostermeijer, G.; Lin, S.-H. Simulating climate change impacts on forests and associated vascular epiphytes in a subtropical island of East Asia. Divers. Distrib. 2012, 18, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Benzing, D.H. Vulnerabilities of tropical forests to climate change: The significance of resident epiphytes. In Potential Impacts of Climate Change on Tropical Forest Ecosystems; Springer: Dordrecht, The Netherlands, 1998; pp. 379–400. [Google Scholar]
- Gehrig-Downie, C.; Obregón, A.; Bendix, J.; Gradstein, S.R. Epiphyte Biomass and Canopy Microclimate in the Tropical Lowland Cloud Forest of French Guiana. Biotropica 2011, 43, 591–596. [Google Scholar] [CrossRef]
- Song, L.; Zhang, Y.-J.; Chen, X.; Li, S.; Lu, H.-Z.; Wu, C.-S.; Tan, Z.-H.; Liu, W.-Y.; Shi, X.-M. Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest. J. Plant Res. 2015, 128, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, N.M. Epiphyte Biomass and Nutrient Capital of a Neotropical Elfin Forest. Biotropica 1984, 16, 249–256. [Google Scholar] [CrossRef]
- Cardelús, C.L.; Mack, M.C. The nutrient status of epiphytes and their host trees along an elevational gradient in Costa Rica. Plant Ecol. 2009, 207, 25–37. [Google Scholar] [CrossRef]
- Mendieta-Leiva, G.; Porada, P.; Bader, M.Y. Interactions of epiphytes with precipitation partitioning. In Precipitation Partitioning by Vegetation; Springer International Publishing: New York, NY, USA, 2020; pp. 133–146. [Google Scholar]
- Liu, W.-J.; Wang, P.-Y.; Li, J.-T.; Liu, W.-Y.; Li, H.-M. Plasticity of source-water acquisition in epiphytic, transitional and terrestrial growth phases of Ficus tinctoria. Ecohydrology 2014, 7, 1524–1533. [Google Scholar] [CrossRef]
- Yoder, J.A.; Zettler, L.W.; Stewart, S.L. Water requirements of terrestrial and epiphytic orchid seeds and seedlings, and evidence for water uptake by means of mycotrophy. Plant Sci. 2000, 156, 145–150. [Google Scholar] [CrossRef]
- Xu, H.-Q.; Liu, W.-Y. Species diversity and distribution of epiphytes in the montane moist evergreen broad-leaved forest in Ailao Mountain, Yunnan. Biodivers. Sci. 2005, 13, 137–147. [Google Scholar] [CrossRef]
- Ma, W.-Z. The Composition and Biomass of Epiphytic Materials and Their Relationships with Ecological Factors in Xujiaba Region from Ailao Mountain, Yunnan. Ph.D. Thesis, Graduate School of the Chinese Academy of Sciences, Beijing, China, 2009. [Google Scholar]
- Zotz, G.; Hietz, P. The physiological ecology of vascular epiphytes: Current knowledge, open questions. J. Exp. Bot. 2001, 52, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.E., Jr.; Mack, M.C.; Sinclair, T.R.; Mulkey, S.S. Ecological and evolutionary consequences of desiccation tolerance in tropical fern gametophytes. New Phytol. 2007, 176, 708–717. [Google Scholar] [CrossRef]
- Benzing, D.H. Air Plants: Epiphytes and Aerial Gardens; Cornell University Press: Ithaca, NY, USA, 2012. [Google Scholar]
- Reyes-García, C.; Mejia-Chang, M.; Griffiths, H. High but not dry: Diverse epiphytic bromeliad adaptations to exposure within a seasonally dry tropical forest community. New Phytol. 2011, 193, 745–754. [Google Scholar] [CrossRef]
- Querejeta, J.I.; Prieto, I.; Torres, P.; Campoy, M.; Alguacil, M.M.; Roldán, A. Water-spender strategy is linked to higher leaf nutrient concentrations across plant species colonizing a dry and nutrient-poor epiphytic habitat. Environ. Exp. Bot. 2018, 153, 302–310. [Google Scholar] [CrossRef]
- Zotz, G. The systematic distribution of vascular epiphytes—A critical update. Bot. J. Linn. Soc. 2013, 171, 453–481. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Long, W.; Zhang, H.; Huang, J.; Cheng, Y.; Jiang, H.; Liao, L.; Tan, Z. Divergent Strategies of Epiphytic Pteridophytes and Angiosperms Responding to Dry and Wet Seasons in a Tropical Cloud Forest. Trop. Conserv. Sci. 2020, 13. [Google Scholar] [CrossRef]
- Villegas, J.C.; Tobón, C.; Breshears, D.D. Fog interception by non-vascular epiphytes in tropical montane cloud forests: De-pendencies on gauge type and meteorological conditions. Hydrol. Process. 2008, 22, 2484–2492. [Google Scholar] [CrossRef]
- Darby, A.; Draguljić, D.; Glunk, A.; Gotsch, S.G. Habitat moisture is an important driver of patterns of sap flow and water balance in tropical montane cloud forest epiphytes. Oecologia 2016, 182, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Freiberg, M.; Turton, S.M. Importance of drought on the distribution of the birds nest fern, Asplenium nidus, in the canopy of a lowland tropical rainforest in north-eastern Australia. Austral Ecol. 2007, 32, 70–76. [Google Scholar] [CrossRef]
- Wu, Y.; Song, L.; Liu, W.; Liu, W.; Li, S.; Fu, P.; Shen, Y.; Wu, J.; Wang, P.; Chen, Q.; et al. Fog Water Is Important in Maintaining the Water Budgets of Vascular Epiphytes in an Asian Tropical Karst Forests during the Dry Season. Forests 2018, 9, 260. [Google Scholar] [CrossRef] [Green Version]
- Watkins, J.E.; Rundel, P.W.; Cardelús, C.L. The influence of life form on carbon and nitrogen relationships in tropical rainforest ferns. Oecologia 2007, 153, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Beckett, R.; Hochman, A.; Nash, T.H. Desiccation-Tolerance in Lichens: A Review. Bryologist 2008, 111, 576–593. [Google Scholar] [CrossRef]
- Green, T.G.A.; Sancho, L.G.; Pintado, A. Ecophysiology of desiccation/rehydration cycles in mosses and lichens. In Plant Desiccation Tolerance; Springer: Berlin/Heidelberg, Germany, 2011; pp. 89–120. [Google Scholar]
- Ah-Peng, C.; Cardoso, A.W.; Flores, O.; West, A.; Wilding, N.; Strasberg, D.; Hedderson, T.A. The role of epiphytic bryophytes in interception, storage, and the regulated release of atmospheric moisture in a tropical montane cloud forest. J. Hydrol. 2017, 548, 665–673. [Google Scholar] [CrossRef]
- Horwath, A.B.; Royles, J.; Tito, R.; Gudiño, J.A.; Allen, N.S.; Farfan-Rios, W.; Rapp, J.M.; Silman, M.R.; Malhi, Y.; Swamy, V.; et al. Bryophyte stable isotope composition, diversity and biomass define tropical montane cloud forest extent. Proc. R. Soc. B Boil. Sci. 2019, 286, 20182284. [Google Scholar] [CrossRef] [Green Version]
- Proctor, M.C.F. The diversification of bryophytes and vascular plants in evolving terrestrial environments. In Photosynthesis in Bryophytes and Early Land Plants; Hanson, D.T., Rice, S.K., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 59–77. [Google Scholar]
- Tsutsumi, C.; Kato, M. Evolution of epiphytes in Davalliaceae and related ferns. Bot. J. Linn. Soc. 2006, 151, 495–510. [Google Scholar] [CrossRef] [Green Version]
- Zotz, G. Plants on Plants: The Biology of Vascular Epiphytes; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Yuan, G.-D.; Liu, W.-Y.; Shi, X.-M.; Fan, X.-Y. Effects of water content change on photosynthetic and fluorescence parameters of four bole epiphytic bryophytes in montane moist evergreen broad-leaved forest in the Ailao Mountains. Plant Sci. J. 2018, 36, 603–611. [Google Scholar]
- Zhao, C.; Deng, X.; Yuan, Y.; Yan, H.; Liang, H. Prediction of Drought Risk Based on the WRF Model in Yunnan Province of China. Adv. Meteorol. 2013, 2013, 295856. [Google Scholar] [CrossRef]
- Williams, C.B.; Murray, J.G.; Glunk, A.; Dawson, T.E.; Nadkarni, N.M.; Gotsch, S.G. Vascular epiphytes show low physiological resistance and high recovery capacity to episodic, short-term drought in Monteverde, Costa Rica. Funct. Ecol. 2020, 34, 1537–1550. [Google Scholar] [CrossRef]
- Song, L.; Lu, H.-Z.; Xu, X.-L.; Li, S.; Shi, X.-M.; Chen, X.; Wu, Y.; Huang, J.-B.; Chen, Q.; Liu, S.; et al. Organic nitrogen uptake is a significant contributor to nitrogen economy of subtropical epiphytic bryophytes. Sci. Rep. 2016, 6, 30408. [Google Scholar] [CrossRef] [Green Version]
- Goswami, P.; Sarkar, S. Analysis and quantification of contrasts in observed meteorological fields for foggy and non-foggy days. Meteorol. Atmos. Phys. 2015, 127, 605–623. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Cao, K.-F.; Goldstein, G. Winter photosynthesis of evergreen broadleaf trees from a montane cloud forest in Subtropical China. In Advanced Topics in Science and Technology in China; Springer: Berlin/Heidelberg, Germany, 2013; pp. 812–817. [Google Scholar]
- Li, S.; Liu, W.-Y.; Li, D.-W. Epiphytic lichens in subtropical forest ecosystems in Southwest China: Species diversity and implications for conservation. Biol. Conserv. 2013, 159, 88–95. [Google Scholar] [CrossRef]
- Lu, H.; Brooker, R.; Song, L.; Liu, W.; Sack, L.; Zhang, J.; Yu, F. When facilitation meets clonal integration in forest canopies. New Phytol. 2020, 225, 135–142. [Google Scholar] [CrossRef]
- Tang, D.-D.; Wu, Y.; Liu, W.-Y.; Chen, Q.; Li, D.-F.; Zhang, T.-T. Diversity and floristic characteristics of vascular epiphytes in montane forests in the Ailao Mountains, Yunnan. Plant Sci. J. 2018, 36, 658–666. [Google Scholar]
- Ehleringer, J.R.; Dawson, T.E. Water uptake by plants: Perspectives from stable isotope composition. Plant Cell Environ. 1992, 15, 1073–1082. [Google Scholar] [CrossRef]
- Dawson, T.E.; Simonin, K.A. The roles of stable isotopes in forest hydrology and biogeochemistry. In Forest Hydrology and Biogeochemistry; Springer: Dordrecht, The Netherlands, 2011; pp. 137–161. [Google Scholar]
- Hietz, P.; Wanek, W.; Popp, M. Stable isotopic composition of carbon and nitrogen and nitrogen content in vascular epiphytes along an altitudinal transect. Plant Cell Environ. 1999, 22, 1435–1443. [Google Scholar] [CrossRef]
- Moreno-Gutierrez, C.; Dawson, T.E.; Nicolas, E.; Querejeta, J.I. Isotopes reveal contrasting water use strategies among coex-isting plant species in a Mediterranean ecosystem. New Phytol. 2012, 196, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.S.O.; Dawson, T.E. The contribution of fog to the water relations of Sequoia sempervirens (D. Don): Foliar uptake and prevention of dehydration. Plant Cell Environ. 2004, 27, 1023–1034. [Google Scholar] [CrossRef]
- Emery, N.C. Foliar uptake of fog in coastal California shrub species. Oecologia 2016, 182, 731–742. [Google Scholar] [CrossRef]
- Eller, C.B.; Lima, A.L.; Oliveira, R.S. Foliar uptake of fog water and transport belowground alleviates drought effects in the cloud forest tree species, Drimys brasiliensis (Winteraceae). New Phytol. 2013, 199, 151–162. [Google Scholar] [CrossRef]
- Hietz, P.; Wanek, W. Size-Dependent Variation of Carbon and Nitrogen Isotope Abundances in Epiphytic Bromeliads. Plant Biol. 2003, 5, 137–142. [Google Scholar] [CrossRef]
- Yang, B.; Meng, X.; Zhu, X.; Zakari, S.; Singh, A.K.; Bibi, F.; Mei, N.; Song, L.; Liu, W. Coffee performs better than amomum as a candidate in the rubber agroforestry system: Insights from water relations. Agric. Water Manag. 2021, 244, 106593. [Google Scholar] [CrossRef]
- Stock, B.C.; Semmens, B.X. MixSIAR: Bayesian Mixing Models in R 2020. [R Package Version 3.1.12]. Available online: https://github.com/brianstock/MixSIAR (accessed on 12 November 2021).
- Wang, J.; Lu, N.; Fu, B. Inter-comparison of stable isotope mixing models for determining plant water source partitioning. Sci. Total Environ. 2019, 666, 685–693. [Google Scholar] [CrossRef]
- Wu, H.; Li, X.-Y.; Li, J.; Zhang, C.; He, B.; Zhang, S.; Sun, W. Age-related water uptake patterns of alpine plantation shrubs in reforestation region of Qinghai-Tibetan Plateau based on stable isotopes. Ecohydrology 2019, 12, e2049. [Google Scholar] [CrossRef] [Green Version]
- Phillips, D.L.; Newsome, S.D.; Gregg, J.W. Combining sources in stable isotope mixing models: Alternative methods. Oecologia 2005, 144, 520–527. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; p. 2. [Google Scholar]
- Li, S.; Liu, S.; Shi, X.-M.; Liu, W.-Y.; Song, L.; Lu, H.-Z.; Chen, X.; Wu, C.-S. Forest Type and Tree Characteristics Determine the Vertical Distribution of Epiphytic Lichen Biomass in Subtropical Forests. Forests 2017, 8, 436. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4: Linear Mixed-Effects Models Using Eigen and S4. [R Package Version 1.1–27.1.]. 2021. Available online: https://cran.r-project.org/web/packages/lme4 (accessed on 12 November 2021).
- Brooks, J.R.; Barnard, H.R.; Coulombe, R.; Mcdonnell, J. Ecohydrologic separation of water between trees and streams in a Mediterranean climate. Nat. Geosci. 2009, 3, 100–104. [Google Scholar] [CrossRef]
- Scholl, M.; Eugster, W.; Burkard, R. Understanding the role of fog in forest hydrology: Stable isotopes as tools for determining input and partitioning of cloud water in montane forests. Hydrol. Process. 2011, 25, 353–366. [Google Scholar] [CrossRef]
- Limm, E.B.; Dawson, T.E. Polystichum munitum (Dryopteridaceae) varies geographically in its capacity to absorb fog water by foliar uptake within the redwood forest ecosystem. Am. J. Bot. 2010, 97, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Rossman, A.; Rosentreter, R.; Ponzetti, J. Lichen ecology and diversity of a sagebrush steppe in Oregon: 1977 to the present. North Am. Fungi 2011, 6, 1–14. [Google Scholar]
- Krömer, T.; Gradstein, S. Vascular epiphytes. In Core Standardized Methods for Rapid Biological Field Assessment; Conservation International: Arlington, VA, USA, 2016; pp. 25–36. [Google Scholar]
- Gauslaa, Y.; Coxson, D. Interspecific and intraspecific variations in water storage in epiphytic old forest foliose lichens. Botany 2011, 89, 787–798. [Google Scholar] [CrossRef]
- Mediavilla, S.; Escudero, A.; Heilmeier, H. Internal leaf anatomy and photosynthetic resource-use efficiency: Interspecific and intraspecific comparisons. Tree Physiol. 2001, 21, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weathers, K.C.; Ponette-González, A.G.; Dawson, T.E. Medium, vector, and connector: Fog and the maintenance of ecosystems. Ecosystems 2020, 23, 217–229. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Feng, Y. Fog water absorption by the leaves of epiphytes and non-epiphytes in Xishuangbanna. J. Appl. Ecol. 2006, 17, 977–981. [Google Scholar]
- Goldsmith, G.R.; Matzke, N.; Dawson, T.E. The incidence and implications of clouds for cloud forest plant water relations. Ecol. Lett. 2013, 16, 307–314. [Google Scholar] [CrossRef]
- Yang, B.; Wen, X.; Sun, X. Seasonal variations in depth of water uptake for a subtropical coniferous plantation subjected to drought in an East Asian monsoon region. Agric. For. Meteorol. 2015, 201, 218–228. [Google Scholar] [CrossRef]
- Fischer, B.M.; van Meerveld, H.J.; Seibert, J. Spatial variability in the isotopic composition of rainfall in a small headwater catchment and its effect on hydrograph separation. J. Hydrol. 2017, 547, 755–769. [Google Scholar] [CrossRef] [Green Version]
- Bedaso, Z.K.; DeLuca, N.M.; Levin, N.; Zaitchik, B.; Waugh, D.W.; Wu, S.-Y.; Harman, C.J.; Shanko, D. Spatial and temporal variation in the isotopic composition of Ethiopian precipitation. J. Hydrol. 2020, 585, 124364. [Google Scholar] [CrossRef]
- Limm, E.B.; Simonin, K.A.; Bothman, A.G.; Dawson, T.E. Foliar water uptake: A common water acquisition strategy for plants of the redwood forest. Oecologia 2009, 161, 449–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querejeta, J.I.; Estrada-Medina, H.; Allen, M.F.; Jiménez-Osornio, J.J. Water source partitioning among trees growing on shallow karst soils in a seasonally dry tropical climate. Oecologia 2007, 152, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wang, P.; You, D.; Liu, W. Coupling evapotranspiration partitioning with root water uptake to identify the water consumption characteristics of winter wheat: A case study in the North China Plain. Agric. For. Meteorol. 2018, 259, 296–304. [Google Scholar] [CrossRef]
- Nadkarni, N.M.; Solano, R. Potential effects of climate change on canopy communities in a tropical cloud forest: An experimental approach. Oecologia 2002, 131, 580–586. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Meteorological conditions of the study site. Average monthly foggy days and precipitation (during 2015–2019; (a)), as well as average monthly number of evaporation and net radiation (b) at the study site. The dry seasons from November to April are remarked by grey bars. Each vertical bar represents an average (±SE) for that month.
Figure 1.
Meteorological conditions of the study site. Average monthly foggy days and precipitation (during 2015–2019; (a)), as well as average monthly number of evaporation and net radiation (b) at the study site. The dry seasons from November to April are remarked by grey bars. Each vertical bar represents an average (±SE) for that month.
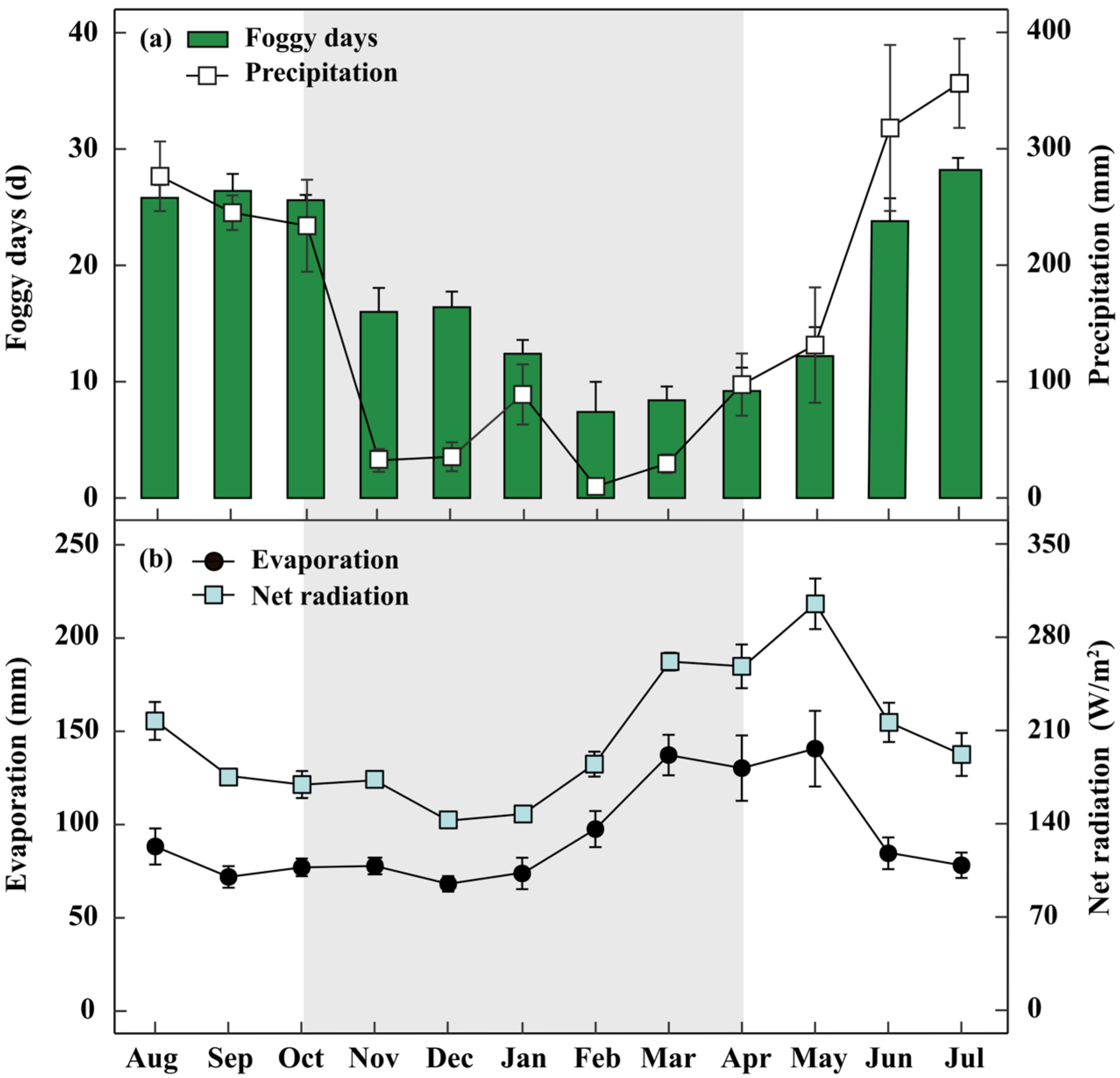
Figure 2.
Average hydrogen and oxygen isotope ratios (δ2H and δ18O) of epiphytes (Epiphytic lichens, n = 4 species; Epiphytic bryophytes, n = 4; Epiphytic ferns, n = 4; Epiphytic seed plants, n = 4) and water sources (Fog water, n = 7; humus, n = 4; and rainwater, n = 5) in the dry season (January 2019). The solid and segmented lines represent the global meteoric water line (GMWL: δ2H = 10 + 8 δ18O) and the local meteoric water line (LMWL: δ2H = 6.23 + 7.55 δ18O, R2 = 0.86, p < 0.001), respectively. The LMWL was calculated by linear regression of the δ2H and δ18O of local precipitation data from 2018 to 2019. Error bars represent mean ± SE of epiphytes and water sources.
Figure 2.
Average hydrogen and oxygen isotope ratios (δ2H and δ18O) of epiphytes (Epiphytic lichens, n = 4 species; Epiphytic bryophytes, n = 4; Epiphytic ferns, n = 4; Epiphytic seed plants, n = 4) and water sources (Fog water, n = 7; humus, n = 4; and rainwater, n = 5) in the dry season (January 2019). The solid and segmented lines represent the global meteoric water line (GMWL: δ2H = 10 + 8 δ18O) and the local meteoric water line (LMWL: δ2H = 6.23 + 7.55 δ18O, R2 = 0.86, p < 0.001), respectively. The LMWL was calculated by linear regression of the δ2H and δ18O of local precipitation data from 2018 to 2019. Error bars represent mean ± SE of epiphytes and water sources.
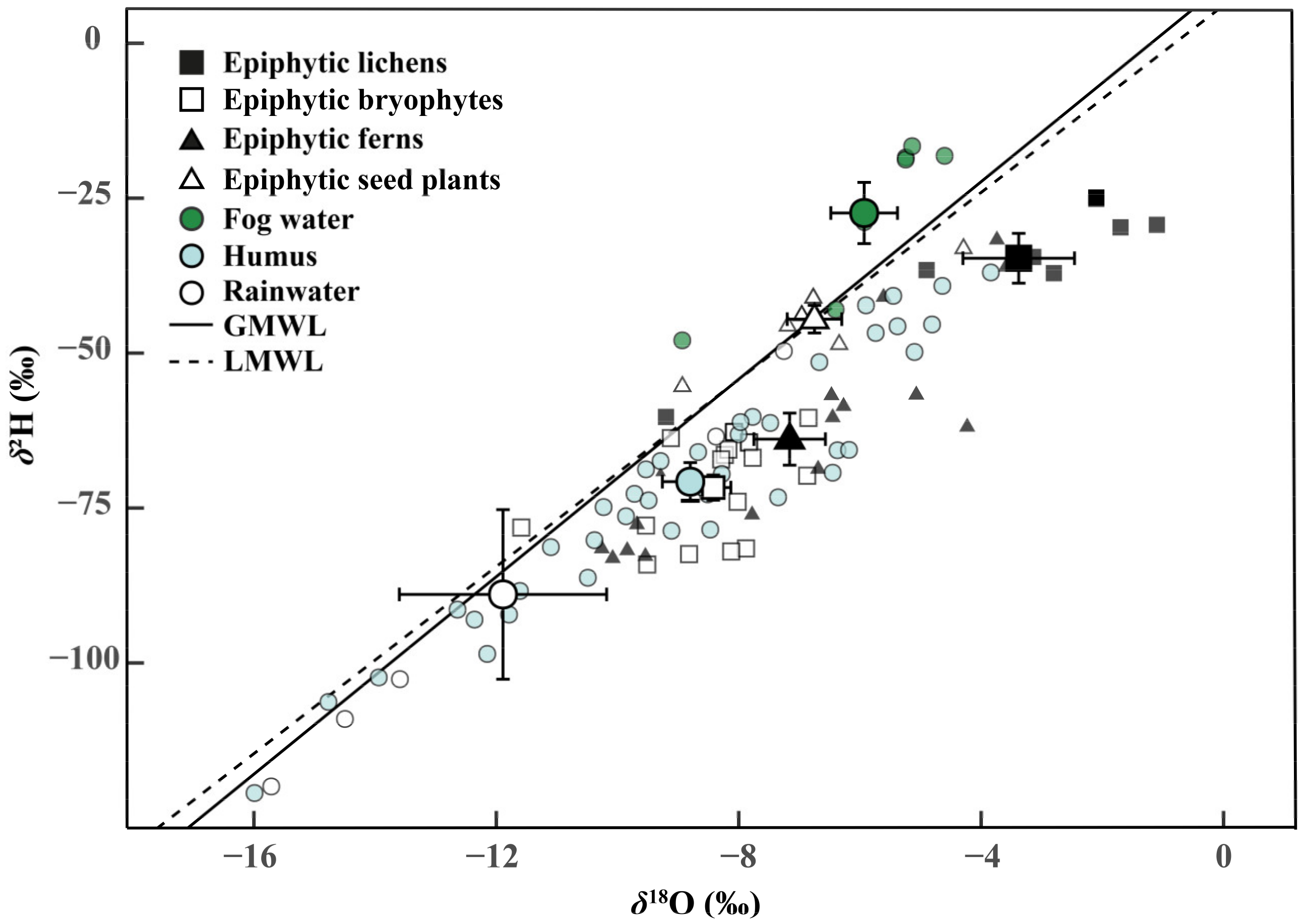
Figure 3.
The δ2H (a) and δ18O (‰) (b) of different water sources (Fog water, n = 7; humus, n = 4; and rainwater, n = 5) in the dry season, January 2019. Wilcoxon rank sum test is used to verify the differences of water source samples (NS > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001); Error bars represent means ± SEs of different water sources.
Figure 3.
The δ2H (a) and δ18O (‰) (b) of different water sources (Fog water, n = 7; humus, n = 4; and rainwater, n = 5) in the dry season, January 2019. Wilcoxon rank sum test is used to verify the differences of water source samples (NS > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001); Error bars represent means ± SEs of different water sources.
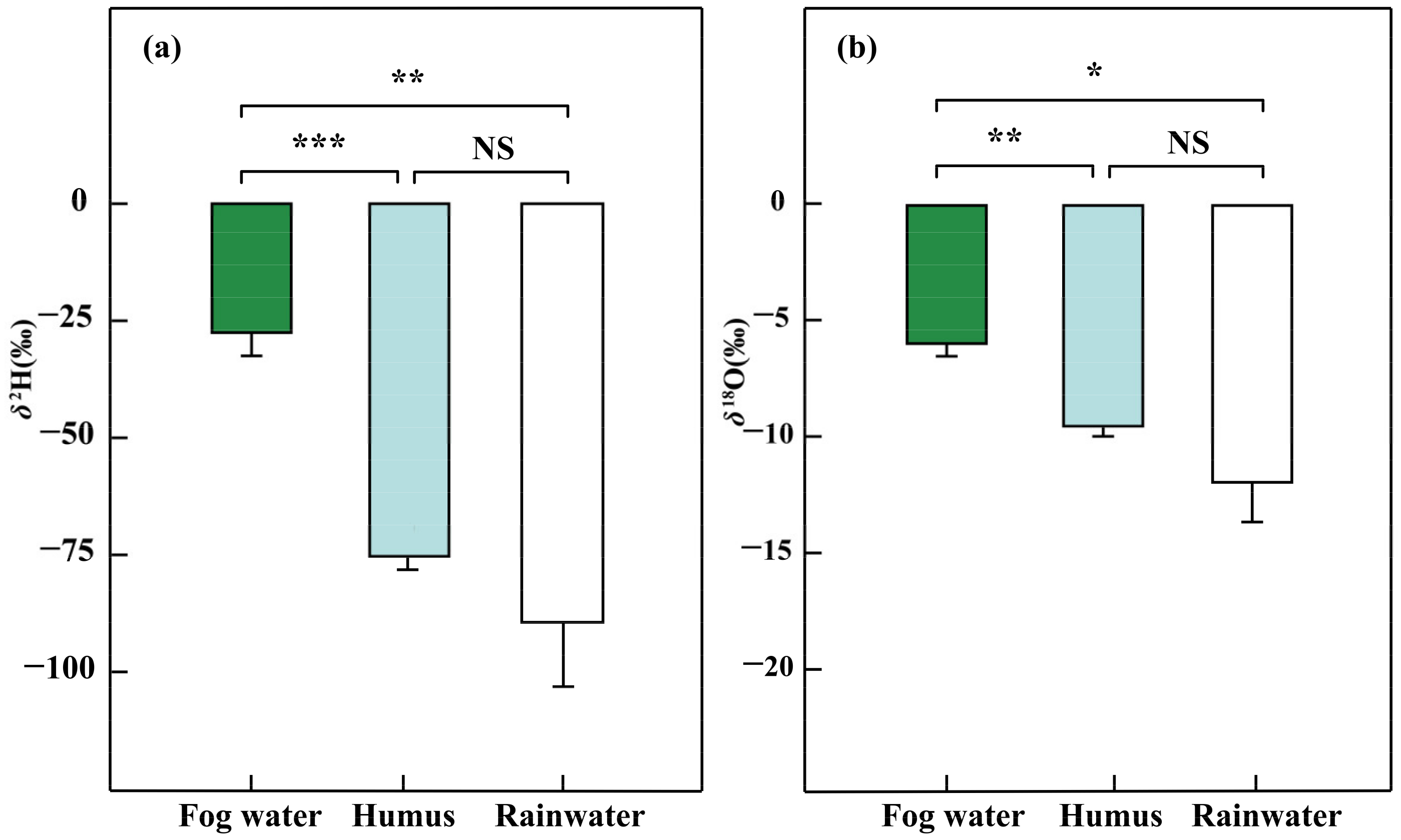
Figure 4.
The δ2H (a) and δ18O (‰) (b) of epiphytes from different groups. Epiphytic lichens (n = 4): NP, Nephromopsis pallescens; LR, Lobaria retigera. Epiphytic bryophytes (n = 4): HM, Hamaliodendron montagneanum; PA, Plagiochila assamica; BH, Bazzania himlayana; TC, Thuidium cymbifolium. Epiphytic ferns (n = 4): AI, Asplenium indicum; LL, Lepisorus loriformis; HP, Hymenophyllum polyanthos; LC, Loxogramme chinensis. Epiphytic seed plants (n = 4): AB, Aeschynanthus buxifolius; AM, Agapetes mannii.) in the dry season, January 2019. Wilcoxon rank sum test is used to verify the differences of epiphyte samples (NS > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001); Error bars represent mean ± SE, and different letters with bars represent significant differences for each species (p < 0.05).
Figure 4.
The δ2H (a) and δ18O (‰) (b) of epiphytes from different groups. Epiphytic lichens (n = 4): NP, Nephromopsis pallescens; LR, Lobaria retigera. Epiphytic bryophytes (n = 4): HM, Hamaliodendron montagneanum; PA, Plagiochila assamica; BH, Bazzania himlayana; TC, Thuidium cymbifolium. Epiphytic ferns (n = 4): AI, Asplenium indicum; LL, Lepisorus loriformis; HP, Hymenophyllum polyanthos; LC, Loxogramme chinensis. Epiphytic seed plants (n = 4): AB, Aeschynanthus buxifolius; AM, Agapetes mannii.) in the dry season, January 2019. Wilcoxon rank sum test is used to verify the differences of epiphyte samples (NS > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001); Error bars represent mean ± SE, and different letters with bars represent significant differences for each species (p < 0.05).
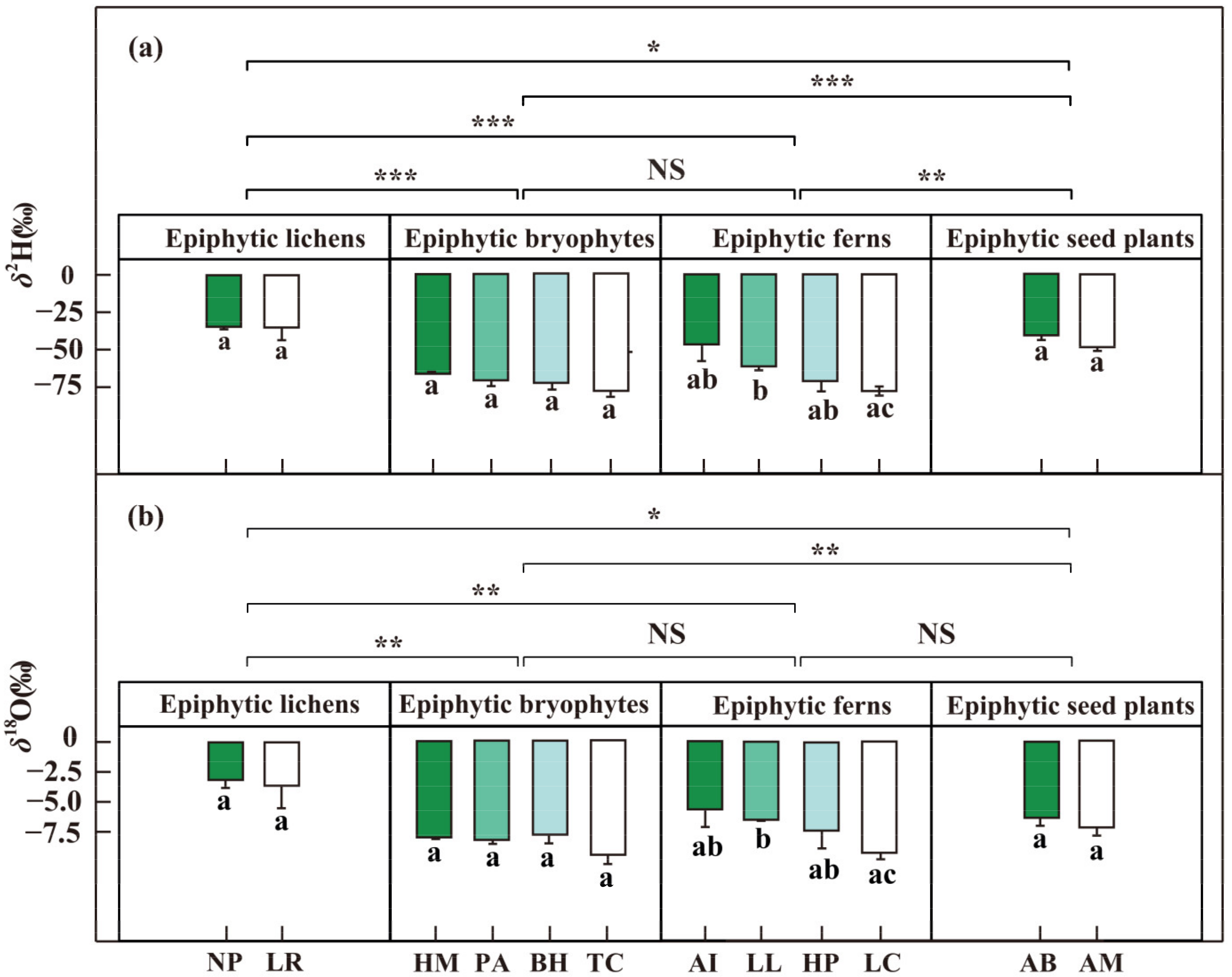
Figure 5.
Partitioning of potential water sources of epiphytes from different groups by MixSIAR model in a subtropical montane cloud forest in the dry season. The abbreviations of species names are the same as in Figure 4.
Figure 5.
Partitioning of potential water sources of epiphytes from different groups by MixSIAR model in a subtropical montane cloud forest in the dry season. The abbreviations of species names are the same as in Figure 4.
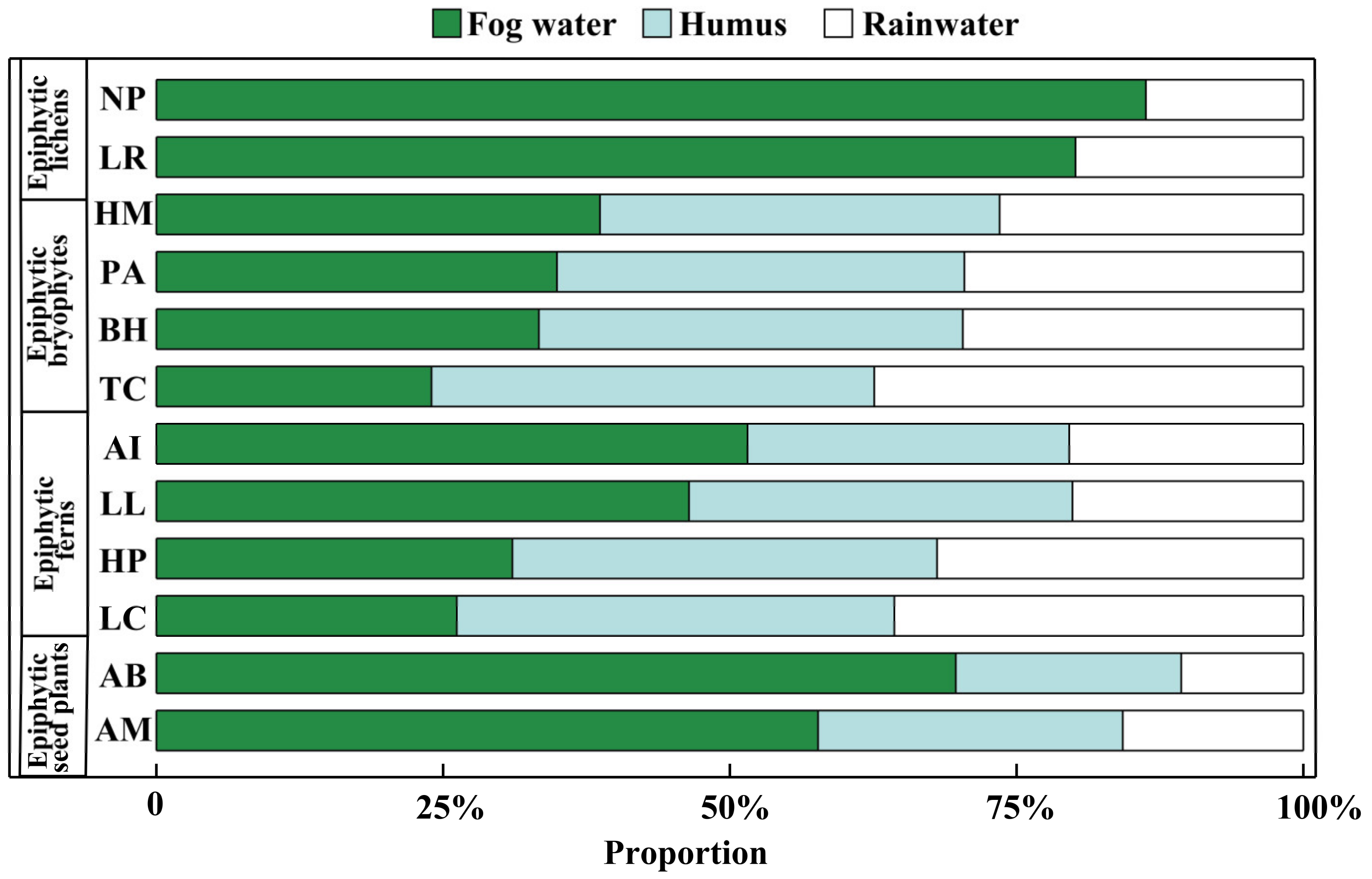
Figure 6.
The δ13C (‰) of mature leaves of epiphytes from different groups in the dry season, January 2019. Wilcoxon rank-sum test is used to verify the differences of epiphyte samples (NS > 0.05, *** p < 0.001); Error bars represent mean ± SE and different letters with bars represent significant differences for each species (p < 0.05). The abbreviations of species names are the same as in Figure 4.
Figure 6.
The δ13C (‰) of mature leaves of epiphytes from different groups in the dry season, January 2019. Wilcoxon rank-sum test is used to verify the differences of epiphyte samples (NS > 0.05, *** p < 0.001); Error bars represent mean ± SE and different letters with bars represent significant differences for each species (p < 0.05). The abbreviations of species names are the same as in Figure 4.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Linear mixed model analysis of the effects of δ2H, δ18O, and their interactions on . (δ13C) of epiphytes from different groups (* p < 0.05). The species of each group was incorporated as a random effect, but were not shown here for simplicity.
Table 1.
Linear mixed model analysis of the effects of δ2H, δ18O, and their interactions on . (δ13C) of epiphytes from different groups (* p < 0.05). The species of each group was incorporated as a random effect, but were not shown here for simplicity.
| Fixed Effect | Epiphytic Lichens | Epiphytic Bryophytes | Epiphytic Ferns | Epiphytic Seed Plants | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SE | χ2 | p | SE | χ2 | p | SE | χ2 | p | SE | χ2 | p | |
| δ2H | 0.115 | 4.624 | 0.032 * | 0.177 | 0.188 | 0.665 | 0.091 | 1.406 | 0.236 | 0.300 | 1.459 | 0.227 |
| δ18O | 0.091 | 2.394 | 0.122 | 0.197 | 0.002 | 0.965 | 0.077 | 0.065 | 0.065 | 0.194 | 3.644 | 0.056 |
| δ2H × δ18O | 1.283 | 9.236 | 0.026 * | 2.935 | 4.258 | 0.235 | 0.601 | 0.139 | 0.139 | 1.698 | 5.048 | 0.168 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, L.-L.; Yang, B.; Lu, H.-Z.; Wu, Y.; Meng, X.-J.; Zhang, Y.-J.; Song, L. Dry-Season Fog Water Utilization by Epiphytes in a Subtropical Montane Cloud Forest of Southwest China. Water 2021, 13, 3237. https://doi.org/10.3390/w13223237
AMA Style
Liu L-L, Yang B, Lu H-Z, Wu Y, Meng X-J, Zhang Y-J, Song L. Dry-Season Fog Water Utilization by Epiphytes in a Subtropical Montane Cloud Forest of Southwest China. Water. 2021; 13(22):3237. https://doi.org/10.3390/w13223237
Chicago/Turabian StyleLiu, Lu-Lu, Bin Yang, Hua-Zheng Lu, Yi Wu, Xian-Jing Meng, Yong-Jiang Zhang, and Liang Song. 2021. "Dry-Season Fog Water Utilization by Epiphytes in a Subtropical Montane Cloud Forest of Southwest China" Water 13, no. 22: 3237. https://doi.org/10.3390/w13223237
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.