Role of Homologous Recombination Genes in Repair of Alkylation Base Damage by Candida albicans


Departamento de Microbiología, Facultad de Ciencias, Universidad de Extremadura, 06071 Badajoz, Spain
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2018, 9(9), 447; https://doi.org/10.3390/genes9090447
Submission received: 14 August 2018
/
Accepted: 27 August 2018
/
Published: 7 September 2018
(This article belongs to the Special Issue Fungal Pathogenesis in Humans: The Growing Threat)
Abstract
:Candida albicans mutants deficient in homologous recombination (HR) are extremely sensitive to the alkylating agent methyl-methane-sulfonate (MMS). Here, we have investigated the role of HR genes in the protection and repair of C. albicans chromosomes by taking advantage of the heat-labile property (55 °C) of MMS-induced base damage. Acute MMS treatments of cycling cells caused chromosome fragmentation in vitro (55 °C) due to the generation of heat-dependent breaks (HDBs), but not in vivo (30 °C). Following removal of MMS wild type, cells regained the chromosome ladder regardless of whether they were transferred to yeast extract/peptone/dextrose (YPD) or to phosphate buffer saline (PBS); however, repair of HDB/chromosome restitution was faster in YPD, suggesting that it was accelerated by metabolic energy and further fueled by the subsequent overgrowth of survivors. Compared to wild type CAI4, chromosome restitution in YPD was not altered in a Carad59 isogenic derivative, whereas it was significantly delayed in Carad51 and Carad52 counterparts. However, when post-MMS incubation took place in PBS, chromosome restitution in wild type and HR mutants occurred with similar kinetics, suggesting that the exquisite sensitivity of Carad51 and Carad52 mutants to MMS is due to defective fork restart. Overall, our results demonstrate that repair of HDBs by resting cells of C. albicans is rather independent of CaRad51, CaRad52, and CaRad59, suggesting that it occurs mainly by base excision repair (BER).
1. Introduction
Methyl-methane-sulfonate (MMS) is used for the analysis of pathways involved in repair/tolerance to methylation [1]. Methyl-methane-sulfonate generates methylated bases on dsDNA whose repair can cause nicks, gaps, and, indirectly, double-strand breaks (DSBs) that can engage in homologous recombination (HR) directly or cause fork stalling during the next replication round [2,3,4]. Although methylated bases can be directly removed by DNA-methyl-transferases, the major repair pathway consists of a step-wise process known as base excision repair (BER) [1,5]. In Saccharomyces cerevisiae, BER is initiated by specific DNA-N-glycosylases that remove the damaged bases. The apurinic/apyrimidic (AP) sites generated are removed by redundant AP endonucleases Apn1 and Apn2, which cleave 5′ the AP-site to form nicks with a 5′ desoxyribose phosphate (5′dRP). Removal of 5′dRP is carried out by the coordinated action of DNA polymerase (δ or ε) and the flap endonuclease Rad27/Fen1, followed by ligation. Alternatively, AP-sites can be processed by unspecific Ntg1, Ntg2, or Ogg1 lyases to generate 3′AP sites (3′-dRP), which are then removed by the 3′-diesterase activity of Apn1/Apn2 or as part of an oligonucleotide generated by the endonuclease Rad1–Rad10. Finally, the gap is filled by DNA Pol and the backbone sealed by DNA ligase [1,4,6]. It is likely that similar enzymes and reactions account for BER in other fungi. However, only one APN endonuclease (APN1) and one NTG lyase (NTG1), in addition to an OGG1 homolog, have been so far identified and partially characterized in Candida albicans [7]. Importantly, null mutants in each of these C. albicans genes as well as the triple mutant exhibited wild type sensitivities to MMS suggesting the presence of redundant enzymes involved in repair of the methylated bases [7].
Importantly, in methylated DNA, AP-sites can arise from elimination of methylated bases by the action of glycosylase Mag1 [6] or, non-enzymatically, through the spontaneous depurination of methylated N3 and N7 purines, a process that is accelerated by heat [8]. It is well known that AP-sites are heat labile [9,10] and can be converted into single-strand breaks (SSBs) at 55 °C (referred to as HDB, for heat dependent breaks), a temperature generally used to prepare plugs for pulse-filed gel electrophoresis (PFGE) [11]. In fact, in S. cerevisiae stationary, or G1-arrested cells, DSBs and chromosome degradation was observed when methylated DNA or DNA carrying AP-sites was incubated at 55 °C [4,6,11]. Methyl-methane-sulfonate treatment can also cause opposed closely-spaced nicks in vivo (referred to as heat independent breaks (HIB) because they are detected when the same DNA samples are incubated at 30 °C), which can also result in secondary DSBs, and therefore in chromosome fragmentation [4,6,11]. For S. cerevisiae wild type, typical MMS treatments generate a large amount of HDBs and a low amount of HIB [12]. However, the latter are substantially increased in some BER-deficient cells as in apn1/apn2 double mutants [4,6,12].
Rad51 and Rad52 are evolutionary conserved proteins that play crucial roles in HR. In yeast, recombinase Rad51 is required for recombination pathways involving strand invasion. These pathways also require Rad52, which is thought to mediate the Rad51-ssDNA nucleofilament assembly. In addition, Rad52 participates in all recombination processes that require single strand annealing. For this reason, the rad52 mutation is epistatic to deletion of any other gene of the RAD52 epistasis group. Rad59 is a yeast paralog of Rad52 that exhibits strand annealing activity but lacks the ability of loading Rad51 onto ssDNA (for reviews see References [3,13]). Several studies have indicated that, although rad51 and rad52 mutants are extremely sensitive to MMS, HR does not play any role in the repair of MMS-born HIB of haploid S. cerevisiae G1 stationary cells which do not have a partner for engaging in HR, and the same was true for diploid G2-arrested cells which do [4,6,11,14]. However, Rad52 was crucial for that repair of HIB in G2-arrested cells in the absence of Anp1 and Anp2 endonucleases, suggesting that HR acts as a backup to repair lesions produced by AP-lyases Ntg1/2 and Ogg1 in the absence of Anp1,2 [6]. Besides, Rad51 and Rad52 are required for replication of methylated DNA [15,16,17] as well as for repair of the gaps generated in the process when cells reach the G2 phase [17]. Methyl-methane-sulfonate lesions and BER intermediates that are not repaired before they encounter the replication fork may cause replication fork stalling and collapse, unless stalled forks are bypassed by translation synthesis (stalled forks) causing increased mutagenesis or faithfully repaired by HR (both stalled and collapsed forks) [6,16,17]. The importance of HR in repair of methylated DNA extends to C. albicans where mutants affected in either HR (Carad52) or resection of DSBs (Carad50, Camre11) were significantly more sensitive to MMS than single mutants in BER genes [7,18].
In addition to MMS, a number of both endogenous and environmental agents including anticancer drugs (i.e., 4-methyl-5-oxo-2,3,4,6,8-pentazabicyclo[4.3.0]nona-2,7,9-triene-9-carboxamide temozolomide) can cause methylation damage [8,19,20], and therefore may affect viability and virulence of commensal opportunistic pathogens such as C. albicans. We previously reported that Carad52-ΔΔ, and to a lesser extent Carad51-ΔΔ cells from C. albicans, exhibit increased sensitivity to MMS [21,22]. In the current study, we have determined methylation base damage and recovery in C. albicans, taking advantage of the secondary DSBs and subsequent chromosome fragmentation generated during preparation of samples for pulse-field gel electrophoresis (PFGE) at 55 °C. We also show that resting cells of C. albicans can repair HDBs in the absence of HR, whereas repair of HDBs (and other BER intermediates) by cycling cell was mostly dependent on efficient HR.
2. Materials and Methods
2.1. Strains
The C. albicans and S. cerevisiae strains used in this study are shown in Table 1. Strain CAF2-1 derives from reference strain SC5314 by deletion of one copy of URA3 whereas CAI4 is the Uri− auxotrophic derivative of CAF2-1. All three strains are wild type for DNA recombination and repair. They were routinely grown in either yeast extract/peptone/dextrose (YPD) medium or synthetic complete (SC) medium supplemented with 33 mM uridine when necessary (i.e., Uri− strains) [23]. The haploid S. cerevisiae strain LSY0695-7D (Table 1) is a W303 derivative kindly provided by Lorraine Symington, from Columbia University [24]. Diploid W303 was obtained by standard genetic crosses [25].
2.2. DNA Extraction and Analysis and Cell Transformation
The DNA preparation for PFGE, as well as resolution of the samples, was carried out as reported before [29,30] using a Bio-Rad Chef Dry III. Gels were stained with ethidium bromide (0.5 µg/mL) for 2–4 h and imaged using a Molecular Imager (Bio-Rad Laboratories, Madrid, Spain). C. albicans cells were transformed using the lithium acetate method [31]. C. albicans chromosome fragments generated during incubation of methylated DNA with proteinase K at 55 °C were sized by using S. cerevisiae chromosomes as molecular weight (MW) markers (316–1091 kb) (see below).
A functional copy of the C. albicans URA3 marker was obtained by digestion of pLUBP plasmid with Pst1-BglII [32]. The resulting 4.9 kb fragment containing the URA3 gene was used to transform the Uri− strains CAI4 and its derivatives rad59-ΔΔ and rad51-ΔΔ. Uri+ transformants were selected on minimal SC medium minus uridine and correct integration was verified by PCR using oligonucleotide URA3 left flank and URA3 right flank [32].
2.3. Sensitivity to DNA-Damaging Agents
For determination of survival following an acute short-term MMS treatment, about 5 × 105 exponentially growing cells suspended in 1 mL YPD were incubated with MMS (0.05%, final concentration) for 30 min. Incubation mixtures were diluted 103-fold with PBS, and 50 µL (containing 300–400 colony forming units (CFU) before treatment) were plated on YPD plates for 48 h to determine the number of colonies. All the assays were done in duplicate and repeated four or more times.
2.4. Generation of MMS-Induced Heat Labile Breaks
Alkylation was induced as described by Lundin et al. [11]. Briefly, the indicated yeast strain was grown overnight in YPD until OD600 ≈ 5–7. MMS was added to a final concentration of 0.05% and cell suspensions were shaken at 30 °C for 15–30 min. The MMS was neutralized with 5% sodium thiosulfate by mixing 1:1 (v/v) ratio with 10% Na2S2O3 and washed twice with phosphate buffer saline (PBS). An aliquot was processed for PFGE (t = 0) and the rest was incubated in MMS-free YPD medium at 30 °C. At regular intervals (1, 2, 4, 8 and 24 h) samples were taken for determination of OD600, CFU, morphology, and repair of DNA damage (PFGE). Plugs for PFGE were prepared as described above and subsequently treated for 24 h with proteinase K (1 mg/mL) at 55 °C for analysis of HDBs or at 30 °C for analysis of HIBs as reported [4,6]. When indicated, thiosulfate neutralized cells were allowed to stand in PBS (resting cells) and samples were taken at the indicated times [4,6,11].
2.5. Calculations of Chromosome Fragments Sizes from Closely Spaced Single-Strand Breaks
Our calculations are based on the assumptions that SSBs (or heat-labile sites, in our assay) are distributed evenly between two DNA strands of a chromosome and that for a given SSB to form a DSB, a second SSB must appear on the opposite strand within an interval ≤S [6]. For further calculations of the number of DSBs, Ma et al. [6] used a circularized ChrIII, which only enters the gel following induction of a single DSB, to quantitatively determine the ratio ChrII to ChrIII using Southern blotting with a probe that hybridizes to both chromosomes. Since C. albicans does not maintain circular plasmids [28], we determined the range of sizes of the smear produced by the MMS treatment (0.05% MMS, 30 min) using the S. cerevisiae chromosomes as size markers. For S. cerevisiae diploid W303 strain (24 Mb), chromosome fragment sizes generated by MMS treatment ranged between 250 and 666 Kb, whose mean is 455 kb. Generation of uniform fragments of this size would require an average of 52 DSB per diploid genome or 26 DSB per haploid genome. For C. albicans (32 Mb), chromosome fragments sizes ranged between 316 and 1091 kb. By analogy, for a mean of 700kb we calculated 46 DSB per diploid genome or 23 DSB per haploid genome. Importantly, these values are not far from the range of 30 and 40 DSB per S. cerevisae haploid genome previously reported [6] for acute (30 min) 0.1% MMS treatments taking into account that we have used half MMS concentration (0.05% MMS) (Figure S1).
To quantify the extent of chromosome restitution during the post-MMS incubation, we estimated the intensities associated to Chr2, Chr5 bands (the same area for each chromosome) and smear from pulsed-field gels stained with ethidium bromide using the ImageJ software. Then, the ratio of the intensities Chr/smear at each time point was graphed versus the post-incubation time using Microsoft Excel. Data obtained with this approach agreed well with visual inspections of the gels. Chr2 and Chr5 were chosen as indicators of chromosome restitution because both are well resolved in PFGE and differ widely in size (2.23 Mb for Chr2 and 1.19 Mb for Chr5).
3. Results
3.1. The Role of HR Genes in Growth Polarization in Response to MMS
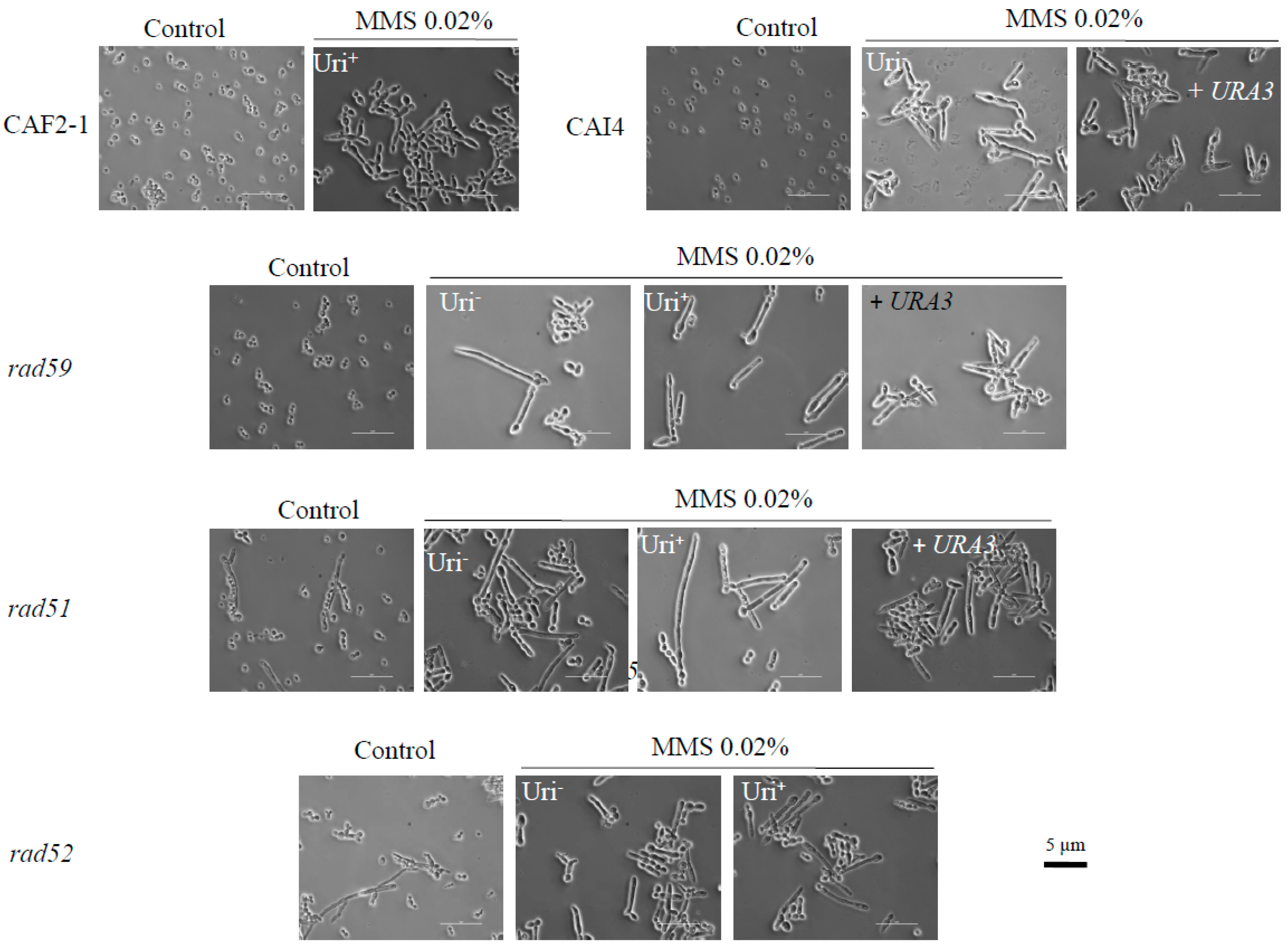
Genotoxic stress, including hydroxyurea and MMS treatments, triggers growth polarization of wild type C. albicans SC5314 and derivatives CAF2-1 (URA3/ura3) and CAI4 (ura3/ura3) (Table 1) generating elongated cells [33,34]. We have recently shown that the Uri− strain CAI4 and its derivatives carrying additional auxotrophies (RM10, RM1000, BWP37 and SN strains) exhibit an enhanced growth polarization and susceptibility to 0.02% MMS compared to parental CAF2-1 [35]. This differential behavior is due to a spontaneous loss of heterozygosity (LOH) event during the generation of CAI4 on the right arm of chromosome 3 (Chr3R) that homozygosed MBP1a, which regulates expression of DNA repair genes at G1/S phase of the cell cycle [35]. To investigate if MMS-induced filamentation was further affected by HR mutations, we subjected wild type and mutant strains to 0.02% MMS for 16 h at 30 °C in liquid YPD (Figure 1). As expected, CAF2-1 displayed chains of elongated cells but no filaments, whereas its Uri− derivative CAI4 showed long filaments. Reintegration of one copy of URA3 in its own locus improved growth rate but did not affect filamentation of the CAI4 strain [35]. All three Carad59-ΔΔ strains filamented as CAI4, regardless of whether they were Uri+ or Uri−, or if one copy of URA3 had been reintegrated into its own locus in the Uri− version of the mutant (Figure 1). The same was true for Carad51-ΔΔ strains in its Uri−, Uri+, or URA3-reintegrated versions (Figure 1). For Carad52-ΔΔ, Uri+ and Uri− versions behaved also similarly (Figure 1); however, as described [28], it was not possible to reintegrate URA3 into its own locus in rad52-ΔΔ strains. Importantly, in addition to the typical filamentous cells, Carad51-ΔΔ and Carad52-ΔΔ cultures also contain yeast cells [23]. In response to MMS, yeast cells also formed “germinative tubes” whose form and length were similar to their counterparts from Carad59-ΔΔ or CAI4 strains. We conclude that recombination mutants retain the ability to filament in response to MMS. It should be, finally, noted that cell elongation was a specific trait of C. albicans since it was not observed when diploid S. cerevisiae W303 or its Scrad52 derivative were incubated in MMS or during their post-MMS incubation in either PBS or YPD (not shown).
3.2. Generation and Repair of MMS-Induced Heat Labile Lesions on Growing C. albicans Wild Type Cells
Although MMS treatment of S. cerevisiae and human cells in vivo does not cause DSBs, methylated bases and AP-sites have been shown to be heat labile and converted into single strand breaks (SSBs) during the 55 °C proteinase K treatment used for preparation of PFGE plugs [6,9,11]. Furthermore, closely spaced SSBs located in opposite DNA strands may result in DSBs, and therefore chromosome fragmentation [6,9,11]. With these premises in mind, we investigated the extent to which MMS causes HDBs and HIBs in DNA from C. albicans.
Parental CAF2-1 strain exhibits the standard C. albicans karyotype. However, when medium-to-late exponentially growing cells (OD600 = 9; of note, when grown in YPD, C. albicans reaches OD600 up to 18) were subjected to an acute MMS treatment (0.05%, 15–30 min in YPD) and chromosome preparation for PFGE (which includes incubation with protease K) was conducted at 55 °C there was complete loss of chromosomal bands, which migrated now below Chr7 as a smear, indicating the presence of DSBs (Figure 2). Furthermore, the average size of the pool of degraded chromosomes obtained at 55 °C decreased with the incubation time in MMS, suggesting progressive chromosome fragmentation (Figure S2, lanes 1–5). By contrast, when following the acute MMS treatment (0.05%, 15–30 min in YPD) proteinase K incubation was conducted at 30 °C, strain CAF2-1 displayed the standard PFGE karyotype, and chromosome degradation was negligible (Figure S3, CAF2-1, lanes 1 and 2). Under these conditions, extension of the acute MMS treatment up to 120 min did not lead to a significant increase in chromosomal degradation (Figure S3, lanes 3–5). Some smear was likely due to the generation of a few closely opposed SSBs during manipulation of the samples, since it also was occasionally shown by preparations of untreated wild type cells incubated at 55 °C; however, the induction of small amounts of HIBs by MMS cannot be ruled out. As expected from previous reports [11,12], MMS-treated late-exponential phase cells of S. cerevisiae W303 subjected to the same process (i.e., preparation of samples for PFGE at 55 °C) displayed also chromosome fragmentation (Figure S1). These results indicate that almost all DSBs were generated during the incubation of C. albicans plugs with proteinase K at 55 °C and not in vivo, and accordingly provide an assessment of the overall number of HDBs [6]. Consistent with this interpretation, incubation of plugs at a lower temperature (50 °C) resulted in a reduced migration of the PFGE smear accompanied by the presence of vestiges of intact chromosomal bands (Figure S4, compare panels A and B, CAF2-1 lanes).
In S. cerevisiae, in vivo repair of the MMS-induced HDB prevented chromosome degradation in the subsequent 55 °C incubation with proteinase K [6]. In order to determine the time window required for repair of HDB in C. albicans, MMS-treated wild type CAF2-1 cells were post-incubated in either YPD or PBS lacking MMS and analyzed for chromosome restoration in time course experiments. Following a standard MMS treatment (0.05% MMS for 30 min in YPD and 55 °C incubations), no vestige of the chromosome ladder was apparent (Figure 3A, lanes 1 and 2). Traces of chromosomal bands were first seen after 2 h of recovery in YPD (Figure 3A, lane 4) and full restoration was accomplished by 4 h (Figure 3A, lane 5), with no significant changes being detected at later times (8–24 h) (Figure 3A, lanes 6 and 7). A similar conclusion was reached by quantification of Chr2 and Chr5 repair kinetics (Figure 3B). As expected from a true repair process, restoration of the chromosome ladder was paralleled by a significant reduction in the intensity of the smear (Figure 3A, compare lanes 2 to 7). It is worthy to mention that when PFGE samples were incubated at 50 °C “restitution” of the chromosomal ladder took only one hour (Figure S4A,B, lanes 1 to 3), as one could expect from the lower amount of SSBs generated at that temperature. Importantly, restitution of chromosomes was accelerated by the supply of metabolic energy (YPD) since it was significantly delayed when post-MMS incubation of CAF2-1 cells was carried out in PBS instead YPD (Figure 3A, lanes 8–12 and Figure 3B,C).
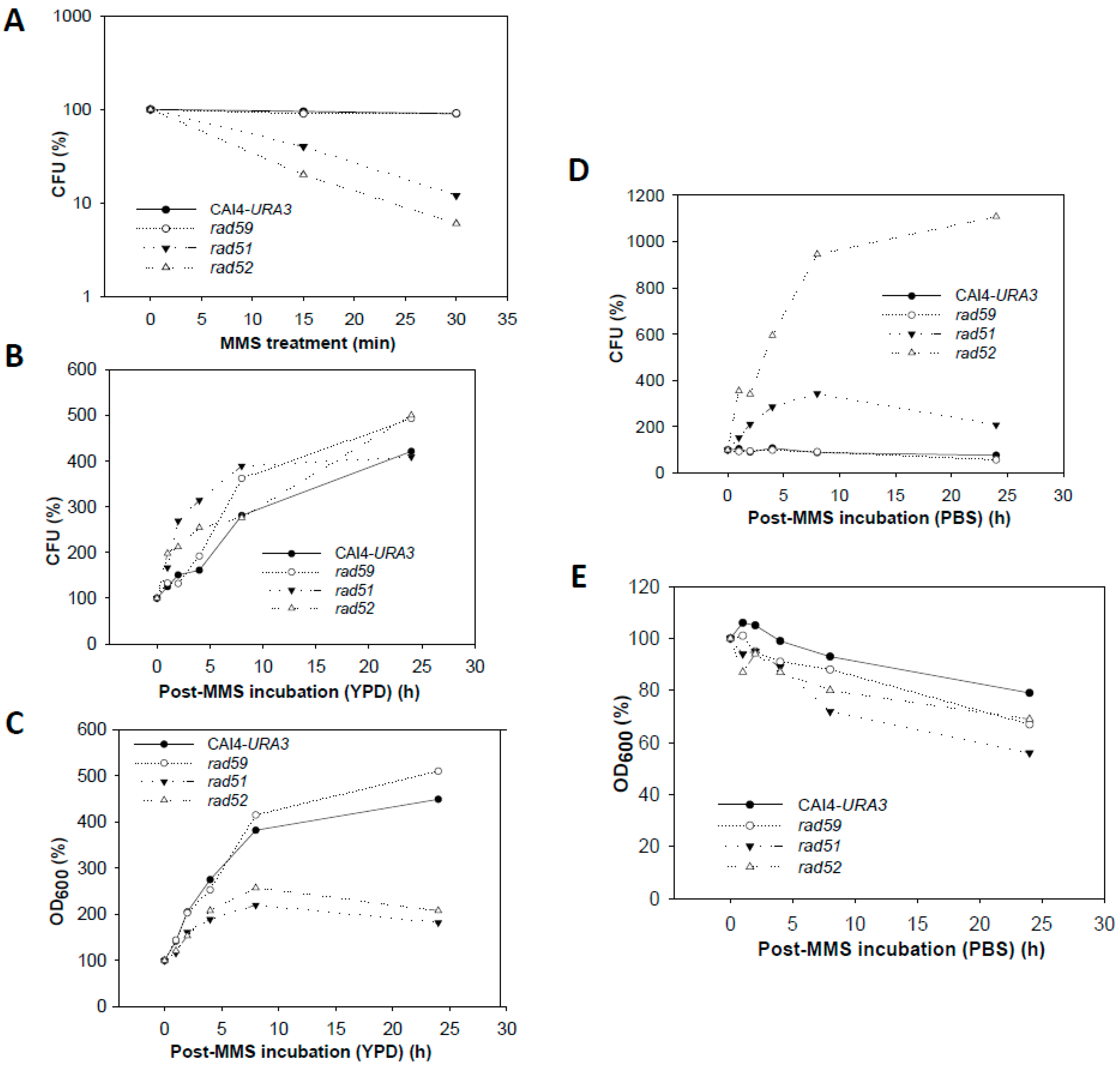
The CFU number of strain CAF2-1, that had been slightly reduced (4%) by the MMS-treatment, increased significantly throughout the post-MMS incubation in YPD (2–8 h) (Figure 3C). Importantly, by 4 h elongated cells carried apical and lateral buds further suggesting that they had undergone mitosis; later on, elongated branched cells steadily returned to the typical yeast form which became predominant by 24 h (Figure 4). In agreement with CFUs, the OD600 value steadily increased throughout the post-MMS incubation, including the first hour (Figure 3C), when few 55 °C -HDB had been repaired as indicated by the absence of chromosome “restitution” (see Figure 3A and Figure S4). It is likely that the initial increase in OD600 was mostly due to the formation of elongated cells (a retarded effect of the acute MMS treatment (Figure 1)), which reached maximal length by 2–4 h (Figure 4) whereas the late increase was due to cell division. Importantly, in contrast to the significant increase in cell number in YPD, neither cell division (Figure 3C) nor cell elongation (Figure 4) were detected when MMS-treated populations were transferred to PBS. Considering that under these conditions repair of HDB was significantly delayed, we conclude that overgrowth of survivors contributed significantly to the fast restitution of chromosomes during the post-MMS incubation in YPD (see Section 4, Discussion).
Because generation of CAI-4 from CAF2-1 resulted in increased sensitivity to low doses of MMS [35] (see above) and HR mutants (see below) were derived from CAI4, we compared chromosome recovery in MMS-treated CAF2-1 and CAI4 strains. As expected, CAI4 exhibited a small delay compared to CAF2-1 since no traces of chromosome bands could be detected in the former after 2 h of incubation in YPD. However, after 4 h both strains showed a full chromosome ladder (Figure S5).
3.3. Role of HR on the Repair of HDB
We have previously reported that C. albicans single mutants Carad51-ΔΔ, Carad52-ΔΔ, and Carad59-ΔΔ retained wild type PFGE karyotype profiles [21,22] (see also Figure 2 and Figure S3). When subjected to a standard acute MMS treatment and PFGE samples were incubated at 55 °C, all the mutants generated a smear similar to that shown by CAF2-1 cells (Figure 2). The average size of the pool of degraded chromosomes also decreased with the incubation time in MMS, suggesting a progressive accumulation of HDBs (Figure 2 and Figure S2). Besides, for each time-point (0–120 min in 0.05% MMS), the extent of chromosomal fragmentation was slightly higher for Carad51-ΔΔ and, to a larger extent, Carad52-ΔΔ strains compared to wild type suggesting that during the acute MMS treatment some repair in wild type is slowed down or blocked in Carad51-ΔΔ and Carad52-ΔΔ mutants (Figure S2). Finally, similarly to wild type, chromosomal fragmentation required in vitro incubations at high temperature since MMS-treated HR mutants also displayed standard chromosomal ladder when PFGE samples were incubated at 30 °C with proteinase K. Besides, prolongation of the MMS treatment up to 120 min did not result a significant increase in chromosomal degradation (Figure S3). We conclude that, as shown for wild type strain CAF2-1, treatment of HR mutants with MMS caused little or no induction of direct DSBs (HIBs).
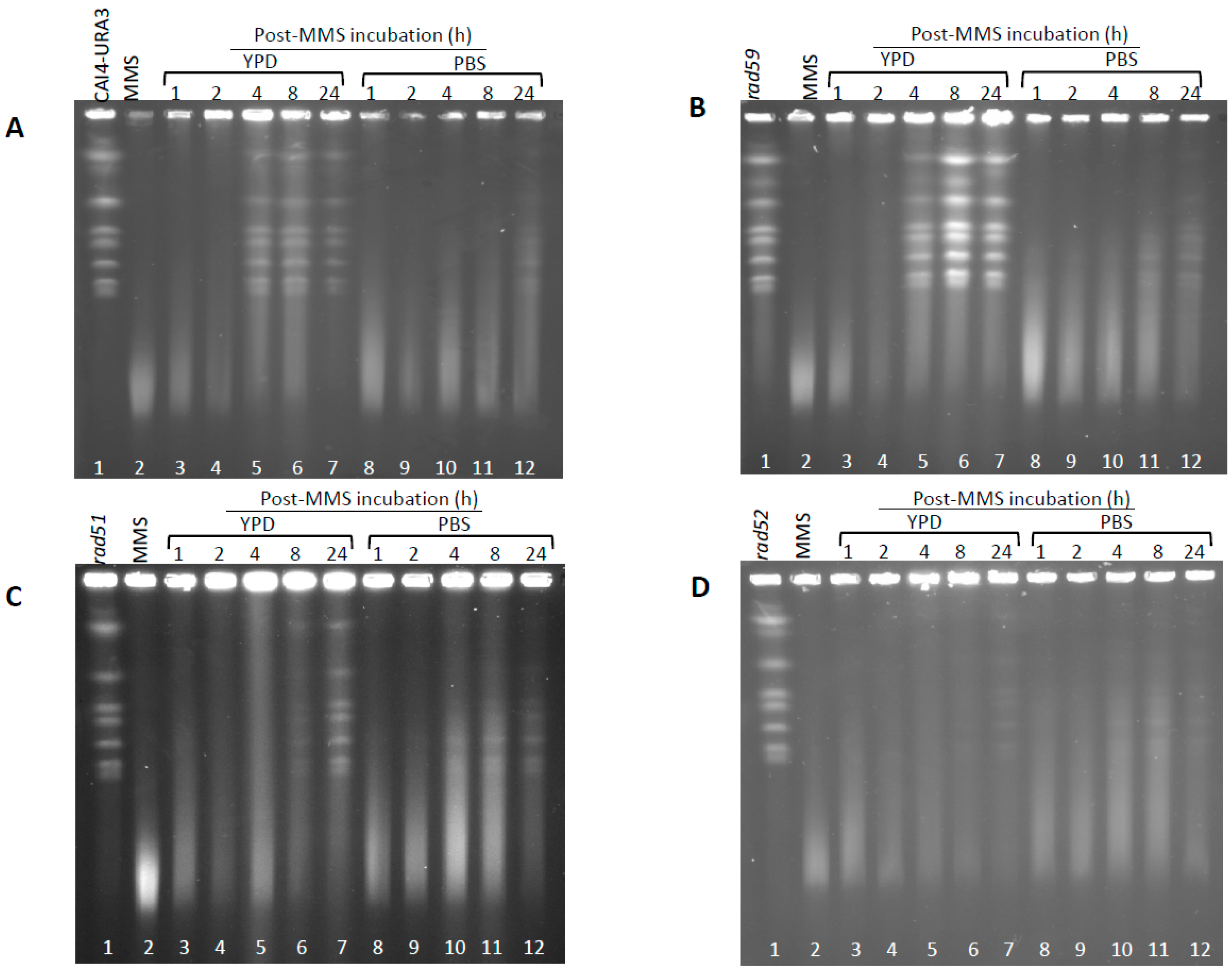
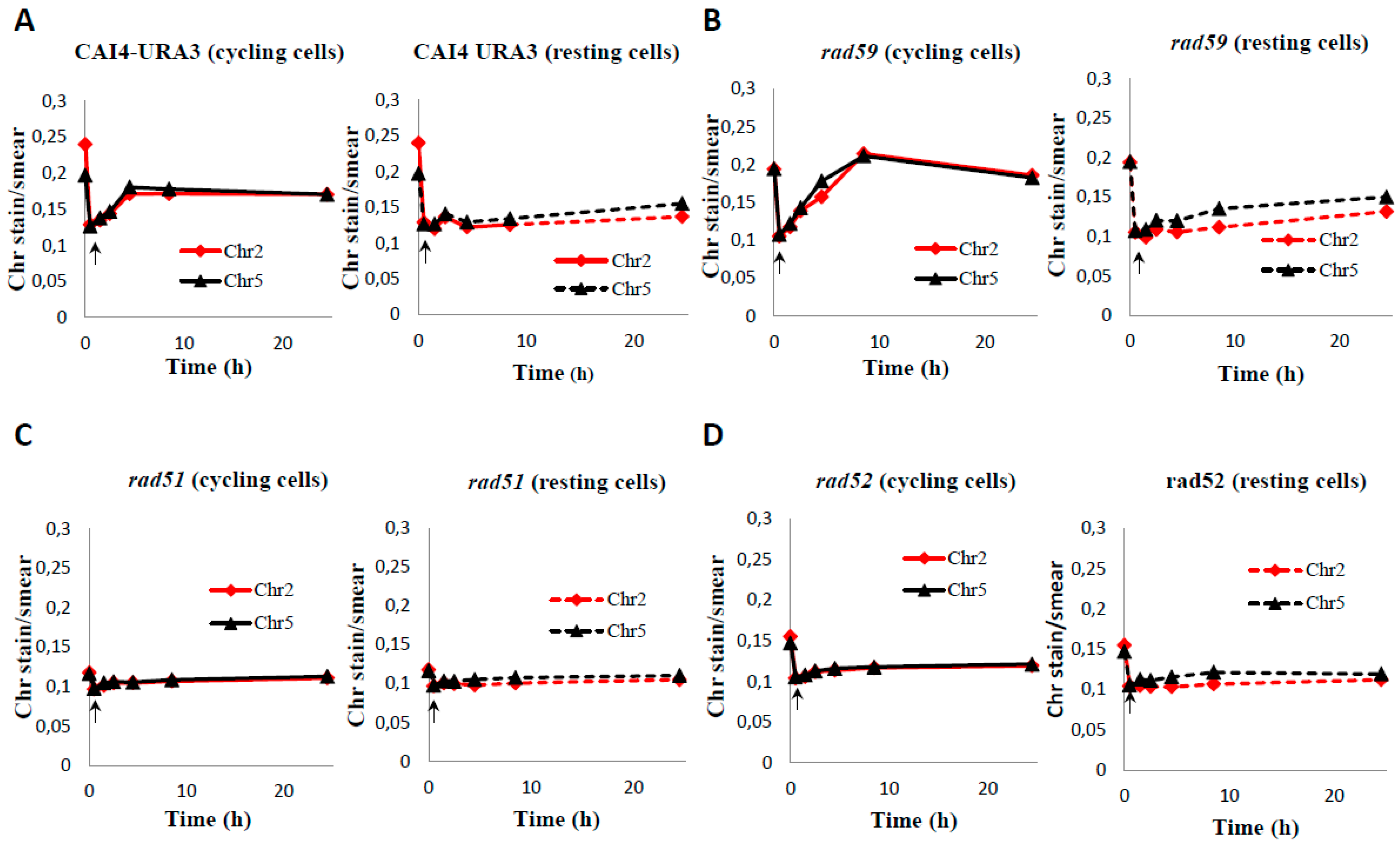
Next, we examined the role of HR proteins in repair of HDBs during the post-MMS incubation at 30 °C in either YPD or PBS. In YPD, the Carad59-ΔΔ mutant showed a delay (4 h) in the repair of HDB/“restitution” of chromosomal bands (incubations of plugs conducted at 55 °C), compared to the wild type CAF2-1 control (2 h) (Figure S4). This delay was also detected in a parallel experiment in which samples were incubated at 50 °C but, under these conditions, repair of HDBs by Carad59-ΔΔ and wild type took only two and one hours respectively (Figure S4). However, no noticeable differences between CAI4-URA3 and Carad59-ΔΔ strains were detected regardless the post-MMS incubation took place in YPD or PBS (Figure 5A,B), suggesting that MBP1a homozygosis in Carad59-ΔΔ and not the absence of Rad59 itself could be responsible for the delay in chromosome restitution in YPD when compared to CAF2-1 (MBP1a/MBP1b). Importantly, as shown above for CAF2-1 (Figure 3), chromosome restitution in CAI4-URA3 and Carad59-ΔΔ was accelerated in YPD (Figure 5 and Figure 6). In order to circumvent unwanted effects derived from the zygotic status of MBP1, CAI4-URA3 instead CAF2-1 was also used as a wild type control to investigate chromosome restitution in Carad51-ΔΔ and Carad52-ΔΔ mutants.
When post-incubated in YPD, MMS-treated Carad51-ΔΔ cultures exhibited a significant delay in chromosome restitution compared to CAI4-URA3 counterparts. As shown in Figure 5C and Figure 6C, Carad51-ΔΔ exhibited a progressive increase in the average size of the smear followed by the appearance of some faint bands by 8 h and distinct clear chromosomal bands by 24 h (Figure 5C, lanes 3–7 and Figure 6C). A similar restitution pattern was shown by the Carad52-ΔΔ mutant (Figure 5D, lanes 3–7 and Figure 6D). However, when the post-MMS incubation took place in PBS chromosome restitution in both mutants occurred with kinetics similar to those of wild type and Carad59-ΔΔ (Figure 5 and Figure 6). It is likely that replication of the few Carad51-ΔΔ and Carad52-ΔΔ survivors left by the MMS-treatment (12% and 7%, respectively) when transferred to YPD accounts by these differences.
3.4. Effect of the MMS Treatment on Cell Number and Morphology during the Post-Incubation Recovery of Mutants Populations
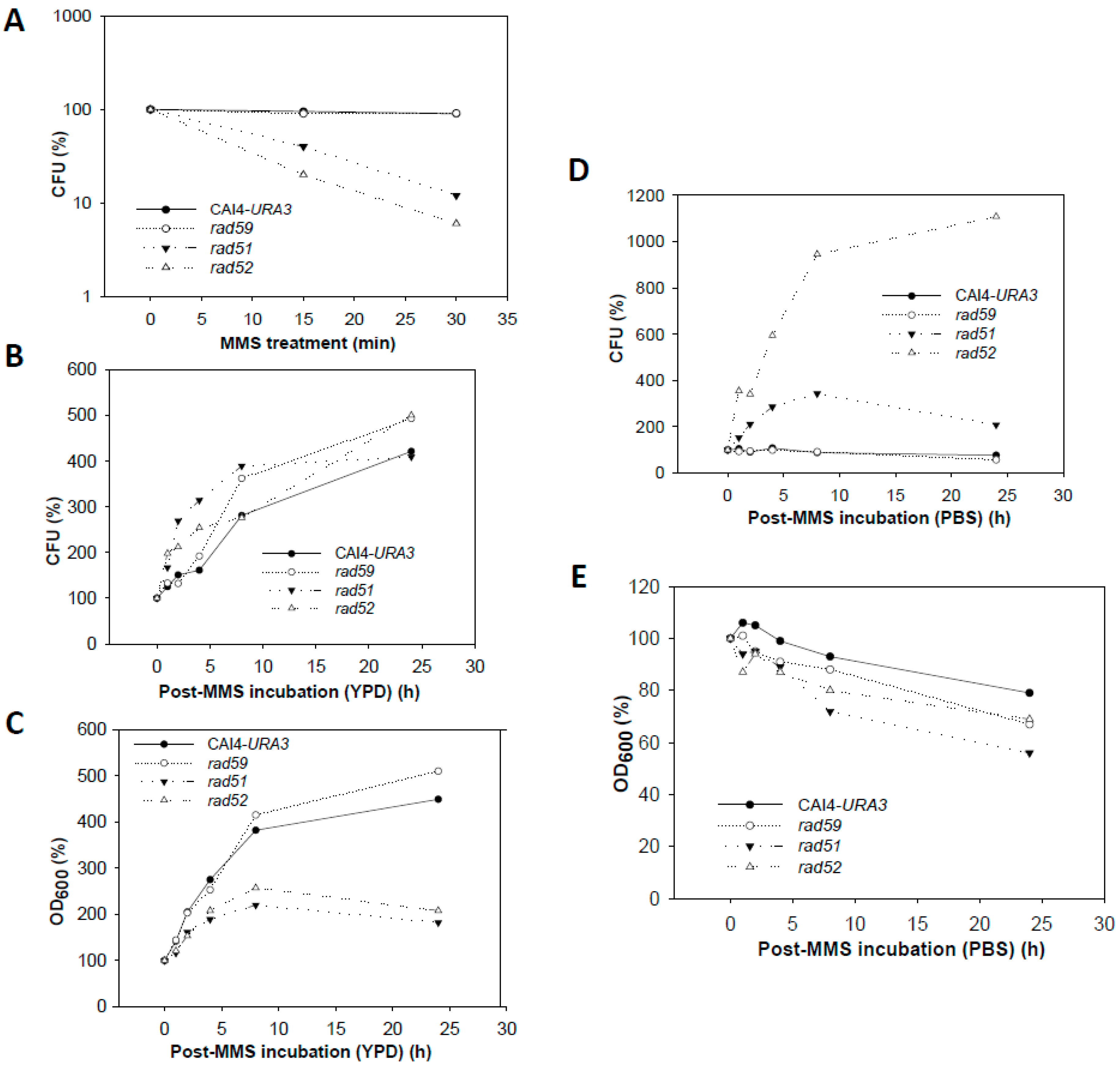
Null Carad59-ΔΔ showed CFU and OD600 values similar to those of wild type CAF2-1 throughout the MMS-treatment and post-MMS incubation (Figure 7), but, as shown for CAI4 (Figure 1), mutant cells displayed a more elongated morphology due to the MBP1a homozygosis (Figure 4). As expected, the MMS treatment strongly reduced Carad51-ΔΔ and Carad52-ΔΔ survival to 12% and 7%, respectively, in terms of CFUs (Figure 7A). For both mutants, CFUs and OD600 continuously increased during the post-MMS recovery in YPD indicating replication of survivors (Figure 7B,C). It is worthy to notice the exacerbated elongation of Carad51-ΔΔ and Carad52-ΔΔ cells throughout the first 8 h of post-recovery as well as their stickiness and subsequent tendency to form aggregates (Figure 4). Importantly, as shown for wild type strains, no changes in morphology and a gentle decrease in OD600 were observed during the post-MMS incubation of CAF2-1, Carad59-ΔΔ, Carad51-ΔΔ and Carad52-ΔΔ cells in PBS (Figure 4 and Figure 7E). However, in contrast to wild type and Carad59-ΔΔ, Carad51-ΔΔ and Carad52-ΔΔ CFUs increased significantly throughout post-MMS incubation in PBS despite the absence of cell replication (Figure 7D). It is likely that in the absence of nutrients damaged cells do not advance in the cell cycle and may repair HDBs by BER before being plated in YPD. Therefore, an increasing fraction of Carad51-ΔΔ and Carad52-ΔΔ cells can enter the S-phase with a repaired genome and give rise to healthy colonies on solid YPD.
4. Discussion
4.1. Role of HR in Repair of HDB by Proliferating C. Albicans Cells
In this study, we have analyzed the generation of HDBs by MMS in the genome of C. albicans as well as the role of HR genes CaRAD51, CaRAD52, and CaRAD59 in their repair. Importantly, we have used MMS concentrations reported not to cause direct DSBs in S. cerevisiae [9,12,36,37]. This was further confirmed by the absence of significant chromosome fragmentation in PFGE gels of MMS-treated C. albicans cells when incubation of plugs was conducted at 30 °C to detect exclusively HIB (Figure S3). As previously described [6], chromosome fragmentation was due to transformation of HDBs into SSBs at high temperature (55 °C). When close enough in opposite strands, SSBs are converted into secondary DSBs, which are manifested as genome shattering in PFGE gels. Our results are consistent with the observation that depurination of methylated DNA is a function of temperature [8]. When temperature dropped from 55 °C to 50 °C less HDBs/DSB were generated in vitro; this was reflected by the lower degree of chromosome fragmentation.
In S. cerevisiae repair of methylated bases by BER is constant and requires the action of the Mag1 glycosylase, which removes methylated bases leaving apurinic heat-sensitive sites throughout all the stages of the cell cycle [4,12,15]. However, Rad51 and Rad52 were also required for DNA replication in the presence of MMS [11,15,16] (see below). We found that for replicating cells of C. albicans, survival of an acute MMS-treatment was strongly dependent on HR proteins CaRad51 and, to a larger extent, CaRad52. In S. cerevisiae, the requirement of ScRad51 and ScRad52 for survival and further growth of survivors could be attributed to the role of both proteins in facilitating fork bypass through methylated DNA and subsequent repair of the resulting ssDNA gaps in G2/M [17]. It is likely that both functions are conserved in C. albicans where they are responsible for the rescue of >90% of damaged cells. In this scenario, the severe drop in survivability observed for cycling Carad51-ΔΔ and Carad52-ΔΔ cells is likely due to defective fork restart of damaged DNA. This S-phase block can potentially be bypassed by translesion synthesis which causes error-prone repair [38] and generates highly mutagenized survivors. In contrast to the high vulnerability of cells that transit S-phase, G1- or G2-cells present in our asynchronic cultures can potentially repair HDBs faithfully using BER and NER enzymes or other pathways before entering the S phase, thus providing additional non-mutagenized survivors [14,39]. In fact, the increase in CFU shown by Carad51-ΔΔ and Carad52-ΔΔ MMS-treated populations when incubated in YPD suggests that survivors have undergone at least one replication round (Figure 7), contributing in this way to restitution of the chromosome ladder. Consistent with this possibility, the relative amount of smear detected in PFGE gels by 8 and 24 h was significantly reduced.
4.2. Role of HR in Repair of HDB by Resting C. albicans Cells
Importantly, not only chromosomes were restituted, albeit with a lower kinetic, during the post-MMS incubation in PBS, but noticeable differences in repair kinetics between wild type and HR mutants Carad51-ΔΔ, Carad52-ΔΔ and Carad59-ΔΔ were not observed. Under these conditions, no cell proliferation was detected in wild type and Carad59-ΔΔ, but CFU increased in Carad51-ΔΔ and Carad52-ΔΔ mutants, indicating that HDBs are being repaired using pathways other than HR. Consistent with the absence of cell proliferation, OD600 did not increased (in fact some decrease was observed for all strains, likely as a consequence of cell lysis) and reduction of the smear throughout the post-MMS incubations was negligible compared to that observed in YPD. We conclude that in the absence of cell proliferation repair of HDBs caused by an acute MMS-treatment is independent of CaRad51, CaRad52, and CaRad59.
In S. cerevisiae, HR was shown to be crucial for the repair of alkylation damage by G2/M cells in the absence of Anp1 and Anp2 endonucleases, but not in its presence, suggesting that HR acts as a backup to repair lesions caused by unspecific Ntg1, Ntg2, and Ogg1 lyases [4]. Nothing is known on BER regulation in C. albicans. In this organism, only one ANP endonuclease (APN1) and one NTG lyase (NTG1), in addition to one OGG1 lyase, have been reported. Furthermore, single (anp1, ntg1, ogg1) and double (anp1 ntg1 and ogg1 ntg1) BER deletion mutants exhibited wild type sensitivity to MMS, while HR mutants were exquisitely sensitive, suggesting the existence of additional BER activities or that BER may be less important for repair of MMS damage in C. albicans compared to S. cerevisiae [7,18]. According to the present results, the single ANP endonuclease present in C. albicans seems to be able to repair methylation damage by resting cells whereas HR is mainly, if not exclusively, needed to allow replication of methylated DNA and further repair of the subsequent lesions when cells reach the G2/M phase. Work is in progress to determine if HR is needed for repair of methylation damage by G2/M-arrested C. albicans cells.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/9/9/447/s1. Figure S1. Determination of chromosome breaks (HDBs) following a MMS treatment. Figure S2. Kinetics of chromosome breaks (HDBs) throughout a prolonged MMS treatment. Figure S3. Determination of heat independent breaks (HIBs) following a prolonged MMS treatment. Figure S4. Effect of temperature on the induction and repair of HDBs by C. albicans cycling cells. C. albicans wild type (CAF2-1) and Carad59-mutant were subjected to an acute MMS treatment (0.05% MMS for 30 min). Figure S5. Influence of the MBP1 homozygosis on the repair of HDBs by C. albicans cycling cells.
Author Contributions
Conceived and designed the experiments: G.L. Performed the experiments: T.C., A.B., E.A. and B.H. Edited the manuscript: G.L. and T.C.
Funding
This research received no external funding.
Acknowledgments
We thank to Andrés Aguilera (CABIMER and Universidad de Sevilla) for allowing us to use his laboratory for several assays and Wenjian Ma for critical reading of an early version of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Boiteux, S.; Jinks-Robertson, S. DNA repair mechanisms and the bypass of DNA damage in Saccharomyces cerevisiae. Genetics 2013, 193, 1025–1064. [Google Scholar] [CrossRef] [PubMed]
- Budzowska, M.; Kanaar, R. Mechanisms of dealing with DNA damage-induced replication problems. Cell Biochem. Biophys. 2009, 53, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Heyer, W.D.; Ehmsen, K.T.; Liu, J. Regulation of homologous recombination in eukaryotes. Annu. Rev. Genet. 2010, 44, 113–139. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Westmoreland, J.W.; Gordenin, D.A.; Resnick, M.A. Alkylation base damage is converted into repairable double-strand breaks and complex intermediates in G2 cells lacking AP endonuclease. PLoS Genet. 2011, 7, e1002059. [Google Scholar] [CrossRef] [PubMed]
- Sedgwick, B.; Bates, P.A.; Paik, J.; Jacobs, S.C.; Lindahl, T. Repair of alkylated DNA: Recent advances. DNA Repair 2007, 6, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Resnick, M.A.; Gordenin, D.A. Apn1 and Apn2 endonucleases prevent accumulation of repair-associated DNA breaks in budding yeast as revealed by direct chromosomal analysis. Nucleic Acids Res. 2008, 36, 1836–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, M.; Chan, C.L.; Jauert, P.A.; Kirkpatrick, D.T. Analysis of base excision and nucleotide excision repair in Candida albicans. Microbiology 2008, 154, 2446–2456. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, M.D.; Pittman, D.L. Methylating agents and DNA repair responses: Methylated bases and sources of strand breaks. Chem. Res. Toxicol. 2006, 19, 1580–1594. [Google Scholar] [CrossRef] [PubMed]
- Valenti, A.; Napoli, A.; Ferrara, M.C.; Nadal, M.; Rossi, M.; Ciaramella, M. Selective degradation of reverse gyrase and DNA fragmentation induced by alkylating agent in the archaeon Sulfolobus solfataricus. Nucleic Acids Res. 2006, 34, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Andersson, A. Rate of chain breakage at apurinic sites in double-stranded deoxyribonucleic acid. Biochemistry 1972, 11, 3618–3623. [Google Scholar] [CrossRef] [PubMed]
- Lundin, E.; North, M.; Erixon, K.; Walters, K.; Jenssen, D.; Goldman, A.S.; Helleday, T. Methyl methane sulfonate (MMS) produces heat-labile DNA damage but no detectable in vivo DNA double-strand breaks. Nucleic Acids Res. 2005, 33, 3799–3811. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Panduri, V.; Sterling, J.F.; Van Houten, B.; Gordenin, D.A.; Resnick, M.A. The transition of closely opposed lesions to double-strand breaks during long-patch base excision repair is prevented by the coordinated action of DNA polymerase delta and Rad27/Fen1. Mol. Cell. Biol. 2009, 29, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- Symington, L.S.; Rothstein, R.; Lisby, M. Mechanisms and regulation of mitotic recombination in Saccharomyces cerevisiae. Genetics 2014, 198, 795–835. [Google Scholar] [CrossRef] [PubMed]
- Begley, T.J.; Rosenbach, A.S.; Ideker, T.; Samson, L.D. Damage recovery pathways in Saccharomyces cerevisiae revealed by genomic phenotyping and interactome mapping. Mol. Cancer Res. 2002, 1, 103–112. [Google Scholar] [PubMed]
- Vázquez, M.V.; Rojas, V.; Tercero, J.A. sMultiple pathways cooperate to facilitate DNA replication fork progression through alkylated DNA. DNA Repair 2008, 7, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Tercero, J.A.; Diffley, J.F. Regulation of DNA replication fork progression through damaged DNA by the Mec1/Rad53 checkpoint. Nature 2001, 412, 553–557. [Google Scholar] [CrossRef] [PubMed]
- González-Prieto, R.; Muñoz-Cabello, A.M.; Cabello-Lobato, M.J.; Prado, F. Rad51 replication fork recruitment is required for DNA damage tolerance. EMBO J. 2013, 32, 1307–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, M.; Chan, C.L.; Jauert, P.A.; Kirkpatrick, D.T. Role of DNA mismatch repair and double-strand break repair in genome stability and antifungal drug resistance in Candida albicans. Eukaryot. Cell 2007, 6, 2194–2205. [Google Scholar] [CrossRef] [PubMed]
- Drablos, F.; Feyzi, E.; Aas, P.A.; Vaagbø, C.B.; Kavli, B.; Bratlie, M.S.; Peña-Diaz, J.; Otterlei, M.; Slupphaug, G.; Krokan, H.E. Alkylation damage in DNA and RNA-repair mechanisms and medical significance. DNA Repair 2004, 3, 1389–1407. [Google Scholar] [CrossRef] [PubMed]
- Larson, K.; Sahm, J.; Shenkar, R.; Strauss, B. Methylation-induced blocks to in vitro DNA replication. Mutat. Res. 1985, 150, 77–84. [Google Scholar] [CrossRef]
- García-Prieto, F.; Gómez-Raja, J.; Andaluz, E.; Calderone, R.; Larriba, G. Role of the homologous recombination genes RAD51 and RAD59 in the resistance of Candida albicans to UV light, radiomimetic and anti-tumor compounds and oxidizing agents. Fungal Genet. Biol. 2010, 47, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Bellido, A.; Andaluz, E.; Gómez-Raja, J.; Álvarez-Barrientos, A.; Larriba, G. Genetic interactions among homologous recombination mutants in Candida albicans. Fungal Genet. Biol. 2015, 74, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Andaluz, E.; Ciudad, T.; Gómez-Raja, J.; Calderone, R.; Larriba, G. Rad52 depletion in Candida albicans triggers both the DNA-damage checkpoint and filamentation accompanied by but independent of expression of hypha-specific genes. Mol. Microbiol. 2006, 59, 1452–1472. [Google Scholar] [CrossRef] [PubMed]
- Bärtsch, S.; Kang, L.E.; Symington, L.S. RAD51 is required for the repair of plasmid double-stranded gaps from either plasmid or chromosomal templates. Mol. Cell. Biol. 2000, 20, 1194–1205. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, C.; Fink, G.R. Guide to Yeast Genetics and Molecular Biology; Academic Press: San Diego, CA, USA, 1991. [Google Scholar]
- Gillum, A.M.; Tsay, E.Y.; Kirsch, D.R. Isolation of the Candida albicans gene for orotidine-5′-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 1984, 198, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar]
- Ciudad, T.; Andaluz, E.; Steinberg-Neifach, O.; Lue, N.F.; Gow, N.A.; Calderone, R.A.; Larriba, G. Homologous recombination in Candida albicans: Role of CaRad52p in DNA repair, integration of linear DNA fragments and telomere length. Mol. Microbiol. 2004, 53, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Andaluz, E.; Gómez-Raja, J.; Hermosa, B.; Ciudad, T.; Rustchenko, E.; Calderone, R.; Larriba, G. Loss and fragmentation of chromosome 5 are major events linked to the adaptation of rad52-DeltaDelta strains of Candida albicans to sorbose. Fungal Genet. Biol. 2007, 44, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Andaluz, E.; Bellido, A.; Gómez-Raja, J.; Selmecki, A.; Bouchonville, K.; Calderone, R.; Berman, J.; Larriba, G. Rad52 function prevents chromosome loss and truncation in Candida albicans. Mol. Microbiol. 2011, 79, 1462–1482. [Google Scholar] [CrossRef] [PubMed]
- Walther, A.; Wendland, J. An improved transformation protocol for the human fungal pathogen Candida albicans. Curr. Genet. 2003, 42, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.M.; Johnson, A.D. Strains and strategies for large-scale gene deletion studies of the diploid human fungal pathogen Candida albicans. Eukaryot. Cell 2005, 4, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.M.; Wang, Y.M.; Zheng, X.D.; Lee, R.T.; Wang, Y. Critical role of DNA checkpoints in mediating genotoxic-stress-induced filamentous growth in Candida albicans. Mol. Biol. Cell 2007, 18, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Legrand, M.; Chan, C.L.; Jauert, P.A.; Kirkpatrick, D.T. The contribution of the S-phase checkpoint genes MEC1 and SGS1 to genome stability maintenance in Candida albicans. Fungal Genet. Biol. 2011, 48, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Ciudad, T.; Hickman, M.; Bellido, A.; Berman, J.; Larriba, G. Phenotypic consequences of a spontaneous loss of heterozygosity in a common laboratory strain of Candida albicans. Genetics 2016, 203, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Covo, S.; Westmoreland, J.W.; Gordenin, D.A.; Resnick, M.A. Cohesin is limiting for the suppression of DNA damage-induced recombination between homologous chromosomes. PLoS Genet. 2010, 6, e1001006. [Google Scholar] [CrossRef] [PubMed]
- Redon, C.; Pilch, D.R.; Rogakou, E.P.; Orr, A.H.; Lowndes, N.F.; Bonner, W.M. Yeast histone 2A serine 129 is essential for the efficient repair of checkpoint-blind DNA damage. EMBO Rep. 2003, 4, 678–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auerbach, P.A.; Demple, B. Roles of Rev1, Pol zeta, Pol32 and Pol eta in the bypass of chromosomal abasic sites in Saccharomyces cerevisiae. Mutagenesis 2010, 25, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Calvo, J.A.; Samson, L.D. Balancing repair and tolerance of DNA damage caused by alkylating agents. Nat. Rev. Cancer 2013, 12, 104–120. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Methyl-methane-sulfonate (MMS) induces constitutive filamentous growth in wild type and HR mutants of Candida albicans. A yeast extract/peptone/dextrose (YPD) overnight culture of exponentially growing cells from the indicated strains was refreshed and adjusted to OD600 = 1. Following a further incubation for 2 h at 30 °C with shaking, one half was suspended in YPD supplemented or not (control) with 0.02% MMS. After 12 h at 30 °C, with gentle shaking, samples were photographed using a Nikon Eclipse 600 microscope with a 60× DIC objective. A CC-12 digital camera interfaced with Soft Imaging System software was used for imaging (Izasa Scientific, Alcobendas, Madrid, Spain). Each bar corresponds to 5 µm.
Figure 1.
Methyl-methane-sulfonate (MMS) induces constitutive filamentous growth in wild type and HR mutants of Candida albicans. A yeast extract/peptone/dextrose (YPD) overnight culture of exponentially growing cells from the indicated strains was refreshed and adjusted to OD600 = 1. Following a further incubation for 2 h at 30 °C with shaking, one half was suspended in YPD supplemented or not (control) with 0.02% MMS. After 12 h at 30 °C, with gentle shaking, samples were photographed using a Nikon Eclipse 600 microscope with a 60× DIC objective. A CC-12 digital camera interfaced with Soft Imaging System software was used for imaging (Izasa Scientific, Alcobendas, Madrid, Spain). Each bar corresponds to 5 µm.
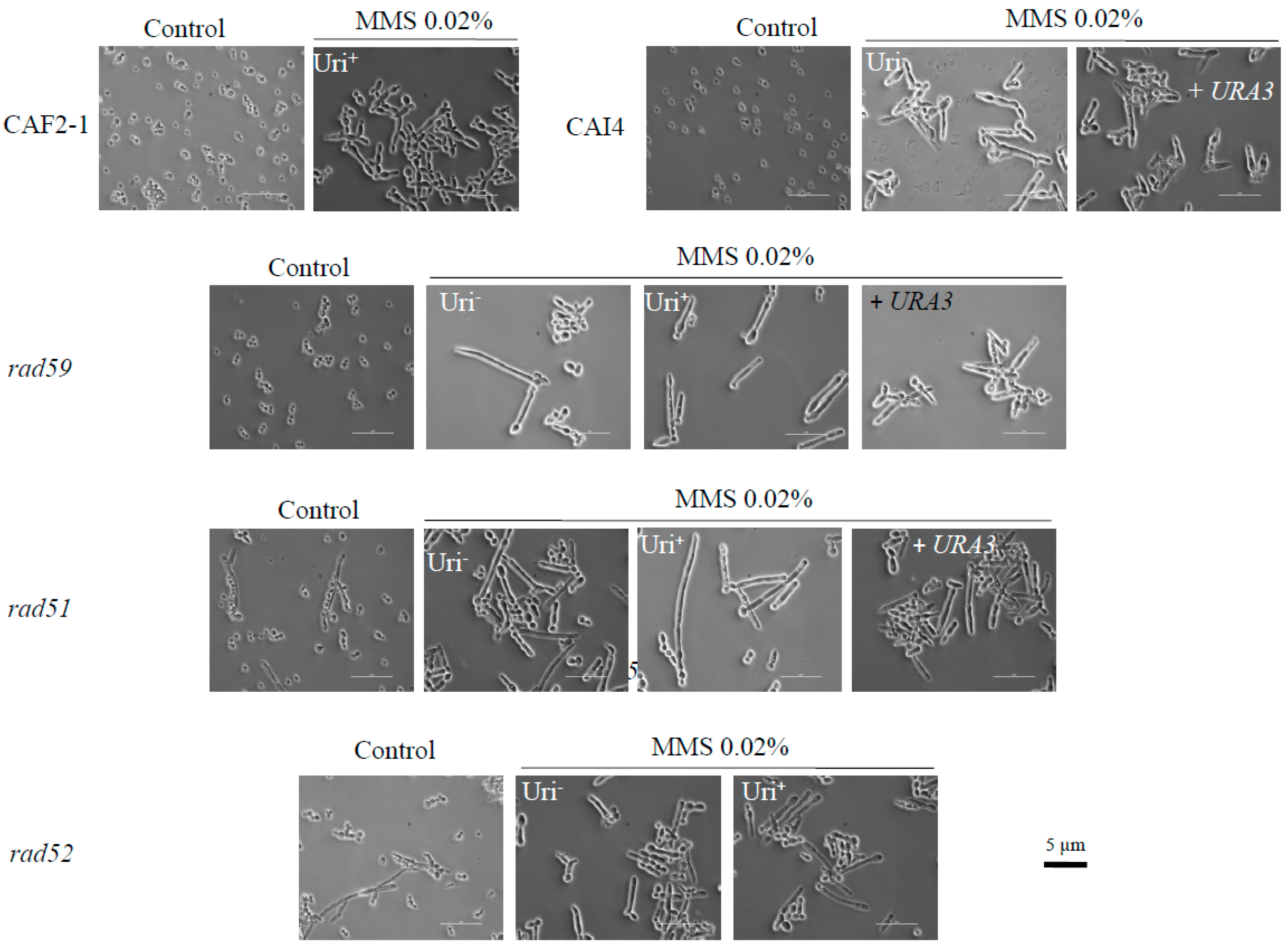
Figure 2.
Determination of chromosome breaks (HDBs) following acute MMS treatments. Cells from wild type CAF2-1 and the several HR mutant derivatives were treated with 0.05% MMS for 15 and 30 min, followed by immediate DNA purification and pulse-field gel electrophoresis (PFGE). Proteinase K digestion was carried out at 55 °C to detect heat dependent breaks (HDBs). Chromosomes were visualized with ethidium bromide staining (see also Figure S2).
Figure 2.
Determination of chromosome breaks (HDBs) following acute MMS treatments. Cells from wild type CAF2-1 and the several HR mutant derivatives were treated with 0.05% MMS for 15 and 30 min, followed by immediate DNA purification and pulse-field gel electrophoresis (PFGE). Proteinase K digestion was carried out at 55 °C to detect heat dependent breaks (HDBs). Chromosomes were visualized with ethidium bromide staining (see also Figure S2).
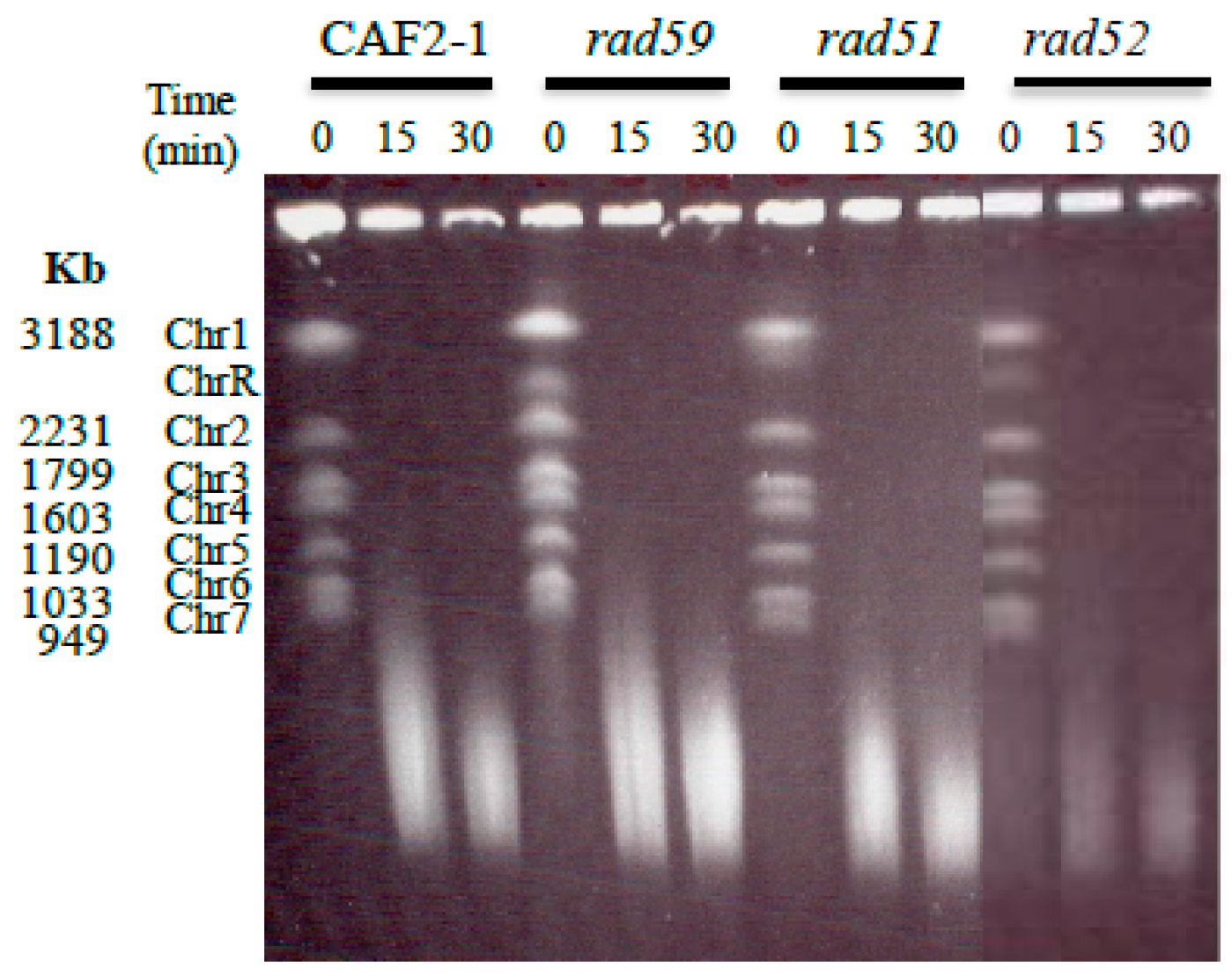
Figure 3.
Induction and repair of HDBs by C. albicans cycling and resting wild type cells. C. albicans wild type (CAF2-1) was subjected to an acute MMS treatment (0.05% MMS for 30 min). Then cells were treated with 5% sodium thiosulphate, by mixing 1:1 (v/v) ratio with 10% Na2S2O3, washed with PBS, and transferred to MMS-free YPD (cycling cells) or PBS (resting cells) at 30 °C. Samples were taken at the indicated times to determine PFGE profiles (A), to quantify Chr2 and Chr5 (B) and to calculate CFU and OD600 (C). For PFGE profiles, DNA plugs were incubated with proteinase K at 55 °C. Of note, in panels B and C, initial time (0) corresponds to samples before MMS treatment. Arrows in panel B shows the MMS-treated sample.
Figure 3.
Induction and repair of HDBs by C. albicans cycling and resting wild type cells. C. albicans wild type (CAF2-1) was subjected to an acute MMS treatment (0.05% MMS for 30 min). Then cells were treated with 5% sodium thiosulphate, by mixing 1:1 (v/v) ratio with 10% Na2S2O3, washed with PBS, and transferred to MMS-free YPD (cycling cells) or PBS (resting cells) at 30 °C. Samples were taken at the indicated times to determine PFGE profiles (A), to quantify Chr2 and Chr5 (B) and to calculate CFU and OD600 (C). For PFGE profiles, DNA plugs were incubated with proteinase K at 55 °C. Of note, in panels B and C, initial time (0) corresponds to samples before MMS treatment. Arrows in panel B shows the MMS-treated sample.
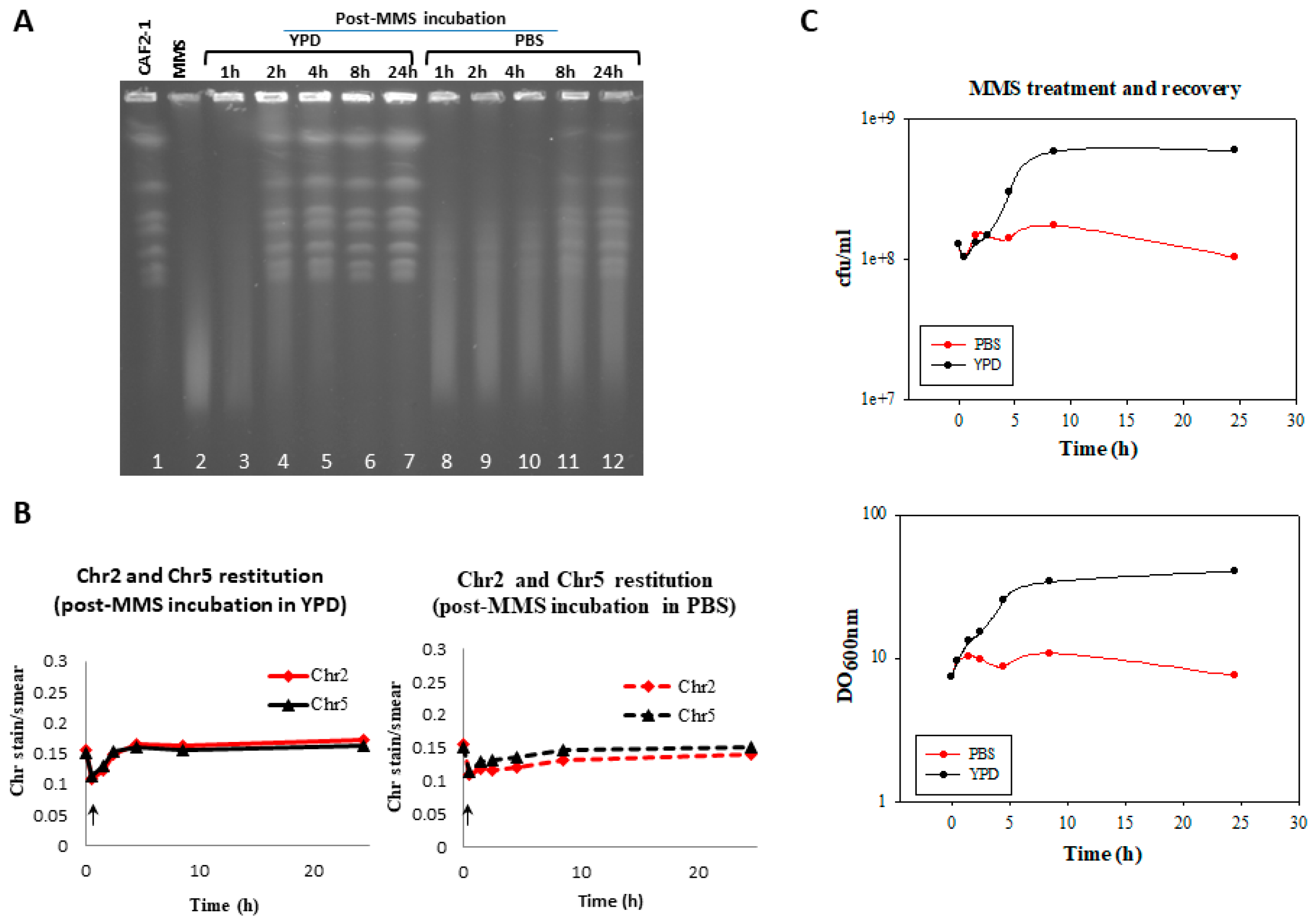
Figure 4.
Cell morphology of MMS-treated exponentially growing cells from wild type and the indicated HR mutants during recovery in MMS-free YPD medium or PBS. For experimental conditions, see legend of Figure 3.
Figure 4.
Cell morphology of MMS-treated exponentially growing cells from wild type and the indicated HR mutants during recovery in MMS-free YPD medium or PBS. For experimental conditions, see legend of Figure 3.

Figure 5.
Induction and repair of HDBs by cycling and resting wild type and HR mutants cells of C. albicans. C. albicans wild type (CAI4-URA3) (A) and the indicated HR mutants (rad59 –B-, rad51 –C- and rad52 –D-) were subjected to an acute MMS treatment (0.05% MMS for 30 min). Then cells were treated with 5% sodium thiosulphate, by mixing 1:1 (v/v) ratio with 10% Na2S2O3, washed with PBS, and resuspended in MMS-free YPD (cycling cells) or PBS (resting cells) at 30 °C. Samples were taken at the indicated times to calculate OD600, CFU (see Figure 7), and to determine PFGE profiles. For PFGE profiles, DNA plugs were incubated with proteinase K at 55 °C.
Figure 5.
Induction and repair of HDBs by cycling and resting wild type and HR mutants cells of C. albicans. C. albicans wild type (CAI4-URA3) (A) and the indicated HR mutants (rad59 –B-, rad51 –C- and rad52 –D-) were subjected to an acute MMS treatment (0.05% MMS for 30 min). Then cells were treated with 5% sodium thiosulphate, by mixing 1:1 (v/v) ratio with 10% Na2S2O3, washed with PBS, and resuspended in MMS-free YPD (cycling cells) or PBS (resting cells) at 30 °C. Samples were taken at the indicated times to calculate OD600, CFU (see Figure 7), and to determine PFGE profiles. For PFGE profiles, DNA plugs were incubated with proteinase K at 55 °C.
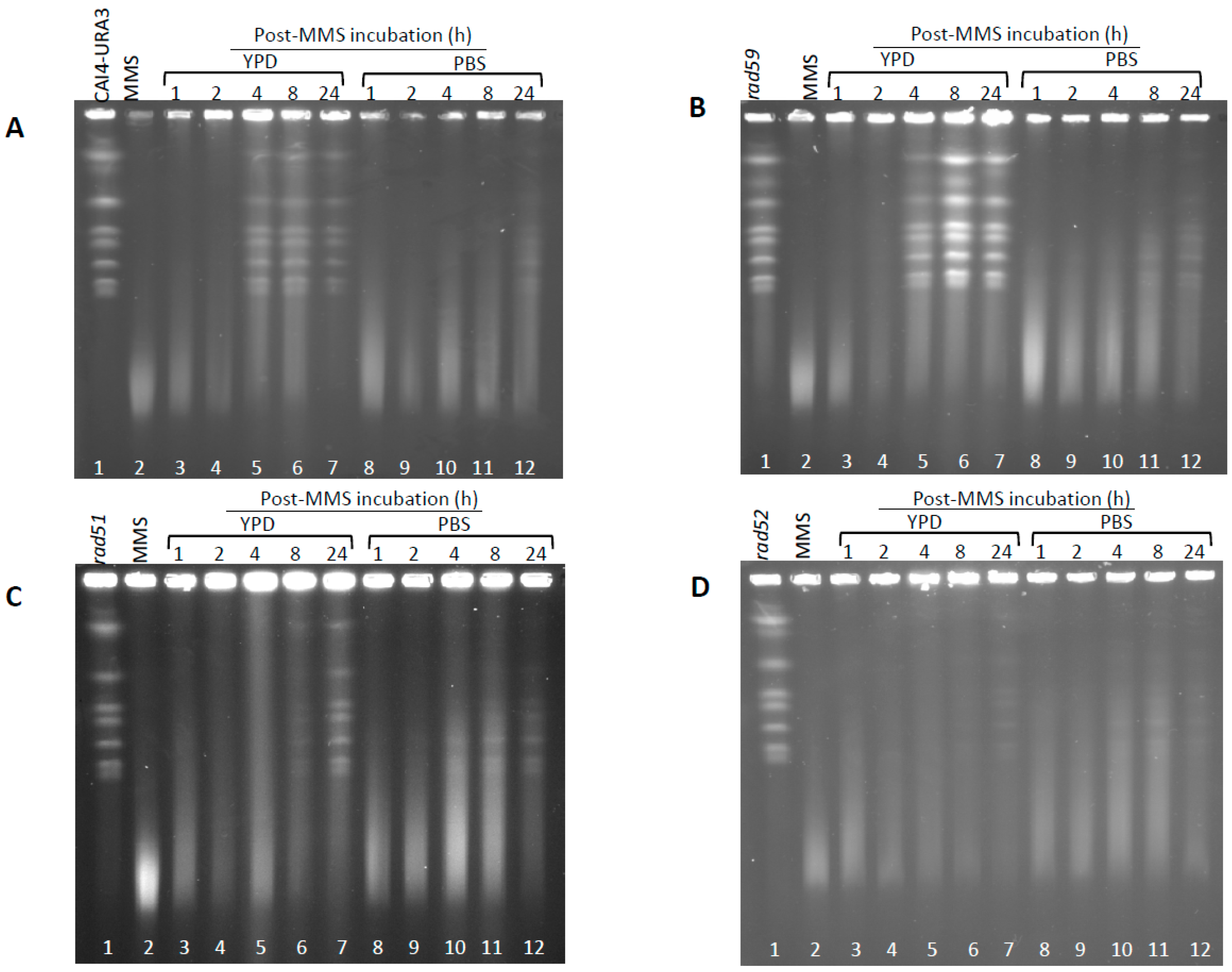
Figure 6.
Quantification of Chr2 and Chr5 intensities during the MMS-treatment and the post-MMS incubation of wild type CAI4-URA3 (A) and HR Uri+ mutants Carad59 (B), Carad51 (C) and Carad52 (D). Post-MMS incubation was carried out in YPD (cycling cells) or PBS (resting cells). PFGE gels shown in Figure 5 were quantified as described in Materials and Methods. Initial time (0) corresponds to samples before MMS treatment. Arrows within each panel shows the MMS-treated sample.
Figure 6.
Quantification of Chr2 and Chr5 intensities during the MMS-treatment and the post-MMS incubation of wild type CAI4-URA3 (A) and HR Uri+ mutants Carad59 (B), Carad51 (C) and Carad52 (D). Post-MMS incubation was carried out in YPD (cycling cells) or PBS (resting cells). PFGE gels shown in Figure 5 were quantified as described in Materials and Methods. Initial time (0) corresponds to samples before MMS treatment. Arrows within each panel shows the MMS-treated sample.
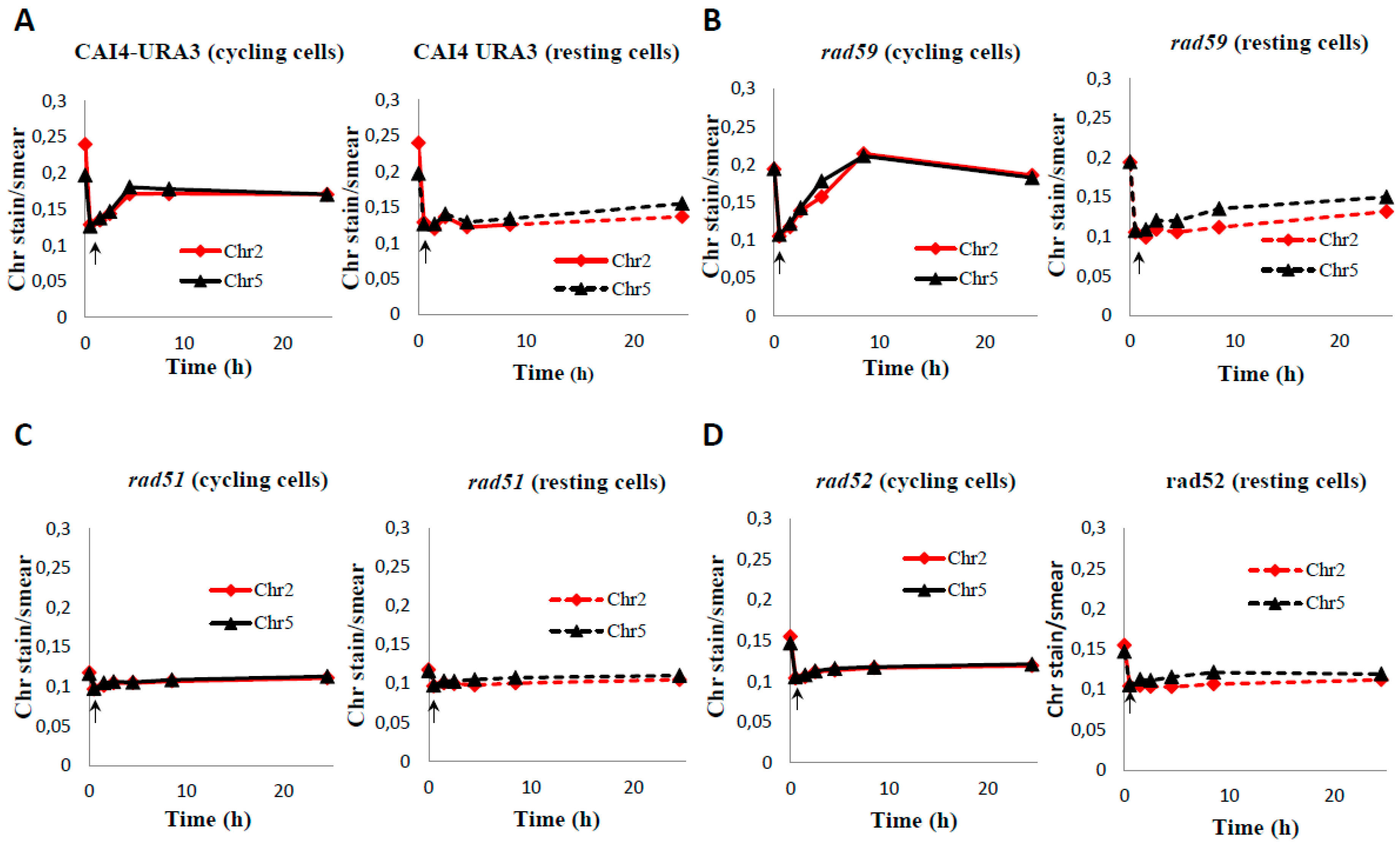
Figure 7.
Changes in CFU and OD600 from wild type and HR mutants during MMS treatment and recovery. (A) Cell survival after the acute MMS-treatment. Survivability in terms of CFU was 95% for wild type (CAI4-URA3) and Carad59-ΔΔ, 12% for Carad51-ΔΔ, and 5% for Carad52-ΔΔ. (B,C) Variation of CFUs and OD600, respectively, during recovery of MMS-treated cells in YPD. (D,E) Variation in CFU and OD600, respectively during recovery in PBS.
Figure 7.
Changes in CFU and OD600 from wild type and HR mutants during MMS treatment and recovery. (A) Cell survival after the acute MMS-treatment. Survivability in terms of CFU was 95% for wild type (CAI4-URA3) and Carad59-ΔΔ, 12% for Carad51-ΔΔ, and 5% for Carad52-ΔΔ. (B,C) Variation of CFUs and OD600, respectively, during recovery of MMS-treated cells in YPD. (D,E) Variation in CFU and OD600, respectively during recovery in PBS.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains used in this study.
| Strains (Old Name) | Genotype | Parental | Reference |
|---|---|---|---|
| Candida albicans | |||
| SC5314 | Wild type | Gillum et al., 1984 [26] | |
| CAF2-1 | Δura3::imm434/URA3 | SC5314 | Fonzi and Irwin, 1993 [27] |
| CAI4 | Δura3::imm434/Δura3::imm434 | CAF2-1 | Fonzi and Irwin, 1993 [27] |
| CAGL4 (TCR2.1) | Δura3::imm434/Δura3::imm434 rad52::hisG/Δrad52::hisG-URA3-hisG | CAI4 | Ciudad et al., 2004 [28] |
| CAGL4.1 (TCR2.1.1) | Δura3::imm434/Δura3::imm434 Δrad52::hisG/Δrad52::hisG | CAGL4 | Ciudad et al., 2004 [28] |
| CAGL17 (BNC1.1) | Δura3::imm434/Δura3::imm434 Δrad59::hisG/Δrad59::hisG-URA3-hisG | CAI4 | García-Prieto et al., 2010 [21] |
| CAGL17.1 (BNC23.1) | Δura3::imm434/Δura3::imm434 Δrad59::hisG/Δrad59::hisG | CAGL17 | Bellido et al., 2015 [22] |
| CAGL19 (JGR5) | Δura3::imm434/Δura3::imm434 Δrad51::hisG/Δrad51::hisG-URA3-hisG | CAI4 | García-Prieto et al., 2010 [21] |
| CAGL19.1 (JGR5A) | Δura3::imm434/Δura3::imm434 Δrad51::hisG/Δrad51::hisG | CAGL19 | García-Prieto et al., 2010 [21] |
| S. cerevisiae | |||
| LSY0695-7D W303 haploid | MATa; ADE2 RAD5 met17-s ade2-1 trp1-1 his3-11,15 can1-100 ura3-1 leu2-3,112 | Bärtsch et al., 2000 [24] | |
| W303 diploid | MATa/α; ADE2 RAD5 met17-s/met17-s ade2-1/ade2-1 trp1-1/trp1-1 his3-11,15/his3-11,15 can1-100/can1-100 ura3-1/ura3-1 leu2-3,112/leu2-3,112 | Bellido et al., 2015 [22] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ciudad, T.; Bellido, A.; Andaluz, E.; Hermosa, B.; Larriba, G. Role of Homologous Recombination Genes in Repair of Alkylation Base Damage by Candida albicans. Genes 2018, 9, 447. https://doi.org/10.3390/genes9090447
AMA Style
Ciudad T, Bellido A, Andaluz E, Hermosa B, Larriba G. Role of Homologous Recombination Genes in Repair of Alkylation Base Damage by Candida albicans. Genes. 2018; 9(9):447. https://doi.org/10.3390/genes9090447
Chicago/Turabian StyleCiudad, Toni, Alberto Bellido, Encarnación Andaluz, Belén Hermosa, and Germán Larriba. 2018. "Role of Homologous Recombination Genes in Repair of Alkylation Base Damage by Candida albicans" Genes 9, no. 9: 447. https://doi.org/10.3390/genes9090447
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.