
Recombinant Production of Arginyl Dipeptides by l-Amino Acid Ligase RizA Coupled with ATP Regeneration


Institut für Lebensmittelchemie, Gottfried Wilhelm Leibniz Universität Hannover, Callinstraße 5, 30167 Hannover, Germany
*
Author to whom correspondence should be addressed.
Catalysts 2021, 11(11), 1290; https://doi.org/10.3390/catal11111290
Submission received: 30 September 2021
/
Revised: 22 October 2021
/
Accepted: 25 October 2021
/
Published: 27 October 2021
(This article belongs to the Special Issue Enzyme Catalysis, Biotransformation and Bioeconomy)
Abstract
:Arginyl dipeptides like Arg-Ser, Arg-Ala, and Arg-Gly are salt-taste enhancers and can potentially be used to reduce the salt content of food. The l-amino acid ligase RizA from B. subtilis selectively synthesizes arginyl dipeptides. However, industrial application is prevented by the high cost of the cofactor adenosine triphosphate (ATP). Thus, a coupled reaction system was created consisting of RizA and acetate kinase (AckA) from E. coli providing ATP regeneration from acetyl phosphate. Both enzymes were recombinantly produced in E. coli and purified by affinity chromatography. Biocatalytic reactions were varied and analyzed by RP-HPLC with fluorescence detection. Under optimal conditions the system produced up to 5.9 g/L Arg-Ser corresponding to an ATP efficiency of 23 g Arg-Ser per gram ATP. Using similar conditions with alanine or glycine as second amino acid, 2.6 g/L Arg-Ala or 2.4 g/L Arg Gly were produced. The RizA/AckA system selectively produced substantial amounts of arginyl dipeptides while minimizing the usage of the expensive ATP.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The reduction of salt intake is one of the most prevalent public health initiatives, as an excessively high salt intake is correlated with high blood pressure and other cardiovascular diseases [1,2]. But success is limited as the recommended amount of five grams by the WHO is far exceeded by the actual salt intake in most western countries surpassing ten grams [3,4]. Reduction of salt intake can substantially lower the blood pressure of both hypertensive and healthy people [5]. However, reductions in salt-content of food are usually not well tolerated, as salt has an important function for the taste of food [6]. Apart from saltiness being one of five tastes, it also enhances the flavor by suppressing bitterness [7]. In 2011, arginyl dipeptides formed by hydrolysis of fish protein [4] were identified as salt-taste enhancers. The strongest effects were found for Arg-Pro, Arg-Ala, Arg-Gly, and Arg-Ser, which increased the perceived salt-taste intensity by 10 to 20% without any off-flavors and are thus suitable candidates to reduce the salt content of food.
Chemical synthesis of dipeptides requires the use of protecting groups to direct the specificity of the reaction which increases costs and produces additional waste [8]. In a biocatalytic approach, peptidases from basidiomycetes were used to produce protein hydrolysates enriched in salt-taste enhancing dipeptides [9]. A relatively novel enzyme class in the biocatalytic toolkit are l-amino acid ligases (LALs; EC 6.3.2.28) which condense two amino acids to their corresponding dipeptide under hydrolysis of ATP to ADP (adenosine diphosphate) [10]. The LAL RizA from Bacillus subtilis NBRC3134 has a high specificity for producing dipeptides with an N-terminal arginine and is an ideal candidate for the synthesis of salt-taste enhancing dipeptides [11]. However, the need for stoichiometric amounts of ATP presents an enormous hurdle for the industrial application due to its high price. Many strategies were developed to regenerate ATP either by using endogenous enzymes of whole cells [12,13] or isolated enzymes [14,15,16]. One very prominent strategy is the use of acetate kinase which regenerates ATP from ADP and acetyl phosphate (AcP) [17]. The necessary acetyl phosphate can be synthesized reasonably cheap by acetylation of phosphoric acid with acetic anhydride [13,18] or through biocatalytic strategies such as the synthesis from pyruvate and phosphate by pyruvate oxidase from Pediococcus sp. [19]. Recently, acetate kinase was used in a multistep biocatalytic cascade to produce the potential HIV drug Islatravir [20].
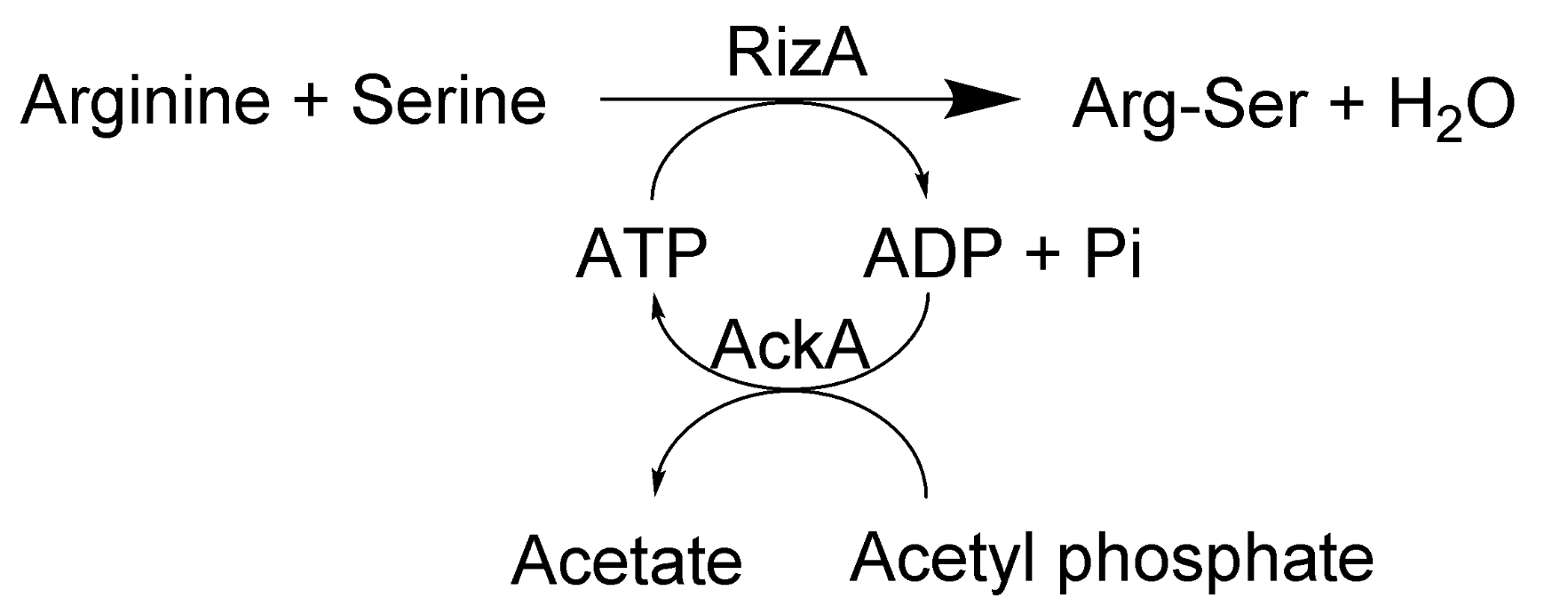
In the present work, a reaction system comprised of RizA and AckA was constructed for the production of the salt-taste enhancing dipeptides Arg-Ser, Arg-Ala, and Arg-Gly with minimal need for ATP (Figure 1).
2. Results and Discussion
2.1. Influence of Substrate Concentrations
RizA and AckA were recombinantly produced in E. coli BL21 (DE3) and purified by affinity chromatography. The yields were 14 mg RizA per 100 mL culture, and 53 mg AckA per 100 mL culture. Biocatalytic reactions were set up with the purified enzymes to optimize the reaction conditions. The production of Arg-Ser was chosen as the optimization target, since Arg-Ser is among the strongest salt-taste enhancers, and initial experiments showed that RizA had the highest activity for this dipeptide.
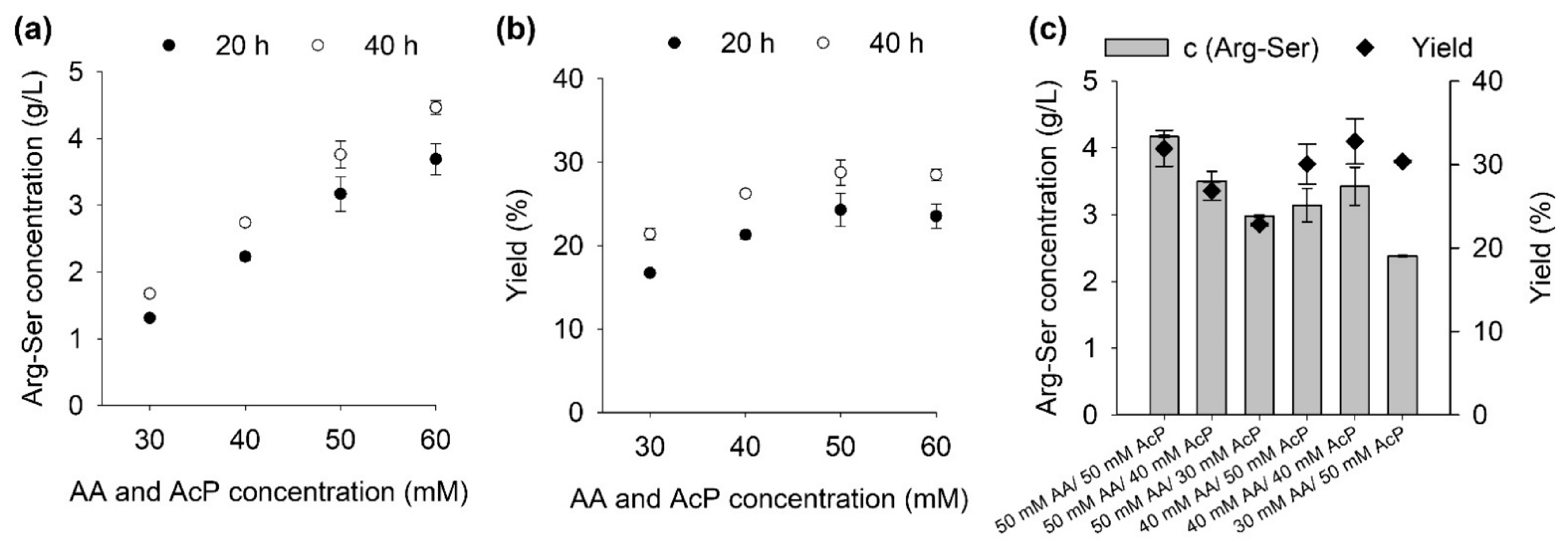
Firstly, the substrate concentrations were examined. Reactions were set up with different concentrations of arginine and serine with equimolar amounts acetyl phosphate for 20 and 40 h reaction time. (Figure 2a,b). Another set of reactions with different ratios of amino acids to acetyl phosphate was also set up (Figure 2c).
In all samples, most of the product (>80%) was already present in the 20 h samples with only a small increase from 20 to 40 h reaction time. Increasing amino acid alongside acetyl phosphate concentrations led to an approximately linear increase in product concentration up to 60 mM with no signs of substrate inhibition (Figure 2a). This is similar to the results reported for the LAL from Bacillus licheniformis, where up to 40 mM increased the reaction velocity [21] while for YwfE up to 100 mM worked [22]. The highest yields were achieved with 50 mM AA and AcP reaching 24% after 20 h and 29% after 40 h (Figure 2b). An equimolar ratio of amino acids to acetyl phosphate was found to be optimal at 50 mM with a yield of 32% (Figure 2c). For RizA without ATP regeneration, a lower yield of 25% was achieved with 12.5 mM AA and 12.5 mM ATP, which was the highest yield reported for RizA [11].
2.2. Influence of Cofactor Concentrations
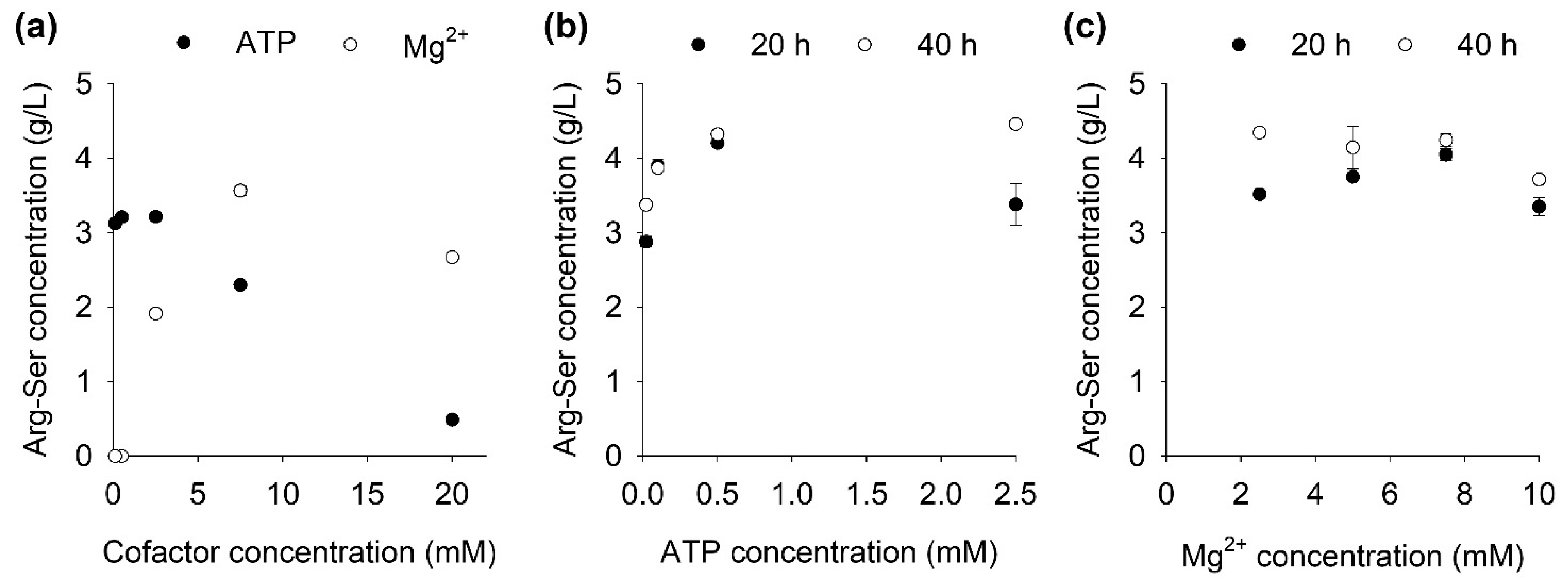
Both RizA and AckA require ATP and Mg2+ as cofactors. In the literature, LAL reactions without ATP regeneration are often set up with both cofactor concentrations equimolar to the amino acids and ATP [11,23,24]. Due to the presence of another enzyme and the objective to reduce the ATP usage, both cofactors were analyzed for their effect on the RizA/AckA system. Reactions were set up with ATP and Mg2+ with concentrations from 0.1 to 20 mM with 40 mM amino acids and 24 h reaction time (Figure 3a). Additionally, reactions were set up with 50 mM amino acids, 20 and 40 h reaction time and different concentration ranges to pinpoint the optimum concentration for both cofactors (Figure 3b,c).
The optimum concentrations for ATP were 0.5 mM (Figure 3a,b) and for Mg2+ 7.5 mM (Figure 3a,c) at 20 h reaction time. Lower or higher concentrations limited the reaction for both cofactors, although to different degrees. At 0.5 and 0.1 mM Mg2+, no product formation was detected, while reducing the ATP concentration to 0.02 mM only reduced the final product concentration by 33% (Figure 3a). Conversely, higher ATP concentrations had a strong inhibiting effect on the reaction system as displayed by the 75% reduction in product formation with 20 mM ATP. Inhibition by ATP was also witnessed during experiments without cofactor regeneration in which 30 mM ATP completely inhibited the reaction (Figure S1). Besides the need to minimize ATP due to its cost, the usage of an ATP-regenerating system also alleviated this apparent substrate inhibition imposed on the RizA enzyme by ATP.
2.3. Influence of Enzyme Concentrations
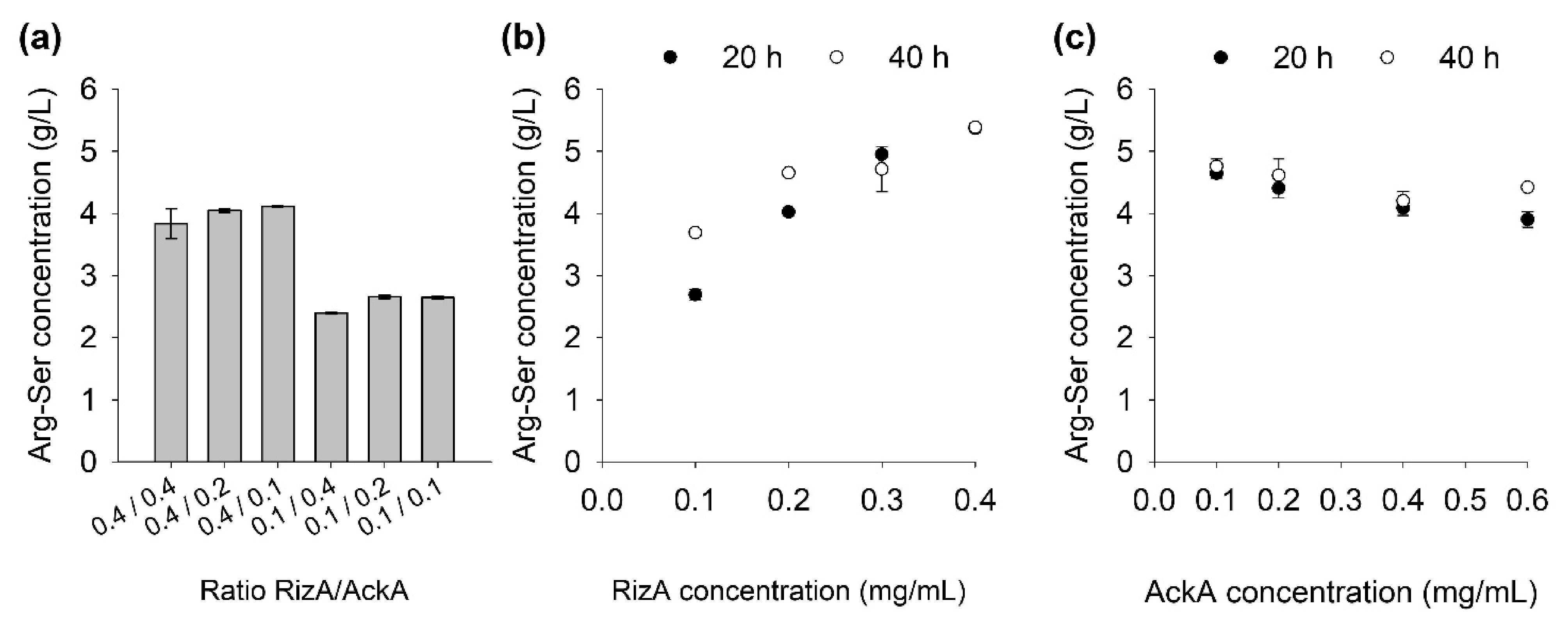
Reactions with different ratios of RizA to AckA were set up (Figure 4a). Additional reactions with varying concentrations of one enzyme were created as well (Figure 4b,c).
In the 20 h RizA samples (Figure 4b), a four-fold increase from 0.1 to 0.4 mg/mL doubled the final product concentration. The size of the product increase from the 20 to the 40 h samples was inversely correlated with the RizA concentration, as the 0.1 mg/mL sample increased from 2.7 to 3.7 g/L and the 0.4 mg/mL showed no increase. These findings imply that the enzymes in the 0.4 mg/mL reactions were still active, but that the product concentration had reached a ceiling. In the reactions with lower RizA concentrations, this ceiling had not been reached yet and the reaction progress continued. The apparent concentration ceiling is further discussed in Section 2.5.
In contrast to RizA, higher concentrations of AckA seemed to decrease the product formation, although the effect of the concentration of this enzyme was far less pronounced than with RizA. Addition of 0.1 mg/mL AckA was still sufficient for the reaction. The reactions with different ratios of RizA to AckA (Figure 4a) confirmed these results as the 0.4 mg/mL RizA and 0.1 mg/mL AckA reactions contained the highest product concentration.
2.4. Influence of Temperature and pH
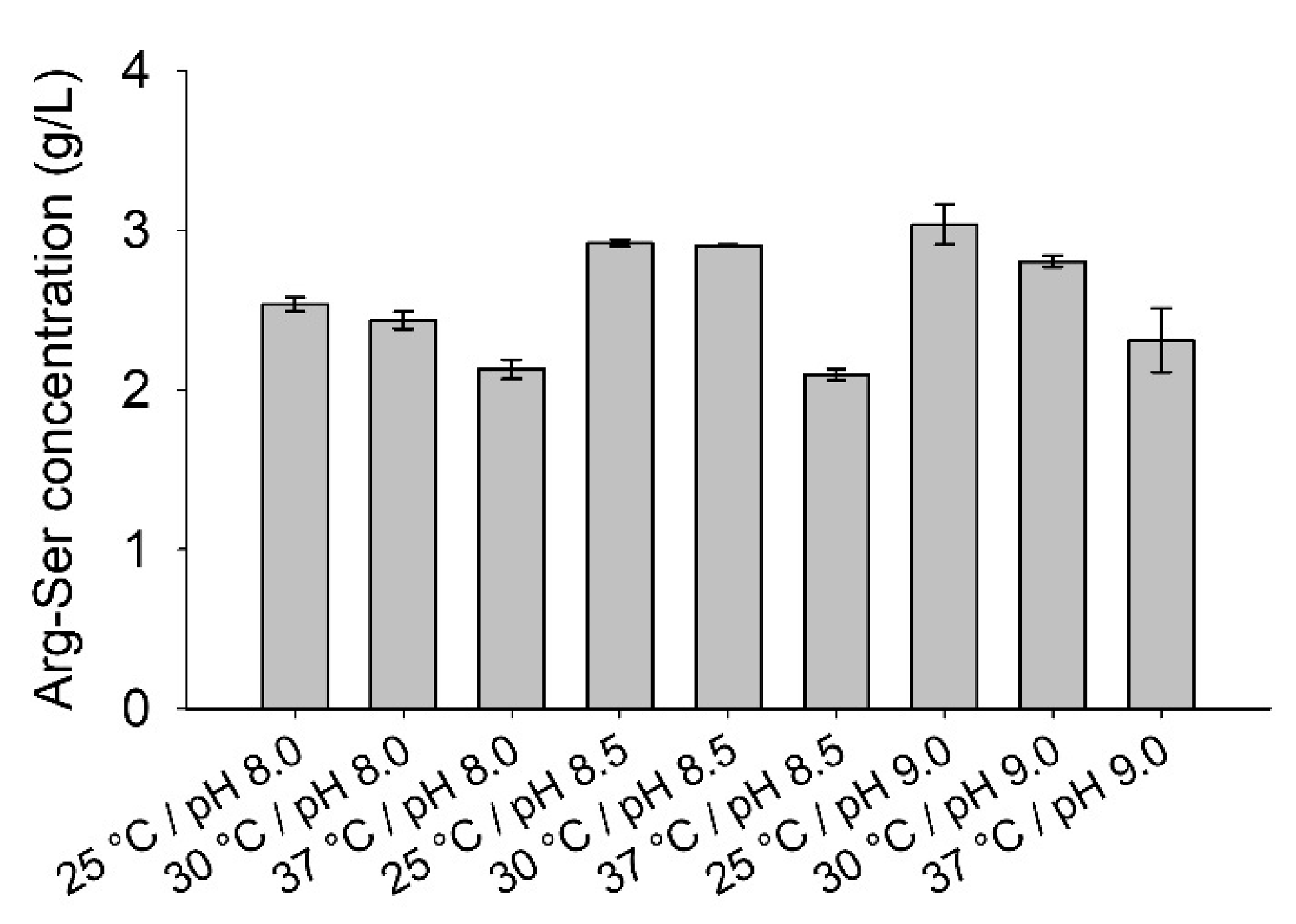
The optimum temperature and pH for isolated RizA were determined to be 37 °C and pH 9.5 with sharp declines towards lower temperatures/pH values [11]. In contrast, the pH optimum of AckA was at 7.3, but with a broader spectrum, still possessing 50% activity at pH 9.0 [25]. To analyze the optimum of the combined system, reactions were set up with combinations of the three reaction temperatures of 25, 30, and 37 °C and three buffers with pH values of 8.0, 8.5, and 9.0 (Figure 5).
At all pH values, 37 °C reaction temperatures led to the lowest product concentrations while the 30 and 25 °C showed similar results. At constant temperature, the reactions with pH 8.0 contained the lowest product concentrations while pH 8.5 and 9.0 performed similarly. The reaction at 25 °C and pH 9.0 contained the highest product concentration, but the difference was not significantly different from the results at 30 °C or pH 8.5. Overall, the system possessed a broader spectrum with temperatures between 25 and 30 °C and pH 8.5 to pH 9.0 being viable.
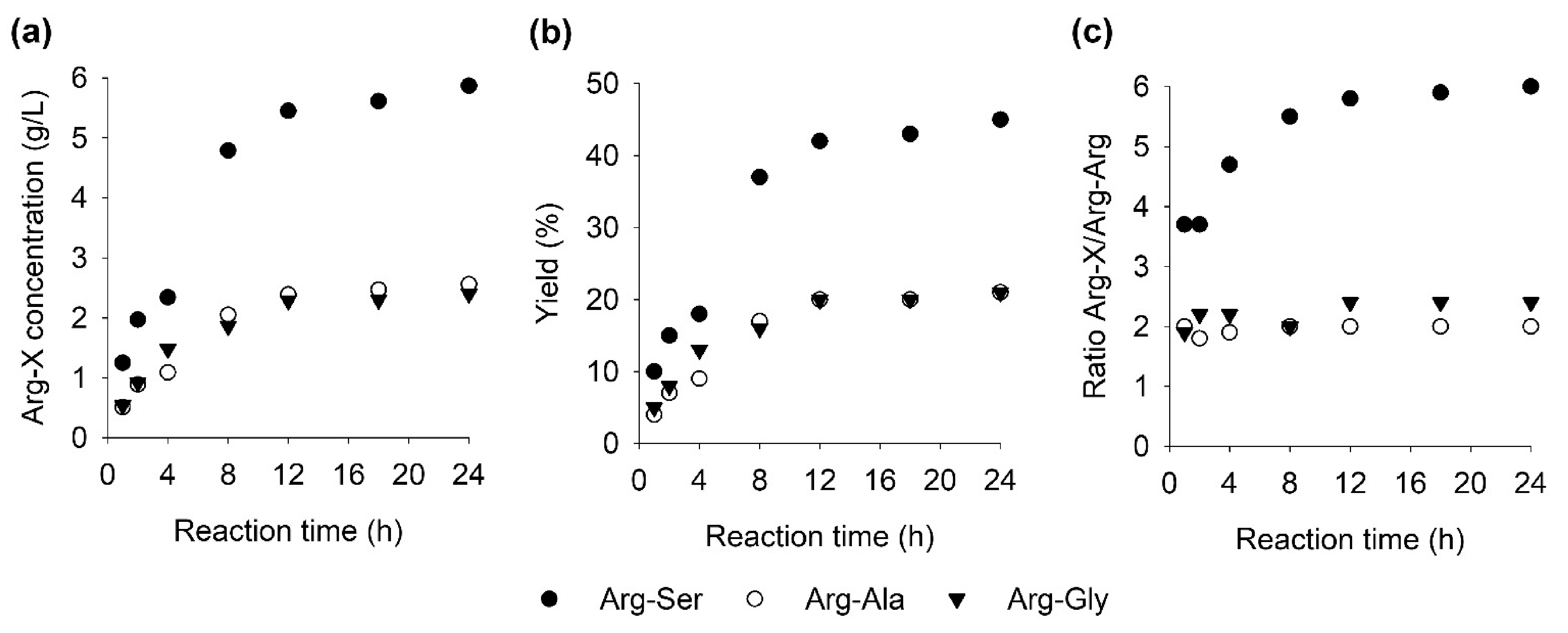
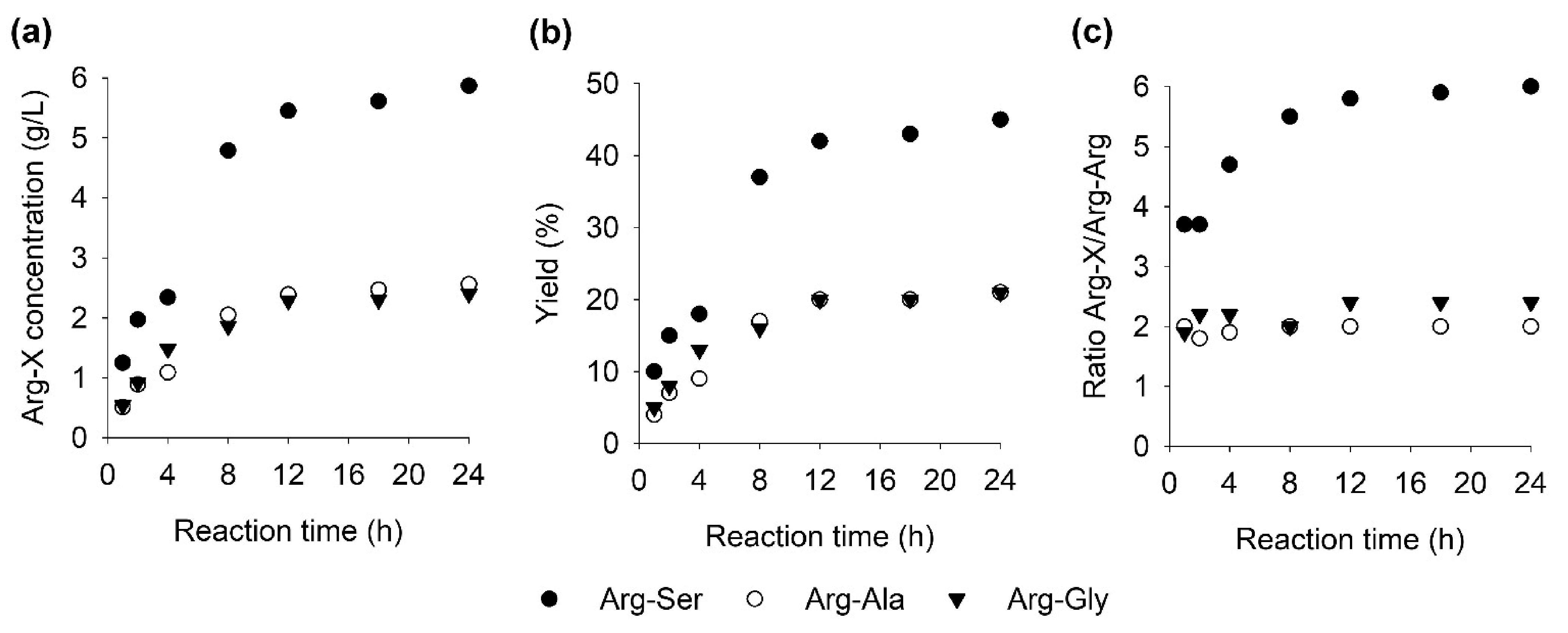
2.5. Time Course of Formation of Arg-Ser, Arg-Ala, Arg-Gly, and Arg-Arg
Reactions were set up with the best observed conditions from the above experiments to optimize the product formation. In addition to arginine and serine, reactions with arginine and either alanine or glycine were also included for the formation of the strong salt-taste enhancers Arg-Ala and Arg-Gly [11], respectively (Figure 6a,b). Apart from these heterodipeptides, formation of the homodipeptide Arg-Arg was also detected as a side product in the reaction. To quantitate the specificity between main product and side product, the ratio Arg-X/Arg-Arg was calculated (Figure 6c).
In general, Arg-Ala and Arg-Gly were formed at similar rates with final product concentrations of 2.4 and 2.6 g/L corresponding to yields of 21% each. Formation of Arg-Ser was significantly higher with a final concentration of 5.9 g/L and a yield of 41%. Similarly, the Arg-Ser reactions contained a higher ratio of Arg-Ser to Arg-Arg over five, while the reactions of Arg-Ala and Arg-Gly had ratios around two. In all cases, over 90% of the product was formed during the first 12 h with only small increases in product concentration after that. The ratio Arg-X/Arg-Arg increased for all reactions with reaction time.
In the original publication on the discovery and recombinant production of RizA, Arg-Arg was only detected in reactions containing only arginine but not in the arginine and serine reaction [11]. It is possible that Arg-Arg was formed but not detected due to the lower substrate concentrations used. Another explanation might be an effect of the his-tag on the reactivity on the enzyme. An N-terminal his6-tag was used in both cases, but the exact placement/linker sequence between tag and RizA could be different and have an effect, although unlikely. Since Arg-Arg is a weak salt-taste enhancer, it does not negatively affect the intended application. To reduce or prevent the formation of Arg-Arg, the substrate specificity of RizA could be engineered by mutagenesis. This was achieved for the production of Met-Gly with the LAL from Bacillus licheniformis, in which the production of the side product Met-Met was successfully prevented by a single mutation of Pro85 [21]. Another target for engineering is the yield of the reaction. As shown, the production of dipeptides slowed considerably after 12 h reaching final yields of around 21% for Arg-Ala and Arg-Gly and 41% for Arg-Ser. Possible reasons for this could be inhibition by phosphate [26] or acetate [18] accumulating during the reaction (Figure 1). In the case of the cascade for the production of Islatravir using acetate kinase, the accumulation of phosphate was reduced by the addition of sucrose and sucrose phosphorylase [17,20]. A ceiling to the final product concentration could also be imposed by the RizA enzyme itself and thus be addressed by protein engineering. This was done for the LAL TabS in which the double mutant S85T/H294D reached a 1.6-fold higher Pro-Gly concentration after 20 h than the wild type [27].
The ATP regeneration in this work increased both the efficiency of ATP usage and enabled the highest reported product concentrations for Arg-Ser by a biocatalytic process. For RizA without ATP regeneration, a product concentration of 3.1 mM or 0.8 g/L Arg-Ser was reported with 12.5 mM of amino acids, Mg2+ and ATP [11]. In terms of ATP efficiency, this corresponds to 0.0064 Arg-Ser per mmole of ATP or 0.13 g Arg-Ser per gram ATP. The optimized system of RizA/AckA used only 0.5 mM ATP to produce 5.9 g/L or 23 g Arg-Ser per gram of ATP corresponding to a 176-fold increase in efficiency. The combination of RizA with AckA presents a large step towards an integrated biocatalytic process [28] for the production of salt-taste enhancers and possibly other bioactive dipeptides like arginyl-phenylalanine, which was found to have antihypertensive properties [29]. As a result, the present work may not only pave the way to a better nutritional value of food, but open access to dipeptides with desired functions in human physiology. It improved the economic viability of RizA by dramatically reducing ATP as a cost factor. On the road towards an industrial process, the next step will be immobilization of the reaction system to increase its stability and reusability for a larger-scale bioprocess [30].
3. Materials and Methods
3.1. Chemicals, Reagents and Strains
All chemicals were purchased from Carl Roth (Karlsruhe, Germany) or Sigma Aldrich (Taufkirchen, Germany) if not otherwise indicated. Enzymes for molecular biology were purchased from Thermo Fisher Scientific (St. Leon-Roth, Germany). The pET28a vector was purchased from Merck KGaA (Darmstadt, Germany). The E. coli strains BL21 (DE3) and TOP10 were maintained in our laboratory. Oligonucleotides were synthesized by Microsynth Seqlab GmbH (Goettingen, Germany).
3.2. Construction of pET28a_his6-rizA and pET28a_his6-ackA Constructs
The sequence of rizA (UniProt accession B5UAT8) was codon-optimized with the software SnapGene version 5.1.7 (2020) from GSL Biotech LLC (Chicago, IL, USA) and an N-terminal 6his-tag sequence was added. The his6-rizA (Sequence S1) gene was produced as a synthetic gene by Thermo Fisher Scientific (St. Leon-Roth, Germany) with restriction sites for NcoI and NotI. The delivered pMA-T-his6-rizA vector and the pET28a vector were digested with FastDigest NcoI and NotI followed by ligation of the his6-rizA fragment into the pET28a vector with T4 DNA ligase. The product was transformed into chemocompetent E. coli TOP10 cells by heat-shock transformation. After overnight incubation at 37 °C, clones were selected for overnight incubation, plasmids were isolated with the innuPREP Plasmid Mini Kit 2.0 from Analytik Jena (Jena, Germany) and the correct sequence confirmed by sequencing by Microsynth Seqlab GmbH (Goettingen, Germany). For the pET28a_ackA construct, genomic DNA of E. coli was isolated by the method of El-Ashram, et al. [31] and the ackA gene was amplified with Phusion High Fidelity Polymerase using the primers “CATCATATGATGTCGAGTAAGTTAGTACTGGTTCTG” and “GTAGCGGCCGCTCAGGCAGTCAGGCGGC” containing NdeI and NotI restriction sites. The amplificate was inserted into the pET28a vector analogously to rizA above with the respective digestion enzymes resulting in an open reading frame coding for his6-ackA (Sequence S2). Both constructs were transformed into E. coli BL21 (DE3) for expression and glycerol stocks were created for storage at −80 °C.
3.3. Cultivation and Expression
Precultures were inoculated from glycerol stocks in 5 mL LB medium with 50 µg/mL kanamycin and incubated overnight at 37 °C, 200 rpm. For cultivation, 400 mL TB medium with 50 µg/mL kanamycin in a shaking flask with baffles were inoculated with 4 mL preculture and incubated at 37 °C and 160 rpm until an OD600 of 0.6–0.8 was reached. Expression was induced by addition of 10 µM IPTG and performed at 20 °C and 160 rpm for 20 h. After harvesting the cells by centrifugation at 5000× g at 4 °C for 15 min, the cell pellets were frozen and stored at −20 °C until purification.
3.4. Purification
A cell pellet from 100 mL culture was resuspended in 10 mL disruption buffer (100 mM tricine pH 8.0, 10 mM imidazole) and sonicated for 7.5 min (50% amplitude; 0.5 cycle) with a UP50H sonicator from Hielscher Ultrasonics GmbH (Teltow, Germany). After centrifugation at 5000× g at 4 °C for 40 min, the supernatant was purified by affinity chromatography with a gravity-flow column containing 2 mL Protino Ni-NTA Agarose (Macherey-Nagel, Dueren, Germany) according to the manufacturer’s instructions. The eluted protein was desalted by PD10 columns from Cytiva (Washington, DC, USA) according to the manufacturer’s instructions. The desalted eluate was frozen in liquid nitrogen and aliquots were stored at −80 °C. Protein concentrations were determined with Bradford solution from Sigma–Aldrich (Taufkirchen, Germany).
3.5. Biocatalysis
Biocatalytic reactions were set up in 0.2 mL PCR tubes with 50 µL reaction volume. If not otherwise stated, reactions contained 40 mM arginine and serine, 40 mM AcP, 0.5 mM ATP, 7.5 mM MgSO4, 0.2 mg/mL RizA, 0.2 mg/mL AckA and 25 mM tricine buffer pH 8.5. Reactions were set up in duplicates and incubated at 25 °C for 20 h in a Biometra thermal cycler from Analytik Jena (Jena, Germany) and inactivated by heating to 70 °C. Samples were stored at −20 °C until analysis.
3.6. Analysis
The amino groups of dipeptides were derivatized with o-phthalaldehyde and analyzed by RP-HPLC with fluorescence detection as previously described by Rottmann, et al. [32] for the analysis of single amino acids. The gradient was modified as follows: 0 min: 90% A, 5 min: 90% A, 25 min: 60% A, 30 min: 0% A, 35 min: 0% A, 40 min: 90% A, 45 min: 90% A. Analytical standards of the dipeptides Arg-Ser, Arg-Ala, Arg-Gly and Arg-Arg were purchased from Bachem (Bubendorf BL, Switzerland) and used for external calibration. Standards were set up in duplicate. Data were visualized with SigmaPlot version 14.5 (2020) from Systat Software GmbH (Erkrath, Germany).
4. Conclusions
ATP regeneration by acetate kinase increased both the yield and economic viability of dipeptide synthesis by RizA. The coupled reaction system provides a foundation for the development of an industrial process for the specific production of salt-taste enhancing arginyl dipeptides.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/catal11111290/s1, Figure S1: Comparison of reaction with or without regeneration; Sequence S1: DNA sequence of his6-rizA; Sequence S2: DNA sequence of his6-ackA
Author Contributions
Conceptualization, S.B. and R.G.B.; methodology, S.B. and T.A.M.; validation, S.B.; formal analysis, S.B.; investigation, S.B. and T.A.M.; writing—original draft preparation, S.B.; writing—review and editing, R.G.B. and F.E.; visualization, S.B.; supervision, R.G.B. and F.E.; project administration, R.G.B. and F.E.; funding acquisition, R.G.B. All authors have read and agreed to the published version of the manuscript.
Funding
The project was supported by funds of the Federal Ministry of Food and Agriculture (BMEL) based on a decision of the Parliament of the Federal Republic of Germany via the Federal Office for Agriculture and Food (BLE) under the innovation support programme. The publication of this article was funded by the Open Access Fund of Leibniz Universität Hannover.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- He, F.J.; MacGregor, G.A. A comprehensive review on salt and health and current experience of worldwide salt reduction programmes. J. Hum. Hypertens. 2009, 23, 363–384. [Google Scholar] [CrossRef] [PubMed]
- Trieu, K.; Neal, B.; Hawkes, C.; Dunford, E.; Campbell, N.; Rodriguez-Fernandez, R.; Legetic, B.; McLaren, L.; Barberio, A.; Webster, J. Salt Reduction Initiatives around the World–A Systematic Review of Progress towards the Global Target. PLoS ONE 2015, 10, e0130247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strazzullo, P.; D’Elia, L.; Kandala, N.-B.; Cappuccio, F.P. Salt intake, stroke, and cardiovascular disease: Meta-analysis of prospective studies. BMJ 2009, 339, b4567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Guideline: Sodium Intake for Adults and Children. Available online: https://www.who.int/publications/i/item/9789241504836 (accessed on 29 September 2021).
- He, F.J.; MacGregor, G.A. Effect of modest salt reduction on blood pressure: A meta-analysis of randomized trials. Implications for public health. J. Hum. Hypertens. 2002, 16, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Desmond, E. Reducing salt: A challenge for the meat industry. Meat Sci. 2006, 74, 188–196. [Google Scholar] [CrossRef]
- Breslin, P.A.S.; Beauchamp, G.K. Salt enhances flavour by suppressing bitterness. Nature 1997, 387, 563. [Google Scholar] [CrossRef]
- Yagasaki, M.; Hashimoto, S.-I. Synthesis and application of dipeptides; current status and perspectives. Appl. Microbiol. Biotechnol. 2008, 81, 13–22. [Google Scholar] [CrossRef]
- Harth, L.; Krah, U.; Linke, D.; Dunkel, A.; Hofmann, T.; Berger, R.G. Salt Taste Enhancing L-Arginyl Dipeptides from Casein and Lysozyme Released by Peptidases of Basidiomycota. J. Agric. Food Chem. 2018, 66, 2344–2353. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, Y.-F.; Ning, L.-X.; Wang, Y.-F.; Liu, X.-H.; Li, R.; Chen, X.-E. L-amino acid ligase: A promising alternative for the biosynthesis of L-dipeptides. Enzyme Microb. Technol. 2020, 136, 109537. [Google Scholar] [CrossRef]
- Kino, K.; Kotanaka, Y.; Arai, T.; Yagasaki, M. A Novel L-Amino Acid Ligase from Bacillus subtilis NBRC3134, a Microorganism Producing Peptide-Antibiotic Rhizocticin. Biosci. Biotechnol. Biochem. 2009, 73, 901–907. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Yin, Z. Highly Efficient Synthesis of Glutathione via a Genetic Engineering Enzymatic Method Coupled with Yeast ATP Generation. Catalysts 2020, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Alissandratos, A.; Caron, K.; Loan, T.D.; Hennessy, J.E.; Easton, C.J. ATP Recycling with Cell Lysate for Enzyme-Catalyzed Chemical Synthesis, Protein Expression and PCR. ACS Chem. Biol. 2016, 11, 3289–3293. [Google Scholar] [CrossRef]
- Andexer, J.N.; Richter, M. Emerging Enzymes for ATP Regeneration in Biocatalytic Processes. ChemBioChem 2015, 16, 380–386. [Google Scholar] [CrossRef]
- Li, Z.; Ning, X.; Zhao, Y.; Zhang, X.; Xiao, C.; Li, Z. Efficient One-Pot Synthesis of Cytidine 5′-Monophosphate Using an Extremophilic Enzyme Cascade System. J. Agric. Food Chem. 2020, 68, 9188–9194. [Google Scholar] [CrossRef]
- Yan, B.; Ding, Q.; Ou, L.; Zou, Z. Production of glucose-6-phosphate by glucokinase coupled with an ATP regeneration system. World J. Microbiol. Biotechnol. 2014, 30, 1123–1128. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Y.-H.P.J. Enzymatic regeneration and conservation of ATP: Challenges and opportunities. Crit. Rev. Biotechnol. 2021, 41, 16–33. [Google Scholar] [CrossRef]
- Crans, D.C.; Whitesides, G.M. A convenient synthesis of disodium acetyl phosphate for use in in situ ATP cofactor regeneration. J. Org. Chem. 1983, 48, 3130–3132. [Google Scholar] [CrossRef]
- Kim, D.-M.; Swartz, J.R. Prolonging cell-free protein synthesis with a novel ATP regeneration system. Biotechnol. Bioeng. 1999, 66, 180–188. [Google Scholar] [CrossRef]
- Huffman, M.A.; Fryszkowska, A.; Alvizo, O.; Borra-Garske, M.; Campos, K.R.; Canada, K.A.; Devine, P.N.; Duan, D.; Forstater, J.H.; Grosser, S.T.; et al. Design of an in vitro biocatalytic cascade for the manufacture of islatravir. Science 2019, 366, 1255–1259. [Google Scholar] [CrossRef]
- Kino, H.; Kino, K. Alteration of the substrate specificity of L-amino acid ligase and selective synthesis of Met-Gly as a salt taste enhancer. Biosci. Biotechnol. Biochem. 2015, 79, 1827–1832. [Google Scholar] [CrossRef]
- Tsuda, T.; Asami, M.; Koguchi, Y.; Kojima, S. Single Mutation Alters the Substrate Specificity of L-Amino Acid Ligase. Biochemistry 2014, 53, 2650–2660. [Google Scholar] [CrossRef]
- Kino, K.; Nakazawa, Y.; Yagasaki, M. Dipeptide synthesis by L-amino acid ligase from Ralstonia solanacearum. Biochem. Biophys. Res. Commun. 2008, 371, 536–540. [Google Scholar] [CrossRef]
- Kino, K.; Noguchi, A.; Nakazawa, Y.; Yagasaki, M. A novel L-amino acid ligase from Bacillus Licheniformis. J. Biosci. Bioeng. 2008, 106, 313–315. [Google Scholar] [CrossRef]
- Nakajima, H.; Suzuki, K.; Imahori, K. Purification and Properties of Acetate Kinase from Bacillus stearothermophilus. J. Biochem. 1978, 84, 193–203. [Google Scholar] [CrossRef]
- Dean, R.L. Kinetic studies with alkaline phosphatase in the presence and absence of inhibitors and divalent cations. Biochem. Mol. Biol. Educ. 2002, 30, 401–407. [Google Scholar] [CrossRef]
- Kino, H.; Nakajima, S.; Arai, T.; Kino, K. Effective production of Pro–Gly by mutagenesis of l-amino acid ligase. J. Biosci. Bioeng. 2016, 122, 155–159. [Google Scholar] [CrossRef]
- Burgener, S.; Luo, S.; McLean, R.; Miller, T.E.; Erb, T.J. A roadmap towards integrated catalytic systems of the future. Nat. Cat. 2020, 3, 186–192. [Google Scholar] [CrossRef]
- Kagebayashi, T.; Kontani, N.; Yamada, Y.; Mizushige, T.; Arai, T.; Kino, K.; Ohinata, K. Novel CCK-dependent vasorelaxing dipeptide, Arg-Phe, decreases blood pressure and food intake in rodents. Mol. Nutr. Food Res. 2012, 56, 1456–1463. [Google Scholar] [CrossRef]
- Yushkova, E.D.; Nazarova, E.A.; Matyuhina, A.V.; Noskova, A.O.; Shavronskaya, D.O.; Vinogradov, V.V.; Skvortsova, N.N.; Krivoshapkina, E.F. Application of Immobilized Enzymes in Food Industry. J. Agric. Food Chem. 2019, 67, 11553–11567. [Google Scholar] [CrossRef]
- El-Ashram, S.; Al Nasr, I.; Suo, X. Nucleic acid protocols: Extraction and optimization. Biotechnol. Rep. 2016, 12, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Rottmann, E.; Hauke, K.F.; Krings, U.; Berger, R.G. Enzymatic acrylamide mitigation in French fries–An industrial-scale case study. Food Control 2021, 123, 107739. [Google Scholar] [CrossRef]
Figure 1.
Reaction scheme for the RizA/AckA system for regeneration of ATP from acetyl phosphate for synthesis of Arg-Ser from argine and serine.
Figure 1.
Reaction scheme for the RizA/AckA system for regeneration of ATP from acetyl phosphate for synthesis of Arg-Ser from argine and serine.
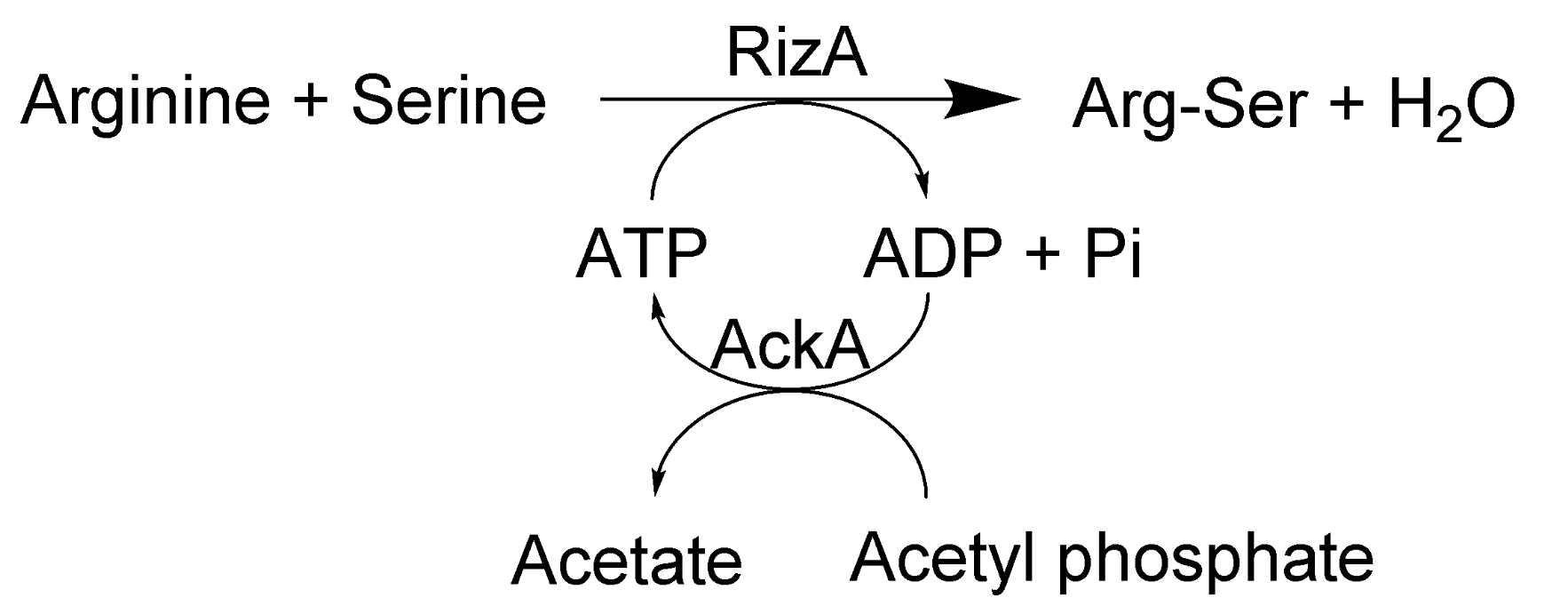
Figure 2.
Effect of substrate amino acids (AA) and acetyl phosphate on the Arg-Ser production. All reactions were performed at 25 °C. (a) Equimolar concentrations of amino acids and acetyl phosphate with 5 mM Mg2+. (b) Yields for data presented in (a). (c) Different ratios of amino acids to acetyl phosphate with 7.5 mM Mg2+ and 20 h reaction time. Reactions were set up in duplicate; data are presented as mean with error bars representing range.
Figure 2.
Effect of substrate amino acids (AA) and acetyl phosphate on the Arg-Ser production. All reactions were performed at 25 °C. (a) Equimolar concentrations of amino acids and acetyl phosphate with 5 mM Mg2+. (b) Yields for data presented in (a). (c) Different ratios of amino acids to acetyl phosphate with 7.5 mM Mg2+ and 20 h reaction time. Reactions were set up in duplicate; data are presented as mean with error bars representing range.
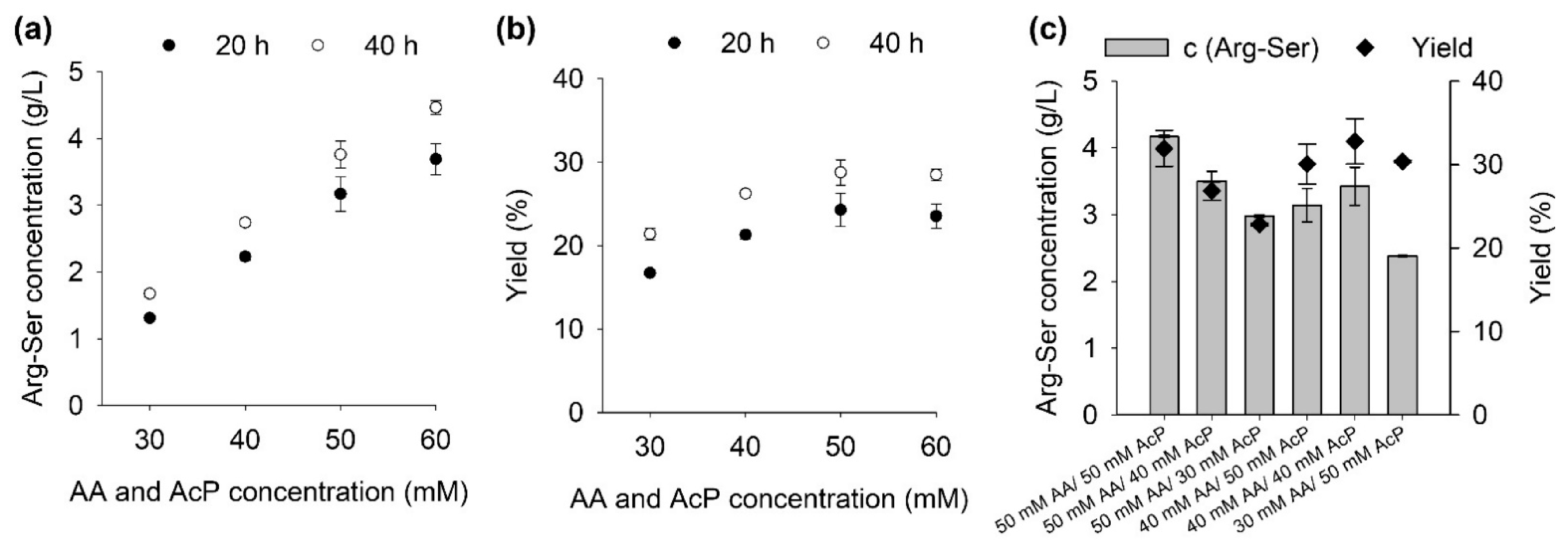
Figure 3.
Effect of two cofactors ATP and Mg2+ on Arg-Ser production. (a) ATP and Mg2+ with 40 mM AA and AcP. (b) ATP with 50 mM AA and AcP at 20/40 h reaction time. (c) Mg2+ with 50 mM AA and AcP at 20/40 h reaction time. Reactions were set up in duplicate; data are presented as mean with error bars representing the range.
Figure 3.
Effect of two cofactors ATP and Mg2+ on Arg-Ser production. (a) ATP and Mg2+ with 40 mM AA and AcP. (b) ATP with 50 mM AA and AcP at 20/40 h reaction time. (c) Mg2+ with 50 mM AA and AcP at 20/40 h reaction time. Reactions were set up in duplicate; data are presented as mean with error bars representing the range.
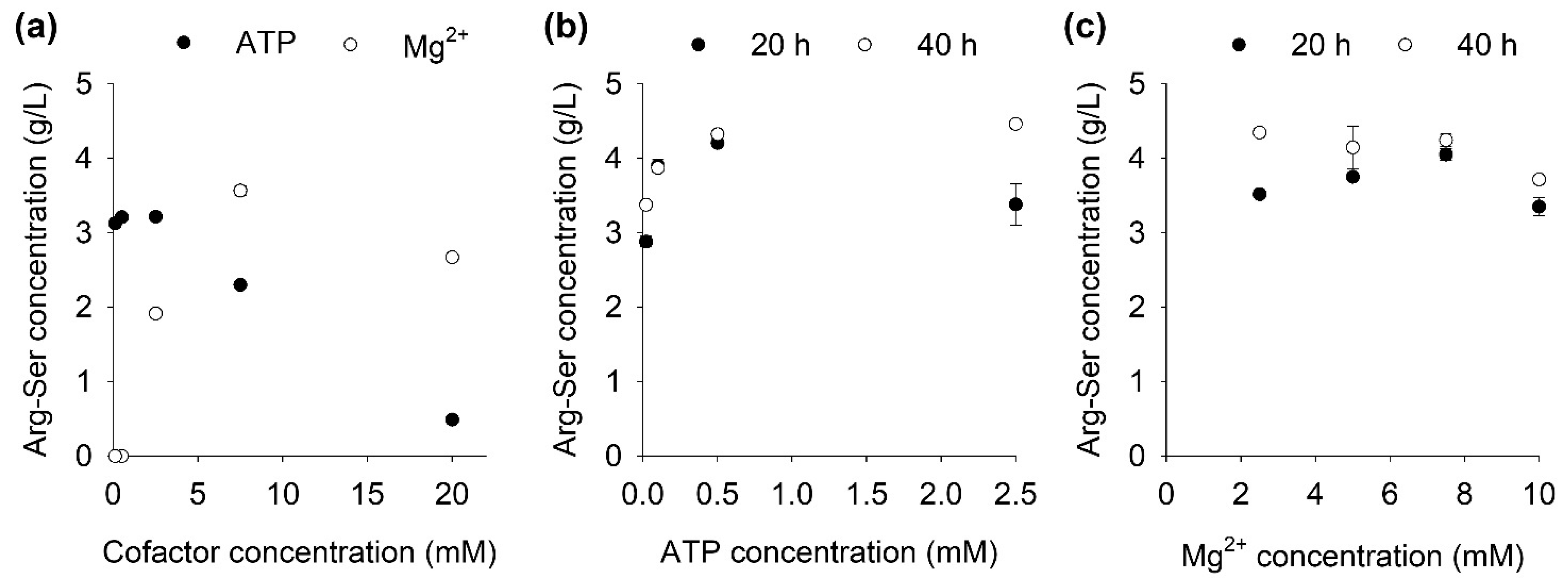
Figure 4.
Effect of two enzymes RizA and AckA on Arg-Ser production. (a) Ratios of RizA to AckA (both in mg/mL) with 40 mM AA and AcP. (b) RizA with 0.2 mg/mL AckA, 50 mM AA and AcP at 20/40 h reaction time. (c) AckA with 0.2 mg/mL RizA, 50 mM AA and AcP at 20/40 h reaction time. Reactions were set up in duplicate; data are presented as mean with error bars representing range.
Figure 4.
Effect of two enzymes RizA and AckA on Arg-Ser production. (a) Ratios of RizA to AckA (both in mg/mL) with 40 mM AA and AcP. (b) RizA with 0.2 mg/mL AckA, 50 mM AA and AcP at 20/40 h reaction time. (c) AckA with 0.2 mg/mL RizA, 50 mM AA and AcP at 20/40 h reaction time. Reactions were set up in duplicate; data are presented as mean with error bars representing range.
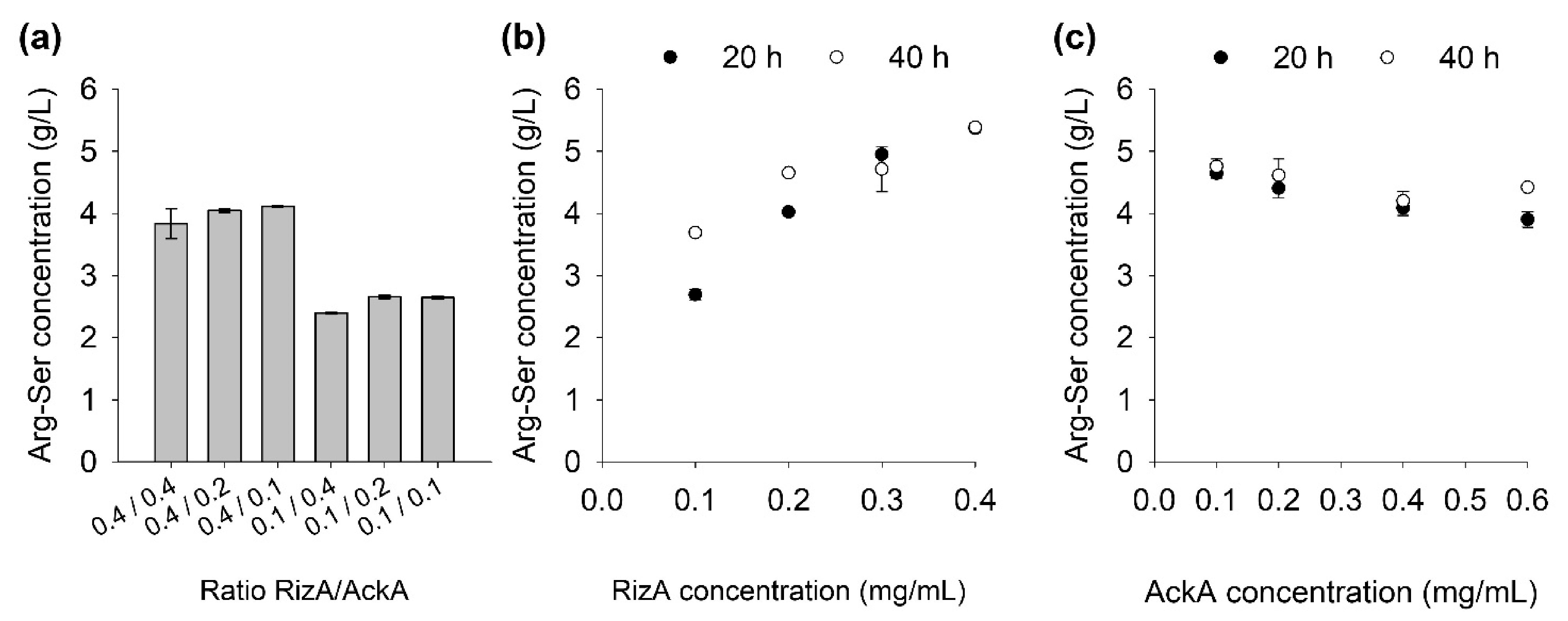
Figure 5.
Effect of temperature and buffer pH on Arg-Ser production. Reactions were set up in duplicate; data are presented as mean with error bars representing range.
Figure 5.
Effect of temperature and buffer pH on Arg-Ser production. Reactions were set up in duplicate; data are presented as mean with error bars representing range.
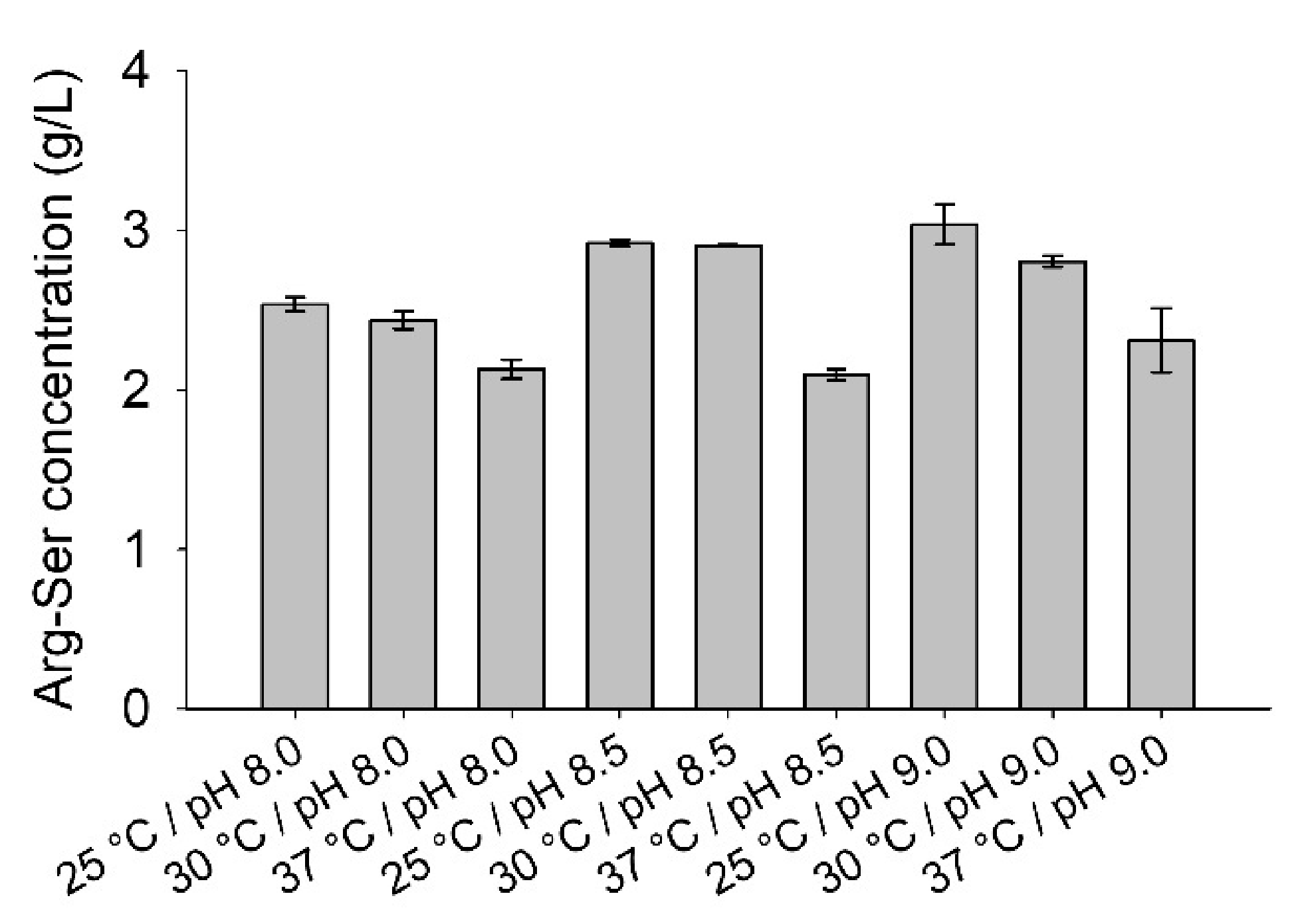
Figure 6.
Time course of dipeptide formation with optimized reaction conditions: 50 mM amino acids and AcP; 0.5 mM ATP; 7.5 mM Mg2+; 0.4 mg/mL RizA; 0.1 mg/mL AckA; 25 °C; pH 9.0 (a) Product concentrations of Arg-Ser, Arg-Ala and Arg-Gly. (b) Yields of respective Arg-X dipeptides. (c) Ratio of main product Arg-X to side product Arg-Arg.
Figure 6.
Time course of dipeptide formation with optimized reaction conditions: 50 mM amino acids and AcP; 0.5 mM ATP; 7.5 mM Mg2+; 0.4 mg/mL RizA; 0.1 mg/mL AckA; 25 °C; pH 9.0 (a) Product concentrations of Arg-Ser, Arg-Ala and Arg-Gly. (b) Yields of respective Arg-X dipeptides. (c) Ratio of main product Arg-X to side product Arg-Arg.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bordewick, S.; Mast, T.A.; Berger, R.G.; Ersoy, F. Recombinant Production of Arginyl Dipeptides by l-Amino Acid Ligase RizA Coupled with ATP Regeneration. Catalysts 2021, 11, 1290. https://doi.org/10.3390/catal11111290
AMA Style
Bordewick S, Mast TA, Berger RG, Ersoy F. Recombinant Production of Arginyl Dipeptides by l-Amino Acid Ligase RizA Coupled with ATP Regeneration. Catalysts. 2021; 11(11):1290. https://doi.org/10.3390/catal11111290
Chicago/Turabian StyleBordewick, Sven, Tim A. Mast, Ralf G. Berger, and Franziska Ersoy. 2021. "Recombinant Production of Arginyl Dipeptides by l-Amino Acid Ligase RizA Coupled with ATP Regeneration" Catalysts 11, no. 11: 1290. https://doi.org/10.3390/catal11111290
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.