SNP-SNP Interaction in Genes Encoding PD-1/PD-L1 Axis as a Potential Risk Factor for Clear Cell Renal Cell Carcinoma

 ,
,  , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. PDCD1 and PD-L1 Polymorphisms
2.2. Linkage Disequilibrium
2.3. The Association of PDCD1 and PD-L1 Genes Polymorphisms with ccRCC Risk
2.4. Haplotype Analysis
2.5. SNP-SNP Interactions between Variations in PDCD1 and PD-L1 Genes in Relation to the ccRCC Risk
2.6. PDCD1 and PD-L1 Polymorphisms and Overall Survival of ccRCC Patients
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Genotyping
4.3. In silico Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwald, R.J.; Freeman, G.J.; Sharpe, A.H. The B7 family revisited. Annu. Rev. Immunol. 2005, 23, 515–548. [Google Scholar] [CrossRef]
- Okazaki, T.; Honjo, T. PD-1 and PD-1 ligands: From discovery to clinical application. Int. Immunol. 2007, 19, 813–824. [Google Scholar] [CrossRef] [Green Version]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Dermani, F.K.; Samadi, P.; Rahmani, G.; Kohlan, A.K.; Najafi, R. PD-1/PD-L1 immune checkpoint: Potential target for cancer therapy. J. Cell. Physiol. 2019, 234, 1313–1325. [Google Scholar] [CrossRef]
- Darvin, P.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Immune checkpoint inhibitors: Recent progress and potential biomarkers. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vuong, L.; Kotecha, R.R.; Voss, M.H.; Hakimi, A.A. Tumor Microenvironment Dynamics in Clear-Cell Renal Cell Carcinoma. Cancer Discov. 2019, 9, 1349–1357. [Google Scholar] [CrossRef] [Green Version]
- Linehan, W.M.; Ricketts, C.J. The Cancer Genome Atlas of renal cell carcinoma: Findings and clinical implications. Nat. Rev. Urol. 2019, 16, 539–552. [Google Scholar] [CrossRef]
- Klatte, T.; Rossi, S.H.; Stewart, G.D. Prognostic factors and prognostic models for renal cell carcinoma: A literature review. World J. Urol. 2018, 36, 1943–1952. [Google Scholar] [CrossRef] [PubMed]
- Salanti, G.; Amountza, G.; Ntzani, E.E.; Ioannidis, J.P. Hardy-Weinberg equilibrium in genetic association studies: An empirical evaluation of reporting, deviations, and power. Eur. J. Hum. Genet. EJHG 2005, 13, 840–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemi, M.; Karami, S.; Sarabandi, S.; Moazeni-Roodi, A.; Malecki, A.; Ghavami, S.; Wiechec, E. Association between PD-1 and PD-L1 Polymorphisms and the Risk of Cancer: A Meta-Analysis of Case-Control Studies. Cancers 2019, 11, 1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Gong, M.; Shi, Z.; Xiao, J.; Zhang, J.; Peng, J. Programmed Cell Death-1 Polymorphisms Decrease the Cancer Risk: A Meta-Analysis Involving Twelve Case-Control Studies. PLoS ONE 2016, 11, e0152448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zining, J.; Lu, X.; Caiyun, H.; Yuan, Y. Genetic polymorphisms of mTOR and cancer risk: A systematic review and updated meta-analysis. Oncotarget 2016, 7, 57464–57480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Xu, Q.; Tu, H.; Sun, L.; Gong, Y.; Liu, J.; Yuan, Y. Polymorphic rs9471643 and rs6458238 upregulate PGC transcription and protein expression in overdominant or dominant models. Mol. Carcinog. 2016, 55, 586–599. [Google Scholar] [CrossRef]
- Tao, L.H.; Zhou, X.R.; Li, F.C.; Chen, Q.; Meng, F.Y.; Mao, Y.; Li, R.; Hua, D.; Zhang, H.J.; Wang, W.P.; et al. A polymorphism in the promoter region of PD-L1 serves as a binding-site for SP1 and is associated with PD-L1 overexpression and increased occurrence of gastric cancer. Cancer Immunol. Immunother. 2017, 66, 309–318. [Google Scholar] [CrossRef]
- Catalano, C.; da Silva Filho, M.I.; Frank, C.; Jiraskova, K.; Vymetalkova, V.; Levy, M.; Liska, V.; Vycital, O.; Naccarati, A.; Vodickova, L.; et al. Investigation of single and synergic effects of NLRC5 and PD-L1 variants on the risk of colorectal cancer. PLoS ONE 2018, 13, e0192385. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liao, P.; He, Y. A gene polymorphism in PD-L1 promoter region is not associated with PD-L1 expression and patients’ survival in gastric cancer. Cancer Immunol. Immunother. 2017, 66, 1379–1381. [Google Scholar] [CrossRef]
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Davis, C.A.; Hitz, B.C.; Sloan, C.A.; Chan, E.T.; Davidson, J.M.; Gabdank, I.; Hilton, J.A.; Jain, K.; Baymuradov, U.K.; Narayanan, A.K.; et al. The Encyclopedia of DNA elements (ENCODE): Data portal update. Nucleic Acids Res. 2018, 46, D794–D801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.M.; Barber, G.P.; Casper, J.; Clawson, H.; Diekhans, M.; Gonzalez, J.N.; Hinrichs, A.S.; Lee, B.T.; Nassar, L.R.; Powell, C.C.; et al. UCSC Genome Browser enters 20th year. Nucleic Acids Res. 2020, 48, D756–D761. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbloom, K.R.; Sloan, C.A.; Malladi, V.S.; Dreszer, T.R.; Learned, K.; Kirkup, V.M.; Wong, M.C.; Maddren, M.; Fang, R.; Heitner, S.G.; et al. ENCODE data in the UCSC Genome Browser: Year 5 update. Nucleic Acids Res. 2013, 41, D56–D63. [Google Scholar] [CrossRef]
- Consortium, G.T. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Ward, L.D.; Kellis, M. HaploReg: A resource for exploring chromatin states, conservation, and regulatory motif alterations within sets of genetically linked variants. Nucleic Acids Res. 2012, 40, D930–D934. [Google Scholar] [CrossRef] [Green Version]
- Ward, L.D.; Kellis, M. HaploReg v4: Systematic mining of putative causal variants, cell types, regulators and target genes for human complex traits and disease. Nucleic Acids Res. 2016, 44, D877–D881. [Google Scholar] [CrossRef]
- Boyle, A.P.; Hong, E.L.; Hariharan, M.; Cheng, Y.; Schaub, M.A.; Kasowski, M.; Karczewski, K.J.; Park, J.; Hitz, B.C.; Weng, S.; et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 2012, 22, 1790–1797. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar] [CrossRef]



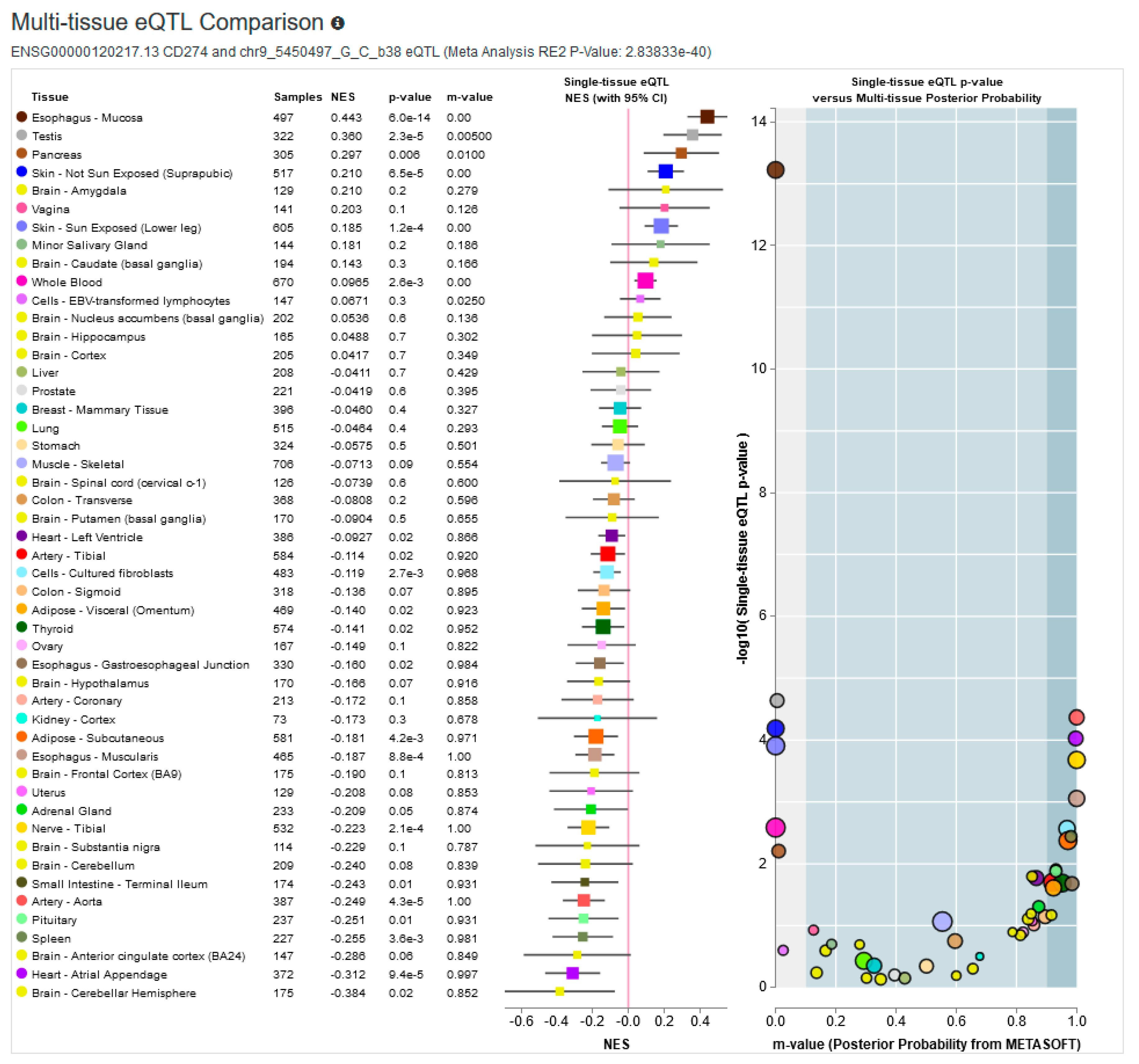{kind=link}
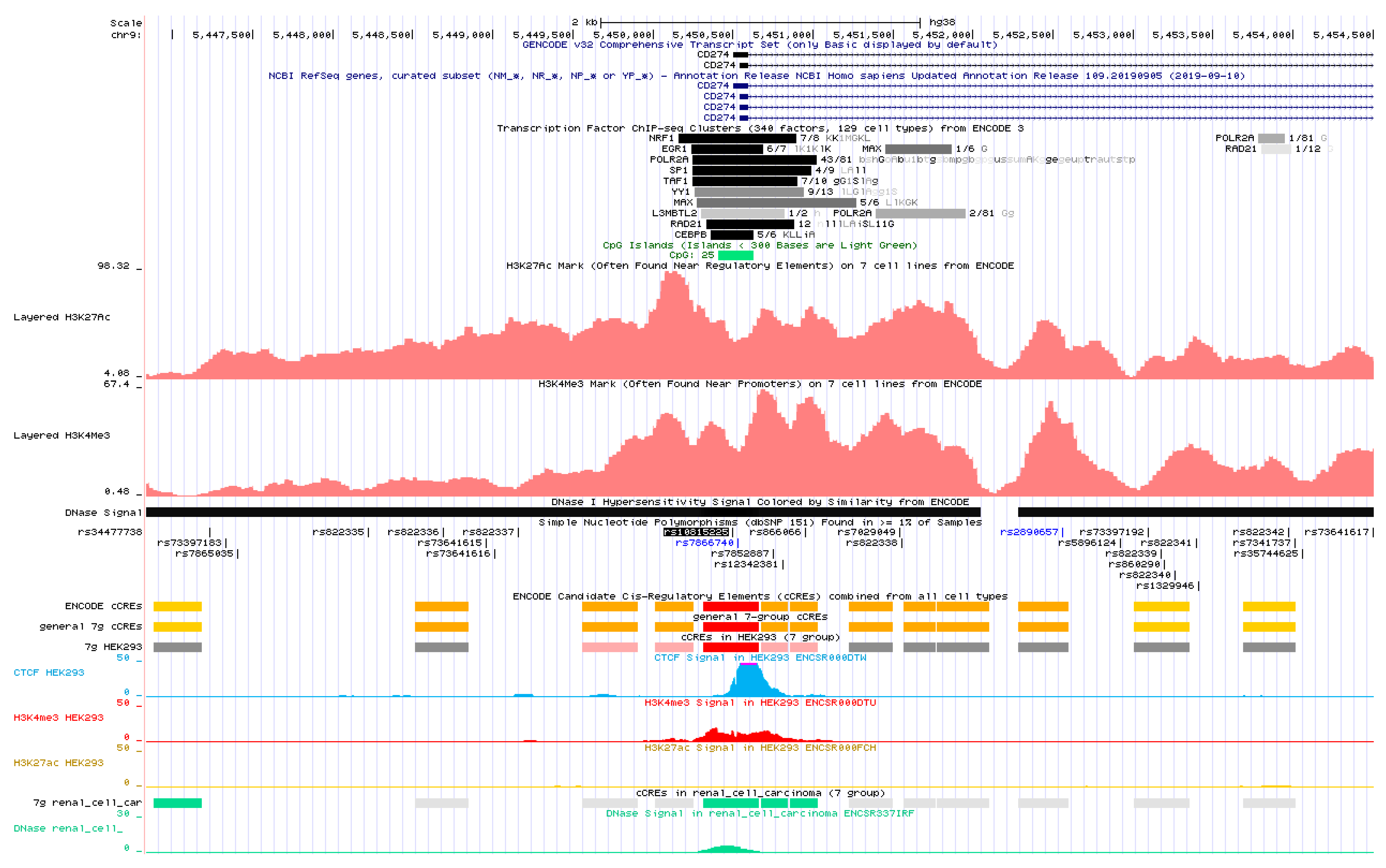{kind=link}
| Gene | Polymorphisms | Genotype | ccRCC Patients a | Controls b | OR (CI95%) | ccRCC vs. Controls | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | % | HWE | N | % | HWE | |||||
| PDCD1 (PD-1) (-strand) | rs36084323 (PD-1.1) | GG | 201 | 96.6 | p = 0.805; f = −0.017 | 249 | 97.3 | p = 0.824; f = −0.014 | 1 c | χ2 = 0.156; p = 0.693 |
| GA | 7 | 3.4 | 7 | 2.7 | 1.24 (0.44; 3.46) | |||||
| AA | 0 | 0.00 | 0 | 0.00 | − | |||||
| rs11568821 (PD-1.3) | GG | 164 | 78.8 | p = 0.050; f = 0.136 | 206 | 80.5 | p = 0.445; f = 0.048 | 1 c | χ2 = 0.974; p = 0.614 | |
| GA | 38 | 18.3 | 46 | 18.0 | 1.04 (0.65; 1.67) | |||||
| AA | 6 | 2.9 | 4 | 1.6 | 1.81 (0.54; 6.14) | |||||
| rs2227981 (PD-1.5) | CC | 70 | 33.7 | p = 0.587; f = 0.038 | 82 | 32.0 | p = 0.361; f = −0.057 | 1 c | χ2 = 1.068; p = 0.586 | |
| CT | 98 | 47.1 | 132 | 51.6 | 0.87 (0.58; 1.31) | |||||
| TT | 40 | 19.2 | 42 | 16.4 | 1.12 (0.65; 1.90) | |||||
| rs10204525 (PD-1.6) | GG | 172 | 82.7 | p = 0.581; f = −0.038 | 208 | 81.2 | p = 0.037; f = 0.131 | 1 c | χ2 = 2.681; p = 0.262 | |
| GA | 35 | 16.8 | 42 | 16.4 | 1.01 (0.62; 1.65) | |||||
| AA | 1 | 0.5 | 6 | 2.3 | 0.28 (0.05; 1.67) | |||||
| rs7421861 | TT | 96 | 46.2 | p = 0.048; f = 0.137 | 99 | 38.7 | p = 0.142; f = −0.052 | 1 c | χ2 = 6.272; p = 0.043 | |
| TC | 81 | 38.9 | 129 | 50.4 | 0.65 (0.44; 0.96) | |||||
| CC | 31 | 14.9 | 28 | 10.9 | 1.14 (0.64; 2.08) | |||||
| PD-L1 (+strand) | PD-L1 (rs822335) | CC | 77 | 37.6 | p = 0.149; f = −0.101 | 102 | 40.5 | p = 0.429; f = 0.050 | 1 c | χ2 = 2.600; p = 0.273 |
| CT | 105 | 51.2 | 112 | 44.4 | 1.24 (0.83; 1.84) | |||||
| TT | 23 | 11.2 | 38 | 15.1 | 0.81 (0.45; 1.46) | |||||
| PD-L1 (rs4143815) | GG | 97 | 47.3 | p = 0.193; f = 0.091 | 122 | 48.4 | p = 0.455; f = 0.047 | 1 c | χ2 = 0.427; p = 0.808 | |
| GC | 82 | 40.0 | 103 | 40.9 | 1.00 (0.68; 1.48) | |||||
| CC | 26 | 12.7 | 27 | 10.7 | 1.21 (0.67; 2.20) | |||||
| PD-L1 (rs4742098) | AA | 122 | 59.5 | p = 0.697; f = 0.027 | 137 | 54.4 | p = 0.737; f = −0.021 | 1 c | χ2 = 1.231; p = 0.540 | |
| AG | 71 | 34.6 | 99 | 39.3 | 0.81 (0.55; 1.19) | |||||
| GG | 12 | 5.9 | 16 | 6.3 | 0.85 (0.39; 1.84) | |||||
| PD-L1 (rs10815225) | GG | 163 | 79.5 | p = 0.102; f = −0.114 | 214 | 84.9 | p = 0.064; f = 0.117 | 1 c | χ2 = 6.981; p = 0.030 | |
| GC | 42 | 20.5 | 34 | 13.5 | 1.62 (0.99; 2.65) | |||||
| CC | 0 | 0.0 | 4 | 1.6 | 0.15 (0.01; 2.73) | |||||
| PD-L1 Haplotype | ccRCC Patients N (%) | Controls N (%) | χ2 | p | OR (CI95%) | |||
|---|---|---|---|---|---|---|---|---|
| rs822335 | rs10815225 | rs4143815 | rs4742098 | |||||
| C | G | C | A | 14.5 (3.5) | 20.3 (4.0) | 0.205 | 0.651 | 0.85 (0.43; 1.70) |
| C | C | C | G | 15.4 (3.7) | 22.3 (4.4) | 0.346 | 0.557 | 0.82 (0.42; 1.59) |
| C | G | C | G | 22.7 (5.5) | 47.3 (9.4) | 5.154 | 0.023 | 0.55 (0.33; 0.93) |
| C | C | G | A | 25.8 (6.3) | 15.8 (3.1) | 4.873 | 0.027 | 2.03 (1.07; 3.86) |
| C | G | G | A | 177.5 (43.3) | 197.2 (39.1) | 1.003 | 0.317 | 1.15 (0.88; 1.50) |
| T | G | C | A | 27.8 (6.8) | 20.9 (4.1) | 2.857 | 0.091 | 1.65 (0.92; 2.95) |
| T | G | C | G | 53.6 (13.1) | 46.2 (9.2) | 3.107 | 0.078 | 1.45 (0.96; 2.21) |
| T | G | G | A | 69.3 (16.9) | 118.8 (23.6) | 7.051 | 0.008 | 0.64 (0.46; 0.89) |
| global χ2 = 21.541; p = 0.003 | ||||||||
| rs10815225 GG | rs10815225 GC + CC | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| ccRCC Patients | Controls | OR (CI95%) | ccRCC Patients | Controls | OR (CI95%) | ||||
| rs7421861 | TT + CC | 92 (56.4%) | 109 (50.9%) | 1 a | χ2 = 1.124; p = 0.289 | 33 (78.6) | 16 (42.1%) | 1 a | χ2 = 11.038; p = 0.0009 |
| TC | 71 (43.6%) | 105 (49.0%) | 0.80 (0.53; 1.31) | 9 (21.4%) | 22 (52.4%) | 0.21 (0.08; 0.54) | |||
| rs7421861 TT + CC | rs7421861 TC | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| ccRCC Patients | Controls | OR (CI95%) | ccRCC Patients | Controls | OR (CI95%) | ||||
| rs10815225 | GG | 92 (73.6%) | 109 (87.2%) | 1 a | χ2 = 7.306; p = 0.007 | 71 (88.7%) | 105 (82.7%) | 1 a | χ2 = 1.415; p = 0.234 |
| GC + CC | 33 (26.4%) | 16 (12.8%) | 2.40 (1.25; 4.61) | 9 (11.3%) | 22 (17.3%) | 0.62 (0.28; 1.41) | |||
| RCC | ccRCC | |
|---|---|---|
| Number of patients | 237 | 208 |
| Gender | ||
| Male | 155 (65.4%) | 135 (64.9%) |
| Female | 82 (34.6) | 73 (35.1%) |
| Age at diagnosis | ||
| Q1 (years) | 57 | 57 |
| Median (years) | 62 | 62 |
| Q3 (years) | 71 | 70.75 |
| Range (min-max) (years) | 21–85 | 21–85 |
| Stage at presentation | ||
| I | 114 (48.1%) | 101 (48.6%) |
| II | 31 (13.1%) | 26 (12.5%) |
| III | 26 (11.0%) | 24 (11.5%) |
| IV | 66 (27.8%) | 57 (27.4%) |
| Grade | ||
| I | 105 (44.3%) | 96 (46.2%) |
| II | 64 (27.0%) | 62 (29.8%) |
| III | 36 (15.2%) | 35 (16.8%) |
| IV | 9 (3.8%) | 9 (4.3%) |
| n.a. a | 23 (9.7%) | 6 (2.9%) |
| Presence of venous thrombus | ||
| absent | 218 (92.0%) | 193 (92.8%) |
| present | 19 (8.0%) | 15 (7.2%) |
| Presence of sarcomatoid component | ||
| absent | 203 (85.7) | 182 (87.5%) |
| present | 9 (3.8%) | 9 (4.3%) |
| n.a. a | 25 (10.5%) | 17 (8.2%) |
| Survival status | ||
| Dead | 117 (49.4%) | 105 (50.5%) |
| Alive | 120 (50.6%) | 103 (49.5%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, M.; Tupikowski, K.; Jasek, M.; Tomkiewicz, A.; Witkowicz, A.; Ptaszkowski, K.; Karpinski, P.; Zdrojowy, R.; Halon, A.; Karabon, L. SNP-SNP Interaction in Genes Encoding PD-1/PD-L1 Axis as a Potential Risk Factor for Clear Cell Renal Cell Carcinoma. Cancers 2020, 12, 3521. https://doi.org/10.3390/cancers12123521
Wagner M, Tupikowski K, Jasek M, Tomkiewicz A, Witkowicz A, Ptaszkowski K, Karpinski P, Zdrojowy R, Halon A, Karabon L. SNP-SNP Interaction in Genes Encoding PD-1/PD-L1 Axis as a Potential Risk Factor for Clear Cell Renal Cell Carcinoma. Cancers. 2020; 12(12):3521. https://doi.org/10.3390/cancers12123521
Chicago/Turabian StyleWagner, Marta, Krzysztof Tupikowski, Monika Jasek, Anna Tomkiewicz, Agata Witkowicz, Kuba Ptaszkowski, Pawel Karpinski, Romuald Zdrojowy, Agnieszka Halon, and Lidia Karabon. 2020. "SNP-SNP Interaction in Genes Encoding PD-1/PD-L1 Axis as a Potential Risk Factor for Clear Cell Renal Cell Carcinoma" Cancers 12, no. 12: 3521. https://doi.org/10.3390/cancers12123521